
Abstract
Cats are hypercarnivorous, opportunistic animals that have adjusted to anthropogenic environments since the Neolithic period. Through humans, either by direct feeding and/or scavenging on food scraps, the diet of cats has been enriched with animals that they cannot kill themselves (e.g., large mammals, fish). Here, we conducted carbon and nitrogen stable isotope ratio analysis to reconstruct the diet of medieval cats and investigate cat-human interactions in two medieval harbor sites (Qalhât, Oman and Siraf, Iran). The analysis included 28 cat individuals and 100 associated marine and terrestrial faunal samples pertaining to > 30 taxa. The isotopic results indicate a high marine protein-based diet for the cats from Qalhât and a mixed marine-terrestrial (C4) diet for the cats from Siraf. Cats at these sites most likely scavenged on both human food scraps and refuse related to fishing activities, with differences in the two sites most likely associated with the availability of marine resources and/or the living conditions of the cats. By shedding light on the dietary habits of cats from two medieval harbors in the Arabian Gulf and Gulf of Oman, this study illustrates the potential of stable isotope analysis in reconstructing human-cat interactions in the past.
Similar content being viewed by others

Introduction
Cats are hypercarnivorous animals, requiring a threefold consumption of protein compared to omnivorous species1. Their high protein requirements stem from their metabolic adaptation to use protein and fat as energy sources2,3, as well as from their increased need for certain amino acids, such as taurine and arginine, which they cannot synthesize themselves4,5,6. With a preference for consuming multiple, small meals throughout the day7, cats prey on small mammals, birds, reptiles, amphibians and invertebrates8,9,10, with prey size decreasing with increasing hunger11. Predation occurs also as part of teaching or playing and, thus, not always results in the consumption of the prey12. However, both the predation rate and the prey diversity seem to be higher in rural areas, where anthropogenic food is scarcer13. Being opportunistic hunters with an ability to adjust rapidly to changing environments14, wild cats seem to have exploited the new hunting grounds that emerged from the development of permanent settlements, following the onset of agricultural activities15,16. This resulted in their adaptation to human presence, and later on to the consumption of foods facilitated by humans17. Either from direct feeding and/or from scavenging on human food scraps, cats have acquired access to animal taxa that they are not able to kill themselves, such as large mammals and fish18,19.
Today, the pet food industry incorporates a wide range of ingredients, including even foods of plant origin, such as grains and vegetables18,19. Unlike dogs20, a cat’s digestive system is not adapted to starch-rich foods; however, milled and cooked plant material can be metabolized7, and constitute part of a healthy diet, provided that this is nutritionally complete and balanced21. Fish is an important component of pet food, with cats consuming an estimated 6% of all wild caught fish22. In nature, however, both domestic and wild cats rarely capture fish. The bulk of evidence on their dietary preferences across four continents shows a very low predation rate (0.3%) of fish for wild, feral and domestic cats23,24. Although actual, active capturing of fish by cats seems limited, they feed on fish when access is facilitated23. Thus, fish is more often consumed by domestic rather than wild cats23. A recent isotopic study by Krajcarz et al.17, for example, revealed the consumption of marine protein by domestic cats populating medieval harbor towns in northern Europe.
Currently, information on the paleodiet of cats is scarce, possibly due to the fact that their bones are not often recovered in human food refuse or other anthropogenic contexts. The rarity of cat remains may also be a reason why its domestication history has been, so far, minimally investigated25,26,27,28. Cat remains are most often isotopically analyzed as single specimens, being part of local isotopic baselines (e.g., Refs.29,30,31,32,33). Recent studies, however, focused on the paleodietary reconstruction of domestic and wild cats by analyzing tens of individuals16,17. Krajcarz et al.16 suggested that the ancestor of the domestic cat, the Near Eastern wildcat Felis silvestris lybica, lived in Poland as a free-roaming cat, feeding on synanthropic pests, in close association with Early Neolithic farming communities. In another study, Krajcarz et al.17 revealed the influence of regional factors (i.e., socio-economic and geomorphological) on cat dietary patterns in medieval northern Europe.
Dietary reconstructions of ancient cats can, thus, provide information on regional food availabilities (i.e., how people adapted to local environments) and, ultimately on human-cat interactions. Aiming at reconstructing the dietary patterns of cats from two medieval harbors in the Arabian Gulf and the Gulf of Oman, in the present study we conducted carbon and nitrogen stable isotope ratio analysis on 47 cat samples and more than 200 associated faunal samples. By developing local baselines and by focusing on two sites with tens of cat remains as well as with historical and archaeological evidence for a long tradition in seafaring and sea fishing34,35,36,37,38,39,40,41, this study seeks to shed light on fish consumption by cats in the past and on medieval human-cat interactions in the Arabian Gulf and Gulf of Oman.
Materials and methods
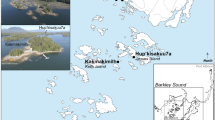
The cat remains and associated fauna analyzed in this study are from two medieval harbor towns, Siraf in Iran and Qalhât in Oman (Fig. 1). The faunal spectra of the two sites can be adequately compared since in both cases, the zooarchaeological analyses were carried out on large animal refuse accumulations that were sampled in the same way (i.e., by hand-collection). Table 1 presents the relative importance of the major food animals at Qalhât and Siraf, based on the zooarchaeological evidence.

Location of Siraf and Qalhât. Background relief map: released under Creative Commons, CC0 (https://maps-for-free.com), no alteration applied. Miniature globe: released under Creative Commons, CC BY-SA 3.0 (Globe_terrestre_Orange_te_Bleu.svg; Commons Wikimedia). The originally orange-colored continental landmasses and the blue sea surfaces (author: Mouh2jijel) have been converted to a black and white version, respectively, using Photoshop CS 5.5. North arrow, scale and text labels were created using Photoshop CS 5.5.
Qalhât, located near the easternmost tip of the Arabian Peninsula, was a large town occupied between the eleventh and sixteenth centuries CE. It was one of the major ports of the region between the thirteenth and fifteenth centuries, when it dominated the maritime trade in the western Indian Ocean and the Arabian Gulf. It was struck by an earthquake at the end of the fifteenth century, then plundered by the Portuguese in 1508 and finally completely abandoned in the second half of the sixteenth century42. Fieldwork since 2008 has revealed the layout of medieval Qalhât and has located the town’s major buildings. The cat remains included in the present study belong to 12 domestic cats (of which 3 were non-adult, weaned individuals, cf. Supplementary Table 1) and date between the fourteenth and sixteenth centuries CE. They were found in a cistern (B2), the Great Mosque (B12) and a large domestic building (B94). The remaining species come from the domestic building B94 (with the exception of a finless porpoise and a red fox, which were found in buildings B13 and B95, respectively) and include 54 fish belonging to 15 taxa (cf. Supplementary Table 3), 24 domestic animals (sheep, goat, chicken, dog and donkey), 5 wild birds (4 taxa), 9 wild terrestrial mammals (gazelle, red fox, hare and rat) and 3 delphinoids (cf. Supplementary Table 2). The cat remains from medieval Qalhât were previously described from a zooarchaeological43,44 and palaeopathological point of view45, while the remaining species are discussed elsewhere by Marrast (fish37) and Monchot (other vertebrates46).
