
Abstract
The world will be dependent on the development of novel feed ingredients from renewable sources to ensure sustainable growth of the aquaculture industry. Zooplankton like Calanus finmarchicus are viable new raw material candidates, as they have optimal nutrient profiles for aquatic animals and may be sustainably harvested in large volumes. In this study, the aim was to investigate if a protein hydrolysate of C. finmarchicus was able to influence the growth performance of fish. The effect of dietary inclusion of hydrolysates was tested in a feeding trial with European sea bass (Dicentrarchus labrax) juveniles, benchmarking calanus hydrolysate (CH) against commercially available hydrolysates. The diet with CH inclusion yielded increased growth, with significantly higher body weight than hydrolysates of sardine and tuna fish at the end of the trial. The observed growth-promoting effects were further examined using an in vitro model with skeletal muscle cells from Atlantic salmon. Through bioactivity experiments with muscle cells grown in media containing CH, low-molecular fractions were found to have the greatest positive effect on proliferation, viability, and expression of muscle-specific genes. Characterization of the most potent fraction revealed an abundance of small peptides, along with amino acids and marine metabolites associated with increased muscle growth.
Similar content being viewed by others

Introduction
The global population is projected to reach 9.8 billion by 20501, which will require a corresponding increase in global food production. Terrestrial food sources are approaching their maximum sustainable capacity2, whereas only 7% of all proteins consumed globally come from seafood3. Seafood is rich in digestible protein and essential amino acids, and aquatic animals generally have a low feed conversion ratio (FCR) compared to terrestrial animals, which allows for high throughput of dietary protein4. The potential to feed the growing population with a larger share of seafood is obvious, especially as the demand for dietary protein sources with low environmental impact is expected to increase considerably in the coming years5.
In consideration of the uncertainty and present exploitation of many global fisheries, aquaculture is predicted to be the major source of seafood in the future3,5. Along with the expansion of aquaculture production comes questions about its sustainability, particularly with respect to the necessary feed ingredients. Fish meal has historically been the protein source of choice in aquaculture, but its seasonal variations and volume limitations have created needs for additional protein sources in the growing industry. The trend in recent years has thus been to formulate feeds with increasing amounts of terrestrial plant proteins such as soy, maize, and rapeseed6. Terrestrial plant protein sources have sub-optimal amino acid profiles compared to marine proteins, may contain anti-nutritional factors and mycotoxins, and generally display lower palatability7,8,9. These drawbacks are shown to reduce growth in farmed fish and have spurred demands for new and sustainable protein ingredients for aquaculture. Of the proposed novel ingredients, low-trophic marine resources such as zooplankton are viewed as attractive and viable options due to their renewable biomass volumes and their role as natural food in the marine food web. One zooplankton species with feed applications is Calanus finmarchicus, found across the Northern Hemisphere. In the Norwegian Sea and adjacent regions alone, the annual biomass production of C. finmarchicus and closely related species is estimated to be approximately 290 million tonnes, making it one of the most significant renewable resources in the region10. A recent review on mesopelagic species as new marine resources remarked that the biomass of C. finmarchicus is well understood and that the management plan, which issues ten commercial harvesting licenses for a total annual quota of 254,000 tonnes, governs its biological sustainability11. The massive volumes, combined with a suitable nutrient composition for aquatic animals12,13, make C. finmarchicus an especially promising raw material for novel aquaculture feed ingredients.
Enzymatic protein hydrolysis (EPH) is a method often used to process raw material into feed ingredients on an industrial scale. EPH is based on food-grade proteases to digest protein-rich biomasses, typically resulting in products that constitute a water-soluble peptide fraction, lipids, and a non-soluble residue solid fraction. This technology provides a gentle and environmentally friendly alternative to processing techniques with chemicals like solvents and acids. The protein fraction isolated after EPH is termed hydrolysate and can serve as an important ingredient in feed formulations for finfish14. In addition to their nutritional and functional properties, hydrolysates are potential sources of specific bioactive peptides that can have biological functions different from their precursor proteins. These bioactive peptides can display hormone or drug-like properties, and among the mode-of-actions they can exert are anabolic (growth-promoting), antioxidative, antimicrobial, antihypertensive, immunomodulatory, and opioid15. The inclusion of hydrolysates in diets for aquatic animals has been shown to promote growth and increase feed efficiency16, and a deeper look into specific fractions of the hydrolysates could help explain some of these bioactive effects. Previously published studies involving EPH of C. finmarchicus have focused on how to extract the oil phase most effectively17, there is thus little knowledge of the protein fraction and how it can benefit fish growth performance.
