
Abstract
Palaeoparasitological studies can provide valuable information on the emergence, distribution, and elimination of parasites during a particular time in the past. In the prehistoric salt mines of Hallstatt, located in the Austrian Alps, human faeces have been conserved in salt. The aim of this study was to recover ancient DNA of intestinal parasites from these coprolites. Altogether, 35 coprolites from the Hallstatt salt mines, dating back to the Bronze Age mining phase (1158–1063 BCE) and the Iron Age mining phase (750–662 BCE), respectively, were analysed by microscopy and molecular methods. In 91% of the coprolite samples, eggs of soil-transmitted helminths (STH), namely of Trichuris and/or Ascaris were detected by light microscopy. The Ascaris eggs were exceptionally well preserved. For further analysis, DNA was extracted from the palaeofaecal samples and species-specific primers targeting different genes were designed. While amplification of Trichuris DNA remained unsuccessful, sequence data of A. lumbricoides species complex were successfully obtained from 16 coprolites from three different genes, the mitochondrial cytochrome c oxidase subunit 1 gene (cox1), the mitochondrial cytochrome B gene (cytB) and the mitochondrial NADH dehydrogenase subunit 1 gene (nadh1). Importantly, these included two Ascaris sequences from a coprolite from the Bronze Age, which to the best of our knowledge are the first molecular data of this genus from this period.
Similar content being viewed by others


Introduction
Hallstatt is a small village idyllically situated at a lake of the same name in the Eastern Austrian Alps and it is one of the most important archaeological sites in the Old World being the type site for the early European Iron Age, the Hallstatt period. The region is a UNESCO World Heritage area and bears one of the oldest known salt mines in the world. Large-scale underground salt mining is evidenced here since the 14th cent. BCE (Bronze Age). Only four prehistoric salt mines are known worldwide—two of them are in Austria, in Hallstatt and Dürrnberg, the third one is in Chehrabad in North-Western Iran, and the fourth one is in Duzdağı in Azberbaijan1.
Palaeoparasitology is a comparably young discipline allowing the reconstruction of the geographical distribution of parasites and their hosts over time, thereby revealing data on migrations of ancient populations, domestication of animals, cultural habits, hygiene, medical practices and other aspects of life such as sanitation and waste management2. The pioneer of palaeoparasitology, Sir Marc Armand Ruffer, discovered calcified eggs of the blood fluke Schistosoma haematobium in the kidneys of two Egyptian mummies dated to 1250 to 1100 BC3. In 1944, Szidat reported the detection of eggs of Trichuris trichiura, the human whipworm, and Ascaris lumbricoides., the human round worm, in a bog mummy from what was then East Prussia4 and in 1960, Callen and Cameron published a method for rehydrating desiccated ancient material using 0.5% trisodium phosphate aqueous solution5. With this method it became possible to also study parasites from ancient faecal materials (coprolites). In Austria, the first palaeoparasitological studies were performed in the early 1970s on coprolites from the ancient salt mines in Hallstatt and Dürrnberg, reporting the findings of eggs of T. trichiura and A. lumbricoides6.
The aim of this study was to recover ancient DNA of intestinal helminths from coprolites of different time periods from the ancient Hallstatt salt mines. The large organic inventories (e.g. tools, working materials) excavated in the mining areas are well investigated (technology, materials analytics)7, but molecular data are still scarce and open up great potential. For the current study, coprolites were obtained under sterile conditions from different sites within the mining areas corresponding to different time periods and compared to archived material. Novel primers were designed to amplify short fragments of three mitochondrial marker genes for identification. As ancient DNA is usually available only at extremely low concentrations8, mitochondrial markers were selected as PCR targets, mitochondrial DNA (mtDNA) being present in a much higher copy numbers than genomic DNA.
Material and methods
Sampling sites
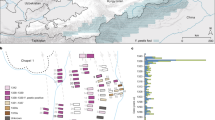
The sampling sites included in this study were the late Bronze Age mining gallery located in the modern-day “Christian von Tuschwerk” site, as well as the early Iron Age sites “Kernverwässerungswerk”, “Kübeck” and “Josef Ritschner Werk” (Fig. 1). The sites in the Bronze Age mining areas, as well as early Iron Age mining areas are located more than 100 m below ground. The production debris of prehistoric mining activity, such as waste rock, left-over salt, burnt torches and broken tools, as well as the miners’ excrements remained in the mines and formed thick layers of waste. After the abandonment of the mine, mountain pressure re-closed all prehistoric mining chambers enclosing all materials left behind and compressing the debris layers to rock-like hardness. Due to the high salt concentration organic objects remained perfectly preserved over the millennia7,9. The Natural History Museum Vienna has been conducting excavations in the prehistoric mine workings since the 1960s. The mining phases have been dated by means of dendrochronology: The late Bronze Age “Christian von Tuschwerk” site dates to (1158–1063 BCE) and the early Iron Age sites mentioned above date to (750–662 BCE)10. The coprolites (Fig. 2) are embedded in the mining debris and can be found at various sites. Temperature and humidity are relatively constant all the year round and there is no natural light.