Siraf is located in the Arabian Gulf and was an important harbor since Sassanid times (between 300 and 600 CE) and in particular between 800 and 1050 CE, when it was a transshipment port for goods coming from India, the Far East and Eastern Africa47. Siraf was a thriving, rich town until the end of the tenth century CE, when the area was struck by an earthquake. Afterwards, parts of the town were rebuilt and remained partly inhabited until the sixteenth century CE. An estimated 200,000 faunal remains were recovered from the excavations carried out between 1966 and 1973, of which about 10% was analyzed36. The studied material is from the area of the Great Mosque (site B) and consists mainly of remains collected from the shops that surrounded the mosque on three sides. About 80% of the faunal remains date to the most flourishing periods of the town, between 300 and 1050 CE, while the remaining 20% date to later periods (1050 until the early 16th c. CE). The material selected for the present study comes mainly from first millennium CE contexts (chronological units 1a and 1b36). This includes 35 domestic cats (of which 2 were non-adult, weaned individuals, cf. Supplementary Table 1), 51 fish belonging to 15 taxa (cf. Supplementary Table 3), 7 sea turtles, 45 domestic animals (sheep, goat, cattle, pig, chicken and dog) and 6 rats (cf. Supplementary Table 2).
Collagen was extracted at the Moesgaard Archaeo-Science Laboratory at Aarhus University (MOS; Denmark) and at the Royal Institute for Cultural Heritage (RICH; Belgium). More specifically, at the Moesgaard Archaeo-Science Laboratory, whole bone pieces were demineralized in 0.5M or 0.25M HCl (depending on preservation) at 4 °C, gelatinized in 0.01M HCl (pH adjusted to ~ 3) at 65 °C for ~ 48 h, purified using an EZEE filter, frozen at − 30 °C and freeze-dried for 48 h. Collagen from bone samples with poor quality indicators was re-extracted, when bone was available, and further purified using pre-cleaned 30 kDa Amicon Ultra Centrifugal Filters (indicated in Supplementary Table 1). In one instance (QALrat04), leftover collagen was ultra-filtered and re-analyzed. Collagen from fish bone samples with poor quality indicators was re-extracted, when bone was available, and treated with 0.1M NaOH prior to gelatinization (indicated in Supplementary Table 3), with the aim to remove humic substances48. Collagen extraction at the Royal Institute for Cultural Heritage is described in detail by Wojcieszak et al.49. Several studies (e.g., Refs.50,51,52,53) have demonstrated that stable isotopic values, and thus paleodietary interpretations, are not influenced by the use of different collagen extraction procedures.
The carbon and nitrogen stable isotope ratios of the samples were determined using a Thermo Flash HT/EA elemental analyzer linked to a Thermo Delta V Advantage isotope ratio mass spectrometer (IRMS) via a ConfloIV interface (Thermo Scientific) at the Department of Earth and Environmental Sciences of KU Leuven (Belgium). Data calibrations were done using one international (IAEA-600) and two in-house standards (Leucine and muscle tissue of Pacific Tuna) that were prior calibrated against certified standards. The international and in-house standards were measured at regular intervals throughout each analytical run, and their standard deviations defined the analytical error to be better than 0.11‰ for δ13C and 0.08‰ for δ15N. The isotopic results are presented as the ratio of the heavier to the lighter isotope (13C/12C, 15N/14N) and are reported relatively to internationally defined standards (VPDB, AIR) as δ values in units per mil (‰).
For estimating the relative contribution of potential food sources to the diet of the cats, we used the Bayesian mixing model FRUITS (Food Reconstruction Using Isotopic Transferred Signals; version 3.0)54. The input parameters used for the mixing model are reported in Supplementary Table S5. Wilcoxon signed rank test was performed using the software R (v. 4.1.3.)55,56 to statistically compare the isotope values of the cats from the two sites. A parametric test was conducted as the size of group samples was small (n < 20)57.
Results
Collagen preservation
All 12 cat samples from Qalhât and 10 out of 35 cats from Siraf yielded good quality collagen (i.e., > 13% for %C, > 4.8% for %N, 2.9–3.6 for C:N and ≥ 1% for collagen yield58,59,60). Another six cats from Siraf (cf. Supplementary Table 1) showed low collagen yields (< 1%), but acceptable %C, %N and C:N. Thus, their values were used for interpretation. Moreover, a total of 42 fish samples, 22 from Qalhât and 20 from Siraf, yielded collagen within the acceptable ranges for %C, %N and C:N, with 22 of them having a collagen yield < 1% (cf. Supplementary Table 3). From the remaining fauna, a total of 58 samples, 36 from Qalhât and 22 from Siraf, yielded acceptable collagen, with one sample from Siraf (SRrat02) having a low yield (cf. Supplementary Table 2). The isotopic values of all the faunal remains from the two sites, which are interpreted, are shown in Figs. 2 and 3.

δ13C and δ15N results of the cats and other faunal remains from Qalhât and Siraf. Supplementary Table S3 presents a list of the coastal and pelagic fish from the two sites.

δ13C and δ15N results of the different fish taxa from Qalhât and Siraf.
Isotopic results from Qalhât
The cats from Qalhât have a mean δ13C value of − 11.8‰ (± 0.9‰) and a mean δ15N value of 17.1‰ (± 0.7‰). These isotopic values are almost one trophic level above the values of the fish from this site61,62, indicating that cats had a diet highly dependent on marine protein. This is further supported by the results of the mixing model (Supplementary Table S6), which suggest that 41 ± 21% of the protein consumed by cats was marine. The fish that yielded acceptable isotopic results have a mean ± SD δ13C value of − 11.4 ± 1.1‰ and a mean ± SD δ15N value of 14.4 ± 1.2‰ (cf. Supplementary Table 3). Two marine mammals (finless porpoise and small delphinid) have δ13C values of − 11.7‰ and − 11.9‰ and δ15N values of 15.0‰ and 15.6‰, respectively, reflecting the small fish, crustaceans, cephalopods and/or other mollusks that these two taxa consume. One rat (QALrat01) and one dog (QALdog01) have values that are similar to the values of the cats (i.e., δ13C: − 13.6‰ and − 11.4‰, respectively, and δ15N: 17.5‰), indicating a significant contribution of marine protein in their diet as well. The remaining four rats have isotope values (δ13C mean ± SD: − 18.6 ± 1.3‰ and δ15N mean ± SD: 10.6 ± 1.7‰) that reveal different amounts of C3, C4 and marine protein in their diet, while the second dog (QALdog02) has one of the lowest δ15N values reported in this study (6.3‰), which could indicate a diet based primarily on C3 and C4 plants (δ13C: − 15.1‰)20,63,64,65.
