
Abstract
Isotope-based records provide valuable information on past climate changes. However, it is not always trivial to disentangle past changes in the isotopic composition of precipitation from possible changes in evaporative enrichment, and seasonality may need to be considered. Here, we analyzed δ2H on n-alkanes and δ18O on hemicellulose sugars in sediments from Bichlersee, Bavaria, covering the Late Glacial and Early Holocene. Our δ2Hn-C31 record documents past changes in the isotopic composition of summer precipitation and roughly shows the isotope pattern known from Greenland. Both records show lower values during the Younger Dryas, but at Bichlersee the signal is less pronounced, corroborating earlier suggestions that the Younger Dryas was mainly a winter phenomenon and less extreme during summer. δ18Ofucose records the isotopic composition of the lake water during summer and is sensitive to evaporative enrichment. Coupling δ2Hn-C31 and δ18Ofucose allows calculating lake water deuterium-excess and thus disentangling changes in the isotopic composition of precipitation and evaporative enrichment. Our deuterium-excess record reveals that the warm Bølling–Allerød and Early Holocene were characterized by more evaporative enrichment compared to the colder Younger Dryas. Site-specific hydrological conditions, seasonality, and coupling δ2H and δ18O are thus important when interpreting isotope records.
Similar content being viewed by others
Introduction
High-altitude ecosystems, such as the European Alps, are highly sensitive to climate change1,2,3,4. Hydrological aspects like melting glaciers and changing precipitation pattern will increase the risk of landscape destabilization, water scarcity and more frequent flooding in the future5,6,7. In this context, paleoclimate studies can provide valuable information to better understand past and predict future climate-landscape interactions8. In particular the Late Glacial–Early Holocene transition is of interest because it is known for its rapid climate changes and has been intensively investigated9,10. The Late Glacial comprises the warm Bølling–Allerød interstadial (starting ~ 14.7 ka BP) and the cold Younger Dryas stadial (from ~ 12.8 to 11.7 ka BP)11,12. Paleoclimate information from the Alps during that time is inferred inter alia from stable oxygen isotope records (δ18O) derived from lake sediments13,14,15,16,17 and speleothems18,19. Those δ18O records mostly resemble the δ18O records from Greenland ice cores20 and are often interpreted to document past temperature changes. However, the interpretation of δ18O alone can be challenging because various factors can influence the isotopic signal, which are difficult to disentangle. Those include changes in atmospheric circulation, precipitation, seasonality, carbonate chemistry, and regarding lake sediments, evaporative enrichment of lake water18,21,22,23. Past changes in evaporative enrichment could have played an important role due to rapidly changing warm and cold conditions during the Late Glacial–Early Holocene transition. Although this effect can complicate the interpretation of δ18O records, evaporative enrichment itself provides very valuable hydrological information, e.g., about wet or dry conditions. This is particularly interesting in view of the scarce and controversial data concerning the hydroclimatic development during the Late Glacial. Pollen from terrestrial and marine sediments, for example, are often interpreted to indicate warm and humid conditions during the Bølling–Allerød and Early Holocene24,25, but drier conditions have also been suggested26,27. For the Younger Dryas, there is also no consensus, although first attempts of paleohydrological reconstructions have been made using novel biomarker and isotope analyses28,29,30.
Over the last few years, compound-specific hydrogen (δ2H) and oxygen (δ18O) isotope analyses on biomarkers, i.e., molecular fossils, have been developed. Long-chain n-alkanes (n-C29 and n-C31), for example, are leaf waxes produced by higher terrestrial plants. Their δ2H signal mainly reflects the isotopic composition of the local precipitation but can be modulated by transpirative enrichment of the leaf water and biosynthetic fractionation31,32,33,34. Shorter-chain n-alkanes (e.g., n-C21 and n-C23), on the other hand, are produced by aquatic organisms and incorporate the δ2H signal of the lake water, i.e., depending on the hydrological setting, evaporation may lead to isotopic enrichment35,36,37,38,39. Like δ2H from n-alkanes, δ18O from hemicellulose sugars provides valuable paleohydrological information34,35,40,41. Hemicellulose sugars are also produced by terrestrial (i.e., arabinose) and aquatic (i.e., fucose) sources42. Their δ18O signal mainly reflects the isotopic composition of the local precipitation (arabinose) and the lake water (fucose) modulated by evapo(transpi)rative enrichment and biosynthetic fractionation32,33,41,43. The “coupled isotope approach”, also dubbed “paleohygrometer approach”, combines δ2Hn-alkane and δ18Osugar analyses to reconstruct deuterium excess (d-excess), which can be used as a proxy for evapo(transpi)rative enrichment30,35,43,44. This approach has been tested and applied successfully in several studies to quantify the evapo(transpi)rative enrichment of leaf water30,35,40,43,44,45 and lake water35.
So far, very few studies have applied biomarker and compound-specific isotope analyses in the European Alps during the Late Glacial-Early Holocene transition23,46, but they have great potential to investigate the common notion of warm and humid interstadials, versus cool and dry stadials. Therefore, the aim of this study is to establish a high-resolution δ2Hn-alkane record for the Late Glacial–Early Holocene sediments from Bichlersee (Bavaria, Germany) and complement this with δ18Osugar analyses. Specifically, we aim to (1) identify the sources of the biomarker compounds (terrestrial versus aquatic) and (2) discuss the role of evaporative and transpirative enrichment of the lake and leaf water, respectively. Afterwards, (3) the coupled isotope approach will be applied to calculate the deuterium excess as a proxy for lake water evaporation.
Geographic setting of Bichlersee
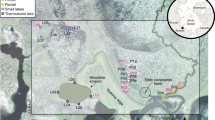
Bichlersee (See = lake) is located in the Northern Limestone Alps ~ 5 km northwest of Oberaudorf in the lower Inn Valley (Fig. 1a,b). It is situated in a small karst depression at 960 m a.s.l. west of the Wildbarren (1,448 m a.s.l.). The circular lake has an area of 0.01 km2 and a maximum water depth of 11 m47. The catchment comprises an area of ~ 0.5 km2 including swampy areas along the lake shore and three small creeks (Fig. 1b,c). The surrounding slopes are covered by dense mountain forest consisting of Picea, Abies, and Fagus. The study site is affected by the Westerly circulation system bringing moisture from the Atlantic Ocean to the Alps. Mean annual temperature (MAT) in Kiefersfelden (~ 7.5 km to the south, 520 m a.s.l.) is 8.8 °C and mean annual precipitation (MAP) is 1308 mm (Reference period 1991–2020, Fig. 1d)48. The isotopic composition of precipitation (δ2Hp, δ18Op) shows a strong seasonal variability: Summer precipitation is isotopically enriched (− 34‰ for δ2Hp and − 5.2‰ for δ18Op in July), whereas winter precipitation is generally strongly depleted with − 108‰ for δ2Hp and − 14.8‰ for δ18Op in January49,50. The isotopic composition of precipitation is very heterogenous across the European Alps and influenced by various effects, including different moisture sources. However, the most important effect explaining ~ 70% of the seasonal variability in δ18Op and δ2Hp is temperature23,51.