Cross-section showing the distribution of prehistoric mining sites in the Hallstatt salt mine. Green—Bronze Age, Blue—late Iron Age, Red—early Iron Age. 15—Kübeck (Aufdeckungsschlag) site, 5—Josef Ritschner Werk (Sinkwerksebentel) site, 2—Kernverwässerungswerk site, 8—Christian von Tuschwerk site (D. Brandner/NHM Vienna). Small window: Map showing the location of the site within Austria and Europe (generated by Julia Klammer).

Coprolites embedded in soil. Coprolites can be distinguished from the surrounding material by their brownish colour and their amorphous texture. (A) archived coprolites. (B) fresh coprolites at the sampling site. (C) sampling after removal of surface layers.
Samples
In total, 35 coprolite samples from the ancient Hallstatt salt mine were analysed in this study. Four samples were from the late Bronze Age (12th to 11th cent. BCE) and 31 samples from the early Iron Age (8th to 5th cent. BCE). The samples were collected on five different dates several years apart (1989, 1993, 2003, 2015, 2018 and 2019). Twenty-six samples were from the collection of the Natural History Museum Vienna (Fig. 2A), four samples had been collected in 2018 for a previous study on coprolites from the Hallstatt salt mine11, and five additional samples were collected in 2019 for this study (Fig. 2B,C). These nine samples were collected under sterile conditions. First, the outermost layer was scraped off with a sterile scalpel and then the ancient faecal material was directly sampled into a sterile tube with a fresh sterile scalpel using double sterile disposable gloves and an FFP2 mask. During transport, the samples were refrigerated. In the laboratory, the samples were processed under sterile conditions in a laminar air flow cabinet (Heraeus HERAsafe® HSP12, Kendro Laboratory Products GmbH, Hanau, Germany).
All samples were stored in individual, labelled plastic containers and opened and processed under sterile conditions. The colour of the coprolites was from greyish-light brown to dark brown (Fig. 2). The samples from the museum collection were dehydrated, the samples from 2018 and 2019 were moist and had a soft, clay-like texture.
Microscopic detection
All samples were screened microscopically. For this, we tried and compared several protocols from the literature12,13,14,15 and established a modified protocol. In brief, aliquots of approximately 30 mg of each sample were transferred into a 1.5 ml Eppendorf tube and rehydrated in 0.5 ml 10% NaOH at room temperature by gentle shaking on a rocking platform overnight. On the following day, the samples were vortexed, and 40 μl of each suspension were placed onto a slide and investigated by light microscopy using a Nikon Eclipse E800 microscope (Nikon Instruments Inc., USA). Three slides per sample were analysed. Micrographs were taken with a microscope camera (Nikon DS-Fi2, Nikon Metrology, Germany) and estimations of egg numbers were made as described previously6,15. Rehydrated samples were stored at 4 °C.
Single egg isolation
In order to prove that the eggs detected by microscopy still contained DNA, single eggs were isolated from samples with intact well-preserved eggs. For this, a drop of 100 μl dH2O was placed centrally into a sterile petri dish and 25 μl of the respective sample were added and mixed with the water. Single eggs were isolated with a micropipette (volume 5–10 μl) using an inverted microscope (Nikon TMS, Nikon Instruments Inc., USA). Single eggs were individually transferred to 1.5 ml tubes containing 50 μl of 70% ethanol for DNA extraction with the DNeasy® PowerSoil® Kit (QIAGEN, Hilden, Germany) as described below.
DNA extraction
Altogether, 33 samples were selected for molecular analyses, from two of these we also had single eggs. As it is known that the yield of aDNA can vary significantly depending on the extraction protocol16, four different DNA extraction methods were evaluated and compared, including the QIAamp® DNA Mini Kit (QIAGEN, Hilden, Germany), the QIAamp® DNA Stool Kit (QIAGEN, Hilden, Germany), the GeneClean® Kit for ancient DNA (MP Biomedicals, USA) and the DNeasy® PowerSoil® Kit (QIAGEN, Hilden, Germany). The latter, especially designed to remove inhibitors commonly found in soil and environmental samples, gave the highest DNA yields and was thus used for all samples. In brief, approximately 30 mg of rehydrated coprolite samples were processed with a Precellys® 24 homogenizer (Bertin Instruments, France). Here, best results were obtained with 0.7 mm garnet beads (compared to 0.5 mm glass beads, 0.1 mm zirconium ceramic oxide beads, and a mix of these both). To remove as much NaOH as possible, samples were washed three times by transferring the samples with the beads to a fresh tube, adding 400 μl of sterile ddH2O, gentle vortexing and centrifugation at 20,000 × g/5 min. After washing, as much liquid as possible was removed, the pellet was resuspended in 200 µl sterile ddH2O and, together with the beads, transferred back to the PowerBead tubes. The samples were gently vortexed and then processed in the same way as the tubes with the single eggs by adding 60 μl of solution C1 (DNeasy® PowerSoil® Kit) and inverting the samples several times. Then, the samples were subjected to 3 cycles of homogenization at 5,800 rpm for 15 s with 60 s breaks in between and incubated overnight at 56 °C and 400 rpm. After vortexing and centrifugation at 10,000 × g/30 s, the supernatants were transferred to clean 2 ml collection tubes and washed according to the manufacturers’ instructions. The DNA was eluted in 20–30 μl Elution buffer, depending on the number of eggs in the sample, and stored at − 20 °C. Total DNA yields as measured by a NanoDrop™ spectrophotometer (ThermoFisher Scientific, Waltham, USA) ranged between roughly 50–500 ng/μl and were highest in the freshly obtained, non-dehydrated samples, as already shown in a previous study11.
