
Abstract
Although larval diet quality may affect adult mosquito fitness, its impact on parasite development is scarce. Plant pollen from Zea mays, Typha latifolia, and Prosopis juliflora was ultraviolet-sterilized and examined for effects on larval development, pupation rate, adult mosquito longevity, survival and infectivity. The control larvae were fed Tetramin fish food as a comparator food. Four treatment and two control groups were used for each pollen diet, and each experimental tray had 25 larvae. Female An. arabiensis were starved overnight and exposed to infectious blood using a membrane-feeding system. The Kaplan–Meier curves and log-rank test were used for analysis. The Z. mays pollen diet increased malaria mosquito survival and pupation rate (91.3%) and adult emergence (85%). Zea mays and Tetramin fish food had comparable adulthood development times. Adults who emerged from larvae fed Z. mays pollen had the longest average wing length (3.72 mm) and were more permissive to P. vivax (45%) and P. falciparum (27.5%). They also survived longer after feeding on infectious blood and had the highest number of P. vivax oocysts. Zea mays pollen improved larval development, adult mosquito longevity, survival and infectivity to Plasmodium. Our findings suggest that malaria transmission in Z. mays growing villages should be monitored.
Similar content being viewed by others

Introduction
Plasmodium falciparum and P. vivax are the two dominant human malaria parasites in Ethiopia, and the principal vector is Anopheles arabiensis. Some mosquito species are efficient and highly permissive to the parasite to complete the cycle and predominantly bite humans to increasing the parasite transmissibility1. There is also a link between the intensity of malaria transmission and the biology of the mosquito vectors. For instance, the quality of larval food may have an impact on adult life histories, including adult survival, the number of emerging adults, and the development of parasites2. Female Anopheles mosquitoes usually select breeding habitats suitable for the survival and development of immature stages. The pollen (fine to a coarse powdery substance comprising pollen grain) from cultivated and wild grasses supplement the natural food for Anopheles larvae in the breeding habitats and might support the development and increase malaria transmission3,4. For example, maize cultivation is mostly claimed to increase malaria transmission5. The odour from the maize pollen also has lured gravid malaria mosquitoes to the oviposition site6. Moreover, nutrition during the larval stage determines the size and metabolic reserves of adult mosquitoes4,7.
There are, however, controversial results with regard to the plant-vector interaction. For example, a study by Muller and his colleagues8 reported higher risk of malaria infection during the flowering season of an invasive plant species Prosopis juliflora. When the flowering branches of this plant were removed, the age structure of the malaria mosquitoes changed, and the number of older malaria mosquitoes was substantially reduced. It might be linked to the sugar that flowering plants provide adult mosquitoes as a source of energy, which can influence their age. The pollen of some plants is more nutritious than others. However, several plants may inhibit disease transmission by affecting vectors in their breeding habitats because of the presence of toxic constituents. For example, Cyperus papyrus and Phragmites australis contains chemicals that can kill the larval stages in breeding habitats9. Some plants have very attractive flowers for mosquitoes and some others have toxic sugar baits10.
Existing knowledge on the interaction of malaria vectors and plants with respect to parasite transmission is scarce. Zea mays (a maize specimen) is the most common crop in the lowlands of Gamo zone where malaria is also endemic11. The crop is cultivated all year round. The shores of the Lake Abaya and Chamo are predominantly covered by T. latifolia. The shores of the two lakes are also ideal breeding sites for the primary and secondary malaria vectors11,12. The shrub P. juliflora is also a very common plant species in the region. While there is little evidence about the interaction between malaria vectors and plants, effective control requires a contextual understanding of the bio-ecology of malaria vector-plant interaction. However, in Ethiopia, there is limited understanding of the impact of pollen on early stages and its long-term effect on adult fitness and susceptibility to the parasite. In light of these factors, the purpose of this study was to demonstrate whether pollen from T. latifolia, Z. mays and P. juliflora enhances An. arabiensis larval development and survivorship, pupation rate, adult size, longevity of malaria vector, survival and infection to Plasmodium parasites.