In the presented study, the overall aim was to investigate if a hydrolysate from C. finmarchicus could improve growth of the commercially important fish species European sea bass (Dicentrarchus labrax) and Atlantic salmon (Salmo salar). The first part consisted of a feeding trial that assessed how dietary inclusion of calanus hydrolysate influenced growth performance of European sea bass, benchmarking it against other commercial marine hydrolysates. The improved growth performance observed with calanus hydrolysate in the feeding trial was studied further in the second part, using a model of skeletal muscle cells from Atlantic salmon to identify bioactive fractions of the hydrolysate associated with increased skeletal muscle growth.
Results and discussion
Dietary inclusion of a novel protein hydrolysate from the marine zooplankter C. finmarchicus led to improved growth performance of European sea bass in a feeding trial that benchmarked marine protein hydrolysates as functional feed ingredients. The observed growth-promoting properties of calanus hydrolysate (CH) were further evaluated using an in vitro cell model for skeletal muscle. Assessments of how supplementing CH in the cell culture medium affected viability, proliferation, and gene expression in cell studies, followed by fractionation of CH, led to the identification and characterization of the most bioactive fraction associated with increased muscle cell growth. A summary of the workflow is presented in Fig. 1.

Schematic representation of the methods used to examine the growth-promoting properties of a hydrolysate from Calanus finmarchicus. (A) Feeding trial with European sea bass. (B) Bioactivity studies on calanus hydrolysate. CCH: Crude calanus hydrolysate; DCH: Desalted calanus hydrolysate; F1-F6: Fraction 1–6; SEC: Size exclusion chromatography; NMR: Nuclear magnetic resonance spectroscopy.
Feeding trial growth performance
The feeding trial aimed to benchmark the growth-promoting effects of dietary calanus hydrolysate (CH) against commercially available marine protein hydrolysates of sardine (SDH), tuna fish (TH), and salmon (SH). Moderate inclusion levels (5–10%) of marine protein hydrolysates to improve growth performance of fish have been well documented14. Marine hydrolysates may also enhance the nutritional properties to stimulate feed consumption18, and increase intake of nutrients and enhance growth, as observed in starter diets for Atlantic salmon19. Benchmarking CH against similar functional ingredients, thus employing diets with commercial marine hydrolysates as controls, would arguably yield more relevant results than comparing it to a standard fish meal control diet.
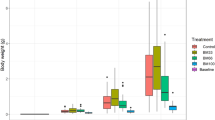
European sea bass juveniles with a starting weight of 8.9 ± 0.4 g fully accepted all the diets. With similar starting weight for all the groups receiving the different diets and equal variances (Table S7.1–S7.3. in supplementary data), one-way ANOVA was applicable to compare the parameters at different time points. No significant effects were observed on survival, whole-body composition, or feed intake (Tables S3–S6 in supplementary data). However, CH inclusion yielded significantly higher body weight than SDH (p = 0.003) and TH (p = 0.047) after 84 days (Fig. 2A) and showed an increased growth rate compared with SDH (p = 0.002) throughout the trial (Fig. 2B). Growth performance is tightly connected with the ability to convert nutrients into body weight, and the relationship between increased body weight and consumed protein is the protein efficiency ratio (PER), which can be used to evaluate the quality of dietary protein ingredients. The fish receiving dietary CH showed higher PER throughout the trial (Fig. 2C), with significantly higher end results compared with the commercial hydrolysates SDH (p = 0.002), TH (p = 0.011), and SH (p = 0.037). Feed conversion ratio (FCR) measures the efficiency in which fish convert their input (feed) into the desired output (body weight), thus it takes the whole feed into account, not only the protein. At the end of the trial, feeds containing the three commercial hydrolysates had FCRs ranging from 1.28 to 1.33, whereas CH had an FCR of 1.22, significantly lower than those of SDH and TH (Fig. 2D). These FCR values correspond well with those found in similar studies with dietary hydrolysates for European sea bass20.

