
Abstract
HBV/HCV co-infection is common in HIV-1-infected prisoners. To investigate the characteristics of HIV co-infections, and to evaluate the molecular heterogeneity of HIV, HBV and HCV in prisoners, we carried-out a multicenter cross-sectional study, including 65 HIV-1-infected inmates enrolled in 5 Italian detention centers during the period 2017–2019. HIV-1 subtyping showed that 77.1% of inmates were infected with B subtype and 22.9% with non-B subtypes. Italian nationals were all infected with subtype B (93.1%), except two individuals, one infected with the recombinant form CRF72_BF1, and the other with the HIV-1 sub-subtype A6, both previously not identified in inmates of Italian nationality. Non-Italian nationals were infected with subtype B (52.6%), CRFs (36.8%) and sub-subtypes A1 and A3 (5.2%). HIV variants carrying resistance mutations to NRTI, NNRTI, PI and InSTI were found in 7 inmates, 4 of which were never exposed to the relevant classes of drugs associated with these mutations. HBV and/or HCV co-infections markers were found in 49/65 (75.4%) inmates, while 27/65 (41.5%) showed markers of both HBV and HCV coinfection. Further, Italian nationals showed a significant higher presence of HCV markers as compared to non-Italian nationals (p = 0.0001). Finally, HCV phylogenetic analysis performed in 18 inmates revealed the presence of HCV subtypes 1a, 3a, 4d (66.6%, 16.7% and 16.7%, respectively). Our data suggest the need to monitor HIV, HBV and HCV infections in prisons in order to prevent spreading of these viruses both in jails and in the general population, and to implement effective public health programs that limit the circulation of different genetic forms as well as of viral variants with mutations conferring resistance to treatment.
Similar content being viewed by others

Introduction
The inmate population represents a model of closed community subject to restrictive conditions that has a multinational composition and a high incidence of infections. It is estimated that around 3.8% of the global prison population is living with HIV1. Prevalence of HIV infection in jail differs greatly in different regions with proportion greater than 10% in low- and middle-income countries2. In Italy, the prevalence of HIV infection among inmates has been estimated to be about 7.5%3.
Resident non–Italian nationals are 8.4% of the Italian general population (Italian National Institute of Statistics, 2020). However, according to the December 2022 official data of the Italian Ministry of Justice, non-Italian national inmates in Italian detention centers represented 31.5% of the total inmates. Therefore, non-Italian national inmates are overrepresented in the Italian prisons. Consequently, prevalence of HIV subtypes in prison can be influenced by the presence of non-Italian nationals, regardless of the geographic areas of origin. Our previous data on the prevalence of HIV-1 genetic forms circulating in the immigrant population in Italy demonstrated that almost 80% of them are represented by non-B subtypes and recombinant forms (CRFs)4. In contrast, the HIV-1 B subtype was the main genetic form found in the Italian general population, although other subtypes and CRFs have been identified and their prevalence is increasing overtime5,6,7.
Because of the similar modes of transmission of HIV, HBV and HCV, patients with behaviors at risk for HIV infection are also at risk for HBV and HCV infection. HBV/HIV-positive individuals are also at increased risk for developing chronic HBV infection8. In addition, HCV may also have an impact on the clinical management of HIV infection9,10. In fact, previous studies suggest that HCV co-infection may have an impact on HIV disease progression also in combined antiretroviral therapy (cART) virally suppressed patients11,12.
Global prevalence of HBV infection in people living with HIV has been recently estimated to be 7.6%, with the greatest burden in sub-Saharan Africa and in people who inject drugs, with odds of HBV infection 1.4 times higher as compared to HIV-negative people13.
Global prevalence of HCV infection in people living with HIV has been estimated to be 2.4% in the general population, increasing up to 82.4% when considering people who inject drugs. Odds of HCV infection are six times higher in HIV-infected people than in their HIV-negative counterparts14.
In prison, the prevalence of HBV and HCV infections among the HIV-seropositive inmates is higher than in the general population. This is due to several factors such as poor living conditions in jails, overcrowding, promiscuous intercourses, reduced perception of risk of infection, and absence of an effective health policy15, but also to the greater prevalence of injecting drug users16.
Worldwide, HBV and HCV co-infections among HIV-people in prison have been estimated to be 12% and 62%, respectively17. However, prevalence rates increase to 15% and 78%, for HBV and HCV infections, respectively, among the HIV-positive drug-injecting inmates17.
A highly variable prevalence of HIV and viral hepatitis infections has been reported in Italian prisons18,19. This varies for HBV and HCV depending on the number of drug users and immigrants present in each detention center. An Italian multicentre study showed, for example, infection of HBV/ HCV and HIV/ HCV, among inmates of 0.4% and 0.9%, respectively19. Furthermore, in the last few years HCV treatment with direct acting antiviral (DAA) seems to be achievable even in prison settings, given the short duration of treatment and its high efficacy. This intervention is important to reduce the circulation of HCV and represent an effective prevention strategy20.
Regarding HBV infection, the most frequent genotypes in the Italian population are D and A21, although other genotypes have been introduced due to immigration, mainly from Africa22,23. Concerning the HCV genotype, distribution of variants differs between Italian and non-Italian national populations, genotype 4 is much more prevalent among non-Italian residents whereas genotypes 1b and 2 are the most frequent ones among Italian natives24.
The prevalence of HIV, HBV and HCV infections in detention centers raises concerns for their spreading within inmates and to the general population. Thus, although public health interventions in jails have been undertaken to limit spreading of these infections, there are limitations due to the lack of adequate information technology, severe budget constraints, frequent inmate transfers among prisons, absent or poor clinical information-sharing among facilities, and the lack of an effective correctional healthcare database25,26.
Health programs may limit the spread of HIV, HBV and HCV infection and the circulation of the different genetic forms of these viruses, as well as of the variants with mutations conferring resistance to treatment. Study of the drug-resistant pattern may, in fact, provide opportunities for improvements of prevention practices and transmission of drug-resistant viruses.
Here we describe a cross-sectional multicenter study to assess the epidemiological, clinical and molecular characteristics of HIV, HBV and HCV infections in HIV-infected inmates during the period 2017–2019, in order to provide date required for targeted public health interventions.
Results
HIV infection
Table 1 reports the demographic, clinical, immunological, virological and behavioral characteristics of the study population.
Fifty-eight out of 65 inmates were males (89.2%) and seven females (10.8%). Median age was 45 years with an Interquartile Range (IQR) of 13 years. Forty-five inmates (69.2%) were Italian nationals and 20 (30.8%) were nationals of other countries including 6 inmates (9.2%) from sub-Saharan Africa (Nigeria and Liberia), 6 (9.2%) from East Europe (Albania, Croatia, Macedonia and Romania), 5 (7.7%) from North Africa (Tunisia, Morocco and Egypt), and 3 (4.6%) from Latin-America (Peru and Brazil).
All participants were on cART for HIV but with different treatment regimens. For 9 inmates, details on the drug regimen were not available. Despite the cART, nine inmates (20.4%) were HIV viremic with a range of detectable HIV plasma viral load from 41 to 85,314 copies/ml. Median lymphocyte counts were 2216 cells/µl, 621 cells/µl and 974 cells/µl, for total lymphocytes, CD4+-T lymphocytes and CD8+ T lymphocytes, respectively.