Materials and methods
Study area description
Anopheles larval sampling was made from the potential breeding habitats in the Arba Minch area, located in the southern Ethiopian Rift Valley system. The climate is hot and humid which is favourable for mosquito breeding and malaria transmission. The average maximum temperature is 30 °C, while the average minimum is 17 °C. The rainfall distribution is bimodal and mostly occurring in March to May and September to November. The annual rainfall average is around 900 mm. The major cash crops cultivated in the area include bananas, maize and mango using both rain and irrigation schemes. In the rainy seasons, the water pools in ditches along the roads, farmlands, tire track pooled water, and marshy areas are the potential breeding sites for Anopheles mosquitoes.
Malaria is a common health problem in the area11,13 and is more prevalent after the end of the rainy season due to the potential breeding sites11. Plasmodium falciparum and P. vivax are the two common species14,15.
Study design
This experiment aimed to assess the effect of various plant pollen diets on key vectorial aspects of An. arabiensis. Newly emerged larvae were provided with equal quantity of pollen from T. latifolia, Z. mays, P. juliflora and standard larval food Tetramin fish food (Cichlid Sticks; Tetra, Maidenhead Aquatics, Leicester, UK). The emerged adults were supplied with10% sugar solution until ready for the infection experiments16. Then, 2–3 days old female Anopheles were starved overnight and provided with P. vivax and P. falciparum parasite infected blood though membrane feeding apparatus (Fig. 1).

The study flow chart.
Study plants
These plants (Fig. 2) were selected based on the scientific evidences documented in relation to the malaria mosquitoes either against or in favour of larval and adult stages5,7. Typha latifolia (common cattail) is a perennial herbaceous plant in the genus Typha and among the most common of all aquatic plants. It is an "obligate wetland" species found close to water bodies. This plant is among the wind-pollinated plants that produces large quantities of pollen. Prosopis juliflora is a shrub or small tree in the family Fabaceae. It is an evergreen and fast-growing invasive species in arid and semi-arid areas. The flowers of P. juliflora are self-incompatible, entomophilous (insect pollinating), depending on insects for seed dispersal. It produces many seeds and is tolerant of a wide range of climatic regimes and soil types which have contributed to making P. juliflora one of the worst invasive alien plants. The leafy stalk of Z. mays (maize) produces pollen inflorescences and separate ovuliferous inflorescences called ears that yield seeds, which are fruits and its pollen, are produced entirely in the staminate inflorescence that is wind pollinated. The male flower matures earlier than the female flower. The plant specimens were collected from their typical habitat by field press and brought to the Arba Minch University Botany laboratory for further confirmation by expertise.

Study plants (a) T. latifolia, (b) Z. mays, (c) P. juliflora.
Pollen collection
Pollen of T. latifolia was collected in the shore of the Lake Abaya, P. juliflora from the shrub dominated area of Lante (local place in Arba Minch area) and the pollen of Z. mays L. (BH660 cultivar) from the farmland of Lemat during the maize flowering season. The pollens were collected by covering the mature flower with a paper bag and shaking. The collected pollen was transferred to vials and sterilized within 24 h of collection using UV light (10850 Harry Hines Blvd Suite 155, Dallas, TX 75220) overnight for 12 h and store on silica gel until used for feeding experiments.
Pollen nutrient analysis and measuring grain size
The C and N concentration of pollen diets were determined by Walkley–Black and Kjeldahl methods in Arba Minch University analytical chemistry laboratory. Approximately 5 mg of sample of pollen from each plant was taken and processed based on the standard protocol developed by Roulston et al17. Thereafter, the C: N ratio of the pollen was determined by %C/%N × 6.2517. The pollen grain sizes were measured according to Roulston et al17. Pollen grains were mounted in paraffin oil and measured at 400 × with an ocular micrometer. Polar axis measurements were taken for all pollen grains. Volumes were calculated using volume equations for spheres (4/3πp3). For the purpose of calculating grain volume, ten randomly selected grains per slide were measured, and these values were averaged.
Mosquito larvae collection and rearing
Anopheles larvae were collected from the potential breeding habitats in the Arba Minch area and reared in Arba Minch University Medical Entomology and Vector Control laboratory. Swamps along the Lake Abaya shoreline and water holding areas at construction sites were the most common breeding habitats where the larvae were collected. Additionally, there were a few small bodies of water gathered alongside ditches and rivers that were Anopheles larvae-positive. The rearing room was maintained at temperature between 25 and 27 °C and relative humidity between 72 and 80%. The larvae rearing trays were inspected twice a day for the presence of pupae, and if present they were placed in adult cages. The larvae collected from natural habitats were not employed in this experiment due to the range of habitats and food sources.