Geographic setting of Bichlersee. (a) Overview map showing the location of the study site. (b) Topographical map of the Bichlersee area (Data: Copernicus EuropeDEM 1.1). (c) Photograph of Bichlersee and the forested Wildbarren (Photo: M. Prochnow). (d) Climate diagram from Kiefersfelden illustrating local climate conditions (1991–2020)48. The maps in (a) and (b) were created with ArcGIS Pro 2.9.2 (www.esri.com/de-de/arcgis/products/arcgis-pro) and all image labels were added using Inkscape 1.2.2 (www.inkscape.org). The climate diagram in (d) was created with Climatol 4.0.5 for R (www.climatol.eu).
Results and discussion
Lithology and chronology
In 2014, we recovered a 9.4 m long sediment core from Bichlersee. For this study, we focus on the sediments between 520 and 420 cm, which comprise the Late Glacial and transition into the Early Holocene. Unit A from 520 to 508 cm shows light colored sediments with TOC contents below 5% (Fig. 2). Unit B (508–487 cm) consists of dark sediments with high TOC contents up to 13%. Unit C (487–467 cm) is then again characterized by lighter colors and lower TOC contents, while Unit D (above 467 cm) is dark-brown with high TOC contents (15%). The two bright mottles in Unit D (458 cm, 445 cm) probably indicate disturbance of the sediment and were avoided during sampling. The rest of the core is finely layered.

Core photograph, age depth model, and results of geochemical as well as stable isotope analyses for the Late Glacial–Early Holocene part of our core from Bichlersee. The graphic was created with Inkscape 1.2.2 (www.inkscape.org).
Radiocarbon ages from eight organic macrofossils and charcoal particles (BS_1 to BS_8) provide a very consistent and robust chronological control (Fig. 2, Table 1). Bayesian age-depth modeling gives a basal median age of 15,118 +980/−860 cal. BP. The age-depth model is corroborated by the pattern of TOC, suggesting that Unit A reflects the Oldest Dryas, Unit B the Bølling–Allerød, Unit C the Younger Dryas and Unit D the Early Holocene12,20. TOC reflects changes in productivity during these periods, with lower TOC during the Younger Dryas because of cooler temperatures and reduced bioproductivity. This seems justified given that similar TOC pattern during the Late Glacial have been reported from other lakes, for example the pre-alpine lakes Steißlingen52 and Mondsee15. A tree cone 14C age (12,130 +600/−770 cal. BP) falls directly into the Younger Dryas, and the Early Holocene is covered by several consistent terrestrial macrofossil 14C ages ranging from 11,430 +1160/−930 to 10,780 +1040/−880 cal. BP (Fig. 2).
n-Alkanes and compound-specific δ2H
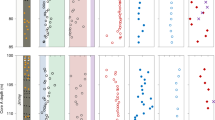
n-Alkanes were obtained in sufficient amounts for high-resolution compound-specific δ2H analyses only for the Late Glacial and Early Holocene part of the core, and we had to focus on the most abundant homologues n-C27 to n-C31. δ2H ranges from − 223.7 to − 170.1‰ for n-C27, from − 222.5 to − 181.3‰ for n-C29, and from − 220.7 to − 180.3‰ for n-C31, respectively (Fig. 2). n-Alkanes are generally more enriched in Unit B and Unit D, and more depleted in Unit C. δ2Hn-C29 and δ2Hn-C31 show a very similar pattern, whereas δ2Hn-C27 is more variable and more positive, which indicates different sources and different paleohydrological implications.
In general, n-C31 and n-C33 tend to be more abundant in grasses and herbs, whereas n-C27 and n-C29 are preferentially produced by deciduous trees and shrubs53. With the beginning of the Bølling–Allerød, the vegetation was dominated by Betula and Pinus forests persisting at least into the Preboreal54,55. Such forests have a dense grassy understory vegetation, as we can confirm by high abundances of Poaceae in three exemplarily investigated pollen samples within the Bølling–Allerød (7–11%; see Supplementary Fig. S3). The most abundant pollen are Betula pendula with 25–43% and Pinus sylvestris with 43–60%. We can therefore assume that n-C31 is primarily derived from grasses, because coniferous trees produce no long-chain n-alkanes, yet Pinus growing in the montane zone of the Alps is known to synthesize only very low amounts of n-alkanes46. While a distinct exclusive attribution is not possible, we can use the δ2Hn-C31 signal as the best available record for terrestrial grasses, which is very relevant with regard to the fact that leaf waxes from grasses are less affected by transpirative enrichment than trees and shrubs56. This is due to the so-called ‘damping-effect’45,56, i.e., grasses grow through the less exposed intercalary meristems57 and are thus less affected by transpirative leaf water enrichment58. δ2Hn-C31 is therefore assumed to mainly reflect changes in the isotopic composition of precipitation during the growing season.
Our δ2Hn-C31 record is in good agreement with results from Meerfelder Maar29 and Gemündener Maar30 in northern Germany, providing a coherent pattern between the existing terrestrial δ2Hn-alkane records (Fig. 4e). This pattern mostly follows δ18O and δ2H in Greenland ice cores20 (Fig. 4f,g), where the reconstructed isotopic composition of precipitation is traditionally explained to reflect northern hemispheric temperature changes11 (i.e., the typical stadial-interstadial pattern): more enriched values during the Bølling–Allerød interstadial and Early Holocene are related to warmer temperatures, interrupted by more depleted values during the cooler Younger Dryas stadial (Fig. 4a,f). Detailed comparison reveals that δ2H in Greenland (Fig. 4g) varies by ~ 40‰ during the Bølling–Allerød and drops markedly into the Younger Dryas59, while δ2Hn-C31 at Bichlersee decreases rather gently by only ~ 20‰ (Fig. 4a). While temperature and related isotope changes were probably more pronounced in Greenland than at lower latitudes like the Alps60, seasonality might have a greater influence on the isotopic signal at our site23. Because temperatures dropped much less in summers than winters during the Younger Dryas61,62, our (summer) δ2Hn-C31 record does not show the pronounced (annual) stadial-interstadial signal as the ice cores from Greenland.