The declared behaviors at risk for blood-borne virus infections were abuse of drugs (injective and/or non-injective), unprotected heterosexual/homosexual intercourses and tattooing. All inmates reported having unprotected intercourses or being drug addicts or having both the at-risk behaviors; specifically, 41.5% of inmates declared drug abuse together with having unprotected sexual (homo- and/or hetero-sexual) intercourses, whereas 40% reported only the drug abuse, and 15.4% only unprotected intercourses.
To investigate the HIV genetic forms, a phylogenetic analysis was carried out using sequences of the HIV-1 Protease (PR)-Reverse Transcriptase (RT) regions of the Pol gene from 48 HIV-infected inmates (Fig. 1, panel A). The Maximum Likelihood (ML) analysis showed that 37 individuals were infected with the HIV-1 B subtype (77.1%), 3 with the sub-subtypes A1, A3 and A6, respectively (2.1% each subtype), and 8 with recombinant forms (16.7%). In particular, 2 inmates were infected with the CRF02_AG (4.1%), 2 with the CRF72_BF1 (4.1%), and 4 with only one among CRF63_02A6, CRF71_BF1, CRF108_BC and CRF06_cpx (2.1% each CRF), respectively (Fig. 1, panel B). Twenty-nine (60.4%) of the 48 characterized inmates were Italian nationals: 27 (93.1%) were infected with B subtype, while the remaining 2 (6.9%) were infected with non-B subtype (A6 and CRF72_BF1). Among the 19 (39.6%) inmates of non-Italian nationality, 10 (52.6%) were infected with HIV-B subtype, 7 (36.8%) with the CRFs, and 2 (10.5%) with the sub-subtypes A1 and A3, respectively.

ML phylogenetic tree of HIV RT-PR sequences obtained from 48 HIV-1-infected inmates and genetic forms distribution. Panel a: The phylogenetic tree was inferred by using the Maximum Likelihood (ML) method and the General Time Reversible model71. The tree with the highest log likelihood (- 20,708.86) is shown. The analysis involved 138 nucleotide sequences, including 48 infected inmates and 91 sequences from the genome reference set available at https://www.hiv.lanl.gov/ (see Methods for details on the reference set construction). Analyses were conducted in MEGA X71. In the tree, clades including sequences from inmates are highlighted in different colours: A1, fuchsia; A3, bright green; A6, brown; B, orange; CRF71_BF1 light blue, CRF72_BF1 red, CRF108_BC green, CRF06_cpx purple, CRF63_02A6 pink, CRF02_AG blue. Clades not including sequences from inmates are represented as cartoons (BF CRFs are indicated with the following labels: BF#: CRF44_BF, CRF38_BF, CRF89_BF, CRF17_BF and CRF12_BF; BF@: CRF40_BF and CRF71_BF). Red labels and black labels indicate sequences from inmates of non-Italian and Italian nationality, respectively. The number indicates the sample code, the first letter is an arbitrary code indicating the detention center, followed by an acronym indicating the risk behavior (HET: unprotected hetero intercourses; MSM: men who have sex with men; DU: drug users; T: tattoo). Diamonds highlight nodes with a bootstrap value > 70%. Panel b: The pie chart shows the distribution of HIV-1 subtypes and recombinant forms in 48 HIV-infected inmates.
We also investigated the presence of mutations in the PR, RT and the Integrase (IN) regions of the HIV pol gene conferring resistance to HIV drugs (detected according to the 2021 HIV drug resistance database of the Stanford University (https://hivdb.stanford.edu/). Seven out of the 48 subtyped inmates (14.6%) presented Major Drug Resistance mutations (DRMs) (Table 2). In 6 of these we found DRMs in the PR and RT regions, and in one inmate a DRM only the IN region. These DRMs were all found in B-subtypes strains isolated from inmates of Italian nationality, but one that was found in a CRF02_AG strain isolated from a inmate of Nigerian nationality. Table 2 shows the DRMs found in the 7 HIV subtyped inmates, according to the drug class mutation and to the current and previous antiretroviral treatment.
In particular, K70R and M184I major mutations, associated with Nucleoside Reverse Transcriptase Inhibitors (NRTIs) resistance, were detected in three inmates treated with NRTI -based regimens, whereas the major mutation M184V, associated with NRTI resistance, was detected in an inmate exposed to Protease Inhibitor (PI) + Integrase Strand Transfer Inhibitors (InSTI) and never exposed to NRTIs.
Further, the K103N mutation, associated with Non-Nucleoside Reverse Transcriptase Inhibitors (NNRTI), was detected in an inmate exposed to NRTI + PI and never exposed to NNRTIs.
Finally, the I84V mutation, associated with PI resistance, was present in one inmate treated with PI and in another inmate never exposed to PI. Lastly, the R263K mutation, associated with InSTI resistance, was detected in an inmate exposed to PI + NRTI and never exposed to InSTIs.
Of note, the K70R and I84V mutations were present in the same sequence from a single inmate.
HBV and HCV coinfections
Inmates were also tested for the presence of HBV and HCV serological markers (Table 3). Four inmates (6.2%) had the HBsAg marker. Twenty-three inmates were positive for the anti-HBs marker (35.4%) and at least another HBV markers with two serological profiles observed (see Table 3), whereas 14 inmates (21.5%) showed the presence of an isolated anti-HBs marker. In agreement with the CDC suggested interpretation of the HBV serologic test results (https://www.cdc.gov/hepatitis/hbv/pdfs/serologicchartv8.pdf), these latter inmates were considered as vaccinated for HBV. Twenty five inmates out of 65 (38.4%) were positive for anti-HBc associated with other HBV markers with two serological profiles observed (see Table 3), whereas 11 inmates out of 65 (17%) showed the presence of an isolated anti-HBc marker. This serological pattern [anti-HBc(+), HBsAg(−) and anti-HBs(−)] often indicates past exposure to HBV with waning anti-HBs immunity, and it is found commonly in HIV-infected individuals, particularly those co-infected with HCV. In fact, 9 of them (81%) were co-infected with HCV (data non shown). No inmates showed the presence of HBV DNA.
Regarding HCV serological markers, 39 inmates out of 65 (60.0%) were positive for anti-HCV antibodies, and 18 out of 65 (27.7%) also showed the presence of HCV RNA in plasma. Of these latter, 14 inmates were positive for both HBV and HCV markers, 3 were positive for both isolated anti-HBs and HCV markers and one only for HCV marker, respectively (data not shown). Notably, out of 39 individuals, 13 received anti-HCV DAA-based treatments (33%), 5 spontaneously cleared the virus (13%), 12 had no therapy (31%) and for 9 individuals no data were available (23%) (data not shown).
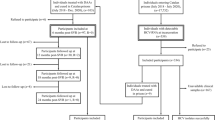
Figure 2 shows the distribution of HBV and/or HCV coinfections and HBV vaccination in the HIV-infected inmates. In detail, 14 inmates (21.5%) showed only HBV markers; of these, 10 (15.4%,) showed HBV infection markers, while the others 4 (6.2%) were vaccinated for HBV. Among the 12 inmates (18.5%) who showed the presence of only HCV infection markers, 2 (3.1%) were co-infected by HCV, while 10 (15.4%) were HCV co-infected and HBV vaccinated. Twenty-seven (41.5%) were HBV/HCV doubly co-infected (previous or current infection). Finally, 12 individuals (18.5%) were negative for all HBV and HCV markers tested, and had no evidence of current or past co-infection.