With a mean ± SD δ13C value of − 15.0 ± 1.1‰ and a mean ± SD δ15N value of 16.2 ± 1.5‰, the isotope values of the eight chickens reveal differential amounts of marine protein in their diet. Two wild bird species have δ13C (Caspian gull: − 18.9‰ and − 22.1‰ and lappet-faced vulture: − 13.2‰) and δ15N (Caspian gull: 14.9‰ and 13.2‰ and lappet-faced vulture: 12.5‰) values that reflect their scavenging nature and the consumption of both terrestrial and marine protein. One lesser white-fronted goose has a δ13C value of − 18.7‰ and a δ15N value of 15.3‰, which perhaps reflect the consumption of coastal plants (including halophytes66) that were affected by sea-sprays and soil salinity67,68, while one cormorant with a δ13C value of − 18.3‰ and a δ15N value of 12.2‰ likely fed on brackish or a mix of freshwater fish and amphibians as well as marine fish. The latter species may have foraged in Khors (i.e., the typical estuarine habitats found along the Arabian Peninsula’s coast)69,70. From the wild terrestrial mammals, two gazelle specimens with similar δ15N values (7.1‰ and 7.9‰) and a 6.8‰ difference in δ13C values (− 19.4‰ and − 12.6‰) reflect the consumption of plants that follow different photosynthetic pathways (i.e., C3 and C4, respectively), while one red fox has a δ13C value of − 13.8‰ and a δ15N value of 10.1‰, which indicate the consumption of small, wild mammals that were highly dependent on C4 plants. From the domestic mammals, sheep and goats have similar isotope values that range between − 10.7‰ and − 16.3‰ for carbon and between 5.8‰ and 10.7‰ for nitrogen, reflecting mainly the consumption of C4 plants, partially at coastal sites71. Apart from a high variety of wild C4 plants in the area66, archaeobotanical evidence from the region reveals the presence of cultivated C4 plants, such as millet72,73 and sorghum74, while historical sources attest that cultivated plants were intended for both human and animal consumption75. One sheep has a δ13C value of − 12.6‰ and a δ15N value of 15.3‰, reflecting the consumption of marine fish, which, based on medieval written testimonies, were used as fodder in the area during the sampled period35. Finally, two donkeys have a δ13C value difference of 6.3‰ (− 19.9‰ and − 13.6‰) and a δ15N value difference of 2.3‰ (8.0‰ and 10.3‰) that reflect a dependence on C3 and C4 plants, respectively, the latter perhaps from coastal environments.
Isotopic results from Siraf
Cats from Siraf have a mean ± SD δ13C value of − 12.1 ± 0.9‰ and a mean ± SD δ15N value of 12.3 ± 0.5‰, suggesting the consumption of protein from C4 sources and perhaps some contribution of marine protein. The lesser contribution of marine protein to the diet of these cats is also supported by the results of the mixing model (Supplementary Table S6), which suggest that 34 ± 19% of the protein consumed by the cats was derived from fish and 37 ± 21% from herbivorous mammals. The 20 fish specimens that yielded acceptable isotopic results have a mean ± SD δ13C value of − 9.8 ± 4.0‰ and a mean ± SD δ15N value of 12.0 ± 2.2‰ (cf. Supplementary Table 3). From the terrestrial animals, three dogs (δ13C mean ± SD: − 18.2 ± 0.9‰, δ15N mean ± SD: 11.0 ± 0.3‰), five chickens (δ13C mean ± SD: − 18.0 ± 1.0‰, δ15N mean ± SD: 11.5 ± 0.8‰) and two pigs (δ13C: − 18.3‰ and − 17.2‰, δ15N: 11.8‰ and 12.3‰) have similar isotope values that indicate a primarily C3-based diet with some contribution of protein from C4 and/or marine sources. Two rats have δ13C values of − 14.7‰ and − 16.5‰ and δ15N values of 14.0‰ and 12.1‰, respectively, indicating differential access to marine protein. For the domestic mammals, sheep and goats have similar isotope values that range between − 19.0‰ and − 20.0‰ for carbon and between 5.5 and 8.4‰ for nitrogen, reflecting the consumption of C3 plants. The five cattle samples have a relatively narrow δ15N value range of 2.4‰ (from 7.2 to 9.6‰), but a much larger δ13C value range of 11.4‰ (from − 18.5 to − 7.1‰), which indicates differential access to C4 plants.
Discussion
Cat diets in Qalhât and Siraf
Due to their peculiar ecological and behavioral features, cats can be both autonomous predators as well as dependent on food provisioning by humans23,76. Hunting of wild prey is more significant in rural areas, where anthropogenic food is scarcer13. Cats that are independent on food availability by humans and their activities, have been shown to exhibit low δ13C and δ15N values and narrow value ranges, which reflect a specialized trophic niche that is based on small wild animals17,23. Cats from urban contexts, however, are expected to exhibit a wide δ13C value range that reflects access to a variety of food sources through humans, as well as high δ15N values, especially if marine resources are available17.
The cats included in the present study come from urban contexts. In Siraf, cat remains were discovered in the commercial heart of the town, the area of the Great Mosque, which was surrounded by shops36,47, while in Qalhât, they were found in various buildings across the medieval town43. Thus, it is expected that, either by direct feeding and/or scavenging on human food scraps, the Siraf and Qalhât cats consumed foods that they would not be able to access otherwise, for example, meat from large mammals, such as sheep and cattle, as well as fish.
In Qalhât, cats have a 0.4‰ lower mean δ13C value, but a 2.8‰ higher mean δ15N value than the marine fish, which indicates that marine protein contributed significantly to the diet of the cats from this site. The fact that cats have a similar mean δ13C value to the finless porpoise and the small delphinid, but a 1.5–2.0‰ higher mean δ15N value, supports this interpretation and further indicates that cats consumed protein of higher trophic level than these two marine mammals, which commonly feed on small fish, crustaceans, cephalopods and other molluscs. Moreover, although cats and red foxes target the same prey groups77, the red fox from Qalhât has a 7.0‰ lower δ15N value than the mean δ15N value of the cats, reflecting the consumption of small prey that was of lesser dietary significance for the domestic cats. Seabirds as well as rats feeding on both fish and seabirds could have also been part of the diet of the Qalhât cats78,79,80,81. According to our FRUITS model, rats and birds (including seabirds) may have constituted, respectively, ~ 16% and ~ 21% of the dietary protein consumed by these cats.
Although the mean δ13C value of the Siraf cats is only 0.3‰ lower than the mean δ13C value of the cats from Qalhât, the 4.8‰ mean δ15N value difference between the cats from the two sites, suggests the consumption of lower trophic level foods by the Siraf cats. This mean value difference in nitrogen is statistically significant (p < 0.001). Having a 3.1‰ lower mean δ13C value and a 0.6‰ higher mean δ15N value compared to the analyzed fish, cats from Siraf were less dependent on marine protein than the cats from Qalhât. This is further supported by the results of the mixing model (Supplementary Table S6), which suggest that ~ 41% of the protein consumed by the cats from Qalhât derived from marine resources, compared to only ~ 34% for the cats from Siraf, with herbivorous mammals comprising their highest dietary contribution (~ 37%). Therefore, the high mean δ13C value of the cats from Siraf most likely indicates their dietary dependence on animals who fed on C4 plants, with a small contribution of protein from marine sources for those with higher δ15N values.