We suggest that the more variable and more positive δ2Hn-C27 signal (up to ~ 20‰) can be explained by the fact that n-C27 is to a higher degree derived from deciduous trees and shrubs than n-C29 or even n-C31. n-C27 is therefore more affected by leaf water transpirative enrichment (Fig. 4a). Betula pendula produces high amounts of n-C2746,53,63 and its occurrence at Bichlersee is documented by high pollen abundances since the Bølling–Allerød (see Supplementary Fig. S3). Vegetation changes such as an expansion of grasses during the Younger Dryas64 might affect the isotopic composition of n-alkanes, i.e. leading to more depleted δ2H, but light Betula and Pinus forests persisted throughout the Younger Dryas at sites < 1700 m a.s.l.26,65. This is corroborated by relatively constant relative abundances of n-C27 and n-C31 during the Bølling–Allerød and Younger Dryas at Bichlersee (see Supplementary Fig. S4).
Recent studies provide evidence that long-chain n-alkanes including n-C27 can also originate from aquatic plants66,67,68. In fact, the relative concentration of n-C27 positively correlates with n-C25 (r = 0.8), but negatively with n-C29 and n-C31 (r = − 0.6 and − 0.4) in our dataset (Table 2). We therefore assume that n-C27 has a mixed origin from aquatic and terrestrial sources and therefore partly reflects lake water, not just leaf water δ2H.
Hemicellulose sugars and compound-specific δ18O
A ternary diagram (Fig. 3) with the relative concentrations of arabinose, fucose and xylose shows that our samples from Bichlersee have a relative abundance of ~ 30% fucose, which is comparable to lake sediment samples from Panch Pokhari, Nepal69, Gemündener Maar, Germany30, as well as some submerged plants from Bichlersee42. Samples from emergent and terrestrial plants from Bichlersee, on the other hand, show almost no fucose42. Fucose is known to be highly abundant in zooplankton, phytoplankton and aquatic bacteria70, but low in terrestrial plants42,69. It is thus a predominantly aquatic compound. Arabinose and xylose, on the other hand, are of mixed aquatic and terrestrial origin at Bichlersee. In the following, we will therefore focus on δ18Ofucose as a proxy for lake water δ18O, as recently suggested by Bittner, et al.41 who showed that δ18Ofucose agrees very well with δ18Odiatom.

Ternary diagram of relative abundances of hemicellulose sugars from Bichlersee. The diagram shows relative abundances of arabinose, fucose, and xylose in the Bichlersee samples (red dots) and data from previous studies: emergent, terrestrial and submerged plants from Bichlersee42, Panch Pokhari69 and Gemündener Maar30. The graphic was created with Inkscape 1.2.2 (www.inkscape.org).
δ18Ofucose ranges from 22.8 to 37.8‰ and is highly variable (Fig. 2). It shows the stadial-interstadial pattern, i.e., more positive values during the Bølling–Allerød and Early Holocene interrupted by more negative values during the Younger Dryas (Fig. 4b). However, the pattern of our δ18Ofucose record reveals a high variability and is less distinct compared to our δ2H record, Greenland δ18O (Fig. 4f), and δ18O from other alpine and pre-alpine lakes, for example, Lake Ammersee14 and Mondsee15 (Fig. 4h). These differences are partly related to specific hydrological settings like surface area, water volume and lakebed geometry. Ammersee, for instance, located 80 km northwest of Bichlersee near Munich, has a large catchment area and a large and deep water body. δ18O on benthic ostracods from Ammersee thus likely represents an annual lake water signal17 and is less sensitive to the effect of evaporative enrichment14. Bichlersee and its catchment, on the other hand, are very small and particularly sensitive to record the isotopic signal of summer precipitation. As mentioned above for δ2H, the Younger Dryas was mainly a winter temperature phenomenon61,62, so one can hypothesize that—analogously to our δ2Hn-C31 record—a summer isotope record like δ18Ofucose shows a less pronounced stadial-interstadial pattern than Greenland. Moreover, the high variability of our δ18Ofucose record can at least partly be explained with the high sensitivity of Bichlersee for evaporative enrichment. Bichlersee has a closed basin (although groundwater outflow is likely), so the ratio of precipitation to evaporation (P/E) in summer strongly controls δ18O of the lake water41 and the fucose. Several studies have shown that lake water 18O enrichment varies highly between lakes and can reach up to several per mille (~ 5‰ in δ18O) even under todays humid climate conditions in Germany and the Alps16,21,71,72. Evaporative enrichment can best be discussed not just looking at one isotope record, but combining δ2H and δ18O.

Late Glacial–Early Holocene compilation of (a) δ2Hn-alkane and (b) δ18Ofucose from Bichlersee. The estimated d-excess for Bichlersee is plotted in (c) and the d-excess from Gemündener Maar30 is shown in (d). δ2Hn-alkane from Meerfelder Maar29 is given in (e), (f) and (g) illustrate ice core δ18O and δ2H from NGRIP (Greenland)20,59 reflecting northern hemispheric temperature, (h) reflects ostracod δ18O from Ammersee14 and Mondsee15, and (i) shows the July summer insolation at 48° N78. The graphic was created with Inkscape 1.2.2 (www.inkscape.org).
Calculating deuterium excess by coupling of δ18Osugar and δ2Hn-alkane
Based on the discussion above, δ2Hn-C31 and δ18Ofucose are the best available terrestrial and aquatic isotope proxies, respectively. Based on these, one can calculate lake water deuterium excess (d-excess), an important proxy for evaporative enrichment, using an adapted version of the “coupled isotope approach” described by Hepp et al.35. As illustrated in the δ18O–δ2H-diagram (Fig. 5), the basic assumption of this approach is that the isotopic composition of lake water can be calculated by applying the specific biosynthetic fractionation for n-alkanes and sugars. Lake water then plots on a local evaporation line (LEL). When the degree of evaporative enrichment is low, the isotopic composition of the lake water plots close to the starting point of the LEL on the Global Meteoric Waterline (GMWL, Eq. 1)60:

Conceptual framework of the coupled isotope approach after Hepp et al.35. The graphic was created with Inkscape 1.2.2 (www.inkscape.org).
With increasing degree of evaporative enrichment, the distance of lake water to the GMWL increases following the LEL. Thus, deuterium excess (d-excess) of the lake water is defined as distance from the GMWL, with more negative values indicating stronger evaporative enrichment. Instead of the GMWL, site-specific Local Meteoric Waterlines (LMWL, Eq. 2) can be used, in our case
based on data from the climate station Garmisch-Partenkirchen73.
