
Abstract
Targeted metabolomics has been widely used in pheromone research but may miss pheromone components in study organisms that produce pheromones in trace amount and/or lack bio-detectors (e.g., antennae) to readily locate them in complex samples. Here, we used non-targeted metabolomics—together with high-performance liquid chromatography–mass spectrometry (HPLC–MS), gas chromatography-MS, and behavioral bioassays—to unravel the sex pheromone of the triangulate cobweb spider, Steatoda triangulosa. A ternary blend of three contact pheromone components [N-4-methylvaleroyl-O-isobutyroyl-l-serine (5), N-3-methylbutyryl-O-isobutyroyl-l-serine (11), and N-3-methylbutyryl-O-butyroyl-l-serine (12)] elicited courtship by S. triangulosa males as effectively as female web extract. Hydrolysis of 5, 11 and 12 at the ester bond gave rise to two mate-attractant pheromone components [butyric acid (7) and isobutyric acid (8)] which attracted S. triangulosa males as effectively as female webs. Pheromone components 11 and 12 are reported in spiders for the first time, and were discovered only through the use of non-targeted metabolomics and GC–MS. All compounds resemble pheromone components previously identified in widow spiders. Our study provides impetus to apply non-targeted metabolomics for pheromone research in a wide range of animal taxa.
Similar content being viewed by others
Introduction
Sexually reproducing organisms commonly attract or locate mates through sexual communication signals. Signals may be uni-, bi- or poly-modal with visual, chemical, acoustic, vibratory, and tactile characteristics1. Pheromones are thought to be the oldest type of sexual communication signals2. They are chemicals, or blends of chemicals, released by a signaler that cause a response by conspecific signal recipients3,4. Pheromones are prevalent in many animal taxa5, including insects6,7,8, myriapods9, crustaceans10, fish11, and mammals12, and may be airborne or substrate-borne and be sensed by olfactory receptors13 or contact chemoreceptors14. Regardless of their physicochemical characteristics, pheromone components commonly occur in complex blends of analyte and are not easily located, isolated, and identified15.
Metabolomics entails the systematic identification and quantitation of metabolites, and their changes over time, in biological samples. Analytical techniques include, but are not limited to, gas chromatography—mass spectrometry (GC–MS), and high-performance liquid chromatography—mass spectrometry (HPLC–MS). The choice of analytical technique is based, in part, on chemical characteristics of target compounds and their abundance in samples. During GC–MS and HPLC–MS analyses, chemicals are separated and broken into mass fragments (ions), resulting in a mass spectrum for each sufficiently abundant compound and in a total ion chromatogram (TIC) that is created by summing up intensities of all mass spectral peaks belonging to the same scan16. Mass spectra then provide information about the molecular structure of compounds16.
Comparative metabolomics in pheromone identification research compares analytes obtained from animals that were capable (e.g. sexually mature), or not (e.g. sexually immature), of producing pheromone15. Indeed, comparative metabolomics has been used to identify the first ever spider pheromone17. Traditional targeted metabolomics compares peaks between TICs and focuses on peaks for pheromone identification that are visually unique in one type of analyte (Fig. 1a)18,19. However, exclusive focus on visually unique peaks may miss pheromone components that co-elute with non-pheromonal compounds (Fig. 1b) or occur at trace quantities (Fig. 1c).

Graphical comparison of total ion chromatograms of a hypothetical case sample (upper trace) and a control sample (lower trace). (a) A unique compound (green) in the case sample is absent in the control sample. (b) A novel compound (blue) in the case sample is masked—and thus easily overlooked—by a compound (brown) present in both samples. (c) A unique trace compound (red) in the case sample might not be visually detected.
Pheromone research has focused on insects, with more than 3000 pheromones identified to date6. This remarkable progress is attributed to a landmark invention 53 years ago that combined gas chromatography with electrophysiology for insect pheromone analyses20,21. In these gas chromatographic-electroantennographic detection (GC-EAD) analyses, an insect’s antenna serves as a bio-detector to help locate candidate pheromone components in complex samples22. Moreover, in many insects, pheromone biosynthetic pathways and receptor sites are well understood13,23, and structural similarities of pheromones among congeners have expedited pheromone identifications24. In contrast, the chemical ecology of animals lacking antennae has hardly been studied, in part, because electrophysiological techniques were not applicable6,15,25,26.
Here, we applied non-targeted metabolomics (XCMS online)—together with GC–MS, HPLC–MS, and behavioral bioassays—to locate and identify both contact and mate-attractant pheromone components of a web building spider. XCMS online is a free and user-friendly metabolomics software (Scripps Research CA, USA)27 that enables analyses of data collected during mass spectrometric analyses such as GC–MS or HPLC–MS. Unlike targeted metabolomics, non-targeted metabolomics considers all detected ions and enables quantitative comparison of ions between samples. The software provides graphs and tables of ions as well as their relative abundance and retention times. This comprehensive approach reduces the probability of erroneously excluding peaks from analyses that are masked by other compounds (Fig. 1b) or occur at trace quantities (Fig. 1c).
Spiders have received little attention in chemical ecology research6. There are some 50,000 spider species but only 15 sex pheromones have been identified to date6,15,25, possibly because spiders lack antennae as pheromone bio-detectors (see above), and the search for pheromone receptors has met with limited success26,28,29. To assess how non-untargeted metabolomics can aid in spider pheromone research, we selected the triangulate cobweb spider, Steatoda triangulosa, a synanthropic, cosmopolitan spider inhabiting buildings30,31. We selected S. triangulosa, a tiny (3–6 mm) cob-web building spider32, because it belongs to a group of widow spiders (Latrodectinae) for which several pheromones have been identified18,33,34,35, anticipating that non-targeted metabolomics would help us find similar pheromone components in S. triangulosa.