The newly emerging adults were provided with 10% sterilized sugar solution. Then, they were allowed feed on live rabbit at 3- to 4-days interval and serving as ‘mother’ cages. The egg dish was placed in the cage one day after the adult fed on blood, and eggs were transferred to the larvae tray for hatching. Using these larvae, an experiment was conducted.
The female adult mosquitoes in the mother cage were morphologically identified to the An. gambiae complex, and a sub-sample (150) was tested for An. gambiae complex by polymerase chain reaction (PCR) in the Medical Entomology laboratory at Arba Minch University following the protocol of Scott et al18. To mention in brief, legs or wings were used for DNA extraction using the NucleoSpin Tissue kit (Macherey Nagel, Germany) according to the manufacturers’ instructions. The species-specific nucleotide sequences of the rDNA intergenic spacers (IGS) were targeted for An. gambiae complex as described by the protocol mention above. The PCR products were visualized on a 1% agarose gel using a 100 bp ladder (Promega, The Netherlands) and species identification was done based on the band height. The entire tested An. gambiae complex was identified as An. arabiensis (hereafter the target mosquito species will be An. arabiensis).
Larval development and survival analyses
After 12 h of hatching, 200 unfed first instar larvae were counted manually and randomly apportioned to larval trays of 10 × 25 cm and 8 cm high filled with one litter of distilled water. Effective dose of pollen diets and TetraMin fish food was determined by optimizing 0.15, 0.3 and 0.6 mg on relative adulthood survival. Then, 0.3 mg was established as the effective dose of each pollen diet and Tetramin fish food.
Six replicates each with 25 larvae for each of the plant pollen diet and two as control group were used. The experiment was repeated three times for each pollen diet and control group of TetraMin fish, using different mosquito batches from the field. A total of 450 larvae were treated for each diet and 150 for each experimental term. For the first and the second instar larvae, the diet was offered only once a day, for the third and fourth, it was twice a day based on procedure of Kivuyo et al19. In each replicate, the larval development and survival was recorded twice a day (01:00 and18:00) and dead individuals were counted and removed. Continuous inspection of the number of larvae surviving, pupating, and emerging adults were recorded.
where s1 the no. of larvae at the begning of day 1, s2 the no. of larvae at the begning of day 2.
Measuring mosquito size/wing length
The same batches and same ages of 30 female adult An. arabiensis were collected from each pollen diet and TetraMin fish food. They were killed by freezing and, then one wing of each mosquito glued onto a slide. Its length from the distal end of the alula to the tip, excluding the fringe scales, was measured with stage graticule micrometer (ruler etched in side) to examine the effect of larval diet on wing length of adult An. arabiensis.
Patient screening for gametocyte stage
Microscopically positive patients with P. vivax and P. falciparum gametocytes were enrolled. The gametocyte detection was done by standard finger prick and Giemsa stained blood smears from thick blood smear under oil immersion microscopy by experienced laboratory technicians. The gametocyte positive patients were asked to provide approximately 3 ml of blood for membrane feeding prior to antimalarial treatment. Blood gametocyte density was estimated microscopically by converting the total number of gametocytes in 1000 leukocytes to 8000 leukocytes/μl. Screening of gametocyte carriers was synchronized into mosquito rearing processes and thus always performed on the same day with the experimental infection. Patients with mixed infection were excluded.
Membrane blood feeding
The same batch of 2 to 3-day-old female An. arabiensis were starved overnight and kept for experimental infection through membrane feeding. A total of 1280 female An. arabiensis were allowed to feed on P. vivax positive blood in four membrane feeding experiments, and another 1280 were exposed to P. falciparum positive blood in four membrane feeding experiments. In each feeding experiment, 320 An. arabiensis were exposed in 8 paper cups each with 40 mosquitoes.
To keep the blood warm and the parasites alive, a circulation water bath was used, and its temperature was set to 37 °C. After carefully pipetting a Plasmodium-positive blood meal into the feeder's neck, it was then allowed to feed simultaneously for 30 min in the dark. Unfed mosquitoes were discarded after carefully inspecting them for abdominal conditions. The fed An. arabiensis were kept in the paper cups and provide with 10% sugar solution under normal insectary condition (27 °C ± 2 and 70 ± 10% relative humidity). Anopheles arabiensis were kept under sugar for eight days for P. vivax and twelve days for P. falciparum oocyst detection. The cups were put inside the small cage as an extra layer of protection in case the mosquitoes that feed on the infectious blood left. The membrane was removed carefully and soaked in 10% bleach to decontaminate and rinse with water.