Primer design
Several primers from the literature, in-house primers and primers from a previous study11 were tested and compared. However, none of these primer pairs gave an amplicon of the correct size in the respective PCRs. Therefore, new primers for each species (Tables 1 and 2) were designed based on reference sequences from the NCBI database (https://www.ncbi.nlm.nih.gov/) and by using GeneDoc (National Resource for Biomedical Supercomputing, Pittsburgh, USA), ClustalX (Conway Institute, Dublin, Ireland) and OligoCalc (http://biotools.nubic.northwestern.edu/OligoCalc.html). The primers were designed to amplify short sequences with a length between 97 and 246 base pairs (bp), as ancient DNA usually is highly fragmented.
For Ascaris, four specific primers pairs were designed, including two primer pairs targeting the mitochondrial cytochrome c oxidase subunit 1 gene (cox1) with overlapping amplicons. The first primer pair was slightly modified from the literature (ACF2 and ACR2)12 and flanks an approximately 182 bp long fragment, the second primer pair amplifies an approximately 172 bp long fragment. The third primer pair targets a ~ 193 bp region within the mitochondrial NADH dehydrogenase subunit 1 gene (nadh1) and the fourth primer pair targets the mitochondrial cytochrome B gene and results in a ~ 138 bp amplicon.
Polymerase chain reactions (PCRs)
All amplifications were carried out in 50 μl reaction volumes and tested in independent setups with different DNA concentrations and using a Hot FIREPol DNA polymerase (5 U/μl, Solis BioDyne, Tartu, Estonia). All PCRs were run in a thermocycler (Eppendorf, Hamburg, Germany) starting with an initial denaturation step (hot start) at 95 °C for 15 min, followed by 35 cycles of 1 min at 95 °C, 45–50 s between 52 and 54 °C depending on primer and 50 s at 72 °C with a final elongation step at 72 °C for 7 min. The cycling conditions for all primer pairs are given in Tables 1 and 2. The PCR results were visualised by agarose gel electrophoresis using a transilluminator (Gel Doc XR +, Bio-Rad Laboratories, Inc., California, USA). Amplicons were stored at 4 °C for further use.
DNA sequencing
All bands of appropriate sizes were excised from the gels with a sterile scalpel. DNA was purified from the bands using the GFX™ PCR DNA and Gel Band Purification Kit (Cytiva Europe GmbH, Wien, Austria) according to the manufacturer´s instructions. The DNA was eluted in 10–50 μl elution buffer depending on the intensity of the bands. Purified DNA was stored at − 20 °C. Sequences were obtained from both strands in at least two independent setups using the BigDyeTM Terminator v1.1 Cycle Sequencing Kit and a SeqStudio automated sequencer (Applied Biosystems, ThermoFisher Scientific, Waltham, USA). The obtained sequences were assembled to consensus sequences using GeneDoc and Chromas (Technelysium Pty Ltd., Australia) and compared to reference sequences using ClustalX. MEGA-X (Molecular Evolutionary Genetics Analysis, Pennsylvania State University, USA) was used for further data analysis.
Results
Microscopy
Altogether, 35 coprolite samples, four from the Bronze Age and 31 samples from the Iron Age, were analysed by microscopy after rehydration. Of these 35 samples, 32 (91%) were positive for eggs of intestinal helminths (Fig. 3). Eggs of Trichuris were detected most frequently, namely in 31 samples. Of these, 14 samples contained only eggs of Trichuris and 17 samples additionally contained Ascaris eggs. One sample contained only eggs of Ascaris. Helminth eggs were detected in three of the four coprolites from the Bronze Age, two samples (sample IDs: 3.4041 and 2019/1) containing only eggs of Trichuris and one sample (ID: 18.215) containing eggs of Trichuris and of Ascaris. As already described in previous microscopic studies on coprolites from these sites6, egg densities were comparably high with up to ~ 7000 eggs/g. Compared to Trichuris eggs, Ascaris eggs were exceptionally well preserved (see also Supplementary Fig. 1).