HBV and/or HCV infection and HBV vaccination in 65 HIV-infected inmates. The flow chart shows the prevalence of HBV and HCV infection in 65 HIV-infected inmates. In red the final estimates for HBV and/or HCV infection are indicated.
The association of the presence of HBV or HCV infection markers with demographic variables was then investigated (Table 4). HBV markers were not significantly associated with either age or nationality of inmates, while the percentage of inmates positive for HCV markers was higher in older than in younger inmates (74.2% vs 44.4%, respectively, p-value = 0.0209). The presence of HCV marker was also more frequently associated with the Italian nationality (75.6% in Italian nationals vs 25.0% in non-Italian nationals, p-value = 0.0001).
In order to evaluate determinants associated with the risk of having at least one co-infection, univariate and multivariate ordered logistic models were performed. In the univariate model, non- Italian nationals appeared to be at more risk of having co-infections, whereas higher CD8 + T cell counts, the elderly and the drug abusers, whether or not combined with unprotected intercourses, had a higher risk of being infected with HIV alone. The multivariate model confirmed the CD8 + T-cell counts, age and drug abuse effects, while nationality seemed not to be a true risk factor for having co-infections (Table 5).
Figure 3 reports the phylogenetic tree inferred from HCV NS5B sequences. Sequences were obtained from 18 individuals showing the HCV RNA marker in plasma (Fig. 3, panel a). Sixteen of these were Italian nationals. The subtype 1a was by far the most frequent since it was detected in 12/18 inmates (66.6%); subtype 3a and subtype 4d were equally detected in 3/18 inmates (16.7%) (Fig. 3, panel b). The two inmates of non-Italian nationality were from Morocco and Macedonia and were infected by subtype 1a and 3a strains, respectively. Analysis of risk factors referred by the 18 HCV infected inmates showed that all but two were drug users. However, 12 of them also reported unprotected sex.

ML phylogenetic tree of HCV NS5B sequences obtained from 18 HCV infected inmates and genetic forms distribution. The tree shows the phylogenetic relationships among the 18 HCV sequences from inmates and 20 reference sequences, representing the major known HCV genotypes/subtypes, downloaded from Los Alamos HCV (https://hcv.lanl.gov/content/index). The best substitution model (T92 + G + I) for the dataset under analysis was preliminarily assessed by the Models function in MEGA. Panel a: subtypes including sequences from inmates are highlighted in different colours: 1a, orange; 3a, blue; 4d, green. The number indicates the sample code, the first letter is an arbitrary code indicating the detention center, followed by an acronym indicating the risk behavior (HET: unprotected hetero intercourses; MSM: men who have sex with men; DU: drug users; T: tattoo). Sequence labels from inmates of non-Italian nationality are reported in red, those from inmates of Italian nationality in black. Diamonds highlight nodes with a bootstrap value > 70%. A black diamond (♦) marks statistically supported nodes (bootstrap value > 70%). Panel b: prevalence of HCV NS5B subtypes in 65 HIV-infected inmates. The prevalence of the genetic forms is expressed as the percentage of the total number.
All samples were negative for HBV DNA.
Discussion
Our study proposes to analyze the inmate population that represents a model of closed community subject to restrictive conditions and characterized by a multinational composition and a high incidence of disease. In prison, emerging problems such as drug use, HIV/AIDS and other infectious diseases are often present and they must be addressed in both Italian and non-Italian national inmates.
For an accurate surveillance, in prison the monitoring of viral epidemiological changes and circulation of virus strains are pivotal for both risk groups and the general population.
In our study, 30.8% of the enrolled inmates were of non-Italian origin. This data is in line with the 2018 official data (the average time period of sample collection) from the Ministry of Justice, which indicated a 33.9% of inmates of non-Italian nationality (https://www.giustizia.it/giustizia/it/mg_1_14_1.page?facetNode_1=1_5_40&contentId=SST165666&previsiousPage=mg_1_14).
It is well known that HIV-1 subtype B is predominant in Western Europe, including Italy, North America and North Africa27,28. However, in recent years, the frequency of non-B clade forms has been reported to increase in subtype B-restricted geographical areas and in Italy6,29.
The HIV strains phylogenetic analysis showed that the majority of inmates of Italian nationality were infected with the HIV-1 subtype B, except two, one infected with the recombinant form CRF72_BF1, and the other one with the HIV-1 sub-subtype A6, both previously not identified in inmates of Italian nationality30,31. CRF72_BF1 was isolated in Brazil32, and it has spread in Spain. HIV-1 sub-subtype A6 is present in the Former Soviet Union (FSU) countries likely originated from A1 strains of African origin, and is now spreading into non-FSU countries33. The presence of new strains within a closed community, such as a prison, can become a public health problem because this would allow the spread of new drug-resistant variants.
Conversely, a different pattern was found in the non-Italian inmate population. Ten out of 19 inmates of non-Italian nationality, were infected by subtype B strains. They came from countries where HIV-1 subtype B circulates with the exception of Liberia and Nigeria in which only very few cases of subtype B have been described in the recent past [Los Alamos https://www.hiv.lanl.gov/components/sequence/HIV/geo/geo.html accessed on march 7, 2023; 29,34]. Of the other non-Italian inmates, six were infected with subtypes and CRFs circulating in their countries of origin, and three were infected with CRF71_BF135, CRF72_BF132 and CRF108_BC36 respectively. Of note, these CRFs were isolated from inmates coming from Tunisia, Romania and Albany, where circulation of these three recombinant forms has not been previously described.
Despite considerable success in the treatment of HIV-1 infection, there continues to be a growing concern about the emergence of HIV-1 drug resistance mutations (DRMs) that can compromise the effectiveness of antiretroviral drugs. It has been described earlier that the prevalence of DRMs in cART naive individuals varies from 0 to 15% and may depend on several variables such as geographical area, HIV prevalence, transmission route and virus subtype37,38,39,40. However, it must be kept in mind that various factors, such as early treatment, use of appropriate drug regimens, and good adherence to therapy play a major role in blocking the emergence of DRMs41.
Since DRMs do not appear during ART in patients with suppressed viremia42, HIV-infected ART-naive individuals are the major source of drug-resistant viruses in both the developed and low- and middle-income countries43,44. Consequently, the DRMs, that can persist for a significant period of time even in the absence of drug treatment, can be transmitted (TDRMs)45. DRMs can be selected during antiviral treatment and persist in proviral DNA as integrated forms in viral reservoirs. Consequently, if treatment is interrupted or adherence compromised, there is a risk that the reactivated virus is drug resistant, potentially facilitating transmission to co-infection partners and hampering the effectiveness of antiviral treatments45.