Dissection to oocyst stage and sporozoite detection
Randomly 40 mosquitoes were taken from each pollen type replicate cages on day 7 post-infection for P. vivax and on day 12 post infection for P. falciparum. A total of 120 female mosquitoes were dissected for P. vivax and P. falciparum. For each of the dissection sessions, the infection rate (infected mosquitoes/total mosquitoes dissected in each feeding (%) was recorded, and intensity of infection (oocyst load/total infected mosquitoes dissected in each feeding) were determined. Non-infected mosquitoes were omitted from the calculation.
The infective stage of the Plasmodium parasite was detected by circum-sporozoite protein (CSP)-ELISA using the kits specific for P. vivax-210, P. vivax-247 and P. falciparum following standard instruction19. A total of 104 and 35 An. arabiensis were tested for CSPs P. vivax and P. falciparum, respectively.
Data analysis
In order to conduct a survival analysis, data from the experiment on the impact of diets on larval survival were transformed to count-time data. Kaplan–Meier curves were used to compare diet-specific survival pattern for each group. Comparison of survival rate in each larval diet to that of TetraMin Fish food was performed using log rank test. The Analysis of Variance (ANOVA) test was conducted to evaluate how larval diets impacted the mean pupation rate, adult emergence rate, and wing lengths. Statistical testing was performed using the mean of each pollen diet, taking into account the origin of the mosquitoes in each batch. Wald’s test was used to determine significant differences between diets on the vectorial parameters of An. arabiensis based on χ2 and P-values.
The proportion of female adult mosquitoes that survived after infected blood meal was determined by counting the number of mosquitoes on their oocyst and sporozoite stage across all larval diets. The median number of oocyst per mosquito mid-gut was compared using a Kruskal–Wallis test, followed by a Dunn's post-hoc test. The prevalence of sporozoite infection, or percentage of mosquitoes reaching to infective stage, was compared with a Fisher's exact test.
All statistical analysis performed using GraphPad Prism 5 (Version 5.01). Significance was considered at 5% level of confidence. Single factor experimental design was employed to assess the impact of pollen of some selected plants on the larval survival, pupation, adult emergence, and vector infectivity to malaria parasites. Survival rate of each treatment case was calculated as survival rate of each day.
Ethical consideration
This study was reviewed and approved by the Institutional Review Board of Arba Minch University (No: CMHS/12036564/111). After briefing the objective of the study, consent was obtained from the study participants. Participants in the study who tested positive for Plasmodium received free treatment in accordance with the national malaria treatment guidelines. The CSP-ELISA test was performed in accordance with standard protocol19. All methods were performed in accordance with the standard protocols and procedures19. Other plant pollens, aside from maize pollen, were collected with the local administration approval in open spaces like the lakeshore and by the side of the road. Through permission of farm owners, the maize pollen was collected on farmland. Despite the fact that only the pollen grains were gathered for the experiment, plant specimens were also gathered in order to be identified by senior botanists before collection. The voucher specimens of the plants (P. juliflora and T. latifolia) have been deposited in a publicly available botanical herbarium at Arba Minch University.
Results
Pollen nutrient content and size
The pollen grain volume was significantly different between plants with higher pollen volume of Z. mays (mean diameter 71.2 ± 1.66 µm) compared to P. juliflora (48.1 ± 0.56 µm) and T. latifolia (25.5 ± 0.32 µm) (F = 1033, DF = 3, P < 0.0001).
The C: N ratio of pollen differed between test plants with the pollen C: N ratio of T. latifolia > Z. mays > P. juliflora. Typha latifolia had a higher proportion of carbon (52%), compared with the two other species Z. mays (44%), and P. juliflora (32%). The higher proportion of nitrogen was belonging to Z. mays (31%) followed by T. latifolia (28%) and P. juliflora (18%).