Applying the coupled isotope approach, we first subtracted the apparent fractionation − 145 ± 12‰ for Central Europe34 from our δ2Hn-C31 values in order to estimate δ2H of precipitation (δ2Hp) used for biosynthesis (Fig. 5). As described in “n-Alkanes and compound-specific δ2H” section, this is based on the assumption that δ2Hn-C31 is primarily derived from grasses, which are less affected by transpirative enrichment and therefore changes in δ2Hn-C31 mainly record changes in the isotopic composition of precipitation. δ2Hp is then used to calculate δ18Op based on the LMWL (Eq. 2). For δ18Osugar, the biosynthetic fractionation factor is ~ 27‰74,75, which can be used to calculate lake water δ18O (δ18Olake). The corresponding δ2Hlake values are derived with local evaporation lines described by Eq. (3):
where m is the temperature-dependent slope and n the intercept with δ2H (Fig. 5). A more detailed methodological description of the d-excess estimation and its potential limitations is provided in the Supplementary section S1.
In general, our d-excess record has a very similar pattern compared to δ18Ofucose (note the reversed axis in Fig. 4c, r = − 0.87, p = 6.86 × 10−10). This illustrates the high sensitivity of δ18O to evapo(transpi)rative enrichment compared to δ2H, which has previously been concluded from transect and growth chamber studies34,56,76,77. Analyzing δ18Osugar and calculating d-excess thus has a major added value compared to only analyzing δ2H on n-alkanes, as it allows to disentangle past changes in the isotopic composition of precipitation and evaporative enrichment.
Our d-excess record shows a long-term trend towards more lake water enrichment from the Late Glacial to the Early Holocene, which can possibly be explained with increasing summer insolation78 (Fig. 4i) and related evaporation. More importantly, the Bølling–Allerød and Early Holocene are characterized by enhanced enrichment compared to the Younger Dryas. This makes sense in view of the higher temperatures driving more evaporation. Our results agree well with a pollen study by Ammann, et al.26 who reconstructed higher evapo(transpi)ration during the Bølling–Allerød at Gerzensee in Switzerland. Similarly, Litt et al.27 suggested more arid conditions during the Early Holocene based on pollen from lake Holzmaar in Germany. We can further compare our record with the d-excess from the Gemündener Maar30, the only other available Late Glacial d-excess record for Central Europe (Fig. 4d). There, d-excess was derived by a coupled isotope approach based on leaf water δ18O and δ2H. Both records show high variability, but no evidence for an overall dry Younger Dryas28,29. Aridity north of the Alps during the Younger Dryas was explained by a southward migration of the Westerlies in relation to cooling and enhanced sea-ice cover in the North Atlantic79. This shift occurred most likely during winter and lead to increased winds and hence dryness in Central Europe, but as it can be expected from variable sea-ice cover in the North Atlantic80, this shift was apparently not persistent throughout the Younger Dryas81,82. It can be moreover only speculated how a shift of the Westerlies affected summer hydrology at Bichlersee, and while our study emphasizes the relevance of seasonality to this issue, we believe that more paleohydrology records are strongly needed to further confirm these atmospheric mechanisms.
Conclusion
This study presents the first lacustrine δ2Hn-alkane record from the Northern European Alps covering the Late Glacial–Early Holocene transition (~ 15–10 ka BP). We complemented this with δ18Osugar analyses to explore the potential of combining δ2H and δ18O for paleohydrological reconstructions at Bichlersee. In view of the scarce and controversial data concerning the hydroclimatic development during the Late Glacial, our study aimed to test the general notion of warm and humid interstadials, versus cool and dry stadials.
δ2Hn-C31 primarily reflects changes in the isotopic composition of summer precipitation because n-C31 is predominantly produced by grasses which are not particularly sensitive to leaf water transpirative enrichment. Our δ2Hn-C31 record shows more negative values during the Younger Dryas compared to the Bølling–Allerød and Early Holocene. It thus agrees with other published leaf-wax records and isotope records from Greenland ice cores showing the typical interstadial-stadial pattern, although the leaf-wax records are somewhat less pronounced, probably due to a summer bias and the fact that the Younger Dryas was particularly a pronounced winter phenomenon. δ2Hn-C27 is more enriched and variable, which likely reflects some transpirative and evaporative enrichment due to higher contributions from deciduous trees and aquatic sources, respectively.
δ18Ofucose is primarily of aquatic origin and reflects changes in δ18O of the lake water during summer. The typical stadial-interstadial pattern known from other δ18O records is not very obvious given the highly variable signal, but one cannot necessarily expect that pattern due to the summer bias of the proxy. Bichlersee is a small and closed basin, and it is sensitive to evaporative enrichment. Higher δ18Ofucose values during the Bølling–Allerød and Early Holocene than during the Younger Dryas may therefore indicate warmer temperatures and enhanced evaporation, but disentangling past changes in evaporative enrichment from changes in the isotopic composition of precipitation requires coupling δ2H and δ18O.
Our d-excess record basically shows the same pattern as δ18Ofucose, which illustrates the high sensitivity of δ18O to evapo(transpi)rative enrichment compared to δ2H and highlights the enormous added value of δ18Osugar analyses for paleohydrological reconstructions. The d-excess documents enhanced evaporative enrichment during the warm Bølling–Allerød and Early Holocene, and somewhat less enrichment during the Younger Dryas, which can be expected from lower temperatures. Furthermore, a long-term trend towards increasing enrichment from the Late Glacial to the Early Holocene follows and maybe explained with summer insolation.
In summary, our study highlights that isotope records have great potential for paleoclimate reconstructions. Site-specific hydrological conditions and seasonality need to be considered, and coupling δ2H and δ18O allows calculating d-excess and thus disentangling some of the many isotope effects.
Methods
Sampling and chronology
Two parallel cores were recovered from Bichlersee by using a piston coring system from UWITEC yielding a composite core length of 9.4 m in 2014. The cores were split in the lab, and one half was then subsampled at 1 cm intervals for further geochemical analyses.
Radiocarbon ages, mostly on macrofossils, were obtained in collaboration with the LARA AMS Laboratory at the University of Bern on a MIni CArbon DAting System (MICADAS) capable of direct analysis of CO2 due to the coupling to an Elementar Analyzer and a gas handling interface83. Prior to 14C analyses, all samples were treated with 1 M HCl for 8 h at 60 °C to remove carbonates. The samples were subsequently washed to pH neutrality with ultrapure water and weighed into tin boats. We performed Bayesian age-depth modelling using the ‘rBacon’ package (v. 2.5.8)84 available for R, and 14C ages were calibrated to cal. BP by applying the IntCal20 calibration curve85. Bayesian age-depth modelling was performed using the radiocarbon ages.
Organic geochemistry and palynological analyses
All samples were dried at 40 °C and homogenized. To quantify total organic carbon (TOC), aliquots were treated with 1 M HCl and measured with a Total Organic Carbon Analyzer (Shimadzu TOC-VCPN) at Technische Universität Dresden.