Different cat diets and local baselines
Although both Siraf and Qalhât are coastal sites (Fig. 1) and were major harbors during the investigated periods42,47, cats from the two sites seem to have had different diets. For urban cats, anthropogenic food comprises an important part of their diet13. Hence, the different diets observed in the cats from the two sites may be related to the composition of waste resulting from human food preparation and consumption on the one hand and its accessibility and corresponding feeding opportunities offered in the cats’ respective home ranges on the other. Although it was not possible to analyze human bone samples from either site, archaeological and historical evidence provide information on the availability of food sources in Siraf and Qalhât during the studied periods.
Historical information and archaeobotanical evidence indicate that fruit (especially dates), cereals (including wheat, rice, sorghum and millet) and vegetables (e.g., garlic and eggplant) were either cultivated and/or imported in pre-Islamic and Islamic Oman72,74,75,82, including medieval Qalhât73,83. The three rat specimens with lower δ15N values that were primarily feeding on crops, corroborate the presence of such plants at the site and likely their importance in human diets. Archaeological evidence of plant remains, such as dates84, grapes85, millet and wheat86,87, is also present in prehistoric Iran. Crops, such as wheat and barley, were paid as taxes during the medieval period88, when water mills close to Siraf were likely used for grinding cereals47. According to the zooarchaeological record, subsistence in Qalhât and Siraf was based on marine fish and domestic animals (Table 1). Hunting of wild mammals and birds was rarely practiced and occasionally larger marine vertebrates (turtles and delphinoids) were captured, probably as a bycatch of fishing activities. The proportion of the domestic animal bones is higher in Siraf, suggesting that fish contributed less to the human diet in this site compared to Qalhât. Historical sources nevertheless support a high consumption of fish both in Oman35 as well as in the coast of the Arabian Gulf34. In Oman, both small fish, such as sardines and anchovies, as well as larger fish, such as sharks and sailfish, were consumed likely after being sun-dried or salted35. Thick layers of fish bones discovered in the domestic building B94, where some of the cat remains analyzed in the present study were also found, may indicate the use of the building for fish processing activities89. Such activities, however, may have also taken place on the beach, providing subsistence opportunities to animals, such as seabirds, rats and cats35.
In Qalhât, pelagic fishing seems to have been more important than coastal, inshore fishing (Table 1). The 18 pelagic and four coastal fish from this site have a lower mean δ13C value (− 11.3 ± 1.1‰ and − 12.1 ± 1.1‰, respectively) compared to the four pelagic (− 9.6 ± 0.9‰) and 16 coastal (− 8.9 ± 1.0‰) fish from Siraf (Fig. 2, Supplementary Table S3). In the Arabian Gulf, high temperatures and high evaporation rates lead to hypersaline conditions90,91, which result in the underdevelopment of the mangrove tree populations92,93. A benthic trophic pathway in microphytobenthos sites, which are 13C-enriched compared to mangrove sites94,95, may, thus, account for the higher δ13C values of the fish from Siraf. Moreover, fish from Qalhât, both coastal and pelagic (Fig. 2), have higher δ15N values compared to fish from Siraf (Fig. 3), which indicates perhaps a longer trophic chain in the Oman Gulf (although the δ15N value differences between fish from the two Gulfs could be also attributed to baseline δ15N variations96). The high trophic level of the fish available in Qalhât is seen in the high δ15N values of the cats that probably consumed fish, such as longtail tuna and requiem shark, which are the most represented fish categories from the site37, and/or seabirds and rats that fed on fish78,80,81.
The zooarchaeological evidence from both sites indicates also that among the domestic species, sheep and goats were the most important, with a representation in the skeletal assemblage of > 90% (Table 1). In Qalhât, the wide value ranges (δ13C: 1.7‰ and δ15N: 2.9‰) for sheep and goats suggest a diet with various amounts of C4 protein (perhaps partially at coastal areas and salty grounds) as well as potentially a dietary contribution of marine protein (especially for one sheep with a δ15N value of > 15.0‰). Similar isotopic signatures were observed in the ovicaprines from the Islamic site of Qalʿat al-Baḥrayn in Bahrain97. The practice of using small fish, especially sardines, as fodder has been reported for Oman by late thirteenth and fourteenth century CE sources34,35. Although slightly more present in Siraf (3.0%) than in Qalhât (0.6%), cattle were rare, probably due to the animal’s size that requires larger quantities of water to be sustained compared to sheep and goats98,99. Cattle are also less tolerant to drinking saline water100,101. Coastal Iran and Oman have an arid climate, with low annual precipitation, leading to freshwater shortages and increased salinity, while the majority of the freshwater used originates from groundwater resources102,103. Of the five cattle from Siraf, three have isotope values that indicate a high dependence on C4 plants. There is a high diversity of wild C4 plants in the region104,105,106, while archaeobotanical evidence of millet proves that it has been cultivated in the area for centuries before the studied period86,87. Although, at first glance, cattle could not have contributed significantly to the diet of the cats given their low overall representation to the faunal assemblage, it was noted that the Siraf cat remains have been collected in the debris deposited near the Great Mosque, more precisely in the commercial heart of the urban center characterized by shops, where butchers likely practiced their craft as well. Cattle butchers in particular not only need large markets with good sales opportunities, but also produce a lot of offal per capita. Therefore, the comparatively large number of cats in the excavation area in question as well as their isotopic signatures could possibly relate to the presence of such a guild. Additionally, considering that the site has been partially excavated47 and only 10% of the recovered faunal remains has been analyzed36, it is likely that large accumulations of cattle bones associated with butchery activities, such as those reported for early Byzantine Sagalassos in Turkey107, were not discovered. An alternative source of C4 protein for the cats from Siraf could have been small prey that fed on C4 plants. Yet, the two rats analyzed have lower δ13C and similar or higher δ15N values than the cats, while the consumption of mainly wild prey would likely result in lower δ15N values, similarly to the red fox from Qalhât. This brings us back to the previous point that economic activities in the city center opened up opportunities for Siraf cats to get hold of food with a high C4 content.
The zooarchaeological evidence from Siraf seems to suggest a lesser dependence on marine protein by humans in the area compared to Qalhât. This is further supported by the isotope values of animals such as dogs and chickens, which have previously been considered as proxies for human diets97,108,109 (however, see also Refs.110,111) and which have mean δ13C values indicative of a terrestrial diet. The higher δ13C values of the cats compared to these two animals (mean value difference of about 6.0‰) suggests that, although the marine dietary signal in the cats from Siraf is not as clear as for the cats from Qalhât, cats from both sites seem to represent a stronger proxy for the marine resources available in the two regions. This has also been suggested for the Islamic site of Qalʿat al-Baḥrayn, where isotopic analysis of both humans and fauna revealed that despite the large numbers of fish bones at the site, humans and chicken (likely consuming human food scraps) have isotope values that indicate a primarily terrestrial diet, while one cat has a higher δ13C value that indicates a higher dietary dependence on marine protein97.