Larval diet impacts on larval survival
Survivorship of the larval instars was found to be diet-dependent. The overall larval survival differed significantly among the pollen diets tested (F = 23, DF = 3, P < 0 0.0001). The survival rate of larvae fed on the Z. mays pollen was 100% until day 8 and slowly declined with the minimum survival rate of 0.4 at day 15. In the follow-up period, the likelihood of survival for larvae fed on diets of Z. mays pollen was 0.89, followed by T. latifolia (0.71). The survivorship of larvae due to P. juliflora pollen diet was found to be the poorest (highest larval death) with 0.51 probability of survival during the follow-up period. The group of An. arabiensis larvae supplied with T. latifolia pollen had accelerated development of larval instars, while those supplied with the P. juliflora pollen diet experienced extended larval period (Fig. 3).

Anopheles arabiensis larvae survivorship and duration of larval development in larval diet of pollens of T. latifolia, Z. mays and P. juliflora and TetraMin fish food.
Effect of larval diets on pupation rate and duration
The effect of pollen diets on the larval average time to complete the larval stage and change to the pupae showed significant differences between the diets (χ2 = 12.8, DF = 3, P < 0.0001). Anopheles arabiensis pupation was faster on T. latifolia, it took 2-to-3-days since the starting day of pupation followed by Tetramin fish and Z. mays. Larval instars took longer time to develop into pupae when larvae supplied with P. juliflora pollen diet 5-to-11 days (Table 1).
Adult emergence rate
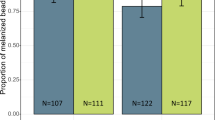
A significant difference was observed between larval diets from different plants on adult emergence rate (x2 = 17.3, DF = 3, P < 0.0001). The highest adult emergence rate was documented in Z. mays diet (P < 0.05) followed by TetraMin fish food and T. latifolia. The number of adults that emerged in P. juliflora was lower compared to the three other diets (Fig. 4). Moreover, high mortality of adult mosquitoes was also documented on this diet. But there was no significant deviation in time needed to reach adulthood in either of the diet (P > 0.05), mostly, emerged with in 48 h after pupation. More females emerged from larvae fed on Z. mays pollen, compared to other diet sources but not significantly different (P > 0.05).

The number adults emerged from initial number (n = 150) of larvae in different larval diets.
Larval diet effects on wing length
Wing length was significantly affected by the type of larval diet (χ2 = 69, DF = 39, p < 0.0001). The female An. arabiensis emerged from larvae fed on Z. mays possessing bigger wings (mean WL = 3.72 mm) than females that fed on Tetramin fish (mean WL = 3.25 mm) and T. latifolia (mean WL = 2.82 mm). The overall mean wing length was 2.9 mm, with a range of 2.06 mm to 3.72 mm. The An. arabiensis that were reared on P. juliflora pollen had the smallest wing size (mean WL = 2.06 mm) (Fig. 5).

The effect of larval diets on wing length (WL) of adult female A. Arabiensis.
Larval diet impacts on adult survival and oocyst density
The proportion of females surviving until the day of dissection for oocysts after feeding on P. vivax and P. falciparum positive blood was higher among adults emerged from larvae fed on Z. mays pollen diet than other diets (Fig. 6).

The percentage of adults survived after feeding on Plasmodium parasite positive blood until day 7 for P. vivax (a) and day 12 for P. falciparum (b) midgut dissection for oocyst detection (boxes show the 95% confedance interval).
The proportion of Plasmodium oocysts found in An. arabiensis following an infected blood meal varied significantly depending on their larval diets (P < 0.05). Anopheles arabiensis adults emerged from larvae fed on Z. mays pollen showed the highest oocyst infection rate (45% for P. vivax and 27.5% for P. falciparum) followed by Tetramin fish food (30% for P. vivax and 20% for P. falciparum) (Table 2). There was no significant difference in Z. mays and Tetramin fish (P > 0.05). Those An. arabiensis fed on T. latifolia had infection rates of 22% for P. vivax and 8.3% for P. falciparum.
The median number of P. vivax oocysts was higher in adult An. arabiensis emerged from larvae reared on Z. mays pollen diet (Fig. 7). The intensity of infection of An. arabiensis reared on Z. mays varied significantly from other pollen diets (P < 0.0001). The highest median number of oocysts was observed in An. arabiensis reared on Z. mays (median ꞊ 43) for P. vivax and (median = 0) for P. falciparum followed by Tetramin fish (median = 6) for P. vivax and (median = 0) for P. falciparum. The lowest mean number of oocysts was reported in those An. arabiensis reared on P. juliflora pollen larval diets (mean = 0.26, 1.04 ± SD, median = 0) for P. vivax and no infection for P. falciparum.