Three samples from 495 cm, 497 cm, and 499 cm sediment depth were prepared for pollen analyses following the procedure described by Schneider, et al.86. This includes acetolysis and treatment with hydrofluoric acid. For pollen counting, the samples were mounted in silicon oil and analyzed using a Zeiss Light microscope at 400-times magnification.
n-Alkanes and compound-specific δ2H analyses
Total lipids were extracted at Friedrich-Schiller-Universität Jena with dichloromethane:methanol (DCM:MeOH, 9:1) from 80 samples (~ 0.1–12.6 g) using ultrasonic extraction over three cycles. The n-alkanes were cleaned over aminopropyl (Supelco, 45 µm) and silver-nitrate (AgNO3; Supelco, 60–200 mesh) columns and then quantified using a gas chromatograph (Agilent 7890B) equipped with an Agilent HP5MS column (30 m × 320 μm × 0.25 μm film thickness) and coupled with a flame ionization detector (GC-FID). External n-alkane standards (n-alkane mix n-C21 to n-C40; Supelco) were measured with each sequence for identification and quantification.
Compound-specific δ2H was analyzed for the most abundant homologues (n-C27, n-C29 and n-C31) using an Isoprime Vision isotope ratio mass spectrometer (IRMS; Elementar, Langenselbold) coupled to an Agilent 7890B gas chromatograph via a GC5 pyrolysis/combustion interface. The GC5 operated in pyrolysis mode with a Cr (ChromeHD) reactor at 1050 °C. All samples were injected in splitless mode and measured in triplicates. Analytical precision is on average 1.1‰ (standard deviation) and always < 3.2‰. Between the samples, a standard n-alkane mix with known isotopic values was measured, and all isotopic values are given in delta notation (δ2Hn-alkane, ‰) versus ‘Vienna Standard Mean Ocean Water’ (V-SMOW). The \({\text{H}}_{3}^{+}\)-correction factor was checked regularly throughout the sequences and yielded stable values of 3.2 ± 0.07‰ (n = 5).
Hemicellulose sugars and compound-specific δ18O analyses
Hemicellulose sugars were extracted from 29 samples (up to ~ 600 mg, depending on TOC content) and processed according to Zech and Glaser87 at the Friedrich-Schiller-Universität Jena. The samples were extracted with 10 ml of 4 M trifluoroacetic acid at 105 °C for 4 h, cleaned using XAD-7 and Dowex 50WX8 columns and subsequently derivatized with methylboronic acid (1 mg in 100 ml pyridine) at 60 °C for 1 h. Myo-Inositol was used as internal standard.
Compound-specific δ18O of arabinose, fucose, and xylose was measured at the Technische Universität Dresden using a Trace GC 2000 coupled to a Delta V Advantage IRMS via an 18O-pyrolysis reactor (GC IsoLink) and a ConFlow IV interface (all Thermo Fisher Scientific, Germany). The samples were measured in triplicates, and standard blocks of derivatized sugars (arabinose, fucose, xylose) at various concentrations and known δ18O values were measured in between. Analytical precision is on average 0.5‰ (standard error) and always < 1.4‰. All measurements were corrected for hydrolytically introduced oxygen atoms that can form carbonyl groups within the sugar molecules87. The oxygen isotopic composition is given in the delta notation (δ18Osugar, ‰) versus V-SMOW.
Data availability
The datasets generated during the current study are available in the PANGAEA repository: https://doi.org/10.1594/PANGAEA.962035.
References
Hock, R. et al. in IPCC Special Report on the Ocean and Cryosphere in a Changing Climate (2019).
Pepin, N. et al. Elevation-dependent warming in mountain regions of the world. Nat. Clim. Change 5, 424–430. https://doi.org/10.1038/nclimate2563 (2015).
Vittoz, P. et al. Climate change impacts on biodiversity in Switzerland: A review. J. Nat. Conserv. 21, 154–162. https://doi.org/10.1016/j.jnc.2012.12.002 (2013).
Beniston, M. et al. The European mountain cryosphere: A review of its current state, trends, and future challenges. Cryosphere 12, 759–794. https://doi.org/10.5194/tc-12-759-2018 (2018).
Brunner, M. I. et al. Present and future water scarcity in Switzerland: Potential for alleviation through reservoirs and lakes. Sci. Total Environ. 666, 1033–1047. https://doi.org/10.1016/j.scitotenv.2019.02.169 (2019).
Köplin, N., Schädler, B., Viviroli, D. & Weingartner, R. Seasonality and magnitude of floods in Switzerland under future climate change. Hydrol. Process. 28, 2567–2578. https://doi.org/10.1002/hyp.9757 (2014).
Wilhelm, B. et al. Impact of warmer climate periods on flood hazard in the European Alps. Nat. Geosci. 15, 118–123. https://doi.org/10.1038/s41561-021-00878-y (2022).
Tierney, J. E. et al. Past climates inform our future. Science (New York, N.Y.) https://doi.org/10.1126/science.aay3701 (2020).
Alley, R. B. Ice-core evidence of abrupt climate changes. Proc. Natl. Acad. Sci. U.S.A. 97, 1331–1334. https://doi.org/10.1073/pnas.97.4.1331 (2000).
Brovkin, V. et al. Past abrupt changes, tipping points and cascading impacts in the Earth system. Nat. Geosci. 105, 1786. https://doi.org/10.1038/s41561-021-00790-5 (2021).
Wolff, E. W., Chappellaz, J., Blunier, T., Rasmussen, S. O. & Svensson, A. Millennial-scale variability during the last glacial: The ice core record. Quatern. Sci. Rev. 29, 2828–2838. https://doi.org/10.1016/j.quascirev.2009.10.013 (2010).
Reinig, F. et al. Precise date for the Laacher See eruption synchronizes the Younger Dryas. Nature 595, 66–69. https://doi.org/10.1038/s41586-021-03608-x (2021).
van Raden, U. J. et al. High-resolution late-glacial chronology for the Gerzensee lake record (Switzerland): δ18O correlation between a Gerzensee-stack and NGRIP. Palaeogeogr. Palaeoclimatol. Palaeoecol. 391, 13–24. https://doi.org/10.1016/j.palaeo.2012.05.017 (2013).
Grafenstein, U. V., Erlenkeuser, H., Brauer, A., Jouzel, J. & Johnsen, S. J. A mid-European decadal isotope-climate record from 15,500 to 5000 years B.P. Science (New York, N.Y.) 284, 1654–1657. https://doi.org/10.1126/science.284.5420.1654 (1999).
Lauterbach, S. et al. Environmental responses to Lateglacial climatic fluctuations recorded in the sediments of pre-Alpine Lake Mondsee (northeastern Alps). J. Quat. Sci. 26, 253–267. https://doi.org/10.1002/jqs.1448 (2011).