Benchmarking marine hydrolysates as feed ingredients for growth performance and feed utilization in European sea bass (Dicentrarchus labrax). (A) Body weight: fish were group weighed for each tank. (B) Specific growth rate (SGR): [(ln final weight – ln initial weight)/number of days] × 100. (C) Protein efficiency ratio (PER): wet weight gain/crude protein intake. (D) Feed conversion ratio (FCR): crude feed intake/weight gain. CH: Calanus hydrolysate; SDH: Sardine hydrolysate; TH: Tuna hydrolysate; SH: Salmon hydrolysate. Bars are means ± standard deviation (n = 3). Different letters denote statistical difference (P < 0.05).
The results from the growth performance trial highlight the potential of CH as a functional ingredient in fish feeds, showing that dietary CH inclusion can improve growth and increase overall protein retention and feed conversion.
To the best of our knowledge, this is the first published study using a protein hydrolysate from C. finmarchicus in a feeding trial, there are thus few direct comparisons to be made. However, a study indicated that supplementation of C. finmarchicus meal in diets for juvenile Atlantic halibut (Hippoglossus hippoglossus) could greatly improve growth rate and nutrient utilization efficiency21. The meal was processed to increase its protein contents, but unlike optimal protein hydrolysates it also contained considerable amounts of lipids. Another study with zooplankton protein showed that dietary inclusion of Antarctic krill (Euphausia superba) meal for European sea bass enhanced growth performance and feed efficiency22. The evident benefits of including zooplankton such as C. finmarchicus and Antarctic krill in diets for fish should come as no surprise, as zooplankton are natural food for most fish species, especially in early life stages. However, little is known about why zooplankton and their products substantially benefit growth, and the subsequent experiments investigate these properties in further detail.
Considering that the diets were isonitrogenous, isolipidic, and isoenergetic (Table 3, Materials and methods), and that there were no significant differences in feed intake between them, any observed differences in growth performance would be based entirely on the composition and properties of the individual hydrolysates. Composition and properties of marine protein hydrolysates can be different depending on the raw material source, the enzymes used for hydrolysis, and the conditions and duration of hydrolysis18. Evidence that certain hydrolysate fractions can influence growth performance of fish in multiple ways have been found in several studies23,24.
Effect of calanus hydrolysate on primary skeletal muscle cells
The first in vitro bioactivity experiment was performed to determine if the natural mineral contents of CH affected the growth of skeletal muscle, using a model with primary skeletal muscle cells from Atlantic salmon. European sea bass and Atlantic salmon are both teleost fish species and share the fundamental events of the myogenesis25. The salmon primary skeletal muscle cells are well-established models for bioactivity experiments to study muscle growth and its mechanisms26,27,28,29,30.
Three concentrations of crude and desalted CH were added to growth media to test if supplementation of these could affect the muscle cell viability, proliferation, and cytotoxicity. The cell metabolic activity increased significantly when the cell culture media was supplemented with desalted CH at concentrations 1 mg/ml and 0.1 mg/ml (Fig. 3A), compared with cells treated with control medium (without added hydrolysates). The cell proliferation was not affected, except when cells were treated with culture medium containing crude CH at 0.1 mg/ml (Fig. 3B). Only the highest concentration of crude CH (1 mg/ml) supplemented in cell culture media induced increased lactate dehydrogenase (LDH) levels in the cells, which could impact cell survival negatively (Fig. 3C).

Viability (A), proliferation (B), and cytotoxicity (C) of Atlantic salmon muscle cells cultured in growth media supplemented with three concentrations (0.01, 0.1, or 1 mg/ml CH in growth medium) of crude or desalted calanus hydrolysate. CH: Calanus hydrolysate; RFU: Relative fluorescence units; LDH: lactate dehydrogenase; CTL: Control (no added hydrolysate to the cell culture growth media). Results presented with box (25th to 75th percentiles with line at median) and whiskers (min to max) plots (n = 4). Stars (*) denote statistically significant differences from control (P < 0.05).
The initial bioactivity experiment showed that the desalted hydrolysates performed better in cell studies (Fig. 3). A putative explanation is that cultured cells are vulnerable to mineral contents higher than their innate concentrations, as high mineral concentrations can cause an osmotic effect where the cells lose water and shrink31. Whereas cultured cells can react negatively to elevated mineral contents, the opposite is true for most aquatic animals, especially those of marine origin, which require significant amounts of micronutrients like minerals in their diets32. Nonetheless, the results suggest the positive effects CH has on muscle growth stem from its inherent peptides, not from its mineral contents, and the desalted CH was thus chosen for the remaining in vitro experiments.