Median number of oocysts of P. vivax (left) and P. falciparum (right) in the mid-gut of female Anopheles arabiensis emerged from different larval diets.
Larval diet impacts on CSP infection rate
The overall CSP rate for P. vivax was 8.6% (95% CI 1.14–21.02; 9 of 104) (Table 2). P. vivax CSP rate of An. arabiensis reared on Z. mays pollen was 13.2% (95% CI 4.4–28.1; 5 of 38) followed by 9.1% (95% CI 1.9–24.3; 3 of 33) for Tetramin fish. No An. arabiensis from larvae fed on P. juliflora was positive for P. vivax. No An. arabiensis were found to be positive for P. falciparum CSPs (Table 3).
Discussion
The effects of plant pollen larval diets on adult emergence, female size, pupation length, larval and adult survival, and parasite infection with P. falciparum and P. vivax were examined in this study. Pollen from Z. mays enhances larval survival, adult emergence, and Plasmodium parasite infectivity. Adult An. arabiensis emerged from larvae fed on Z. mays pollen were more susceptible to P. vivax and P. falciparum parasites and had the highest mean number of P. vivax oocysts and the longest survival times. Adults that emerged from larvae fed P. juliflora pollen had the lowest infection rate.
The development, survival, and adult emergence rates of Anopheles larvae are all influenced by the quantity and quality of their diet7. In farmland-associated habitats, maize pollen grains had a positive correlation with larvae abundance20. The grasses including T. latifolia also determine the abundance of larval stages of Anopheles mosquitoes and adult productivity21. Prosopis juliflora, an invasive plant, has been found to increase the number of Anopheles mosquitos8. There is evidence that maize pollen has beneficial effects on mosquito longevity, vector density, and larval development5. Additionally, the odour of maize pollen is utilised to attract gravid female malaria mosquitoes to lay their eggs6. This study investigated the effects of larval diets on survival of adults until oocyst development, the density of oocyst infections, and Plasmodium parasite susceptibility using the artificial membrane feeding experiment.
In this study, a substantial difference in larval survival rate was observed between larvae fed on different pollen diets. The pollen of Z. mays increased the survival of malaria mosquito larvae and adults. It has been demonstrated that larvae fed Z. mays pollen survived better21. In contrast, larvae fed T. latifolia pollen took less time to pupate than Z. mays, but those fed P. juliflora pollen diet took longer to mature into pupae. The ratio of carbon to nitrogen in the diet mat influence the growth and survival of the An. arabiensis22. The protein content of Z. mays pollen may be the most important element impacting An. arabiensis larval development and survival5. The larvae need enough food for ecdysis in order to avoid having a high mortality rate due to a prolonged larval period caused by a poor diet23.
When compared to the other pollen diets, larvae fed Z. mays pollen produced the largest female adults, which is proportional to the length of its wing. Numerous studies have demonstrated that one could produce small, intermediate, and large-sized mosquitoes by varying the amount and type of food24. This could be due to the nutritional values. An earlier study has shown that adult mosquitoes may grow to reasonably large sizes when their dietary needs are met25. Also, larger An. arabiensis live longer and lay more eggs than smaller An. arabiensis26.
Adult An. arabiensis emerged from larvae fed Z. mays pollen were more permissive to infection with P. vivax and P. falciparum parasites, and had the highest number of P. vivax oocysts. Adult body size, as well as the larval quality diets, may explain this7,25. The nutrient content of Z. mays may be superior to other diets and result in large-size female adults. Larger Anopheles mosquitoes ingest more blood meals, which correlate positively with the number of oocyst development7,27. Larger infected mosquitos have a better chance of surviving and may harbour more oocysts, resulting in the largest incidence of infective mosquitoes. In Ethiopia, communities that grow maize have been linked to higher rates of malaria infection3. Therefore, additional malaria vector control measures like larval source management could be strengthen in malaria endemic areas, particularly during the maize flowering seasons.