Grafenstein, U. V. et al. The oxygen and carbon isotopic signatures of biogenic carbonates in Gerzensee, Switzerland, during the rapid warming around 14,685 years BP and the following interstadial. Palaeogeogr. Palaeoclimatol. Palaeoecol. 391, 25–32. https://doi.org/10.1016/j.palaeo.2013.08.018 (2013).
Cartier, R. et al. Paleohydrological history of Lake Allos (2200 m a.s.l) since 13,500 cal a bp in the Mediterranean Alps inferred from an ostracod δ18O record. J. Quat. Sci. 37, 1044–1055. https://doi.org/10.1002/jqs.3425 (2022).
Luetscher, M. et al. North Atlantic storm track changes during the Last Glacial Maximum recorded by Alpine speleothems. Nat. Commun. 6, 6344. https://doi.org/10.1038/ncomms7344 (2015).
Li, H., Spötl, C. & Cheng, H. A high-resolution speleothem proxy record of the Late Glacial in the European Alps: Extending the NALPS19 record until the beginning of the Holocene. J. Quat. Sci. 36, 29–39. https://doi.org/10.1002/jqs.3255 (2021).
Rasmussen, S. O. et al. A new Greenland ice core chronology for the last glacial termination. J. Geophys. Res. 111, 527. https://doi.org/10.1029/2005jd006079 (2006).
Grafenstein, U. V. & Labuhn, I. in Paleoclimatology Vol. 137 (eds Gilles Ramstein et al.) 179–195 (Springer, 2021).
Weber, M. et al. Opposite trends in holocene speleothem proxy records from two neighboring caves in Germany: A multi-proxy evaluation. Front. Earth Sci. 9, 17. https://doi.org/10.3389/feart.2021.642651 (2021).
Wirth, S. B. & Sessions, A. L. Plant-wax D/H ratios in the southern European Alps record multiple aspects of climate variability. Quat. Sci. Rev. 148, 176–191. https://doi.org/10.1016/j.quascirev.2016.07.020 (2016).
Ponel, P., Guiter, F., Gandouin, E., Peyron, O. & Beaulieu, J.-L.D. Late-Glacial palaeotemperatures and palaeoprecipitations in the Aubrac Mountains (French Massif Central) reconstructed from multiproxy analyses (Coleoptera, chironomids and pollen). Quat. Int. 636, 39–51. https://doi.org/10.1016/j.quaint.2022.02.005 (2022).
Fletcher, W. J. et al. Millennial-scale variability during the last glacial in vegetation records from Europe. Quat. Sci. Rev. 29, 2839–2864. https://doi.org/10.1016/j.quascirev.2009.11.015 (2010).
Ammann, B. et al. Vegetation responses to rapid warming and to minor climatic fluctuations during the Late-Glacial Interstadial (GI-1) at Gerzensee (Switzerland). Palaeogeogr. Palaeoclimatol. Palaeoecol. 391, 40–59. https://doi.org/10.1016/j.palaeo.2012.07.010 (2013).
Litt, T., Schölzel, C., Kühl, N. & Brauer, A. Vegetation and climate history in the Westeifel Volcanic Field (Germany) during the past 11 000 years based on annually laminated lacustrine maar sediments. Boreas 38, 679–690. https://doi.org/10.1111/j.1502-3885.2009.00096.x (2009).
Muschitiello, F. et al. Fennoscandian freshwater control on Greenland hydroclimate shifts at the onset of the Younger Dryas. Nat. Commun. 6, 8939. https://doi.org/10.1038/ncomms9939 (2015).
Rach, O., Brauer, A., Wilkes, H. & Sachse, D. Delayed hydrological response to Greenland cooling at the onset of the Younger Dryas in western Europe. Nat. Geosci. 7, 109–112. https://doi.org/10.1038/ngeo2053 (2014).
Hepp, J. et al. How dry was the Younger Dryas? Evidence from a coupled δ2H–δ18O biomarker paleohygrometer applied to the Gemündener Maar sediments, Western Eifel, Germany. Clim. Past 15, 713–733. https://doi.org/10.5194/cp-15-713-2019 (2019).
Sachse, D. et al. Molecular paleohydrology: Interpreting the hydrogen-isotopic composition of lipid biomarkers from photosynthesizing organisms. Annu. Rev. Earth Planet. Sci. 40, 221–249. https://doi.org/10.1146/annurev-earth-042711-105535 (2012).
Struck, J. et al. Leaf waxes and hemicelluloses in topsoils reflect the δ2H and δ18O isotopic composition of precipitation in Mongolia. Front. Earth Sci. https://doi.org/10.3389/feart.2020.00343 (2020).
Strobel, P. et al. The potential of δ2Hn-alkanes and δ18Osugar for paleoclimate reconstruction—A regional calibration study for South Africa. Sci. Total Environ. 716, 137045. https://doi.org/10.1016/j.scitotenv.2020.137045 (2020).
Hepp, J. et al. Evaluation of bacterial glycerol dialkyl glycerol tetraether and 2H–18O biomarker proxies along a central European topsoil transect. Biogeosciences 17, 741–756. https://doi.org/10.5194/bg-17-741-2020 (2020).
Hepp, J. et al. Reconstructing lake evaporation history and the isotopic composition of precipitation by a coupled δ18O–δ2H biomarker approach. J. Hydrol. 529, 622–631. https://doi.org/10.1016/j.jhydrol.2014.10.012 (2015).
Hou, J., D’Andrea, W. J. & Huang, Y. Can sedimentary leaf waxes record D/H ratios of continental precipitation? Field, model, and experimental assessments. Geochim. Cosmochim. Acta 72, 3503–3517. https://doi.org/10.1016/j.gca.2008.04.030 (2008).
Strobel, P. et al. Precipitation and lake water evaporation recorded by terrestrial and aquatic n-alkane δ2H isotopes in Lake Khar Nuur, Mongolia. Geochem. Geophys. Geosyst. 23, e2021GC010234. https://doi.org/10.1029/2021GC010234 (2022).
Bliedtner, M. et al. Late Holocene climate changes in the Altai Region based on a first high-resolution biomarker isotope record from Lake Khar Nuur. Geophys. Res. Lett. 1, 1. https://doi.org/10.1029/2021gl094299 (2021).
Aichner, B. et al. Hydroclimate in the Pamirs was driven by changes in precipitation-evaporation seasonality since the last glacial period. Geophys. Res. Lett. 46, 13972–13983. https://doi.org/10.1029/2019gl085202 (2019).
Tuthorn, M. et al. Coupling δ2H and δ18O biomarker results yields information on relative humidity and isotopic composition of precipitation—A climate transect validation study. Biogeosciences 12, 3913–3924. https://doi.org/10.5194/bg-12-3913-2015 (2015).