Within the Latrodectinae, females deposit contact pheromone components on their webs that elicit courtship by males upon contact. During courtship, males cut and bundle up sections of the female’s web, adding their own silk in the process36. Contact sex pheromone components have been identified for females of the redback spider, Latrodectus hasselti [N-3-methylbutyroyl-O-(S)-2-methylbutyroyl-l-serine methyl ester (1)]18, the western black widow spider, L. hesperus [N-3-methylbutanoyl-O-isobutyroyl-l-serine methyl ester (2)]33, the brown widow, L. geometricus [N-3-methylbutyroyl-O-propionyl-l-serine-methyl ester (3)]35, and the false black widow spider, Steatoda grossa [N-4-methylvaleroyl-O-butyroyl-l-serine (4), N-4-methylvaleroyl-O-isobutyroyl-l-serine (5) and N-4-methylvaleroyl-O-hexanoyl-l-serine (6)]34 (Fig. 2a). In S. grossa, contact pheromone components—web pH-dependently—hydrolyse at the ester bond and give rise to airborne mate-attractant pheromone components: butyric acid (7), isobutyric acid (8), and hexanoic acid (9) (Fig. 2a)34. N-4-Methylvaleroyl-l-serine (10), as another hydrolysis breakdown product, accumulates on webs and has no pheromonal activity34.

Phylogeny and comparison of pheromone components (contact & airborne) in widow spiders (Latrodectinae). (a) Previously known pheromone components of Latrodectus hasselti18, L. hesperus33, L. geometricus35, and Steatoda grossa34: N-3-methylbutyroyl-O-(S)-2-methylbutyroyl-l-serine methyl ester (1), N-3-methylbutanoyl-O-isobutyroyl-l-serine methyl ester (2), N-3-methylbutyroyl-O-isobutyroyl-l-serine-methyl ester (3), N-4-methylvaleroyl-O-butyroyl-l-serine (4), N-4-methylvaleroyl-O-isobutyroyl-l-serine (5), and N-4-methylvaleroyl-O-hexanoyl-l-serine (6). The contact pheromone components 4–6 of S. grossa hydrolyse at the ester bond and give to three airborne mate-attractant pheromone components [butyric acid (7), isobutyric acid (8), and hexanoic acid (9)], whereas the amide N-4-methylvaleroyl-l-serine (10), as another hydrolysis breakdown product, remains on webs and has no behavioural activity. (b) Pheromone components of Steatoda triangulosa identified in this study. The contact pheromone components N-4-methylvaleroyl-O-isobutyroyl-l-serine (5), N-3-methylbutyroyl-O-isobutyroyl-l-serine (11), and N-3-methylbutyroyl-O-butyroyl-l-serine (12) hydrolyse at the ester bond and give rise to two airborne mate-attractant pheromone components [butyric acid (7) and isobutyric acid (8)], whereas N-4-methylvaleroyl-l-serine (10) and N-3-methyl-butyroyl-l-serine (13) accumulate on webs. Blue-coloured parts of molecules are phylogenetically conserved, whereas green-coloured parts are unique to Steatoda. Orange parts are shared between Latrodectus spp. and S. triangulosa.
In this study, we applied four analytical tools—non-targeted metabolomics (XCMS online), HPLC–MS, GC–MS, and behavioural bioassays—to identify the contact and mate-attractant pheromone components of S. triangulosa. We demonstrate that these tools in combination, but not on their own, provided decisive analytical capability to unravel the complete pheromonal communication system of S. triangulosa.
Methods
Spider rearing and web collection
Steatoda triangulosa spiders used in experiments were the F1 and F3 offspring of females collected at the Black Widow Winery [Penticton, British Columbia (B.C.), Canada, 49.5467° N, 119.5698° W]. The spiders were reared in the insectary of the Burnaby campus of Simon Fraser University at 22 °C under a reversed light cycle (12:12 h). Spiderlings were kept individually in petri dishes (100 × 20 mm)37 fitted with moist cotton, and fed Drosophila melanogaster vinegar flies once per week. Virgin females and naïve adult males were randomly chosen for web-building and bioassays, respectively. As female spiders produce little silk per day, we extended the web-building period, allowing each female to build her web for seven days, instead of three days34, on a triangular prism (10 × 10 × 10 cm) of bamboo skewers (Goodcook, USA)36. Webs were collected using a methanol-cleaned glass rod and were extracted overnight in 25 µL/web of acetonitrile (ACN, 99%, Sigma-Aldrich, USA). All behavioral bioassays were run during the reversed scotophase (0900 to 1700).
Analyses of web extracts by high-performance liquid chromatography—mass spectrometry (HPLC–MS)
Aliquots (2 µL) of web extracts of adult and subadult female S. triangulosa were analyzed, and compared, using coupled high-performance liquid chromatography—mass spectrometry (HPLC/MS). The Bruker maXis Impact Quadrupole Time-of-Flight LC/MS system consisted of an Agilent 1200 LC fitted with a Spursil C18 column (30 mm × 3.0 mm, 3 µm; Dikma Technologies, Foothill Ranch, CA, USA) and a Bruker maXis Impact Ultra-High Resolution tandem TOF (UHR-Qq-TOF) mass spectrometer. The LC/MS was operated with positive electrospray ionisation (+ ESI) at a gas temperature of 200 °C and a flow of 9 L/min. The nebuliser was set to 4 bar and the capillary voltage to 4200 V. The column was eluted with a 0.4 mL/min flow of a solvent gradient, starting with 80% water and 20% acetonitrile, and ending with 100% acetonitrile after 4 min. The solvent system contained 0.1% formic acid to improve the peak shape of compounds.