Additionally, it is possible that increasing the cultivation of crops like Z. mays could enhance the larval diet and perhaps generate favourable conditions for increased vector populations and transmission of malaria. This is because of the influence of larval nutrition on larval survival, adult fitness, oocyst density, and infection. Adult mosquitoes are the target for contemporary vector control. However, during a favourable transmission season, such as the growing season to maize, larval and pupal control efforts may be combined with adult control methods. Based on this study, some plants located close to breeding habitats can contribute to an increase in the number of vectors and the transmission of parasites since the food from these plants impacts the vector's capacity. In regions where maize is grown, intensified larval source management may be encouraged to reduce the number of mosquito larvae.
Although An. arabiensis from the wild were employed in the study, it was conducted in a laboratory setting; hence, field trials would be the next step. The failure to link the mechanisms of the various food source components to the vectorial components of malaria vectors was another limitation of the study. More study on this area is encouraged for the formulation of a tailored intervention.
Conclusions
In conclusion, our results provide new insights into the effect of larval diet on mosquito larval survival, pupation, adult size and infectivity to Plasmodium parasites in Ethiopia. Of the diets, Z. mays pollen diet was superior to the other diets including the standard Tetramin fish food. Larvae fed on this diet produced adults with larger wing size and longer longevity, and greatly increasing the infectivity to malaria parasites. In contrast, the larval development time was significantly longer and mortality rate was higher when the larvae fed on P. juliflora pollen diet. These conditions also resulted in a reduction in adult longevity and adult fitness. Therefore, further investigations on the nutritional components of plant pollen as a source of mosquito larva diet, as well as the link between larval nutrition and parasite growth in vectors, should be conducted.
Data availability
The datasets used and/or analysed during the current study available from the corresponding author on reasonable request.
References
Cohuet, A., Harris, C., Robert, V. & Fontenille, D. Evolutionary forces on Anopheles: What makes a malaria vector?. Trends Parasitol. 26, 130–136 (2010).
Moller-Jacobs, L. L., Murdock, C. C. & Thomas, M. B. Capacity of mosquitoes to transmit malaria depends on larval environment. Parasit. Vectors 7, 1–12 (2014).
Kebede, A., McCann, J. C., Kiszewski, A. E. & Ye-Ebiyo, Y. New evidence of the effects of agro-ecologic change on malaria transmission. Am. J. Trop. Med. Hyg. 73, 676–680 (2005).
Carvajal-Lago, L., Ruiz-López, M. J., Figuerola, J. & Martínez-de la Puente, J. Implications of diet on mosquito life history traits and pathogen transmission. Environ. Res. 195, 110893 (2021).
Ye-Ebiyo, Y., Pollack, R. J., Kiszewski, A. & Spielman, A. A component of maize pollen that stimulates larval mosquitoes (Diptera: Culicidae) to feed and increases toxicity of microbial larvicides. J. Med. Entomol. 40, 860–864 (2003).
Wondwosen, B. et al. A(maize)ing attraction: Gravid Anopheles arabiensis are attracted and oviposit in response to maize pollen odours. Malar. J. 16, 1–9 (2017).
Takken, W. et al. Larval nutrition differentially affects adult fitness and plasmodium development in the malaria vectors Anopheles gambiae and Anopheles stephensi. Parasit. Vectors 6, 860–864 (2013).
Muller, G. C. et al. The invasive shrub Prosopis juliflora enhances the malaria parasite transmission capacity of Anopheles mosquitoes: A habitat manipulation experiment. Malar. J. 16, 1–9 (2017).
Fillinger, U., Sonye, G., Killeen, G. F., Knols, B. G. J. & Becker, N. The practical importance of permanent and semipermanent habitats for controlling aquatic stages of Anopheles gambiae sensu lato mosquitoes: Operational observations from a rural town in western Kenya. Trop. Med. Int. Heal. 9, 1274–1289 (2004).
Müller, G. C. et al. Field experiments of Anopheles gambiae attraction to local fruits/seedpods and flowering plants in Mali to optimize strategies for malaria vector control in Africa using attractive toxic sugar bait methods. Malar. J. 9, 1–11 (2010).
Loha, E. & Lindtjørn, B. Predictors of Plasmodium falciparum malaria incidence in Chano Mille, SouthEthiopia: A longitudinal study. Am. J. Trop. Med. Hyg. 87, 450–459 (2012).