A higher dependence on marine protein for cats, compared to animals that were perhaps mainly feeding on the waste of human food preparation and consumption, likely indicates that these felids were roaming freely, scavenging both on human food scraps as well as on refuse related to on-site processing of fish. The clearer isotopic signal of a marine diet for the cats from Qalhât probably relates to more extended fishing activities in this site, which further included large pelagic species. Finally, cats (and animals in general) are viewed favorably in Islamic culture112,113, however, the high mortality of juvenile cats in Qalhât along with the presence of skeletal pathologies (including traumatic lesions) observed in at least two cats43,45, may be partially indicative of occasional human violence towards cats. An overall lower human tolerance of cats in this town could have restricted them to the harbor, where large fish were probably gutted prior to further processing. Conversely, the cats that roamed the city center of Siraf may have frequented mainly the immediate surroundings of the market place with its shops, where the processing and sale of fish may have been one activity among many.
Cat diets at seaport sites
A marine dietary signal has also been reported for cats from medieval port towns in Europe. In a recent study, Krajcarz et al.17 analysed cat remains from various sites, both coastal and inland, in Poland, Germany and Belgium and reported higher δ13C and δ15N values for the cats from the coastal sites. Krajcarz et al.17 further reported δ13C and δ15N differences between cats from port sites located in the Baltic Sea region (Puck, Gdańsk and Kołobrzeg in Poland) and those found in the North Sea region (Bremen in Germany and Nieuwpoort in Belgium), analogous to isotopic value differences reported for fish by Barrett et al.114, i.e., both δ13C and δ15N values being lower for the Baltic Sea, where there is a larger freshwater input. Although cats from Qalhât and Siraf have higher δ13C and δ15N values than the Polish, German and Belgian cats reported by Krajcarz et al.17 (Fig. 4, Supplementary Table S4), these differences are most likely related to local isotopic baselines. What remains evident, however, from both studies, is that cats are opportunistic animals that adjust their diets based on the availability of local resources.

δ13C and δ15N values of cats from the medieval harbor sites of Qalhât (Oman) and Siraf (Iran) (present study) as well as from Bremen (Germany), Nieuwpoort (Belgium) and Puck, Gdańsk and Kołobrzeg (Poland)17.
Conclusions
By establishing local isotopic baselines, this study generated new evidence on the diet of cats from the medieval harbor sites of Siraf in Iran and Qalhât in Oman. The δ13C and δ15N results suggest different dietary habits, with the cats from Qalhât being highly dependent on fish and animals feeding on fish, and the cats from Siraf having a mixed marine-terrestrial (C4) diet. This C4 signal is potentially related to the consumption of cattle meat and offal, however, additional isotopic analyses (i.e., hydrogen stable isotope analysis115 and compound-specific stable isotope analysis of amino acids116,117) may help to provide a clearer dietary reconstruction in the future.
With an augmented interest in keeping cats as pets118 and research showing the benefits for mental health in living with companion animals (e.g., Refs.119,120), it becomes increasingly important to learn more about the interactions between humans and cats in space and time. By reconstructing the diet of domestic cats from two harbors in the Arabian Gulf and the Gulf of Oman, this study revealed that these felids were most likely roaming freely, scavenging on both the waste of human food consumption as well as refuse related to fishing activities, and, thus, representing an enhanced proxy for the marine dietary protein component of the human diet. The dietary differences between cats from Siraf and Qalhât indicate either differences in the availability of marine resources and/or distinct living conditions, induced by the specific feeding opportunities offered in the anthropogenic space of the two sites. A dietary reconstruction of domestic cats from various sites and periods could shed more light on how humans and cats interacted, thereby creating both unique bonds and mutual dependencies121,122 that resulted in cats being one of the most popular pets in the world today118.
Data availability
All data generated or analyzed during this study are included in this published article (and its Supplementary Information file).
References
Morris, J. G. Idiosyncratic nutrient requirements of cats appear to be diet-induced evolutionary adaptations. Nutr. Res. Rev. 15, 153–168 (2002).
Zoran, D. L. The carnivore connection to nutrition in cats. J. Am. Vet. Med. Assoc. 221, 1559–1567 (2002).
Eisert, R. Hypercarnivory and the brain: Protein requirements of cats reconsidered. J. Comp. Physiol. B 181, 1–17 (2011).
Morris, J. G. & Rogers, Q. R. Arginine: An essential amino acid for the cat. J. Nutr. 108, 1944–1953 (1979).
Knopf, K., Sturman, J. A., Armstrong, M. & Hayes, K. C. Taurine: An essential nutrient for the cat. J. Nutr. 108, 773–778 (1978).
Zaghini, G. & Biagi, G. Nutritional peculiarities and diet palatability in the cat. Vet. Res. Commun. 29, 39–44 (2005).
Spotte, S. Free-Ranging Cats: Behavior, Ecology, and Management (Wiley, 2014).
Castelló, J. R. Felids and Hyenas of the World. Wildcats, Panthers, Lynx, Pumas, Ocelots, Caracals, and Relatives (Princeton University Press, 2020).
Loss, S. R., Will, T. & Marra, P. P. The impact of free-ranging domestic cats on wildlife of the United States. Nat. Commun. 4, 1396 (2013).
Woinarski, J. C. Z. et al. How many birds are killed by cats in Australia?. Biol. Conserv. 214, 76–87 (2017).
Biben, M. Predation and predatory play behaviour of domestic cats. Anim. Behav. 27, 81–94 (1979).
Loyd, K. A. T., Hernandez, S. M., Carroll, J. P., Abernathy, K. J. & Marshall, G. J. Quantifying free-roaming domestic cat predation using animal-borne video cameras. Biol. Conserv. 160, 183–189 (2013).
Piontek, A. M. et al. Analysis of cat diet across an urbanisation gradient. Urban Ecosyst. 24, 59–69 (2021).
Natoli, E., Litchfield, C. & Pontier, D. Coexistence between humans and ‘misunderstood’ domestic cats in the anthropocene: Exploring behavioural plasticity as a gatekeeper of evolution. Animals 12, 1717 (2022).
Driscoll, C. A. et al. The Near Eastern origin of cat domestication. Science 317, 519–523 (2007).
Krajcarz, M. et al. Ancestors of domestic cats in Neolithic Central Europe: Isotopic evidence of a synanthropic diet. Proc. Natl. Acad. Sci. 117, 17710–17719 (2020).
Krajcarz, M. et al. Stable isotopes unveil one millennium of domestic cat paleoecology in Europe. Sci. Rep. 12, 12775 (2022).
Alexander, P., Berri, A., Moran, D., Reay, D. & Rounsevell, M. D. A. The global environmental paw print of pet food. Glob. Environ. Change 65, 102153 (2020).
McDonald, B. W. et al. High variability within pet foods prevents the identification of native species in pet cats’ diets using isotopic evaluation. PeerJ 8, e8337 (2020).
Axelsson, E. et al. The genomic signature of dog domestication reveals adaptation to a starch-rich diet. Nature 495, 360–364 (2013).
Knight, A. & Leitsberger, M. Vegetarian versus meat-based diets for companion animals. Animals 6, 57 (2016).
De Silva, S. S. & Turchini, G. M. Towards understanding the impacts of the pet food industry on world fish and seafood supplies. J. Agric. Environ. Ethics 21, 459–467 (2008).
Széles, G. L., Purger, J. J., Molnár, T. & Lanszki, J. Comparative analysis of the diet of feral and house cats and wildcat in Europe. Mammal Res. 63, 43–53 (2018).