Analyses of web extracts by gas chromatography—mass spectrometry (GC–MS)
Web extracts of adult virgin females were also analyzed by coupled gas chromatography—mass spectrometry (GC–MS), using an Agilent 7890B GC fitted with a DB-5 GC–MS column (30 m × 0.25 mm ID, film thickness 0.25 µm) and coupled to a 5977 A MSD. The injector port of the GC was set to 250 °C, the transfer line to 280 °C, the MS source to 230 °C, and the MS quadrupole to 150 °C. Helium was used as the carrier gas at a flow rate of 35 cm s−1. The following temperature program was used: 50 °C held for 5 min, a 10 °C/min increase to 280 °C (held for 10 min). Compounds were identified by comparing their mass spectra and retention indices with those of authentic standards that were synthesized in our laboratory. To improve chromatography of potential acids in web extracts, acids were transformed into trimethylsilyl-esters using BSTFA prior to analyses38.
Analyses of web extracts by XCMS online
In search of further pheromone components, LC–MS analyses of web extracts of 10 adult females and 10 subadult female S. triangulosa were compared by XCMS online. The following parameters were used for the pairwise comparison: bw = 5, ppm = 10, peak width = c(2, 20), mzwidth = 0.01, and mzdiff = 0.01. Detected masses were sorted by “fold change” between the two groups and an arbitrary fold change of greater 35 × was selected as the threshold (see suppl. table).
Behavioural testing of contact pheromone components—T-rod bioassays
General experimental design
The ability of web extract and of specific candidate contact pheromone components to induce courtship by male S. triangulosa was tested in T-rod bioassays, drawing on an established protocol34,37,39. The T-rod apparatus consisted of a horizontal beam (8 × 0.4 cm) and a vertical beam (8 × 0.4 cm) held together by labelling tape (3 × 1.9 cm, Fisher Scientific, Ottawa, ON, CA). A piece of filter paper (2 cm2) was attached to each distal end of the horizontal beam. The vertical beam of T-rods was inserted into plasticine (Craftsmart, Irving, TX, USA) placed in a tray (45 × 35 × 2.5 cm) partially filled with water to prevent spider males from escaping.
For each bioassay, three web equivalents in ACN or synthetic candidate pheromone components in ACN at three web-equivalents were applied to the randomly assigned treatment filter paper, whereas ACN was applied to the control filter paper. Using ACN as a corresponding control stimulus allowed us to account for potential solvent effects on male spider behaviour, and to demonstrate that male spiders do not court in response to ACN only. ACN was allowed to evaporate for 1 min before the onset of a 15-min bioassay. A randomly selected naïve male spider was then placed at the base of the vertical beam, and the time he spent courting on each filter paper was recorded. In response to the presence of female-produced or synthetic pheromone on a filter paper, the male engaged in courtship, pulling silk with his hindlegs from his spinnerets and adding it to the paper. Sensing contact pheromone, the male essentially behaves as if he were courting on the web of a female. Replicates of experiments as part of specific research objectives were run in parallel to eliminate day effects on responses of spiders. Treatment and control arms were alternated between replicates. T-rods and filter paper were discarded after use.
Specific experiments
The effect of web extract and of specific candidate contact pheromone components on courtship behaviour by male S. triangulosa was tested in three sets of T-rod bioassays. In set 1 (summer 2019), parallel experiments 1 and 2 (n = 20 each) tested web extract (three web equivalents in ACN) versus an ACN control (Exp. 1), and synthetic candidate contact pheromone component 5 versus an ACN control (Exp. 2). Synthetic 5 was tested at the same amount (387 ng) as present in three web extract equivalents. In set 2 (summer 2022), parallel experiments 3 and 4 (n = 20 each) tested web extract (three web equivalents in ACN) versus an ACN control (Exp. 3), and a blend of synthetic candidate contact pheromone components 5 (239 ng), 11 (8 ng) and 12 (11 ng) versus an ACN control. In set 3 (summer 2022), parallel experiments 5–8 tested 5, 11 and 12 in ternary combination (Exp. 5; lure composition as in Exp. 4; total lure dose: 259 ng) and singly (Exps. 6–8), each at 259 ng.
Chemical inferences and calculations of mate-attractant pheromone components
Drawing on previous findings that the contact pheromone components of S. grossa hydrolyse at the ester bond and give rise to acid mate-attractant pheromone components (Fig. 2b)34, we predicted that the contact pheromone components 5, 11 and 12 of S. triangulosa would also hydrolyse and release butyric acid (7) and isobutyric acid (8) as mate-attractant pheromone components. However, we could not detect 7 and 8 in GC–MS analysis of silyl-ester derivatized web extract, and needed to estimate 7 and 8 based on amounts of the amide breakdown products 10 (N-4-methylvaleroyl-l-serine) and 13 (N-3-methylbutanoyl-l-serine) that originate from the hydrolysis and remain on webs (Fig. 2). With 10 quantified at 272 ng (1.34 mol) per web, and anticipating equal stoichiometric amounts of 8 and 10, we decided to bioassay 8 at 124 ng per web. Moreover, with 13 not quantifiable in web extracts, we inferred the amount of 7 (5 ng) based on the 4% of 12 (which gives rise to 7) in the 3-component contact pheromone blend.
Sources of synthetic chemicals
Butyric acid (7) and isobutyric acid (8) (both 99% chemically pure) were purchased from Sigma-Aldrich (USA), N-4-methylvaleroyl-O-isobutyroyl-l-serine (5) was available from previous work in our laboratory34, and N-3-methylbutyroyl-O-isobutyroyl-l-serine (11), and N-3-methylbutyroyl-O-butyroyl-l-serine (12) were synthesized following the established synthesis of acylated serine derivates18,33,34. We report the 1H and 13C NMR spectra of 11 and 12, as well as the mass spectra of esterified 11 and 12, in the Supplementary Information.