In our study, we found DRMs in 7 HIV-1 subtyped inmates (6 with subtypes B isolated from inmates of Italian nationality, and 1 with CRF02_AG isolated from a inmate of Nigerian nationality). Six inmates had major DRMs in the PR and RT genes, and one inmate had major DRM in the IN gene. Of note, two mutations, K70R and I84V, were present in the same sequence from a single inmate. The simultaneous presence of multiple DRMs within the same viral genome can lead to a significant increase in resistance to antiviral drugs, as well as cross-resistance to inhibitors of the same class46. Three out of 6 inmates showed resistance mutations to drugs belonging to classes never administered to them. For one inmate the previous therapeutic regimen was unknown, therefore it cannot be excluded that the DRM could be related to the previous therapy. One inmate, treated with PIs and with InSTI showed a resistance mutation to NRTI drugs. It has been recently described that transmitted mutations to NRTI drugs in patients treated with InSTI can increase the risk of viral failure if treated with InSTI-based regimens47. We also found an inmate with the R263K mutation against InSTIs, a class of drugs never administered to him, which confers low-level resistance to InSTIs48,49. Other authors have also described a very low prevalence of transmitted drug resistance to InSTIs50,51,52. However, it has been observed that increased resistance rates is associated with incomplete adherence to therapy and low CD4 T-cells regardless of which INSTI was administered53. Thus, baseline resistance to INSTIs should be evaluated. In the case of emergence of DRMs to one class of drugs the retreatment with an antiviral of the same class could cause poor response, suggesting that switching to another combination type/class of drugs is preferable46. However, it must be appreciated that there are resistance mutations that may appear in patients receiving drugs that affect regions other than those in which the mutations were found. As in the case of DDA resistance for HCV infection, we can define these extra target mutations46. This suggests that DRM studies are important when deciding to treat HIV-infected inmates.
We found that 75.4% of HIV-infected inmates showed also markers of HBV and/or HCV infection. In particular, 56.9% of individual had infection markers for HBV, 60.0% for HCV and 41.5% + for both HBV and HCV co-infection. Of note, 14 inmates (13 Italian nationals and 1 Peruvian national, 21.5%) were found positive only for anti-HBs antibodies. Therefore, although we did not have information on the HBV vaccination status of the 65 inmates, we considered these inmates as vaccinated for HBV, as suggested by the CDC (https://www.cdc.gov/hepatitis/hbv/pdfs/serologicchartv8.pdf). Although the Ministerial Decree n. 251 of 25 October 1991 specifies that vaccination against hepatitis B must be administered to all prisoners, this is severely hampered by the high turnover of inmates in Italian prisons. For this reason, rapid vaccination schedules have been proposed and are being applied54,55. Stasi and colleagues found a similar prevalence of inmates (15.9%) with isolated anti-HBs marker in detention facilities in the Tuscany region55, and considered them as vaccinated.
We found that HBV infected inmates were mostly Italian nationals (81.3% vs 57.9% of non-Italian nationals). This highlights the importance of screening for HBV in accelerating vaccination in order to ensure short-term protection in the prison population55.
In our study, 11 (17%) inmates showed an isolated anti-HBc marker. This prevalence is in line with the frequency of anti-HBc marker in HIV-1-infected patients observed by Chang and colleagues. This frequency varied from 17 to 40%, representing, in most of cases, a waning host immune response to HBV infection that can result in HBV reactivation with varying consequences in morbidity56. Nine out of 11 inmates in our study showed an isolated anti-HBc pattern associated to HCV marker. The isolated anti-HBc pattern is particularly relevant in HIV/HCV co-infected patients and, most often, HCV is dominant, suppressing HBV replication57. When HCV is treated, its inhibitory effects on HBV replication is released possibly resulting in HBV reactivation. In fact, HBV reactivation has emerged in the era of DAA treatment of HCV infection. Due to this, screening for HBV should be performed also in prison prior to initiation of HCV therapy.
In our study, HBV sequencing was not attempted since all samples were found to be negative for HBV DNA. This was expected because HIV treatment includes inhibitors of reverse transcriptase that are also effective against the reverse transcriptase activity of the HBV polymerase.
Our data confirm the results of other studies that showed a high prevalence of HIV/HCV coinfection in inmates. HCV co-infection was significantly associated with the Italian nationality. In Italy, a high prevalence of HCV infection has been reported in drug users, both in the general and in the inmate populations58,59,60. In fact, we found a higher prevalence of this coinfection in inmates older than 45 years (74.2%), corresponding to inmates born in the years at major risk for drug abuse, one of the main transmission routes of HIV/HCV. The multivariate model showed that higher CD8 + T-cell counts, elderly age and drug abuse were associated with HIV infection alone, while nationality seemed not to be a risk factor for having co-infections. In agreement with our data, Schmidt et al.61 reported CD8 T-cell failure in HCV and HBV infections, due to T-cell exhaustion, deletion, and viral escape.
The prevalence of HBV and HCV infection markers in HIV-infected prisoners, although high, is lower than what we found in our previous study31. The greatest difference was observed considering the HCV infection that was present in 60.0% of inmates in this study, as compared to the 78.3% of the previous study. Similarly, 27.7% of inmates had HCV RNA, a much lower prevalence compared to the 65.2% found in our previous work. It must be specified that inmates enrolled in our previous work were not under anti-HCV DAA-based treatments31. Therefore, even if the detention centers involved in our two studies are different, we can speculate that the lower prevalence of HCV markers can be, at least partially, due to administration of DAAs against HCV, as observed in our present study and as already reported by others62,63. A recent study indicates that the prevalence of HCV markers among inmates is less than 20%, possibly due to the availability of DAAs64. However, other factors may also play a role, such as the improving strategies for the control of infectious diseases, and/or a different distribution of behaviors at-risk for HCV infection among the detention centers. More tailored studies are needed to evaluate the efficacy of DAAs drugs in reducing HCV infection prevalence in prison.
HCV genotype and subtype heterogeneity may be influenced by the route of infection and coinfection, regardless of the geographic variable65,66. In this work we found that the subtype 1a is the most frequent variant (66% prevalence), followed by subtypes 3a and 4d that are equally represented with a prevalence around 16.7%. In our previous study31, we found a similar HCV subtype distribution with the exception of the prevalence of HCV subtype 3a which was higher than in the present study. In addition, in both studies, we found a similar prevalence of the 4d subtype. This distribution of HCV subtypes is in line with the distribution observed in the population of drug addicts outside the prisons, both in Italy and in Europe58,67. However, in the general population, subtype 1b is predominant with a prevalence of more than 50%, followed by genotype 2 (35%), and by subtypes 1a and 3a that show a very low prevalence68.
Some limitations of our study should be addressed. Firstly, our study was based on voluntary enrollment and, therefore, we were unable to obtain an estimate of the percentage of HIV-positive people who were enrolled in the study, as compared to the total HIV-positive people in the 5 detention facilities. However, the prisons participating in the study were uniformly distributed in all the Italian territory.
Another limitation of our study is that HIV sequences were obtained from proviral DNA since the majority of the enrolled inmates had negative plasma viral load. In fact, the proviral DNA sequence can contain a variety of multiple DRMs, not present in plasma viral RNA69, which may thus reveal viral variants with mutations that are not actively expressed70. However, HIV DNA sequencing may provide a good method to gain unique preliminary insights into HIV‐1 subtype diversity or for an overall evaluation of HIV DRM in a population70. Data from previous studies have also shown that the proviral compartment can be reliably used for the investigation of DRMs in ART-naive patients71.