Massebo, F., Balkew, M., Gebre-Michael, T. & Lindtjorn, B. Entomologic inoculation rates of Anopheles arabiensis in southwestern ethiopia. Am. J. Trop. Med. Hyg. 89, 466–473 (2013).
Getawen, S. K. Exploring the impact of house screening intervention on entomological indices and incidence of malaria in Arba Minch town, southwest Ethiopia: A randomized control trial. Acta Trop. 181, 84–94 (2018).
Esayas, E., Woyessa, A. & Massebo, F. Malaria infection clustered into small residential areas in lowlands of southern Ethiopia. Parasite Epidemiol. Control 10, e00149 (2020).
Abraham, M., Massebo, F. & Lindtjørn, B. High entomological inoculation rate of malaria vectors in area of high coverage of interventions in southwest Ethiopia: Implication for residual malaria transmission. Parasite Epidemiol. Control 2, 61–69 (2017).
World Health Organization. Malaria Entomology and Vector Control 192 (World Health Organization, 2013).
Monographs, E. What governs protein content of pollen : Pollinator preferences, pollen-pistil interactions, or phylogeny ? In (eds. Roulston, T. H, Cane, J. H., & Buchmann, S. L.). 70, 617–643. Wiley Stable. http://www.jstor.org/stable/2657188 (2016).
Scott, J. A., Brogdon, W. G. & Collins, F. H. Identification of single specimens of the Anopheles gambiae complex by the polymerase chain reaction. Am. J. Trop. Med. Hyg. 49, 520–529 (1993).
Kivuyo, H. S. et al. Performance of five food regimes on Anopheles gambiaesenso stricto larval rearing to adult emergence in insectary. PLoS ONE 9, e110671 (2014).
Kweka, E. J. et al. Anopheline larval habitats seasonality and species distribution: A prerequisite for effective targeted larval habitats control programmes. PLoS ONE 7, e52084 (2012).
Asmare, Y., Hill, S. R., Hopkins, R. J., Tekie, H. & Ignell, R. The role of grass volatiles on oviposition site selection by Anopheles arabiensis and Anopheles coluzzii. Malar. J. 16, 1–9 (2017).
Raubenheimer, D., Simpson, S. J. & Mayntz, D. Nutrition, ecology and nutritional ecology: Toward an integrated framework. Funct. Ecol. 23, 4–16 (2009).
Telang, A., Frame, L. & Brown, M. R. Larval feeding duration affects ecdysteroid levels and nutritional reserves regulating pupal commitment in the yellow fever mosquito Aedes aegypti (Diptera: Culicidae). J. Exp. Biol. 210, 854–864 (2007).
Ng’habi, K. R. et al. Sexual selection in mosquito swarms: May the best man lose?. Anim. Behav. 76, 105–112 (2008).
Russell, T. L. et al. Linking individual phenotype to density dependent population growth: The influence of body size on the population dynamics of malaria vectors. Proc. R. Soc. B Biol. Sci. 278, 3142–3151 (2011).
Ameneshewa, B. & Service, M. W. The relationship between female body size and survival rate of the malaria vector Anopheles arabiensis in Ethiopia. Med. Vet. Entomol. 10, 170–172 (1996).
Koella, J. C. Relationship between body size of adult Anopheles gambiae s.l. and infection with the malaria parasite Plasmodium falciparum. Parasitology 104, 233–237 (1992).
Acknowledgements
Study participants are sincerely thanked for their kind assistance by donating blood for feeding experiments. We thank botanist Mulugeta Kebebew for his assistance with the study's plant identification.
Funding
This work was financially supported by the Norwegian Programme for Capacity Development in Higher Education and Research for Development-Arba Minch University project (ETH-13/0025).
Author information
Authors and Affiliations
Contributions
Conceptualisation and study design: S.A., T.W., F.M.; Formal analysis and interpretation of data: S.A., F.M.; Laboratory work: S.A., G.T., N.E.; Supervision: F.M., T.W., BL.; Project administration: F.M., B.L.; Validation: F.M., B.L.; Drafting and revising the manuscript: F.M., B.L. All authors reviewed the manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Ayele, S., Wegayehu, T., Eligo, N. et al. Maize pollen diet enhances malaria mosquito longevity and infectivity to Plasmodium parasites in Ethiopia. Sci Rep 13, 14490 (2023). https://doi.org/10.1038/s41598-023-41826-7
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-023-41826-7
This article is cited by
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