Micrographs of helminth eggs from coprolites. (A) Ascaris egg from a sample from the Bronze Age. (B) Trichuris egg from a sample from the Iron Age with damage on the egg shell. Interference contrast microscopy, magnification: 400 ×.
As based on morphology only, it is not possible to differentiate between eggs of Trichuris trichiura and T. suis and between Ascaris lumbricoides and A. suum, respectively, we refer to Trichuris eggs and Ascaris eggs here.
No eggs of other helminths were detected in any of the samples.
PCR results
Specific PCRs were run with all 18 samples that were microscopically positive for Ascaris (of two of these we also had single eggs), and with 10 samples microscopically positive for Trichuris (see also Supplementary Table 1). From 16 Ascaris samples, including one sample also as single egg, amplicons of the correct lengths were obtained; in some cases, fragments of more than one gene were successfully obtained. Of the Trichuris samples, three did give amplicons, however not of the appropriate size. For all other Trichuris samples, no bands were retrieved. All visible PCR products were excised from the respective gels for DNA sequencing.
Ascaris specific PCRs
In total, 31 PCR runs resulted in amplicons of the correct lengths. Cytochrome c oxidase subunit 1 (cox1) amplicons were obtained from 15 samples, cytochrome B (cytB) amplicons from nine samples and NADH dehydrogenase subunit 1 (nadh1) amplicons from seven samples. All amplicons were subjected to Sanger sequencing. Reliable sequences were obtained from 29 amplicons, including 15 cox1, eight cytB and six nadh1 sequences. Of the cox1 sequences, 11 samples were of high quality and consensus sequences were generated from both strands. The sequences of the remaining four PCR amplicons were too short and/or had too many ambiguous bases and were therefore excluded from further analyses. The obtained cox1 consensus sequences have a length of 142 bp. Sequence identities ranged from 129 to 141/142 bp (92.81% to 99.30%). Of the eight cytB sequences, five gave reliable consensus sequences with a length of 97 bp and identities ranging from 90 to 96/97 bp (93.75% to 98.97%). Nadh1 sequences were obtained from six samples of which five were of good quality. The obtained consensus sequences had a length of 152 bp and sequence identities ranged from 136 to 148/152 bp (90.67–98.00%). For six samples, sequences of at least two different genes were obtained. With these fragments, the samples were reliably identified as A. lumbricoides species complex, those that had the highest number of unambiguous bases showed highest identity to A. lumbricoides species complex clade A and lowest identity to clade C.
Discussion
This study provides the first two ancient DNA sequences of the intestinal parasite A. lumbricoides species complex from the Bronze Age. Moreover, several sequences of different genes of A. lumbricoides species complex of coprolites from the Iron Age were obtained. This study revealed a high detection frequency of soil-transmitted intestinal helminths in the coprolites of the Hallstatt miners of the Bronze and the Iron Age, however only eggs of Trichuris and Ascaris were found.
Overall, 91% (32/35) of the palaeofaecal samples investigated revealed eggs of intestinal helminths, with Trichuris sp. being most abundant, detected in 31 samples. Fourteen samples also contained eggs of Ascaris sp. and one sample only contained Ascaris eggs. Importantly, eggs of both genera were detected in coprolites from both, the Bronze and the Iron Age. Thus, based on the current archaeological dating of the sampling sites10, it can be stated with certainty that the Hallstatt miners were infested with whipworms and roundworms, at least in the period between 1,158 and 662 BC. The actual parasite burden of the prehistoric miners or even more so of the general population of Hallstatt is difficult to assess, however our results and also those of previous studies6,11 indicate that both helminths were rather common at the time. Light infestations with these worms usually cause no symptoms, but people with heavy infestations or children may suffer from abdominal pain, diarrhoea and rarely also more severe symptoms. As the worms are comparably large and excreted when they die, it can be assumed that the miners knew about them. Preserved leaves of butterbur (Petasites officinalis and Petasites paradoxus) have been found in the mines, which are still today used in folk medicine to treat abdominal pain17. Both worm species are soil-transmitted helminths (STH) without an intermediate host, adult female worms producing thousands of eggs daily that are shed with the human stool. The eggs have to mature in the soil into infective eggs, which are then taken up orally via contaminated hands or foods by the next human host. Depending on the external conditions, it takes around 3–6 weeks for the eggs to become infective. The occurrence of roundworms and whipworms is an indication of poor sanitation and hygiene18. In Hallstatt, particularly during the Iron Age, hygiene conditions in the mines were obviously low, as cooking, eating and defecation were accomplished at the same spots. This is corroborated by the fact that palaeofeces can be found distributed throughout the salt mines6,7,10,11,19. So far, only one larger pile of palaeofaeces was found in the mining areas, namely at the “Kernverwässerungswerk” site, which might indicate that this spot was used more than once. A constant temperature of 8 °C and comparably high humidity prevails inside the mines and prehistoric miners worked closely together in working groups7,10,19, thus conditions were favourable for survival and transmission of helminth eggs. Ascaris and Trichuris are among the most common helminths found in paleoparasitological samples, and together with the hookworms, they are still the most common helminths in humans today, although no longer endemic in most parts of Europe, including Austria. Worldwide, approximately 807-1,121 million people are infested with Ascaris