Plantinga, E. A., Bosch, G. & Hendriks, W. H. Estimation of the dietary nutrient profile of free-roaming feral cats: Possible implications for nutrition of domestic cats. Br. J. Nutr. 106, S35–S48 (2011).
Baca, M. et al. Human-mediated dispersal of cats in the Neolithic Central Europe. Heredity 121, 557–563 (2018).
Driscoll, C. A., Macdonald, D. W. & O’Brien, S. J. From wild animals to domestic pets, an evolutionary view of domestication. Proc. Natl. Acad. Sci. 106, 9971–9978 (2009).
Ottoni, C. et al. The palaeogenetics of cat dispersal in the ancient world. Nat. Ecol. Evol. 1, 0139 (2017).
Ottoni, C. & Van Neer, W. The dispersal of the domestic cat: Paleogenetic and zooarcheological evidence. East. Archaeol. 83, 38–45 (2020).
Brozou, A. et al. Leprosy in medieval Denmark: Exploring life histories through a multi-tissue and multi-isotopic approach. Am. J. Phys. Anthropol. 176, 36–53 (2021).
Linderholm, A. & Kjellström, A. Stable isotope analysis of a medieval skeletal sample indicative of systemic disease from Sigtuna Sweden. J. Archaeol. Sci. 38, 925–933 (2011).
Polet, C. & Katzenberg, M. A. Reconstruction of the diet in a mediaeval monastic community from the coast of Belgium. J. Archaeol. Sci. 30, 525–533 (2003).
van der Sluis, L. G., Hollund, H. I., Kars, H., Sandvik, P. U. & Denham, S. D. A palaeodietary investigation of a multi-period churchyard in Stavanger, Norway, using stable isotope analysis (C, N, H, S) on bone collagen. J. Archaeol. Sci. Rep. 9, 120–133 (2016).
Webb, E. C. et al. Compound-specific amino acid isotopic proxies for distinguishing between terrestrial and aquatic resource consumption. Archaeol. Anthropol. Sci. 10, 1–18 (2018).
Zazzo, A., Munoz, O. & Saliége, J.-F. Diet and mobility in a Late Neolithic population of coastal Oman inferred from radiocarbon dating and stable isotope analysis. Am. J. Phys. Anthropol. 153, 353–364 (2014).
ElMahi, A. T. Traditional fish preservation in Oman: The seasonality of a subsistence strategy. In Proceedings of the Seminar for Arabian Studies vol. 30 99–113 (Archaeopress, 2000).
von den Driesch, A. & Dockner, A. Animal exploitation in medieval Siraf, Iran, based on the faunal remains from the excavations at the Great Mosque (seasons 1966–1973). Bonn. Zool. Beitr. 50, 227–247 (2002).
Marrast, A. Qalhat: Preliminary results of the B94 fish bones study. In Qalhat, a Medieval Port City of Oman. From a Field of Ruins to UNESCO (ed. Rougeulle, A.) (Archaeopress) (in press).
Ghidoni, A. & Vosmer, T. Boats and ships of the Arabian Gulf and the Sea of Oman within an archaeological, historical and ethnographic context. In The Arabian Seas: Biodiversity, Environmental Challenges and Conservation Measures (ed. Jawad, L. A.) 957–989 (Springer, 2021).
Carter, R. The Neolithic origins of seafaring in the Arabian Gulf. Archaeol. Int. 6, 44–47 (2002).
Biagi, G. The prehistoric fishermen settlements of RH5 and RH6 at qurum, sultanate of Oman. Proc. Semin. Arab. Stud. 17, 15–19 (1987).
Agius, D. A. Omani seafaring identity before the early 1600s: Ethnic and linguistic diversity. In Oman and Overseas (Studies on Ibadism and Oman) (eds. Hoffmann-Ruf, M. & Al Salimi, A.) (Georg Olms Verlag, 2013).
Rougeulle, A. The Qalhāt Project: New research at the medieval harbour site of Qalhāt, Oman (2008). Proc. Semin. Arab. Stud. 40, 303–319 (2010).
Monchot, H. Stray cats: Pets or pests? Cats and kittens everywhere in the medieval harbour site of Qalhāt (Oman). Arab. Arcliaeol. Epigr. https://doi.org/10.1111/aae.12204 (2022).
Monchot, H. & Béarez, P. Des ossements dans les citernes. Les exemples de Dharih (Jordanie) et de Qalhāt (Oman). Syr. Archéologie Art Hist. 93, 339–352 (2016).
Monchot, H. & Guintard, C. Chronic diseases in cats from the medieval site of Qalhât (Oman). Int. J. Paleopathol. 17, 1–9 (2017).
Monchot, H. The mammal and bird remains of the building B94. In Qalhat, a Medieval Port City of Oman. From a Field of Ruins to UNESCO (ed. Rougeulle, A.) (Archaeopress) (in press).
Whitehouse, D., Whitcomb, D. S. & Wilkinson, T. J. Siraf. History, Topography and Environment (The British Institute of Persian Studies, 2009).
UCI AMS Facility. Acid Base/Acid (ABA) Sample pre-treatment. (2011).
Wojcieszak, M., Van den Brande, T., Ligovich, G. & Boudin, M. Pretreatment protocols performed at the Royal Institute for Cultural Heritage (RICH) prior to AMS 14C measurements. Radiocarbon 62, e14–e24 (2020).
Sealy, J., Johnson, M., Richards, M. & Nehlich, O. Comparison of two methods of extracting bone collagen for stable carbon and nitrogen isotope analysis: Comparing whole bone demineralization with gelatinization and ultrafiltration. J. Archaeol. Sci. 47, 64–69 (2014).
Rand, A. et al. Comparison of three bone collagen extraction procedures: The effect of preservation on δ13C and δ15N values. N. Atl. Archaeol. 4, 93–113 (2015).
Cersoy, S., Zazzo, A., Lebon, M., Rofes, J. & Zirah, S. Collagen extraction and stable isotope analysis of small vertebrate bones: A comparative approach. Radiocarbon 59, 679–694 (2017).
Talamo, S., Fewlass, H., Maria, R. & Jaouen, K. “Here we go again”: The inspection of collagen extraction protocols for 14C dating and palaeodietary analysis. Sci. Technol. Archaeol. Res. 7, 62–77 (2021).
Fernandes, R., Millard, A. R., Brabec, M., Nadeau, M.-J. & Grootes, P. Food reconstruction using isotopic transferred signals (FRUITS): A Bayesian Model for diet reconstruction. PLoS ONE 9, e87436 (2014).
RStudio Team. RStudio: Integrated Development Environment for R. RStudio, PBC, Boston (2020).
R Core Team. R: A Language and Environment for Statistical Computing (R Foundation for Statistical Computing, 2021).
Madrigal, L. Statistics for Anthropology (Cambridge University Press, 2012).
Ambrose, S. H. Preparation and characterization of bone and tooth collagen for isotopic analysis. J. Archaeol. Sci. 17, 431–451 (1990).
DeNiro, M. J. Postmortem preservation and alteration of in vivo bone collagen isotope ratios in relation to palaeodietary reconstruction. Nature 317, 806–809 (1985).
van Klinken, G. J. Bone collagen quality indicators for palaeodietary and radiocarbon measurements. J. Archaeol. Sci. 26, 687–695 (1999).