Behavioural testing of mate-attractant pheromone components—olfactometer bioassays
Attraction of males to webs of mature females spiders and to synthetic candidate mate-attractant pheromone components 7 (butyric acid) and 8 (isobutyric acid) was tested in still-air dual-choice olfactometers (winter 2022)40. Large Plexiglass arenas (180 cm × 12 cm × 13 cm, , Fig. 3h)40 lined with printer plot recorder paper (180 × 13 cm, Agilent, Santa-Clara, CA, USA) served as olfactometers. Two wooden prisms bearing (i) a female web or no web (Exp. 9, n = 30), or (ii) artificial (Halloween) web (45.05 ± 0.4 mg)41 treated with synthetic 7 (5 ng) and 8 (124 ng) (Sigma-Aldrich) in ACN (75 µL), or an ACN control (75 µL) (Exp. 10, n = 30), were placed at opposite ends of the arena. For each bioassay, a single naïve male spider was placed into the centre of the arena, and allowed 30 min to approach and contact a prism, a behavioural response recorded as first choice. After each bioassay, the paper lining, webbing, and prisms were discarded. Treatment and control sides were alternated between replicates, and the same number of replicates was run for each of two experiments to eliminate potential day effects. Each male was tested only once.

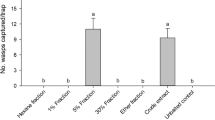
Chromatograms, experimental designs, and behavioural bioassay results. (a) Total ion chromatogram (TIC) of web extract of female Steatoda triangulosa analysed by high-performance liquid chromatography—mass spectrometry. (b) TIC of silyl ester-derivatized web extract of female S. triangulosa analysed by gas chromatography—mass spectrometry. (c) Comparative XCMS online Cloud Plots of web extracts of mature and immature female S. triangulosa (depicted by solid and dotted lines, respectively), with circles denoting a > 35-fold abundance increase of fragment ions in compounds; the larger the circle, the greater the fold-change of a particular ion. (d) T-rod bioassay apparatus. (e) Effects of female S. triangulosa web extract (Exp. 1) and contact pheromone component 5 (N-4-methylvaleroyl-O-isobutyroyl-l-serine) (Exp. 2) on courtship by S. triangulosa males. (f) Effects of female S. triangulosa web extract (Exp. 3), and a ternary blend of contact pheromone components 5, 11 (N-3-methylbutyoyl-O-isobutyroyl-l-serine, and 12 (N-3-methylbutyroyl-O-butyroyl-l-serine) (Exp. 4), on courtship by S. triangulosa males. (g) Effects of contact pheromone components 5, 11 and 12 presented in ternary combination (Exp. 5), and singly (Exps. 6–8), on courtship by S. triangulosa males. (h) Arena olfactometer with prisms carrying test stimuli. (i) Attraction of male S. triangulosa to webs of female S. triangulosa (Exp. 9), and to synthetic mate-attractant pheromone components 7 (butyric acid) and 8 (isobutyric acid) in arena olfactometers. In each of subpanels e–g, different letters indicate statistical differences between test stimuli across experiments (rank sum test; p < 0.05). In experiments 9 and 10 (subpanel i), the asterisk (*) indicates a significant preference for the test stimulus (binomial test; p < 0.05).
Statistical analysis
We analysed data using R (v. 4.3.1) and R-studio (v. 2303.06.0). A Wilcoxon Rank Sum test was used to compare the amount of time male S. triangulosa spent courting on filter paper treated with web extract (Exps. 1, 3) or a synthetic pheromone blend (Exps. 2, 4). A Kruskal–Wallis Rank Sum test with Benjamini–Hochberg correction was used to compare the amount of time male S. triangulosa spent courting on filter paper treated with a ternary pheromone blend or single components (Exps. 4–8). A one-sided binomial test34 with Benjamini–Hochberg correction for multiple testing was used to test effects of spider webs (Exp. 9), or a binary blend of synthetic mate-attractant pheromone components (Exp. 10), on attraction of male S. triangulosa.
Results
Analyses of web extracts by HPLC–MS, GC–MS and XCMS online
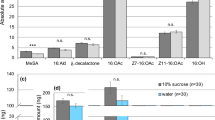
HPLC–MS analyses of web extract revealed a candidate contact pheromone component with fragment ions 296.1542 (M + Na), 274.1725 (M + 1) and 186.1179 (Fig. S1), matching the fragment ions and retention times of synthetic co-eluting 4 (N-4-methylvaleroyl-O-butyroyl-l-serine) and 5 (N-4-methylvaleroyl-O-isobutyroyl-l-serine)37. Coelution of 4 and 5 made it difficult to determine which compound was present but subsequent GC–MS analysis of esterified web extract determined that the compound was 5 (Fig. S3).
XCMS online confirmed the presence of 5 in web extracts (Fig. 3b). As female spiders progressed from subadults to adults, compound 5 ions 186.1179 and 274.1725 in web extracts increased 560-fold and 341-fold, respectively (Fig. 3b; Fig. S6). Another unknown compound (X), with retention time 4.42 min and fragment ion 260.1559, increased 37-fold (Fig. 3b).
GC–MS analysis of esterified extract in selected ion monitoring mode, searching for the 331.1815 ion of unknown X, revealed two isomers with retention time 21.05 min and 21.52 min. These isomers were identified as the trimethylsilyl-derivatives of N-3-methylbutyroyl-O-isobutyroyl-l-serine (11) (Fig. S4) and N-3-methylbutyroyl-O-butyroyl-l-serine (12) (Fig. S5).