and approximately 604–795 million with Trichuris20. Eggs of T. trichiura were also detected in intestinal samples of the famous frozen “Iceman” mummy found in the Ötztal Alps dated to the 1st half of the 4th mil. BC21,22. Interestingly, no eggs of other intestinal helminths were found in this study, particularly no eggs of parasites associated with the consumption of meat or fish. In a previous study on coprolites from this salt mine11, structures resembling eggs of Taenia sp. and Dibothriocephalus latus (= Diphyllobothrium latum), were found, but could not be reliably identified. In the other prehistoric Austrian salt mine, which is located in Dürrnberg (Salzburg) around 60 km from Hallstatt, eggs of Fasciola hepatica, Dicrocoelium dendriticum and Taenia sp. have been found together with eggs of Ascaris and Trichuris in coprolites6,19,23. The tapeworms Taenia spp. and D. latus are transmitted via infested pork/beef and fish, respectively, if consumed raw or undercooked. Archaeological and archaeozoological studies suggest, that already during the Bronze Age, not only salt production but also meat (mainly pork) processing and conservation was an important economic branch in Hallstatt7,24. Butchered and cut meat parts were delivered to the mining community and cured with salt-rich mining debris in large wooden basins. In addition, archaeozoological and molecular analyses evidence that meat from different animals (pork, goat, beef and mutton) was consumed25,26. Earlier studies on palaeofaeces and the investigation of ancient cooking utensils have revealed that the Iron Age miners typically consumed a stew that mainly consisted of millet, barley, beans and meat and was cooked for many hours27. However there is also molecular evidence for substantial complexity in the prehistoric miners diet including the consumption of fermented foods (blue cheese) and beverages (beer) in Iron Age26.
In this study, a total of 19 A. lumbricoides species complex-specific sequences from different genes were successfully amplified and sequenced. This is the first time that ancient Ascaris DNA has been recovered from coprolites dated to the Bronze Age in Austria. In a previous study, ancient Ascaris DNA was detected by PCR, but no reliable sequence data was obtained11. It is noteworthy, that in the current study, only DNA from Ascaris was successfully amplified, while Trichuris DNA was not, either from the Bronze Age, or from the Iron Age. All PCRs with Trichuris-specific primers targeting different genes were unsuccessful. Molecular analysis of ancient material is challenging, the rate of DNA degradation depends on various external and internal factors28. We hypothesize that the specifically thick-walled Ascaris eggs protected the DNA better than the plugged Trichuris eggs. Morphologically, the Trichuris eggs showed more damage than the Ascaris eggs, and most of the eggs had lost the polar plugs and thus the internal contents were fully exposed to the salt. In other archaeological sites (but without high salt content), aDNA of parasites has been detected even in the absence of microscopically visible parasitic structures29,30.
Two aDNA fragments of A. lumbricoides species complex were obtained from a coprolite dated to the Bronze Age between 1158 and 1063 BC10, including a 142 bp long cox1 sequence and a 152 bp long fragment of the nadh1 gene. While several studies report the recovery of ancient parasitic DNA from archaeological sites dated to the Iron Age and later31,32,33,34,35, there is only a limited number of reports on intestinal helminth remains from the Bronze Age worldwide, and even fewer with molecular data. In a recent study, eggs of Dibothriocephalus sp., Trichuris sp., Dioctophyme renale (a dog parasite, only very rarely affecting humans), Echinostoma sp. and Capillaria sp. were detected by digital light microscopy in coprolites from a Late Bronze Age Must Farm in wetlands in England, UK36. On the island of Kea, Greece, eggs of Ascaris sp. and Trichuris sp. were detected in soil sediment samples from the sacrum and iliac area of burials from the Neolithic to Byzantine period37. In Sardinia, Italy, eggs of Ascaris and Trichuris were found in Bronze Age pits and wells38. Earlier studies on Bronze Age samples from Europe include findings of Ascaris, Trichuris, Ancylostoma duodenale (one of the two human hookworms) and Dicrocoelium dendriticum (the lancet liver fluke)39,40,41. Outside Europe, parasite eggs from archaeological excavations dated to the Bronze Age have been reported particularly also from Iran42,43,44. In a burial of a Bronze Age cemetery (2600–2200 BC), eggs of D. dendriticum were found, which represent the oldest finding of this parasite in the Near East42. Findings of D. dendriticum eggs in human faeces indicate a close relationship between humans and herbivores, as the life cycle of this parasite includes sheep, snails and ants. In Tel Meggido, Israel, a poorly preserved Trichuris egg was detected during microscopic investigations of faecal material found near the remains of a large palace dated to the Late Bronze Age45. But all these data were obtained by microscopy only. In sediment samples from the Neolithic (5320–4980 BC) lakeside settlement of La Draga in Spain, DNA of Ascaris sp., T. trichiura, E. vermicularis, T. saginata and D. dendriticum has been detected by multiplex PCRs. Again, two taxa (D. dendriticum and E. vermicularis) were not detectable by microscopy, which however revealed also Dibothriocephalus sp., Capillaria sp., Paramphistomum sp. (parasite of ruminants) and Macracanthorhynchus sp. (animal parasite, affecting mostly pigs and wild boar)30. In 2018, eggs of Toxascaris sp. were detected in a mammalian carnivore coprolite from an archaeological site in Argentina dated to 14,981–14,552 BC, confirmed by ancient mitochondrial DNA analysis, which identified the parasite eggs as Toxascaris leonina, a parasite of dogs, cats and other carnivores that can also infest humans, representing the oldest molecular record of a parasite worldwide46.