Finally in our study, we utilized Sanger sequencing as the standard HIV genotyping method. However, this methodology may not detect low–abundance HIV variants mutations, that are less represented within the viral pool. Although Next Generation Sequencing (NGS) is a more sensitive method capable of identifying minor resistant mutants within viral populations72,73, a high concordance exists between Sanger sequencing and NGS. Nevertheless, further investigation are needed for detection of low‐abundance HIV‐1 variants by NGS.
In conclusion, our data indicate the need for a systematic screening and monitoring both at the beginning of incarceration and during incarceration to limit the spread of HIV, HCV, and HBV.
Although public health interventions are underway, effective prevention programs should be implemented that can limit the spread and circulation of the different genetic forms of these viruses, in prisons and to the general population, as well as of variants with mutations that confer resistance to treatment.
Materials and methods
Study population
A multicenter study was performed from 2017 to 2019 in detention centers of 5 Italian cities (from North to South: Brescia, Milan, Civitavecchia, Viterbo, Sassari). The study was approved by the ISS ethical committees [prot. PRE-866/16 (November 8th, 2016)] and the local ethical committees [Viterbo, Prot. n. 769/CE Lazio1 (March 31st, 2017); Sassari, Prot. n. 2495/CE (May 23rd, 2017); Brescia, Prot. n. 2699 (May 26th, 2017); Civitavecchia, Prot. n. 2287/CE Lazio 1 (November 30th, 2018), Milan, Prot. n. ST-173, 2018 (November 17th, 2018)]. All participants gave written informed consent in accordance with the Declaration of Helsinki. Demographic, behavioral, clinical, immunological and virological data were collected for each patient in complete anonymity in each center in a case report form provided by the National HIV/AIDS Research Center (CNAIDS) of the Istituto Superiore di Sanità (ISS). Sixty-five inmates already diagnosed with HIV infection accepted to participate to the study. After signature of the informed consent, 2 ml of plasma from whole blood, collected in EDTA during the routine testing, were obtained, using standardized procedures in each clinical center. Plasma samples and blood components were kept at-80 °C in each center until shipment to CNAIDS (ISS) in Rome for serological HBV and HCV analyses and sequencing of HIV, HBV and HCV.
Serological assays
Anti-HCV, anti-HBc (IgM and IgG) and HBsAg markers were determined by chemiluminescent assays on an automated analyzer (COBAS Elecsys e401, Roche Diagnostics, Basel, Switzerland), using Elecsys anti-HCV II, Elecsys anti-HBc and Elecsys HBsAg II kits, the last one with a sensitivity of 20–30 mUI/ml. Anti-HBs antibodies were detected by Enzygnost Anti-HBs II (Siemens Healthcare Diagnostics Products, Germany) and results evaluated according to the manufacturer's instructions.
Viral nucleic acid quantification
HIV viral load was determined at each center where enrollment was performed, as a common routine analysis for HIV-infected inmates, using different commercial kits.
HBV DNA quantification was performed at ISS on plasma using the High Pure System Viral Nucleic Acid Kit (Roche Diagnostics, Basel, Switzerland), followed by amplification and detection on a COBAS AmpliPrep/COBAS TaqMan Instrument using the COBAS Taq Screen MPX Test v2.0; this assay is currently used for blood screening and has a sensitivity of 2.1 IU/ml.
HCV RNA quantification was performed on plasma using the High Pure System Viral Nucleic Acid Kit (Roche Diagnostics, Basel, Switzerland), at ISS, followed by amplification and detection on a COBAS TaqMan 48 Analyzer using the COBAS TaqMan HCV Test v2.0; this test has a sensitivity of about 9.3 IU/ml and a linear range from 25 to 3.91 × 108 IU/ml.
Virus genotyping
According to data reported by each center, a few inmates had a detectable HIV viral load, most of them being under successful cART. HIV subtyping was therefore carried out on HIV proviral DNA. To this purpose, HIV proviral DNA was extracted from 400 ul of whole blood using the QIAamp DNA blood mini kit (Qiagen, Hilden, Germany) according to the manufacturer’s instructions. A portion of the pol gene, encompassing the Protease and Reverse Transcriptase-encoding region (the PR-RT region) was amplified by a nested Polymerase Chain Reaction (PCR), following a previously described protocol74, with modifications. Briefly, primers for PR-RT region amplification were for PCR outer: NES3 5’GAC AGG CTA ATT TTT TAG GG 3’ located at 2075–2094 (gag); NS4 5’ GGC TCT TGA TAA ATT TGA TAT GT 3’ located at 3561–3583 (pol),; PCR inner FK1 5’ AGC AGA CCA GAG CCA ACA GC 3’(2140–2159 gag), FK7 5’ CTA TTA AGT CTT TTG ATG GGT CA 3’ (3506–3528 pol) (with reference to the HIV-1 HXB2 strain from the Los Alamos National Laboratory database using the Sequence Locator and QuickAlign tools: http://www.hiv.lanl.gov/content/ sequence/HIV/mainpage.html).
The region of the pol gene encompassing the Integrase-coding region (nucleotides 4150 to 5263 according to the HIV-1 HXB2 sequence) was also amplified. To this purpose, primers for the nested PCR amplification were the following: PCR outer FS1:5’-CATGGGTACCAGCACACAAAG-3’FS6: 5’-CTGTATGCAGACCCCAATATG-3’; PCR inner FS2:5’-GGAGGAAATGAACAAGTAGA-3’ FS5: 5’-GGGATGTGTACTTCTGAACT-3’. The optimized thermocycler conditions for outer and inner PCR assays were: initial denaturation at 94 °C for 2′, 37 cycles at 94 °C for 1′, 55 °C for 1′, 72 °C for 2′ and a final extension at 72 °C for 2′75.
The resulting amplicons were purified using Gel Extraction Kit (Qiagen, Hilden, Germany) and directly sequenced using an ABI 3730 automated sequencer.
For HBV, no patient showed the presence of HBV DNA (HBV DNA below the limit of detection of the test), therefore amplification of HBV genome sequences was not attempted.
For HCV, viral RNA was extracted from 140 μl of serum samples using the QIAamp Viral RNA Extraction Kit (Qiagen, Hilden, Germany), according to the manufacturer’s instructions. Healthy donor serum samples were used as a negative control. The RNA was reverse transcribed using the High-Capacity cDNA Reverse Transcription Kit protocol (Thermo Fischer Scientific, Waltham, Massachussetts, US) and the resulting cDNA amplified by nested PCR using the FastStart High Fidelity PCR system (Roche Diagnostics, Basel, Switzerland). The specific primers used to amplify the NS5B region (nt 8256–8632) of HCV for the first and second rounds have been previously described76. PCR conditions for both rounds were 94 °C for 2’, followed by 28 cycles of denaturation at 94 °C for 15″, annealing at 60 °C for 30″, extension at 72 °C for 45″, and a last extension step at 72 °C for 7′. PCR products were analyzed on a 2% agarose gel stained with GelRed (Biotium). Both strands were sequenced using the Genome Lab DTCS Quick Start KiT (Beckman Coulter, Inc., Fullerton, CA). Purified sequencing reactions were run on an automated DNA sequencer (Beckman Coulter, Inc., Fullerton, CA).