Behavioural testing of contact pheromone components—T-rod bioassays
Web extract of adult females elicited more sustained courtship behaviour than synthetic 5 as a single contact pheromone component (W = 272, p = 0.026; Fig. 3e, Exps. 1, 2), indicating the presence of additional contact pheromone components in web extracts. However, the ternary blend of contact pheromone components 5, 10 and 11 was as effective as web extract in eliciting courtship behaviour by males (W = 215.5, p = 0.665; Fig. 3f, Exps. 3, 4), indicating that all essential components were present in the synthetic blend. The duration of male courtship on filter paper treated with 5, 10 and 11 as a ternary blend, and singly, differed (χ2 = 14.52, df = 3, p < 0.001, Fig. 3g, Exps. 5–8). Statistically (but not numerically), 5 was as effective as the ternary blend, and more effective than 12 but not than 11, in prompting and sustaining courtship by males (Fig. 3g).
Behavioural testing of mate-attractant pheromone components—olfactometer bioassays
In arena olfactometers (Fig. 3h), prisms bearing webs of a female spider attracted more males than empty prisms (19 vs 9, N = 30, p = 0.043; Fig. 3i, Exp. 9), indicating the dissemination of airborne mate-attractant pheromone components from webs. Similarly, prisms bearing artificial (Halloween) web treated with synthetic mate-attractant pheromone components 7 and 8 attracted more males than prisms bearing Halloween web treated with a corresponding solvent control (17 vs 7, N = 30, p = 0.043; Fig. 3i, Exp. 10), indicating that 7 and 8 are the essential, and likely the only, mate-attractant pheromone components of S. triangulosa.
Discussion
Our study provides proof of concept that a comprehensive analytical approach, entailing non-targeted metabolomics (XCMS online), HPLC–MS, GC–MS, and behavioural bioassays, was effective for unravelling the sex pheromone of the web-building spider S. triangulosa.
Whereas contact pheromone component 5 (N-4-methylvaleroyl-O-isobutyroyl-l-serine) would have been detected by conventional HPLC–MS, contact pheromone components 11 (N-3-methyl-butyryl-O-propionyl-l-serine) and 12 (N-3-methyl-butyryl-O-butyroyl-l-serine) were discovered only through the combined application of XCMS online, GC–MS, and behavioural bioassays. Through XCMS, we found that the fragment ion 260.1559 of an unknown compound (X) was 37-fold more abundant in web extracts of adult females than in web extracts of subadult females. GC–MS analyses of esterified extract, selectively scanning for the indicative ion of X, then revealed that X consisted of two isomers: 11 and 12. In T-rod behavioural bioassays with male S. triangulosa, a ternary blend of synthetic 5, 11, and 12 was as effective as adult female web extract in eliciting courtship by males, indicating that all essential contact pheromone components were present in the synthetic blend.
Metabolomics has become a routine analytical tool to screen samples for the presence or relative abundance of compounds in ‘case’ samples relative to reference (control) samples15,27. Non-targeted metabolomics has been applied e.g. in studies of diet and health43, sport and exercise44, host-microbiota45, drug discovery46, plant metabololisms47, organismal responses to environmental toxicants48, and biomarker discovery in disease diagnosis49.
Non-targeted metabolomics as a pheromone research tool seems to have been used in only a recent single study50. Liu et al.50 hypothesized that the uropygial gland of ducks secretes chemicals that mediate sexual communication. Using LC–MS and principal component analyses, the authors found numerous metabolites in gland secretions, and noticed a gender-bias in metabolite secretions. Five compounds, in particular, were significantly more abundant in secretions of males than females: picolinic acid, 3-hydroxypicolinic acid, indolacetaldehyde, 3-hydroxymethyl-glutaric acid, and 3-methyl-2-oxovaleric acid. Although this male-biased abundance of compounds is helpful to select candidate sex pheromone components, their potential pheromonal signal function has still to be demonstrated in behavioural bioassays. In our S. triangulosa study, we applied non-targeted metabolomics, together with mass spectrometry and behavioural bioassays, to unravel new contact pheromone components and demonstrate their pheromonal signal function. To some extent, non-targeted metabolomics substituted for the analytical function of antennal bio-detectors that are lacking in spiders.
Prior pheromone chemistry knowledge of theridiid widow spiders (Fig. 2a)18,33,34,35 aided the identification of the S. triangulosa sex pheromone. Steatoda triangulosa and S. grossa share N-4-methylvaleroyl-O-isobutroyl-l-serine (5) as a contact pheromone component, but 5 is a minor component in S. grossa and the major component in S. triangulosa (Fig. 3). The two minor components of S. triangulosa—N-3-methylbutyroyl-O-propionyl-l-serine (11) and N-3-methylbutyroyl-O-butyroyl-l-serine (12)—are reported here for the first time as spider pheromone components. Notably, all currently known contact pheromone components of Latrodectus and Steatoda are acylated serine derivates with a conserved N-amide-O-ester core. Whereas pheromone components of female Latrodectus spp. have a methyl ester functionality and an N-4-methylvaleroyl-l-serine amide (10) rest, pheromone components of S. grossa and S. triangulosa have a free carboxylic acid—instead of a methyl ester—and either a N-4-methylvaleroyl-l-serine amide rest or a N-3-methyl-butyroyl-l-serine amide (13) rest (Fig. 2). All data combined reveal astounding structural similarity between theridiid pheromones, and imply a shared biosynthetic pathway. However, despite their common N-amide-O-ester serine motif, Latrodectus and Steatoda pheromones have unique characteristics that support the taxonomic assignment of these spiders to different genera51. While it has long been known that insect congeners produce structurally related pheromones, as shown in Lymantria moths52,53,54 and Dendroctonus bark beetle55, our study reveals an analogous phenomenon in two genera of web-building widow spiders.