Limitations of this study were that the sample set was comparably small, with only 35 coprolites included. Also, the samples had been collected at different time points and stored for different periods of time. Moreover, this study only attempted to amplify aDNA of those parasites, of which eggs were detected by microscopy. Currently available molecular data do not allow for a reliable differentiation of A. lumbricoides and A. suum and rather suggest that Ascaris infecting humans is a species complex47,48, we thus refered to A. lumbricoides species complex throughout.
In conclusion, this study presents the first successfully amplified and sequenced ancient Ascaris DNA sequences from the Bronze Age and also the first micrographs of intestinal helminth eggs from the Bronze Age in Austria. Additionally, several sequences of different genes of A. lumbricoides species complex were obtained from coprolites from the Iron Age. Microscopically, Ascaris and Trichuris were detected in 91% of the palaeofaecal samples, indicating a high frequency of these parasites in at least a part of the prehistoric population. Interestingly, however, only these two parasite species were found.
Data availability
All sequence data have been submitted to GenBank and are available under the following accession numbers: OQ721964-OQ721974 (cox1), OQ626561-OQ626564 (cytB), OQ626565-OQ626569 (nadh1). Details of the methodological procedures are available in the Master Thesis that derived from this work49. All other data generated are available through the authors upon request.
References
Gonon, T. et al. Early salt-mining systems and strategies at Duzdağı, Nakhchivan. In On Salt, Copper and Gold: The Origins of Early Mining and Metallurgy in the Caucasus (eds Marro, C. & Stöllner, T.) (MOM Éditions, 2021).
Le Bailly, M. & Araújo, A. Past intestinal parasites. In Paleomicrobiology of Humans (eds Drancourt, M. & Raoult, D.) 143–154 (American Society for Microbiology, 2016). https://doi.org/10.1128/9781555819170.chap14.
Ruffer, M. A. Note on the presence of Bilharzia haematobia in Egyptian mummies of the twentieth dynasty. Brit. Med. J. 2557, 16 (1910).
Szidat, L. Über die Erhaltungsfähigkeit von Helmintheneiern in Vor- und Frühgeschichtlichen Moorleichen. Zeitschr. f. Parasitenkunde 13(3), 265 (1944).
Callen, E. O. & Cameron, T. W. M. A prehistoric diet revealed in coprolites. N. Sci. 8(190), 35–40 (1960).
Aspöck, H., Flamm, H. & Picher, O. Darmparasiten in menschlichen Exkrementen aus prähistorischen Salzbergwerken der Hallstatt-Kultur (800–350 v. Chr.). Zbl. Bakt. Hyg. I Abt Orig A, 549–558. (1973).
Reschreiter, H. & Kowarik, K. Bronze age mining in hallstatt. A new picture of everyday life in the salt mines and beyond. Archaeol. Austriaca 103, 99–136. https://doi.org/10.1553/archaeologia103s99 (2019).
Loreille, O., Roumat, E., Verneau, O., Bouchet, F. & Hänni, C. Ancient DNA from Ascaris: Extraction amplification and sequences from eggs collected in coprolites. Int. J. Parasitol. 31(10), 1101–1106 (2001).
Tintner, J. et al. Aging of wood under long-term storage in a salt environment. Wood Sci. Technol. 50(5), 953–961 (2016).
Grabner, M. et al. Prehistoric salt mining in Hallstatt, Austria New chronologies out of small wooden fragments. Dendrochronologia https://doi.org/10.1016/j.dendro.2021.125814 (2021).