HIV and HCV phylogenetic analyses
To perform HIV phylogenetic analyses, a genome reference set was constructed starting from the reference sequences available at the HIV Los Alamos database (www.hiv.lanl.gov) that includes 428 genomes. The sequences were aligned by Clustal Omega (https://www.ebi.ac.uk/Tools/msa/clustalo/).
and a redundancy removal procedure was performed. On the basis of the identity matrix for each pair of sequences with a percentage of identities (%ID) higher than 95%, only one sequence was considered. This resulted in a set of genome sequences including 46 pure subtypes and sub-subtypes (A1-A6 12; B 5; C 3; D 4; F1-F2 7; G 4; H 4; K 2; J 3; L 2) and 186 CRFs. These sequences were aligned with the 48 PR-RT sequences obtained from 48 inmates by Clustal Omega. The resulting alignment were manually edited by using BioEdit 7.277 and gaps were removed. A phylogenetic analysis was carried out by MEGA X by using the Maximum Likelihood (ML) method and the General Time Reversible (GTR) model78, the statistical robustness and reliability of the inferred tree was estimated by a bootstrap test (1000 bootstrap replications), and it was visually inspected to remove CRF sequences having no phylogenetic relationships with our 48 sequences. The resulting final reference set comprises 91 sequences (including 46 pure subtypes and sub-subtypes and 45 CRFs). The phylogenetic inference procedure (ML method; GTR model; 1000 bootstrap replications) was repeated on the novel set of sequences to assign inmate sequences to subtypes.
The HCV genome reference set, representing the major known HCV genotypes/subtypes, includes 20 reference sequences downloaded from the HCV Los Alamos databases ((www.hcv.lanl.gov). These sequences were aligned to 18 inmates sequences by ClustalW in BioEdit 7.277, the resulting multiple alignment was manually edited and gaps were removed. Genotype assignments were achieved by constructing a phylogenetic tree inferred using a ML method by MEGA X, using the best substitution model describing the observed data, preliminarily determined by the Models function in MEGA X. The statistical robustness and the reliability of the phylogenetic tree were confirmed by bootstrap analysis using 1000 replicates.
The resulting phylogenetic trees for HCV and HIV data sets were visualized by Fig Tree 1.4.4 (http://tree.bio.ed.ac.uk).
Determination of HIV Major Drug Resistance Mutations (DRMs) against HIV ART
The presence of major DRMs against HIV ART in the protease, RT and integrase genes was investigated in all inmates, for whom either the PR-RT or the Integrase region nucleotide sequences, or both, were known, according to the WHO-2009 list of resistance mutations79, the 2019 IAS-USA Drug resistance mutations in HIV-1 list80, and the Stanford HIV Drug Resistance database (Stanford University, version 9.0, 2021; https://hivdb.stanford.edu/hivdb/by-mutations/)].
Nucleotide sequence accession numbers
The nucleotide sequences of the HIV-1 protease and reverse transcriptase (PR-RT) regions and of the HCV NS5B region obtained from all subtyped inmates are available in GenBank, with the following accession numbers ON611640-ON611686 (HIV-1 PR-RT) and ON055295-ON055312 (HCV NS5B).
Statistical analysis
Descriptive statistics summarizing quantitative variables included median and interquartile range (IQR). Qualitative variables were presented as absolute and relative frequencies. Two-tailed Pearson’s chi-squared test was used to evaluate the difference in the prevalence of viral hepatitis markers between groups based on demographic. p-values less than 0.05 were considered statistically significant.
Univariate and multivariate ordered logistic models were performed in order to evaluate determinants associated to the risk of having 1 or 2 coinfections vs only HIV. In the multivariate model all the variables are each other adjusted.
All the statistical procedures were performed using the SAS®, Version 9.4 (SAS Institute Inc. 100 SAS Campus Drive Cary, NC, USA).
Ethical approval
The study was approved by the ISS ethical committees [prot. PRE-866/16 (November 8th, 2016)] and the local ethical committees [Viterbo, Prot. n. 769/CE Lazio1 (March 31st, 2017); Sassari, Prot. n. 2495/CE (May 23rd, 2017); Brescia, Prot. n. 2699 (May 26th, 2017); Civitavecchia, Prot. n. 2287/CE Lazio 1 (November 30th, 2018), Milan, Prot. n. ST-173, 2018 (November 17th, 2018)]. All participants gave written informed consent in accordance with the Declaration of Helsinki.
Data availability
The datasets generated and analysed during the current study are available in the NCBI repository (https://www.ncbi.nlm.nih.gov/genbank/; Accession numbers: ON611640-ON611686 for HIV-1 PR-RT sequences and ON055295-ON055312 for HCV NS5B).
References
Dolan, K. et al. Global burden of HIV, viral hepatitis, and tuberculosis in prisoners and detainees. The Lancet 388, 1089–1102 (2016).
Dolan, K. et al. HIV in prison in low-income and middle-income countries. Lancet Infect. Dis. 7, 32–41 (2007).
Babudieri, S. B. et al. Correlates of HIV, HBV, and HCV infections in a prison inmate population: Results from a multicentre study in Italy. J. Med. Virol. 76, 311–317 (2005).
Maggiorella, M. T. et al. Italian network for HIV characterization. High HIV-1 diversity in immigrants resident in Italy (2008–2017). Sci. Rep. 10, 3226 (2020).
Lai, A. et al. Changing patterns in HIV-1 non-B clade prevalence and diversity in Italy over three decades. HIV Med. 11, 593–602 (2010).
Ciccozzi, M. et al. HIV-1 non-B subtypes in Italy: A growing trend. New Microbiol. 5, 377–386 (2012).
Lorenzin, G. et al. Prevalence of non-B HIV-1 subtypes in North Italy and analysis of transmission clusters based on sequence data analysis. Microorganisms. 23;8(1), 36 (2019).
Sarmati, L. & Malagnino, V. HBV infection in HIV-driven immune suppression. Viruses 11, 1077 (2019).
Merchante, N., Jiménez-Saenz, M. & Pineda, J. A. Management of HCV-related end-stage liver disease in HIV-coinfected patients. AIDS Rev. 9, 131–139 (2007).
Kim, H. N. et al. Treatment of HCV in Persons with HIV Coinfection. Hepatitis C Online. https://www.hepatitisc.uw.edu/go/key-populations-situations/treatment-hiv-coinfection/core-concept/all. Last updated: February 17th (2021).
Moron- Lopez, S. et al. Short-term treatment with interferon alfa diminishes expression of HIV-1 and reduces CD4+ T-cell activation in patients coinfected with HIV and hepatitis C virus and receiving antiretroviral therapy. J. Infect. Dis. 213(6), 1008–12 (2016).
Curry, M. P. HIV and hepatitis C virus: Special concerns for patients with cirrhosis. J. Infect. Dis. 207, 40–44 (2013).
Platt, L. et al. Prevalence and burden of HBV co-infection among people living with HIV: A global systematic review and meta-analysis. J. Viral Hepat. 27, 294–315 (2019).
Platt, L., Easterbrook, P., Gower, E., McDonald, B. & Sabin, K. Prevalence and burden of HCV co-infection in people living with HIV: A global systematic review and meta-analysis. Lancet Infect. Dis. 16, 797–808 (2016).