In S. grossa, the contact pheromone components 4, 5 and 6—web pH-dependently—hydrolyze at the ester bond, giving rise to the airborne mate-attractant pheromone components butyric acid (7), isobutyric acid (8) and hexanoic acid (9) (Fig. 2)34. We have shown that this hydrolysis is likely catalyzed by a web-borne carboxyl ester hydrolase34, but did not search for this type of enzyme on webs of S. triangulosa, anticipating a similar pheromone breakdown mechanism. We inferred that the hydrolysis of both 5 (N-4-methylvaleroyl-O-isobutyroyl-l-serine) and 11 (N-3-methylbutanoyl-O-isobutyroyl-l-serine) would release isobutyric acid and that the hydrolysis of 12 (N-3-methylbutanoyl-O-butyroyl-l-serine) would release butyric acid as mate-attractant pheromone components (Fig. 2b). This inference proved correct because a binary blend of synthetic 7 and 8 attracted S. triangulosa males in arena olfactometers as effectively as female webs (Fig. 3i). The release of 7 and 8 as mate-attractant pheromone components is reminiscent of pheromonal communication in the spider Linyphia triangularis17, where both the dimer (R)-3-[(R)-3-hydroxybutyryloxy]-butyric acid and its breakdown monomer (R)-3-hydroxybutyric acid induce courtship by males, and where the monomer likely also attracts males56.
Non-targeted metabolomics was the key analytical tool to help us locate trace candidate pheromone components in complex S. triangulosa web extracts. However, many other analytical tools were still required for pheromone identification, with each tool contributing to the identification process. Moreover, various analytical steps of the identification process were carefully informed through behavioural bioassays with spiders. A similar analytical approach—albeit not yet including non-targeted metabolomics—was recently taken for the identification of the S. grossa sex pheromone34. Historically, however, bioassay-guided pheromone identifications, with the application of multiple sophisticated analytical tools, were pioneered in insects. The study unveiling the oviposition-deterring pheromone of the coccinellid beetle Cheilosomones sexmaculata exemplifies this type of multi-tool analytical approach57.
The integration of insect antennae as pheromone bio-detectors in GC-EAD analyses of complex odor samples58 has expedited insect pheromone identifications and enabled the discovery of trace pheromone components59. In contrast, pheromone research in animal taxa that lack antennae, such as spiders, or that rely on olfactory epithelia in nasal cavities and/or on vomeronasal organs for odor reception, such as mammals, reptiles and amphibians5, has progressed at a much slower pace6. Pheromones are known for only 15 spiders15,34, and for relatively few mammals60, reptiles61, and amphibians62. This paucity of progress may be attributed to the fact that olfactory receptors are not known for spiders29,63,64, or are deemed fully functional for vertebrates only in-vitro, which would require challenging preparations for pheromone research. However, there is now emerging evidence that metabolomics may be able—to some extent—to assume the analytical role of antennal bio-detectors in pheromone research. Non-targeted metabolomics was effectively applied to help select candidate pheromone components of ducks50 and spidersthis study, both groups lacking antennae or equivalently effective bio-detectors for pheromone tracing. As a result, there is incentive now to apply non-targeted metabolomics for pheromone research in marine and terrestrial mammals, fish, birds, reptiles and amphibians, or even in signaling studies within and among plants.
In conclusion, we have applied non-targeted metabolomics, in combination with HPLC–MS, GC–MS, and behavioural bioassays, to unravel the sex pheromone of S. triangulosa. In this proof-of-concept study, we demonstrate that these four tools in combination, but not on their own, provide the analytical resolution to unravel the complete pheromonal communication system of a web-building spider. Our study provides impetus to take a similar analytical approach for pheromone research in other taxa that lack antennae or have odor receptors deemed fully functional only in-vivo.
Data availability
All data and code used in this manuscript have been added to the suppl. materials.
References
Clutton-Brock, T. Sexual selection in males and females. Science 318, 1882–1885 (2007).
Breithaupt, T. & Thiel, M. Chemical Communication in Crustaceans (Springer, 2011).
Karlson, P. & Lüscher, M. ‘Pheromones’: A new term for a class of biologically active substances. Nature 183, 55–56 (1959).
Wyatt, T. D. Primer pheromones. Curr. Biol. 27, 739–743 (2017).
Wyatt, T. D. Pheromones and Animal Behavior. University of Oxford (Cambridge University Press, 2014). https://doi.org/10.1017/CBO9781139030748.
Symonds, M. R. E. & Elgar, M. A. The evolution of pheromone diversity. Trends Ecol. Evolut. 23, 220–228 (2008).
Francke, W. & Schulz, S. Pheromones of terrestrial invertebrates. in Comprehensive Natural Products II (eds Mander, L. & Liu, H.-W.) 153–223 (2010).
Ayasse, M., Stökl, J. & Francke, W. Chemical ecology and pollinator-driven speciation in sexually deceptive orchids. Phytochemistry 72, 1667–1677 (2011).
Littlewood, P. M. H. The chemosensory behaviour of Lithobius forficatus (Myriapoda: Chilopoda). 2. Bioassay and chemistry of the coxal pheromone. J. Zool. 215, 523–535 (1988).
Beauche, F. & Richard, F.-J. The best timing of mate search in Armadillidium vulgare (Isopoda, Oniscidea). PloS One 7, e57737 (2013).
Scott, A. M. et al. Spermine in semen of male sea lamprey acts as a sex pheromone. PLoS Biol. 17, 1–24 (2019).
Varner, E., Gries, R., Takács, S., Fan, S. & Gries, G. Identification and field testing of volatile components in the sex attractant pheromone blend of female house mice. J. Chem. Ecol. 45, 18–27 (2019).