Bittner, L. et al. The Holocene lake-evaporation history of the afro-alpine Lake Garba Guracha in the Bale Mountains, Ethiopia, based on δ18O records of sugar biomarker and diatoms. Quat. Res. 105, 23–36. https://doi.org/10.1017/qua.2021.26 (2022).
Hepp, J. et al. A sugar biomarker proxy for assessing terrestrial versus aquatic sedimentary input. Organ. Geochem. 98, 98–104. https://doi.org/10.1016/j.orggeochem.2016.05.012 (2016).
Strobel, P. et al. Reconstructing Late Quaternary precipitation and its source on the southern Cape coast of South Africa: A multi-proxy paleoenvironmental record from Vankervelsvlei. Quaternary Science Reviews 284, 107467. https://doi.org/10.1016/j.quascirev.2022.107467 (2022).
Yang, Y., Zhang, Y., Zhang, H. & Huang, X. Quantitative reconstruction of the relative humidity by a coupled δ18O–δ2H approach during the Younger Dryas in central China. Quat. Sci. Rev. 299, 107879. https://doi.org/10.1016/j.quascirev.2022.107879 (2023).
Lemma, B. et al. δ2Hn-alkane and δ18Osugar biomarker proxies from leaves and topsoils of the Bale Mountains, Ethiopia, and implications for paleoclimate reconstructions. Biogeochemistry 153, 135–153. https://doi.org/10.1007/s10533-021-00773-z (2021).
Zech, M. et al. Revisiting the subalpine Mesolithic site Ullafelsen in the Fotsch Valley, Stubai Alps, Austria—New insights into pedogenesis and landscape evolution from leaf-wax-derived n-alkanes, black carbon and radiocarbon dating. E&G Quat. Sci. J. 70, 171–186. https://doi.org/10.5194/egqsj-70-171-2021 (2021).
Kuefner, W., Hofmann, A. M., Geist, J., Dubois, N. & Raeder, U. Algal community change in mountain lakes of the alps reveals effects of climate warming and shifting treelines. J. Phycol. 57, 1266–1283 (2021).
DWD. Climate Data Center—Monthly mean precipitation and air temperature for Kiefersfelden-Gach, Observation Germany. https://opendata.dwd.de/climate_environment/CDC/observations_germany/climate/monthly/ (2023).
Bowen, G. J. & Revenaugh, J. Interpolating the isotopic composition of modern meteoric precipitation. Water Resour. Res. 39, 23. https://doi.org/10.1029/2003wr002086 (2003).
Bowen, G. J., Wassenaar, L. I. & Hobson, K. A. Global application of stable hydrogen and oxygen isotopes to wildlife forensics. Oecologia 143, 337–348. https://doi.org/10.1007/s00442-004-1813-y (2005).
Mariani, I. et al. Temperature and precipitation signal in two Alpine ice cores over the period 1961–2001. Clim. Past 10, 1093–1108. https://doi.org/10.5194/cp-10-1093-2014 (2014).
Mayer, B. & Schwark, L. A 15,000-year stable isotope record from sediments of Lake Steisslingen, Southwest Germany. Chem. Geol. 161, 315–337. https://doi.org/10.1016/S0009-2541(99)00093-5 (1999).
Schäfer, I. K. et al. Leaf waxes in litter and topsoils along a European transect. SOIL 2, 551–564. https://doi.org/10.5194/soil-2-551-2016 (2016).
Schwörer, C. et al. Holocene climate, fire and vegetation dynamics at the treeline in the Northwestern Swiss Alps. Veg. Hist. Archaeobotany 23, 479–496. https://doi.org/10.1007/s00334-013-0411-5 (2014).
Rey, F. et al. Climate impacts on vegetation and fire dynamics since the last deglaciation at Moossee (Switzerland). Clim. Past 16, 1347–1367. https://doi.org/10.5194/cp-16-1347-2020 (2020).
Hepp, J. et al. Validation of a coupled δ2Hn-alkane–δ18Osugar paleohygrometer approach based on a climate chamber experiment. Biogeosciences 18, 5363–5380. https://doi.org/10.5194/bg-18-5363-2021 (2021).
Helliker, B. R. & Ehleringer, J. R. Establishing a grassland signature in veins: 18O in the leaf water of C3 and C4 grasses. Proc. Natl. Acad. Sci. 97, 7894 (2000).
McInerney, F. A., Helliker, B. R. & Freeman, K. H. Hydrogen isotope ratios of leaf wax n-alkanes in grasses are insensitive to transpiration. Geochim. Cosmochim. Acta 75, 541–554. https://doi.org/10.1016/j.gca.2010.10.022 (2011).
Gkinis, V. et al. A 120,000-year long climate record from a NW-Greenland deep ice core at ultra-high resolution. Sci. Data 8, 141. https://doi.org/10.1038/s41597-021-00916-9 (2021).
Dansgaard, W. Stable isotopes in precipitation. Tellus 16, 436–468. https://doi.org/10.1111/j.2153-3490.1964.tb00181.x (1964).
Lücke, A. & Brauer, A. Biogeochemical and micro-facial fingerprints of ecosystem response to rapid Late Glacial climatic changes in varved sediments of Meerfelder Maar (Germany). Palaeogeogr. Palaeoclimatol. Palaeoecol. 211, 139–155. https://doi.org/10.1016/j.palaeo.2004.05.006 (2004).
Schenk, F. et al. Warm summers during the Younger Dryas cold reversal. Nat. Commun. 9, 1634. https://doi.org/10.1038/s41467-018-04071-5 (2018).
Strobel, P., Struck, J., Zech, R. & Bliedtner, M. The spatial distribution of sedimentary compounds and their environmental implications in surface sediments of Lake Khar Nuur (Mongolian Altai). Earth Surf. Process. Landf. 46, 611–625. https://doi.org/10.1002/esp.5049 (2021).
Ilyashuk, B. et al. Lateglacial environmental and climatic changes at the Maloja Pass, Central Swiss Alps, as recorded by chironomids and pollen. Quat. Sci. Rev. 28, 1340–1353. https://doi.org/10.1016/j.quascirev.2009.01.007 (2009).
Ivy-Ochs, S., Monegato, G. & Reitner, J. M. in European Glacial Landscapes (eds David Palacios, Philip D. Hughes, José M. García-Ruiz, & Nuria Andrés) 525–539 (Elsevier, 2023).
Aichner, B., Herzschuh, U. & Wilkes, H. Influence of aquatic macrophytes on the stable carbon isotopic signatures of sedimentary organic matter in lakes on the Tibetan Plateau. Organ. Geochem. 41, 706–718. https://doi.org/10.1016/j.orggeochem.2010.02.002 (2010).