Bick, J. A. Infection control in jails and prisons. Clin. Infect. Dis. 45, 1047–1055 (2007).
Falla, A. M. et al. Hepatitis B/C in the countries of the EU/EEA: A systematic review of the prevalence among at-risk groups. BMC Infect. Dis. 18, 79 (2018).
Ahmadi Gharaei, H. et al. The global and regional prevalence of hepatitis C and B co-infections among prisoners living with HIV: A systematic review and meta-analysis. Infect. Dis. Poverty 10, 93 (2021).
Sagnelli, E. et al. Blood born viral infections, sexually transmitted diseases and latent tuberculosis in italian prisons: A preliminary report of a large multicenter study. Eur. Rev. Med. Pharmacol. Sci. 16, 2142–2146 (2012).
Stasi, C., Silvestri, C., Fanti, E., Di Fiandra, T. & Voller, F. Prevalence and features of chronic viral hepatitis and HIV coinfection in Italian prisons. Eur. J. Intern. Med. 34, 21–22 (2016).
Pontali, E. et al. Treatment with direct-acting antivirals in a multicenter cohort of HCV-infected inmates in Italy. Int. J. Drug Policy 59, 50–53 (2018).
Zuccaro, O. et al. Clinical, epidemiological and virological features of acute hepatitis B in Italy. Infection 43, 431–441 (2015).
Coppola, N. et al. Factors affecting the changes in molecular epidemiology of acute hepatitis B in a Southern Italian area. J. Viral Hepat. 17, 493–500 (2010).
Sagnelli, E., Sagnelli, C., Pisaturo, M., Macera, M. & Coppola, N. Epidemiology of acute and chronic hepatitis B and delta over the last 5 decades in Italy. World J. Gastroenterol. 20, 7635–7643 (2014).
Quaranta, M. G. et al. Clinical features and comorbidity pattern of HCV infected migrants compared to native patients in care in Italy: A real-life evaluation of the PITER cohort. Dig. Liver Dis. 53, 1603–1609 (2021).
Babudieri, S. et al. HIV e infezioni correlate negli Istituti Penitenziari Italiani: Note di epidemiologia e di organizzazione sanitaria. Ann. Ist Super. Sanità 39, 251–257 (2003).
Dell’Isola, S. et al. Detention and incompatibility of HIV patients in Italy. Ann. Ist Super. Sanità 49, 332–333 (2013).
Hemelaar, J. et al. Global and regional epidemiology of HIV-1 recombinants in 1990–2015: A systematic review and global survey. Lancet HIV 7, 772–781 (2020).
Hemelaar, J. et al. Country level diversity of the HIV-1 pandemic between 1990 and 2015. J. Virol. 95, e01580-e1620 (2020).
Bbosa, N., Kaleebu, P. & Ssemwanga, D. HIV subtype diversity worldwide. Curr. Opin. HIV AIDS 14, 153–160 (2019).
Longo, B. et al. HIV-1 diversity among inmates of Italian Prisons. J. Med. Virol. 80, 1689–1694 (2008).
Sanarico, N. et al. Correlates of infection and molecular characterization of blood-borne HIV, HCV, and HBV infections in HIV-1 infected inmates in Italy: An observational cross-sectional study. Medicine (Baltimore) 95, e5257 (2016).
Pessôa, R., De Freitas Carneiro Proietti, A. B., Busch, M. P. & Sanabani, S. S. Identification of a novel HIV-1 circulating recombinant form (CRF72_BF1) in deep sequencing data from blood donors in Southeastern Brazil. Genome Announc. 2, e00386-14 (2014).
Abidi, S.H. et al. Origin and evolution of HIV-1 subtype A6. Plos One. 16 (2021).
Giovanetti, M., Ciccozzi, M., Parolin, C. & Borsetti, A. Molecular epidemiology of HIV-1 in African countries: A comprehensive overview. Pathogens 9(12), 1072 (2020).
Pessoa, R. et al. Deep sequencing of HIV-1 near full-length proviral genomes identifies high rates of BF1 recombinants including two novel circulating recombinant forms (CRF) 70_BF1 and a disseminating 71_BF1 among blood donors in Pernambuco, Brazil. PLOS ONE 9, 11 (2014).
Cañada, J. E. et al. Identification of a new HIV-1 BC intersubtype circulating recombinant form (CRF108_BC) in Spain. Viruses 13, 93 (2021).
Kim, M. H. et al. HIV antiretroviral resistance mutations among antiretroviral treatment-naive and -experienced patients in South Korea. AIDS Res. Hum. Retrovir. 29, 1617–1620 (2013).
Frentz, D. et al. Increase in transmitted resistance to non-nucleoside reverse transcriptase inhibitors among newly diagnosed HIV-1 infections in Europe. BMC Infect. Dis. 14, 407 (2014).
Jiamsakul, A. et al. Transmitted drug resistance in recently infected HIV-positive Individuals from four urban locations across Asia (2007–2010)—TASER-S. AIDS Res. Ther. 12, 3 (2015).
Alexiev, I. et al. Low HIV-1 transmitted drug resistance in Bulgaria against a background of high clade diversity. J. Antimicrob. Chemother. 70, 1874–1880 (2015).
Ndashimye, E., Reyes, P. S. & Arts, E. J. New antiretroviral inhibitors and HIV-1 drug resistance: More focus on 90% HIV-1 isolates?. FEMS Microbiol. Rev. 16(47), 1–22 (2023).
Pitman, M. C., Lau, J. S. Y., McMahon, J. H. & Lewin, S. R. Barriers and strategies to achieve a cure for HIV. Lancet HIV 5, e317–e328 (2018).
Baxter, J. D. et al. Global HIV-1 transmitted drug resistance in the INSIGHT Strategic Timing of AntiRetroviral Treatment (START) trial. HIV Med. 16, 77–87 (2015).
Gupta, R. K. et al. HIV-1 drug resistance before initiation or re-initiation of first-line antiretroviral therapy in low-income and middle-income countries: A systematic review and meta-regression analysis. Lancet Infect. Dis. 18, 346–355 (2017).
Bandera, A. et al. Phylogenies in ART: HIV reservoirs, HIV latency and drug resistance. Curr. Opin. Pharmacol. 48, 24–32 (2019).
Chen, Q. et al. Deep-sequencing reveals broad subtype-specific HCV resistance mutations associated with treatment failure. Antivir. Res. 174, 104694 (2020).
Borghetti, A. et al. Transmitted drug resistance to NRTIs and risk of virological failure in naïve patients treated with integrase inhibitors. HIV Med. 22, 22–27 (2021).
Quashie, P. K. et al. Characterization of the R263K mutation in HIV-1 Integrase that confers low-level resistance to the second-generation Integrase strand transfer inhibitor Dolutegravir. J. Virol. 86, 2696–2705 (2012).
Garrido, C. et al. Broad phenotypic cross-resistance to elvitegravir in HIV-infected patients failing on raltegravir-containing regimens. Antimicrob. Agents Chemother. 56, 2873–2878 (2012).
Doyle, T. et al. Integrase inhibitor (INI) genotypic resistance in treatment-naive and raltegravir-experienced patients infected with diverse HIV-1 clades. J. Antimicrob. Chemother. 70, 3080–3086 (2015).