Malainey, M. E. A Consumer’s Guide to Archaeological Science: Analytical Techniques. (2011).
Bocherens, H. & Drucker, D. Trophic level isotopic enrichment of carbon and nitrogen in bone collagen: Case studies from recent and ancient terrestrial ecosystems. Int. J. Osteoarchaeol. 13, 46–53 (2003).
Albizuri, S. et al. Dogs that ate plants: Changes in the canine diet during the late bronze age and the first iron age in the Northeast Iberian Peninsula. J. World Prehistory 34, 75–119 (2021).
Albizuri, S. et al. Dogs in funerary contexts during the Middle Neolithic in the northeastern Iberian Peninsula (5th–early 4th millennium BCE). J. Archaeol. Sci. Rep. 24, 198–207 (2019).
Grandal-d’Anglade, A. et al. Dogs and foxes in Early-Middle Bronze Age funerary structures in the northeast of the Iberian Peninsula: Human control of canid diet at the sites of Can Roqueta (Barcelona) and Minferri (Lleida). Archaeol. Anthropol. Sci. 11, 3949–3978 (2019).
Patzelt, A. Synopsis of the flora and vegetation of Oman, with special emphasis on patterns of plant endemism. Abh. Braunschw. Wiss. Ges. 282–317 (2015).
Heaton, T. H. E. The 15N/14N ratios of plants in South Africa and Namibia: Relationship to climate and coastal/saline environments. Oecologia 74, 236–246 (1987).
Virginia, R. A. & Delwiche, C. C. Natural 15N abundance of presumed N2-fixing and non-N2-fixing plants from selected ecosystems. Oecologia 54, 317–325 (1982).
von den Driesch, A. & Obermaier, H. The Hunt for Wild Dromedaries during the 3rd and 2nd Millennia BCE on the United Arab Emirates Coast: Camel Bone Finds from the Excavation at Al Sufouh 2, Dubai, UAE. In Skeletal Series and Their Socio-Economic Context (eds. Grupe, G. & Peters, J.) 133–167 (M. Leidorf, Rahden/Westf., 2007).
Heide, M. & Peters, J. Camels in the Biblical World. History, Archaeology, and Culture of the Levant (Eisenbrauns, Penn State University Press, 2021).
Ghazanfar, S. A. Coastal vegetation of Oman. Estuar. Coast. Shelf Sci. 49, 21–27 (1999).
Bellini, C., Pavan, A., Pignotti, L., Gonnelli, T. & Lippi, M. M. Food plants in pollen records from ancient Southern Arabia: The evidences from Sumhuram (Southern Oman). J. Arid Environ. 177, 104131 (2020).
Dabrowski, V., Tengberg, M., Creissen, T. & Rougeulle, A. Plant supplying strategies in an Islamic Omani harbour city: Archaeobotanical Analysis from a Workshop (B39) in Qalhat (XIVth-XVIth c. AD). J. Islam. Archaeol. (2018) https://doi.org/10.1558/jia.37690.
Dabrowski, V., Bouchaud, C., Tengberg, M., Zazzo, A. & Priestman, S. Archaeobotanical analysis of food and fuel procurement from Fulayj fort (Oman, 5th–8th c. CE) including the earliest secure evidence for sorghum in Eastern Arabia. J. Arid Environ. 190, 104512 (2021).
Ubaydli, A. The Agrarian economy of Oman (132–280/749–893) in Arabic sources. J. Islam. Stud. 4, 33–51 (1993).
Crowley, S. L., Cecchetti, M. & McDonald, R. A. Our wild companions: Domestic cats in the Anthropocene. Trends Ecol. Evol. 35, 477–483 (2020).
Castañeda, I., Zarzoso-Lacoste, D. & Bonnaud, E. Feeding behaviour of red fox and domestic cat populations in suburban areas in the south of Paris. Urban Ecosyst. 23, 731–743 (2020).
Bicknell, A. W. J. et al. Stable isotopes reveal the importance of seabirds and marine foods in the diet of St Kilda field mice. Sci. Rep. 10, 6088 (2020).
Guiry, E. & Buckley, M. Urban rats have less variable, higher protein diets. Proc. R. Soc. B 285, 20181441 (2018).
Hobson, K. A., Piatt, J. F. & Pitocchelli, J. Using stable isotopes to determine seabird trophic relationships. J. Anim. Ecol. 63, 786–798 (1994).
Stapp, P. Stable isotopes reveal evidence of predation by ship rats on seabirds on the Shiant Islands, Scotland. J. Appl. Ecol. 39, 831–840 (2002).
Hammer, K., Gebauer, J., Khanjari, S. A. & Buerkert, A. Oman at the cross-roads of inter-regional exchange of cultivated plants. Genet. Resour. Crop Evol. 56, 547–560 (2009).
Dabrowski, V., Ros, J., Tengberg, M. & Rougeulle, A. De l´origine et de l´utilisation des ressources végétales en Oman Medieval: Premiere étude archeobotanique à Qalhat. Assoc. Routes L´Orient 2, 1–13 (2014).
Tengberg, M. Beginnings and early history of date palm garden cultivation in the Middle East. J. Arid Environ. 86, 139–147 (2012).
Milanesi, C. et al. Geometric morphometry and archaeobotany: Characterisation of grape seeds (Vitis vinifera L.) by analysis of form. Open Access Libr. J. 1, e634 (2014).
Scott, A. et al. Exotic foods reveal contact between South Asia and the Near East during the second millennium BCE. Proc. Natl. Acad. Sci. 118, e2014956117 (2021).
Nesbitt, M. & Summers, G. D. Some recent discoveries of millet (Panicum Miliaceum L. and Setaria Italica (L.) P. Beauv) at excavations in Turkey and Iran. Anatol. Stud. 38, 85–97 (1988).
Choksy, J. K. Muslims and Zoroastrians in Medieval Iran and Western Inner Asia: Cultural Transition and Religious History (Harward University, 1991).
Rougeulle, A. Medieval Qalhāt, historical vs archaeological data. Arab. Humanit. https://doi.org/10.4000/cy.3442 (2017).
Ibrahim, H. D. Investigation of the Impact of Desalination on the Salinity of the Persian Gulf (Massachusetts Institute of Technology, 2017).
Smith, R., Purnama, A. & Al-Barwani, H. H. Sensitivity of hypersaline Arabian Gulf to seawater desalination plants. Appl. Math. Model. 31, 2347–2354 (2007).
Ball, M. C. Interactive effects of salinity and irradiance on growth: Implications for mangrove forest structure along salinity gradients. Trees 16, 126–139 (2002).
Wilson, S. C. Northwest Arabian sea and Gulf of Oman. In Seas at the Millennium: An Environmental Evaluation (ed. Sheppard, C. R. C.) vol. 3 17–33 (Elsevier Science, 2000).
Hemmati, M. R., Shojaei, M. G., Mirghaed, A. T., Farahani, M. M. & Weigt, M. Food sources for camptandriid crabs in an arid mangrove ecosystem of the Persian Gulf: a stable isotope approach. Isotopes Environ. Health Stud. 57, 457–469 (2021).