Liu, W. et al. Influence of aquatic plants on the hydrogen isotope composition of sedimentary long-chain n-alkanes in the Lake Qinghai region, Qinghai-Tibet Plateau. Sci. China Earth Sci. 59, 1368–1377. https://doi.org/10.1007/s11430-016-5263-2 (2016).
Andrae, J. W., McInerney, F. A. & Sniderman, J. M. K. Carbon isotope systematics of leaf wax n-alkanes in a temperate lacustrine depositional environment. Organ. Geochem. 150, 104121. https://doi.org/10.1016/j.orggeochem.2020.104121 (2020).
Zech, M. et al. A 16-ka δ18O record of lacustrine sugar biomarkers from the High Himalaya reflects Indian Summer Monsoon variability. J. Paleolimnol. 51, 241–251. https://doi.org/10.1007/s10933-013-9744-4 (2014).
Ogier, S., Disnar, J.-R., Albéric, P. & Bourdier, G. Neutral carbohydrate geochemistry of particulate material (trap and core sediments) in an eutrophic lake (Aydat, France). Organ. Geochem. 32, 151–162. https://doi.org/10.1016/S0146-6380(00)00138-8 (2001).
Flaim, G., Camin, F., Tonon, A. & Obertegger, U. Stable isotopes of lakes and precipitation along an altitudinal gradient in the Eastern Alps. Biogeochemistry 116, 187–198. https://doi.org/10.1007/s10533-013-9855-z (2013).
Aichner, B. et al. Spatial and seasonal patterns of water isotopes in northeastern German lakes. Earth Syst. Sci. Data 14, 1857–1867. https://doi.org/10.5194/essd-14-1857-2022 (2022).
IAEA/WMO. Global Network of Isotopes in Precipitation. The GNIP Database., https://nucleus.iaea.org/wiser (2019).
Cernusak, L. A., Wong, S. C. & Farquhar, G. D. Oxygen isotope composition of phloem sap in relation to leaf water in Ricinus communis. Funct. Plant Biol. 30, 1059–1070. https://doi.org/10.1071/FP03137 (2003).
Gessler, A. et al. Tracing carbon and oxygen isotope signals from newly assimilated sugars in the leaves to the tree-ring archive. Plant Cell Environ. 32, 780–795. https://doi.org/10.1111/j.1365-3040.2009.01957.x (2009).
Cernusak, L. A. et al. Do 2H and 18O in leaf water reflect environmental drivers differently?. New Phytol. 235, 41–51. https://doi.org/10.1111/nph.18113 (2022).
Sachse, D., Radke, J. & Gleixner, G. Hydrogen isotope ratios of recent lacustrine sedimentary n-alkanes record modern climate variability. Geochim. Cosmochim. Acta 68, 4877–4889. https://doi.org/10.1016/j.gca.2004.06.004 (2004).
Laskar, J. et al. A long-term numerical solution for the insolation quantities of the Earth. Astron. Astrophys. 428, 261–285. https://doi.org/10.1051/0004-6361:20041335 (2004).
Brauer, A., Haug, G. H., Dulski, P., Sigman, D. M. & Negendank, J. F. W. An abrupt wind shift in western Europe at the onset of the Younger Dryas cold period. Nat. Geosci. 1, 520–523. https://doi.org/10.1038/ngeo263 (2008).
Cabedo-Sanz, P., Belt, S. T., Knies, J. & Husum, K. Identification of contrasting seasonal sea ice conditions during the Younger Dryas. Quat. Sci. Rev. 79, 74–86. https://doi.org/10.1016/j.quascirev.2012.10.028 (2013).
Isarin, R. F. B., Renssen, H. & Vandenberghe, J. The impact of the North Atlantic Ocean on the Younger Dryas climate in northwestern and central Europe. J. Quat. Sci. 13, 447–453. https://doi.org/10.1002/(SICI)1099-1417(1998090)13:5%3c447::AID-JQS402%3e3.0.CO;2-B (1998).
Pauly, M. et al. Subfossil trees suggest enhanced Mediterranean hydroclimate variability at the onset of the Younger Dryas. Sci. Rep. 8, 13980. https://doi.org/10.1038/s41598-018-32251-2 (2018).
Szidat, S. et al. 14 C analysis and sample preparation at the New Bern laboratory for the analysis of radiocarbon with AMS (LARA). Radiocarbon 56, 561–566. https://doi.org/10.2458/56.17457 (2014).
Blaauw, M. & Christen, J. A. Flexible paleoclimate age-depth models using an autoregressive gamma process. Bayesian Anal. 6, 457–474. https://doi.org/10.1214/11-ba618 (2011).
Reimer, P. J. et al. The IntCal20 northern hemisphere radiocarbon age calibration curve (0–55 cal kBP). Radiocarbon 62, 725–757. https://doi.org/10.1017/rdc.2020.41 (2020).
Schneider, H. et al. Holocene estuary development in the Algarve Region (Southern Portugal)—A reconstruction of sedimentological and ecological evolution. Quat. Int. 221, 141–158. https://doi.org/10.1016/j.quaint.2009.10.004 (2010).
Zech, M. & Glaser, B. Compound-specific δ18O analyses of neutral sugars in soils using gas chromatography–pyrolysis–isotope ratio mass spectrometry: problems, possible solutions and a first application. Rapid Commun. Mass Spectrom. 23, 3522–3532. https://doi.org/10.1002/rcm.4278 (2009).
Acknowledgements
We thank Theresa Henning, Moritz Mäding and Samuel Getachew for their assistance in the lab. M.P. gratefully acknowledges the scholarship of the State of Thuringia (Landesgraduiertenstipendium). Ernst Kroemer and Torsten Haberzettl are thanked for their contribution during sediment coring.
Funding
Open Access funding enabled and organized by Projekt DEAL.
Author information
Authors and Affiliations
Contributions
M.P., R.Z., M.B., and M.Z. designed this study. M.P., M.B., J.S., and P.S. performed the biomarker analyses and H.S. performed the palynological analyses at Friedrich-Schiller-Universität Jena. M.P., P.S., L.B., and M.Z. performed the sugar analyses at Technische Universität Dresden and Friedrich-Schiller-Universität Jena. S.S. and G.S. performed radiocarbon analyses at University of Bern. M.P. wrote the manuscript with contributions of all coauthors.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Prochnow, M., Strobel, P., Bliedtner, M. et al. Summer paleohydrology during the Late Glacial and Early Holocene based on δ2H and δ18O from Bichlersee, Bavaria. Sci Rep 13, 18487 (2023). https://doi.org/10.1038/s41598-023-45754-4
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-023-45754-4
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