Rodler, K. M. Identification of Gastrointestinal Parasites in Ancient Fecal Samples from the Hallstatt Salt Mine (Master thesis, University of Vienna, 2019). 93 pages
Søe, M. J. et al. Ancient DNA from latrines in Northern Europe and the Middle East (500 BC–1700 AD) reveals past parasites and diet. PLoS ONE 13(4), 1–17. https://doi.org/10.1371/journal.pone.0195481 (2018).
Dufour, B. & Le Bailly, M. Testing new parasite egg extraction methods in paleoparasitology and an attempt at quantification. Int. J. Paleopathol. 3(3), 199–203. https://doi.org/10.1016/j.ijpp.2013.03.008 (2013).
Anastasiou, E. & Mitchell, P. D. Simplifying the process of extracting intestinal parasite eggs from archaeological sediment samples: A comparative study of the efficacy of widely-used disaggregation techniques. Int. J. Paleopathol. 3(3), 204–207. https://doi.org/10.1016/j.ijpp.2013.04.004 (2013).
Camacho, M., Araújo, A., Morrow, J., Buikstra, J. & Reinhard, K. Recovering parasites from mummies and coprolites: An epidemiological approach. Parasit. Vectors 11(1), 248. https://doi.org/10.1186/s13071-018-2729-4 (2018).
Hagan, R. W. et al. Comparison of extraction methods for recovering ancient microbial DNA from paleofeces. Am. J. Phys. Anthropol. 171(2), 275–284. https://doi.org/10.1002/ajpa.23978 (2020).
Kromer, K. Funde von Pestwurzblättern im alten Bergbau von Hallstatt. Wissenschaftliche Arbeiten aus dem Burgenland 71, 145–148 (1985).
Flammer, P. G. & Smith, A. L. Intestinal helminths as a biomolecular complex in archaeological research: Parasites as archaeological biomarkers. Philos. Trans. R. Soc. Lond. B. Biol. Sci. 375(1812), 20190570. https://doi.org/10.1098/rstb.2019.0570 (2020).
Stöllner, T. et al. The economy of Dürrnberg-Bei-Hallein: An Iron Age salt-mining centre in the Austrian Alps. Antiqu. J. 83, 123–194. https://doi.org/10.1017/S0003581500077684 (2003).
CDC. Parasites. Global Health, Division of Parasitic Diseases and Malaria. https://www.cdc.gov/parasites/ (2021).
Aspöck, H., Auer, H. & Picher, O. The mummy from the Hauslabjoch: A medical parasitology perspective. Alpe Adria Microbiol. J. 4(2), 105–114 (1995).
Aspöck, H., Auer, H. & Picher, O. Trichuris trichiura eggs in the neolithic glacier mummy from the Alps. Parasitol. Today 12(7), 255–256 (1996).
Aspöck, H., Auer, H. & Picher, O. Parasites and parasitic diseases in prehistoric human populations in Central Europe. Helminthologia 36(3), 139–145 (1999).
Pucher, E., Barth, F. E., Seemann R. & Brandstätter, F. (Eds.) Bronzezeitliche Fleischverarbeitung im Salzbergtal bei Hallstatt. Mitteilungen der Prähistorischen Kommission 80, Wien Verlag der Österreichischen Akademie der Wissenschaften (2013).
Pucher, E. Ältereisenzeitliche Tierknochenfunde aus dem Hallstätter Salzberg. Festschrift für Fritz Eckart Barth. Mitteilungen der Anthropologischen Gesellschaft in Wien 139, 123–133 (2009).
Maixner, F. et al. Hallstatt miners consumed blue cheese and beer during the Iron Age and retained a non-Westernized gut microbiome until the Baroque period. Curr. Biol. 31(23), 5149–5162. https://doi.org/10.1016/j.cub.2021.09.031 (2021).
Barth, F. E. Das Ritschert, eine urzeitliche Reminiszenz. Archäologie Österreich 10(2), 54–58 (1999).
Lindhal, T. Recovery of antediluvian DNA. Nature 365(6448), 700 (1993).
Côté, N. M. L. et al. A new high-throughput approach to genotype ancient human gastrointestinal parasites. PLoS ONE 11(1), 1–18 (2016).
Maicher, C. et al. Paleoparasitological investigations on the Neolithic lakeside settlement of La Draga (Lake Banyoles, Spain). Holocene 27(11), 1659–1668. https://doi.org/10.1177/0959683617702236 (2017).
Tams, K. W. et al. Parasitic infections and resource economy of Danish Iron Age settlement through ancient DNA sequencing. PLoS ONE 13(6), e0197399. https://doi.org/10.1371/journal.pone.0197399 (2018).
Askari, Z. et al. Genomic palaeoparasitology traced the occurrence of Taenia asiatica in ancient Iran (Sassanid Empire, 2th cent. CE-6th cent. CE). Sci. Rep. 12(1), 12045. https://doi.org/10.1038/s41598-022-10690-2 (2022).