Vosshall, L. B., Amrein, H., Morozov, P. S., Rzhetsky, A. & Axel, R. A spatial map of olfactory receptor expression in the Drosophila antenna. Cell 96, 725–736 (1999).
Caskey, J. L., Watson, G. M. & Bauer, R. T. Studies on contact sex pheromones of the caridean shrimp Palaemonetes pugio: II. The role of glucosamine in mate recognition. Invertebr. Reprod. Dev. 53, 105–116 (2009).
Fischer, A. Chemical communication in spiders—A methodological review. J. Arachnol. 47, 1–27 (2019).
Harris, D. C. Quantitative Chemical Analysis (W. H. Freeman and Company, 2016).
Schulz, S. & Toft, S. Identification of a sex pheromone from a spider. Science 260, 1635–1637 (1993).
Jerhot, E., Stoltz, J. A., Andrade, M. C. B. & Schulz, S. Acylated serine derivatives: A unique class of arthropod pheromones of the Australian Redback spider, Latrodectus hasselti. Angew. Chem. 49, 2037–2040 (2010).
Sramkova, A., Schulz, C., Twele, R., Francke, W. & Ayasse, M. Fertility signals in the bumblebee Bombus terrestris (Hymenoptera: Apidae). Naturwissenschaften 95, 515–522 (2008).
Moorhouse, J. E., Yeadon, R., Beevor, P. S. & Nesbitt, B. F. Method for use in studies of insect chemical communication. Nature 223, 1174–1175 (1969).
Arn, H., Städler, E. & Rauscher, S. The electroantennographic detector—A selective and sensitive tool in the gas chromatographic analysis of insect pheromones. Z. Naturforsch. 30, 722–725 (1975).
Ma, T. et al. Analysis of tea geometrid (Ectropis grisescens) pheromone gland extracts using GC-EAD and GC×GC/TOFMS. J. Agric. Food Chem. 64, 3161–3166 (2016).
Blomquist, G. J. & Vogt, R. G. Insect Pheromone Biochemistry and Molecular Biology (Academic Press, 2003).
Holman, L., Lanfear, R. & D’Ettorre, P. The evolution of queen pheromones in the ant genus Lasius. J. Evolut. Biol. 26, 1549–1558 (2013).
Schulz, S. Spider pheromones—A structural perspective. J. Chem. Ecol. 39, 1–14 (2013).
Tichy, H., Gingl, E., Ehn, R., Papke, M. & Schulz, S. Female sex pheromone of a wandering spider (Cupiennius salei): Identification and sensory reception. J. Comp. Physiol. A 187, 75–78 (2001).
Tautenhahn, R., Patti, G. J., Rinehart, D. & Siuzdak, G. XCMS online: A web-based platform to process untargeted metabolomic data. Anal. Chem. 84, 5035–5039 (2012).
Fischer, A., Schulz, S., Ayasse, M. & Uhl, G. Pheromone communication among sexes of the garden cross spider Araneus diadematus. Sci. Nat. 108, 1–11 (2021).
Müller, C. H. G., Ganske, A. S. & Uhl, G. Ultrastructure of chemosensory tarsal tip-pore sensilla of Argiope spp. Audouin, 1826 (Chelicerata: Araneae: Araneidae). J. Morphol. 281, 1634–1659 (2020).
Bradley, R. A. Common Spiders of North America (University of California Press, 2012).
Bellmann, H. Der Kosmos-Spinnenführer (Kosmos, 2010).
Braun, R. Zur Biologie von Teutana triangulosa. Z. wiss. Zool. 159, 255–318 (1956).
Scott, C., McCann, S., Gries, R., Khaskin, G. & Gries, G. N-3-methylbutanoyl-O-methylpropanoyl-l-serine methyl ester—Pheromone component of western black widow females. J. Chem. Ecol. 41, 465–472 (2015).
Fischer, A. et al. Origin, structure and functional transition of sex pheromone components in a false widow spider. Commun. Biol. 5, 1156 (2022).
Baruffaldi, L. Function and diversity of sex pheromones in representative species of the black widow spiders (genus Latrodectus, Araneae: Theridiidae) (University of Toronto, 2016).
Fischer, A. et al. Multimodal and multifunctional signaling?—Web reduction courtship behavior in a North American population of the false black widow spider. PloS One 15, e0228988 (2020).
Fischer, A., Lee, Y., Stewart, J. & Gries, G. Dodging sexual conflict?—Sub-adult females of a web-building spider stay cryptic to mate-seeking adult males. Ethology 124, 838–843 (2018).
Stalling, D., Gehrke, C. W. & Zumwalt, R. W. A new silylation reagent for amino acids bis (trimethylsilyl) trifluoroacetamide (BSTFA). Biochem. Biophys. Res. Commun. 31, 616–622 (1968).
Scott, C., Gerak, C., McCann, S. & Gries, G. The role of silk in courtship and chemical communication of the false widow spider, Steatoda grossa (Araneae: Theridiidae). J. Ethol. 36, 191–197 (2018).
Gries, R. et al. Bed bug aggregation pheromone finally identified. Angew. Chem. Int. Ed. 54, 1135–1138 (2015).
Fischer, A., Hung, E. & Gries, G. Female false black widow spiders, Steatoda grossa, recognize webs based on physical and chemical cues. Entomol. Exp. Appl. 167, 803–810 (2019).
Ruxton, G. D. & Neuhäuser, M. When should we use one-tailed hypothesis testing?. Methods Ecol. Evolut. 1, 114–117 (2010).
Zhang, X., Li, Q., Xu, Z. & Dou, J. Mass spectrometry-based metabolomics in health and medical science: A systematic review. RSC Adv. 10, 3092–3104 (2020).