De Francesco, M. A. et al. Prevalence of integrase strand transfer inhibitors resistance mutations in integrase strand transfer inhibitors-naive and -experienced HIV-1 infected patients: A single center experience. AIDS Res. Hum. Retrovir. 34, 570–574 (2018).
Alvarez, M. et al. Surveillance of transmitted drug resistance to integrase inhibitors in Spain: Implications for clinical practice. J. Antimicrob. Chemother. 74, 1693–1700 (2019).
Lepik, K. J. et al. Emergent drug resistance with integrase strand transfer inhibitor-based regimens. AIDS 31, 1425–1434 (2017).
Van Herck, K., Leuridan, E. & Van Damme, P. Schedules for hepatitis B vaccination of risk groups: Balancing immunogenicity and compliance. Sex. Transm. Infect. 83, 426–432 (2007).
Stasi, C. et al. Screening for hepatitis B virus and accelerated vaccination schedule in prison: A pilot multicenter study. Vaccine 37, 1412–1417 (2019).
Chang, J. J., Mohtashemi, N. & Bhattacharya, D. Significance and management of isolated hepatitis B core antibody (anti-HBc) in HIV and HCV: Strategies in the DAA era. Curr. HIV/AIDS Rep. 15, 172–181 (2018).
Wu, T., Kwok, R. M. & Tran, T. T. Isolated anti-HBc: The relevance of hepatitis B core antibody—A review of new issues. Am. J. Gastroenterol. 112, 1780–1788 (2017).
Stroffolini, T. et al. Hepatitis C virus infection among drug addicts in Italy. J. Med. Virol. 84, 1608–1612 (2012).
Gardini, I., Bartoli, M., Conforti, M., Mennini, F. S. & Marcellusi, A. Estimation of the number of HCV-positive patients in Italy. PLoS ONE 14, e0223668 (2019).
Kondili, L. A. et al. Estimated prevalence of undiagnosed HCV infected individuals in Italy: A mathematical model by route of transmission and fibrosis progression. Epidemics 34, 100442 (2021).
Schmidt, J., Blum, H. E. & Thimme, R. T-cell responses in hepatitis B and C virus infection: Similarities and differences. Emerg. Microbes Infect. 2, e15 (2013).
Giuliani, R. et al. HCV micro-elimination in two prisons in Milan, Italy: A model of care. J. Viral Hepat. 27, 1444–1454 (2020).
Rodríguez-Tajes, S. et al. Significant decrease in the prevalence of hepatitis C infection after the introduction of direct acting antivirals. J. Gastroenterol. Hepatol. 35, 1570–1578 (2020).
Fiore, V. et al. Chronic viral hepatitis in the Italian prison setting: Prevalence, outcomes, literature needs and perspectives. Healthcare 9, 1186 (2021).
Tresó, B. et al. Molecular epidemiology of hepatitis C virus genotypes and subtypes among injecting drug users in Hungary. Euro Surveill. 18, 20639 (2013).
Salemovic, D. et al. Intravenous drug use - an independent predictor for HCV genotypes 3 and 4 infection among HIV/HCV co-infected patients. Arch. Med. Sci. 13, 652–658 (2017).
Tyczyno, M., Halota, W., Nowak, W. & Pawlowska, M. Distribution of HCV genotypes in the populations of inmates in polish prison Potulice and patients hospitalised in Bydgoszcz. Hepat. Mon. 14, e14559 (2014).
Petruzziello, A. et al. Nine-year distribution pattern of hepatitis C virus (HCV) genotypes in Southern Italy. PLoS ONE 14, e0212033 (2019).
Bon, I. et al. Genotypic resistance in plasma and peripheral blood lymphocytes in a group of naive HIV1 patients. J. Clin. Virol. 38, 313–320 (2007).
Kwon, E. H. et al. HIV-1 subtypes and drug resistance mutations among female sex workers varied in different cities and regions of the Democratic Republic of Congo. PLoS ONE 15, e0228670 (2020).
Chew, C. B. et al. Assessment of drug resistance mutations in plasma and peripheral blood mononuclear cells at different plasma viral loads in patients receiving HAART. J. Clin. Virol. 33, 206–216 (2005).
Arias, A. et al. Sanger and next generation sequencing approaches to evaluate HIV-1 virus in blood compartments. Int. J. Environ. Res. Public Health 15, 1697 (2018).
Manyana, S. et al. HIV-1 drug resistance genotyping in resource limited settings: Current and future perspectives in sequencing technologies. Viruses 13(6), 1125 (2021).
Fokam, J. et al. Performance evaluation of an in house human immunodeficiency virus type-1 protease-reverse transcriptase genotyping assay in Cameroon. Arch. Virol. 156, 1235–1243 (2011).
Giovanetti, M. et al. Analysis of HIV-1 integrase genotypes and polymorphisms among integrase inhibitors-based antiretroviral treatment naïve patients in South Sudan. J. Med. Virol. 94, 3320–3327 (2022).
Pybus, O. G. et al. Genetic history of hepatitis C virus in East Asia. J. Virol. 83, 1071–1082 (2009).
Hall, T. A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 41, 95–98 (1999).
Nei, M. & Kumar, S. Molecular Evolution and Phylogenetics (Oxford University Press, 2000).
Bennett, D. E. et al. Drug resistance mutations for surveillance of transmitted HIV-1 drug-resistance: 2009 update. PLoS ONE 4, e4724 (2009).
Wensing, A. M. et al. 2019 update of the drug resistance mutations in HIV-1. Top. Antivir. Med. 27, 111–121 (2019).
Acknowledgements
We would like to thank Dr. F. Cammisa for support in study management and editorial assistance; Dr. N. Galea for discussion and English reviewing of the manuscript; S. De Menna, S. Tobelli and F. Fedeli for administrative support; F. Costa, P. Cocco and P. Arciero for technical support.
Funding
The work was funded by grants from the Gilead Fellowship Program Supporting Research to advance Patient Care (2016).
Author information
Authors and Affiliations
Contributions
M.T.M. designed the study, wrote the manuscript, provided supervision. L.S., participated in the experimental procedures and in the interpretation of results. O.P. analyzed the data and performed the statistical analysis. E.P performed phylogenetic and clustering HIV analyses. R. Be, D.F., C.R., C.F. performed HIV amplification and sequencing. AR.C. provided critical revision of the manuscript. R.Br. supervised HBV and HCV analyses and performed HCV phylogenetic analysis. A.C., S.T. performed HBV and HCV amplification and sequencing. P. C., E.M. performed HBV and HCV serological assays. A.B. provided critical advice on data. R.R., R. Ba., A.P., F.C., S. Ba., G.M., G.S., S.D.I., P.C. organized sample collection at the centres and sample shipment to CNAIDS and provided information on the patients. B.E. contributed in analysing the data and revising the manuscript. S.B. conceived the study and contributed to manuscript writing. All authors have read and approved the final manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Maggiorella, M.T., Sernicola, L., Picconi, O. et al. Epidemiological and molecular characterization of HBV and HCV infections in HIV-1-infected inmate population in Italy: a 2017–2019 multicenter cross-sectional study. Sci Rep 13, 14908 (2023). https://doi.org/10.1038/s41598-023-41814-x
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-023-41814-x
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