Roche, K. et al. Gastrointestinal parasite burden in 4th–5th c. CE Florence highlighted by microscopy and paleogenetics. Infect. Genet. Evol. 90, 104713. https://doi.org/10.1016/j.meegid.2021.104713 (2021).
Flammer, P. G. et al. Epidemiological insights from a large-scale investigation of intestinal helminths in medieval europe. PLoS Negl. Trop. Dis. 14(8), 1–20. https://doi.org/10.1371/journal.pntd.0008600 (2020).
Wood, J. R. DNA barcoding of ancient parasites. Parasitology 145(5), 646–655 (2018).
Ledger, M. L. et al. Intestinal parasites at the Late Bronze Age Settlement of Must Farm, in the fens of East Anglia, UK (9th century B.C.E.). Parasitology 146(12), 1583–1594 (2019).
Anastasiou, E., Papathanasiou, A., Schepartz, L. A. & Mitchell, P. D. Infectious disease in the ancient Aegean: Intestinal parasitic worms in the Neolithic to Roman Period inhabitants of Kea Greece. J. Archaeol. Sci. Rep. 17, 860–864 (2018).
Sabato, D. et al. Archaeobotanical analysis of a Bronze Age well from Sardinia: A wealth of knowledge. Plant Biosyst. 149, 205–215. https://doi.org/10.1080/11263504.2014.998313 (2015).
Le Bailly, M. & Bouchet, F. Ancient dicrocoeliosis: Occurrence, distribution and migration. Acta Trop. 115, 175–180 (2010).
Le Bailly, M. & Bouchet, F. Diphyllobothrium in the past: Review and new records. Int. J. Paleopathol. 3, 182–187 (2013).
Šebela, L., Vojtková, L. & Vojtek, J. Intestinal Parasites in man of old Bronze Age. Moravian Museum Stable 28(1), 105–107; https://www.jstor.org/stable/26294824?seq=1 (1990).
Mowlavi, G., Mokhtarian, K., Sadat, M., Mobedi, I. & Masoumian, M. Dicrocoelium dendriticum found in a Bronze Age cemetery in western Iran in the pre-Persepolis period: The oldest Asian palaeo finding in the present human infection hottest spot region. Parasitol. Int. 64(5), 251–255 (2015).
Nezamabadi, M., Aali, A., Stöllner, T., Mashkour, M. & Le Bailly, M. Paleoparasitological analysis of samples from the Chehrabad salt mine (Northwestern Iran). Int. J. Paleopathol. 3(3), 229–233 (2013).
Makki, M. et al. First paleoparasitological report on the animal feces of bronze age excavated from Shahr-e Sukhteh, Iran. Korean J. Parasitol. 55(2), 197–201. https://doi.org/10.3347/kjp.2017.55.2.197 (2017).
Langgut, D. et al. Micro-archaeological indicators for identifying ancient cess deposits: An example from Late Bronze Age Megiddo, Israel. J. Archaeol. Sci. Rep. 9, 375–385 (2016).
Petrigh, R. S., Martínez, J. G., Mondini, M. & Fugassa, M. H. Ancient parasitic DNA reveals Toxascaris leonina presence in final pleistocene of South America. Parasitology 146(10), 1284–1288 (2019).
Easton, A. et al. Molecular evidence of hybridization between pig and human Ascaris indicates an interbred species complex infecting humans. Elife 9, e61562. https://doi.org/10.7554/eLife.61562 (2020).
Leles, D., Gardner, S. L., Reinhard, K., Iñiguez, A. & Araujo, A. Are Ascaris lumbricoides and Ascaris suum a single species?. Parasit. Vectors 5, 42. https://doi.org/10.1186/1756-3305-5-42 (2012).
Barsch, E. Molecular Analyses of Prehistoric Faecal Samples for Intestinal Parasites (Master thesis, University of Vienna, 2021). 101 pages
Acknowledgements
The authors wish to thank the team of the Molecular Parasitology lab at the Institute of Specific Prophylaxis and Tropical Medicine, Medical University of Vienna, for help and advice.
Author information
Authors and Affiliations
Contributions
K.K., H.R., H.S. and J.W.: Study design. E.B., K.K., K.R., H.R. and J.W.: Sampling. E.B., K.R., C.H. and H.S.: Laboratory work and photography. E.B., K.K. and J.W.: Data analysis. E.B., K.K., and J.W.: Writing of the manuscript. All authors: Review and approval of the final version of the manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.




Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Barsch, E., Kowarik, K., Rodler, K. et al. First molecular data on the human roundworm Ascaris lumbricoides species complex from the Bronze and Iron Age in Hallstatt, Austria. Sci Rep 13, 12055 (2023). https://doi.org/10.1038/s41598-023-38989-8
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-023-38989-8
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.

{kind=link}
{kind=link}
{kind=link}
{kind=link}