Heaney, L. M., Deighton, K. & Suzuki, T. Non-targeted metabolomics in sport and exercise science. J. Sports Sci. 37, 959–967 (2019).
Zhao, X. et al. Host-microbiota interaction-mediated resistance to inflammatory bowel disease in pigs. Microbiome 10, 115 (2022).
Alarcon-Barrera, J. C., Kostidis, S., Ondo-Mendez, A. & Giera, M. Recent advances in metabolomics analysis for early drug development. Drug Discov. Today 27, 1763–1773 (2022).
Anzano, A., Bonanomi, G., Mazzoleni, S. & Lanzotti, V. Plant metabolomics in biotic and abiotic stress: A critical overview. Phytochem. Rev. 21, 503–524 (2022).
Liu, L. et al. Study on toxicity effects of environmental pollutants based on metabolomics: A review. Chemosphere 286, 131815 (2022).
Baima, G. et al. Salivary metabolomics for the diagnosis of periodontal diseases: A systematic review with methodological quality assessment. Metabolomics 17, 1 (2021).
Liu, H. et al. Metabolic profiling reveals that the olfactory cues in the duck uropygial gland potentially act as sex pheromones. Animals 12, 413 (2022).
Liu, J., May-Collado, L. J., Pekár, S. & Agnarsson, I. A revised and dated phylogeny of cobweb spiders (Araneae, Araneoidea, Theridiidae): A predatory cretaceous lineage diversifying in the era of the ants (Hymenoptera, Formicidae). Mol. Phylogenet. Evolut. 94, 658–675 (2016).
Gries, G. et al. Specificity of nun and gypsy moth sexual communication through multiple-component pheromone blends. Naturwissenschaften 83, 382–385 (1996).
Gries, G. et al. 2-Methyl-(Z)-7-octadecene: Sex pheromone of allopatric Lymantria lucescens and L. serva. J. Chem. Ecol. 28, 469–478 (2002).
Gries, R., Khaskin, G., Gotoh, T., Schaefer, P. W. & Gries, G. (7Z,9E)-2-Methyl-7,9-octadecadiene: A sex pheromone component of Lymantria bantaizana. J. Chem. Ecol. 31, 879–891 (2005).
Symonds, M. R. E. & Elgar, M. A. The mode of pheromone evolution: Evidence from bark beetles. Proc. R. Soc. B 271, 839–846 (2004).
Schulz, S. Semiochemistry of spiders. In Advances in Insect Chemical Ecology (eds Carde, R. T. & Millar, J. G.) 110–150 (Cambridge University Press, London, 2004). https://doi.org/10.1017/cbo9780511542664.005.
Klewer, N., Růžička, Z. & Schulz, S. (Z)-Pentacos-12-ene, an oviposition-deterring pheromone of Cheilomenes sexmaculata. J. Chem. Ecol. 33, 2167–2170 (2007).
Roelofs, W. L. Electroantennogram assays: Rapid and convenient screening procedures for pheromones. In Techniques in Pheromone Research (eds Hummel, H. E. & Miller, T. A.) 131–159 (Springer, New York, 1984). https://doi.org/10.1007/978-1-4612-5220-7_5.
Gries, R. et al. Limoniic acid—Major component of the sex pheromones of the click beetles Limonius canus and L. californicus. J. Chem. Ecol. 47, 123–133 (2021).
Varner, E. et al. Brown rats and house mice eavesdrop on each other’s volatile sex pheromone components. Sci. Rep. 10, 17701 (2020).
Shine, R. & Mason, R. T. An airborne sex pheromone in snakes. Biol. Lett. 8, 183–185 (2012).
Poth, D., Wollenberg, K. C., Vences, M. & Schulz, S. Volatile amphibian pheromones: Macrolides from mantellid frogs from Madagascar. Angew. Chem. Int. Ed. 51, 2187–2190 (2012).
Ganske, A.-S. & Uhl, G. The sensory equipment of a spider—A morphological survey of different sensillum types in both sexes of Argiope bruennichi (Araneae, Araneidae). Arthropod Struct. Dev. 47, 144–161 (2018).
Uhl, G. Spider olfaction: Attracting, detecting, luring and avoiding. In Spider Ecophysiology (ed. Nentwig, W.) 141–157 (Springer, 2013). https://doi.org/10.1007/978-3-642-33989-9.
Acknowledgements
We thank the Lancaster family of the Black Widow Winery for allowing us to collect spiders, and Sharon Oliver for some word processing. We thank Scripps Research for providing free access to XCMS online which was used in data analysis and to create subpanel c in Figure 3. A.F. was supported by Graduate Fellowships from SFU, the H.R. McCarthy Bursary, and an Alexander Graham Bell Scholarship from the Natural Sciences and Engineering Research Council of Canada (NSERC). The project was funded by an NSERC—Industrial Research Chair to G.G., with Scotts Canada Ltd. and BASF Canada Inc. as the industrial sponsors. The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
Funding
Open Access funding enabled and organized by Projekt DEAL.
Author information
Authors and Affiliations
Contributions
Conceptualization, A.F., H.C. and G.G.; Methodology, A.F., R.G., and G.G.; Investigation, A.F., A.C.R-T, J.V., Y.L., N.B., R.G., S.A., H.C.; Data Curation, A.F.; Writing—Original Draft, A.F and G.G.; Writing—Review & Editing, G.G. and A.F; Funding Acquisition, G.G.; Resources, G.G., A.F.; Supervision, A.F.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Fischer, A., Roman-Torres, A.C., Vurdela, J. et al. Non-targeted metabolomics aids in sex pheromone identification: a proof-of-concept study with the triangulate cobweb spider, Steatoda triangulosa. Sci Rep 13, 18426 (2023). https://doi.org/10.1038/s41598-023-44948-0
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-023-44948-0
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