As the next step to further narrow down the list of potentially bioactive compounds in CH, the desalted CH was fractionated to investigate how differently sized peptide fractions affect muscle growth. CH was separated using size-exclusion chromatography (SEC) and yielded six fractions with average molecular weights of >742 Da (F1), 742 Da (F2), 527 Da (F3), 407 Da (F4), 316 Da (F5), and 260 Da (F6). The muscle cells were then cultured in growth media containing each of the peptide fractions to measure their individual influence, while the control cells were cultured in growth medium without supplementation of peptide fractions. F3, F4, and F6 were associated with the highest cell viability, significantly higher than the control and fractions F1 and F2, while F5 showed an intermediate effect (Fig. 4A). Likewise, cell culture medium supplemented with F6 induced the highest proliferative capacity, significantly higher than control and F5 (Fig. 4B). Overall, the results showed that fractions of lower molecular weight affected muscle growth more positively than those of higher molecular weight fractions. This is consistent with previous studies that showed growth-promoting effects of small molecular weight peptides33. Bakke et al.23 revealed that small peptides and amino acids formed during hydrolysis can facilitate the absorption of molecules in the intestinal tract by increasing the expression of peptide transporters. Rashidia, et al.34 found that low-molecular fractions from shrimp waste hydrolysate can improve growth performance of rainbow trout (Oncorhynchus mykiss), and Zheng et al.24 indicated a positive effect on growth and feed utilization in juvenile turbot (Scophthalmus maximus) by the use of low-molecular weight compounds from fish protein hydrolysate. Low-molecular weight fractions from protein hydrolysates have also been shown to be more bioactive in mammalian primary muscle cell studies35,36.

Viability (A) and proliferative activity (B) of Atlantic salmon muscle cells cultured in growth media supplemented with six size-separated fractions of desalted CH (0.1 mg/mL). RFU: Relative fluorescence units; CTL: Control; F1-F6: CH fractions 1–6. Results presented with box (25th to 75th percentiles with line at median) and whiskers (min to max) plots (n = 5). Lower case letters (a, b, c) denote statistically significant differences (P < 0.05).
Gene expression analysis
The two most promising fractions from the viability (F4 and F6) and proliferation (F6) assays were chosen for gene expression analysis. Expression of genes associated with muscle growth was analyzed for cells cultured in media supplemented with F4 and F6, to explore the influence of these potent fractions in more detail. Proliferation and differentiation of myoblasts to multinuclear myotubes require the expression of genes coding for muscle regulatory factors such as myogenic factor 5 (Myf5) and myoblast determination protein 1a (MyoD1a)37,38. Myf5 and MyoD1a are early markers responsible for specification of cells to the muscle lineage25. The muscle regulatory factors are essential for the transcription of several muscle-specific genes, resulting in increased expression of the structural muscle proteins like myosin heavy chain (MHC). In differentiating muscle cells, early markers will be downregulated while late markers like structural muscle proteins will be upregulated25. Muscle cells cultured in growth media with F4 did not affect the gene expression of either myf5 or myod1a, whereas F6 induced downregulation of both genes (Fig. 5A and B). Although the results were just below the statistical limit set for significance, there was a trend toward higher expression of mhc (Fig. 5C), with F6 showing the highest upregulation (p = 0.0518). A downregulation of the early muscle markers myf5 and myod1a, concomitantly with a trending upregulation of mhc, may indicate that CH peptide fractions positively affected the muscle cells by increasing their rate of differentiation. The positive effects observed on in vitro muscle cell differentiation could be explained by the composition of certain small peptides and amino acids in F4 and F6. Supplementation of amino acids glutamate and arginine to in vitro cultured Atlantic salmon muscle cells has previously been found to influence gene expression of both early and late muscle-specific markers, including myogenin, Myf5, MyoD1a, Myf6 and myosin light chain27.