Shahraki, M., Fry, B., Krumme, U. & Rixen, T. Microphytobenthos sustain fish food webs in intertidal arid habitats: A comparison between mangrove-lined and un-vegetated creeks in the Persian Gulf. Estuar. Coast. Shelf Sci. 149, 203e212 (2014).
Kruger, B. R. et al. Organic matter transfer in Lake Superior’s food web: Insights from bulk and molecular stable isotope and radiocarbon analyses. Limnol. Oceanogr. 61, 149–164 (2016).
Smith, C. B. & Littleton, J. Multi-species analysis of stable carbon and nitrogen isotope data from Qalʿat al-Baḥrayn. Proc. Semin. Arab. Stud. 51, 35–54 (2022).
Al-Ramamneh, D., Riek, A. & Gerken, M. Effect of water restriction on drinking behaviour and water intake in German black-head mutton sheep and Boer goats. Animal 6, 173–178 (2012).
Wagner, J. J. & Engle, T. E. Invited review: Water consumption, and drinking behavior of beef cattle, and effects of water quality. Appl. Anim. Sci. 37, 418–435 (2021).
Runa, R. A., Brinkmann, L., Riek, A., Hummel, J. & Gerken, M. Reactions to saline drinking water in Boer goats in a free-choice systemReactions to saline drinking water in Boer goats in a free-choice system. Animal 13, 98–105 (2019).
Squires, V. R. Water requirements of livestock fed on halophytes and salt tolerant forage and fodders. In Halophytic and Salt-Tolerant Feedstuffs. Impacts on Nutrition, Physiology and Reproduction of Livestock (eds. El Shaer, H. M. & Squires, V. R.) (CRC Press, 2015).
Abdel-Rahman, H. A. & Abdel-Magid, I. M. Water conservation in Oman. Water Int. https://doi.org/10.1080/02508069308686 (1993).
Zehtabian, G., Khosravi, H. & Ghodsi, M. High demand in a land of water scarcity: Iran. In Water and Sustainability in Arid Regions. Bridging the Gap Between Physical and Social Sciences (eds. Schneier-Madanes, G. & Courel, M.-F.) 75–86 (Springer, 2010).
Hatami, E. & Khosravi, A. R. Mapping of geographic distribution of C3 and C4 species of the family Chenopodiaceae in Iran. Iran. J. Bot. 19, 263–276 (2013).
Mahdavi, P. & Bergmeier, E. Distribution of C4 plants in sand habitats of different climatic regions. Folia Geobot. 53, 201–211 (2018).
Rudov, A., Mashkour, M., Djamali, M. & Akhani, H. A review of C4 plants in Southwest Asia: An ecological, geographical and taxonomical analysis of a region with high diversity of C4 eudicots. Front. Plant Sci. 11, (2020).
Waelkens, M. et al. Archaeometrical research at Sagalassos in 2013. Arkeom. Sonuç. Toplantısı 30, 147–156 (2014).
Rogers, B. et al. Isotopic analysis of the Blick Mead dog: A proxy for the dietary reconstruction and mobility of Mesolithic British hunter-gatherers. J. Archaeol. Sci. Rep. 24, 712–720 (2019).
Guiry, E. J. & Grimes, V. Domestic dog (Canis familiaris) diets among coastal Late Archaic groups of northeastern North America: A case study for the canine surrogacy approach. J. Anthropol. Archaeol. 32, 732–745 (2013).
Guiry, E. J. Dogs as analogs in human stable isotope based paleodietary reconstructions: A review and consideration for future use. J. Archaeol. Method Theory 19, 351–376 (2012).
Guiry, E. J. & Gaulton, B. C. Inferring human behaviors from isotopic analyses of rat diet: a critical review and historical application. J. Archaeol. Method Theory 23, 399–426 (2016).
Tlili, S. Animals in Islamic law and Muslim culture. In The Oxford Encyclopedia of Philosophy, Science, and Technology in Islam (2014).
Tlili, S. Animal ethics in Islam: A review article. Religions 9, 269 (2018).
Barrett, J. H. et al. Interpreting the expansion of sea fishing in medieval Europe using stable isotope analysis of archaeological cod bones. J. Archaeol. Sci. 38, 1516–1524 (2011).
Soto, D. X. et al. Terrestrial contributions to Afrotropical aquatic food webs: The Congo River case. Ecol. Evol. 9, 10746–10757 (2019).
Brozou, A. et al. Aquatic resource consumption at the Odense leprosarium: Advancing the limits of palaeodiet reconstruction with amino acid δ13C measurements. J. Archaeol. Sci. 141, 105578 (2022).
Ma, Y. et al. Aminoisoscapes and palaeodiet reconstruction: New perspectives on millet-based diets in China using amino acid δ13C values. J. Archaeol. Sci. 125, 105289 (2021).
FEDIAF. Facts & Figures 2021. 11 www.fediaf.org (2022).
Brooks, H. L. et al. The power of support from companion animals for people living with mental health problems: A systematic review and narrative synthesis of the evidence. BMC Psychiatry 18, 31 (2018).
Miles, J. N. V., Parast, L., Babey, S. H., Griffin, B. A. & Saunders, J. M. A propensity-score-weighted population-based study of the health benefits of dogs and cats for children. Anthrozoös 30, 429–440 (2017).
Onar, V. et al. A Cat Skeleton from the Balatlar Church Excavation, Sinop, Turkey. Animals 11, 288 (2021).
Vigne, J. D., Guilaine, J., Debue, K., Haye, L. & Gérard, P. Early taming of the cat in Cyprus. Science 304, 259 (2004).
Acknowledgements
This project has received funding from the European Research Council (ERC) under the European Union's Horizon 2020 research and innovation programme (project FELIX, grant agreement n° 101002811). The work undertaken at the Moesgaard Archaeo-Science Laboratory was also funded through a starting grant (project number: AUFF-E-2015-FLS-8-2) awarded by the Aarhus University Research Foundation to Marcello A. Mannino (Aarhus University). The authors are thankful to Mathieu Boudin (Royal Institute for Cultural Heritage, Brussels) for providing laboratory facilities, to Steven Bouillon (KU Leuven) for conducting the isotopic measurements, to Tess Van Den Brande (Royal Institute for Cultural Heritage, Brussels), Zita Kelemen (KU Leuven) and Yannick Stroobandt (KU Leuven) for technical support.
Author information
Authors and Affiliations
Contributions
Conceived and designed the study: W.V.N., B.D.C. Acquired funding: W.V.N., C.O., M.A.M. Provided materials and context: B.D.C., A.M., H.M., J.P., W.V.N. Performed research: A.B., B.T.F., K.V.d.V., W.V.N., O.L., M.A.M. Analyzed the data: A.B., B.T.F., W.V.N. Wrote the original draft: A.B., W.V.N. All authors reviewed the manuscript and approved the final version.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Brozou, A., Fuller, B.T., De Cupere, B. et al. A dietary perspective of cat-human interactions in two medieval harbors in Iran and Oman revealed through stable isotope analysis. Sci Rep 13, 12316 (2023). https://doi.org/10.1038/s41598-023-39417-7
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-023-39417-7
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