Relative gene expression of the muscle regulatory factors myf5 (A) and myod1a (B) and the muscle structural protein mhc (C) upon 72 h of incubation with cell culture media supplemented with different CH fractions (F4 and F6). myf5: Myogenic factor 5; myod1A: Myoblast determination protein 1A; mhc: Myosin heavy chain; Relative gene expression (RGE); CTL: control, F4; CH fraction 4; F6: CH fraction F6. Results presented with box plots (min to max with line at mean, n = 3). Different letters denote statistical differences (P < 0.05).
Characterization of bioactive fraction
The two low-molecular weight fractions of CH that showed the highest effect on viability (F4 and F6) and proliferation (F6) of Atlantic salmon muscle cells were further analyzed to study the molecular weight distribution of the peptides. Compared to crude CH, both F4 and F6 were shown to constitute relatively low amounts of early-eluting large proteins and high amounts of late-eluting smaller molecular weight peptides (Fig. 6). Average molecular weights of peptides in the fractions were calculated to be 407 and 260 g/mol for F4 and F6, respectively, which approximately correspond to peptides with an average length of 3.6 and 2.3 amino acid residues (calculated using a weighted average molecular weight of 113 g/mol for single amino acids). These results indicate that the growth-promoting effects observed in the feeding trial and in vitro studies can potentially be attributed to low-molecular weight peptides in CH, which are known to have efficient uptake by peptide transporters in the intestines39 and improve growth performance34.

Molecular weight distribution of crude CH and fractions F4 and F6 acquired using size-exclusion chromatography with UV monitoring (214 nm).
Identified as the most bioactive fraction in terms of viability, proliferation, and gene expression of in vitro cultured muscle cells, F6 was further characterized with 1H NMR spectroscopy (Fig. 7). Tentative assignments of the proton NMR peaks were performed according to published chemical shift values for closely related samples40,41,42. In addition to characteristic signals assigned to side chains of aromatic and aliphatic amino acids, significantly large signals were assigned to common marine metabolites such as trimethylamine N-oxide (TMAO) and dimethyl glycine (DMG). The abundance of DMG in combination with the branched-chain amino acids (BCAAs) was particularly interesting. Supplementation of DMG-salt in diets has been shown to improve growth performance and skeletal muscle function in terrestrial animals43,44 and inclusion of dietary BCAAs in aquafeeds to contribute to growth and flesh quality45. Glutamate and arginine, the aforementioned amino acids found to increase the expression of muscle-specific genes27, were also identified as part of several abundant peaks in the fraction. Lactate is yet another metabolite identified in the fraction that has been associated with growth stimulation. It has been reported to play a role in the exercise-induced human growth hormone response46, to stimulate hypertrophy and regeneration of mouse skeletal muscle47, and to stimulate growth when supplemented to the diet of Arctic charr (Salvelinus alpinus)48.

1H NMR spectra of CH-F6 with tentative assignments based on previously reported chemical shift values. Assignments were done for the abundant peaks, and only peaks diagnostic to a given molecule were assigned. Unassigned peaks are marked with “*”.
Conclusions
Dietary inclusion of calanus hydrolysate (CH) in a feeding trial with European sea bass juveniles led to increased growth performance and feed efficiency compared with other commercial marine hydrolysates, highlighting the potential of CH as a functional ingredient in fish feed. In the following in vitro study, low-molecular weight fractions of CH were associated with increased viability and proliferative activity in a salmon skeletal muscle cell model. Furthermore, a gene expression experiment showed downregulation of early muscle markers and a trend towards higher expression of a structural muscle marker, which may indicate a positive effect on muscle differentiation. Characterization of the most bioactive fraction by 1H NMR revealed that it contained small peptides and important amino acids, but also metabolites previously found to be associated with increased muscle growth. Further studies are required to quantify the direct contribution of such metabolites to the observed growth-promoting effects. To the authors knowledge, the present study is the first to demonstrate the promising potential of CH as a feed ingredient with specific growth-promoting constituents.
Materials and methods
Marine hydrolysate test products
Calanus finmarchicus raw material was harvested and processed into calanus hydrolysate (CH) by Calanus AS (Tromsø, Norway), and the commercial hydrolysates from salmon (SH), sardine (SDH), and tuna fish (TH) were bought and supplied by SPAROS, Lda. (Olhão, Portugal). Calanus and salmon hydrolysates were used in liquid form, while those of sardine and tuna were dry powders. The nutrient composition of the test hydrolysates is presented in Table 1.
Formulation of experimental diets
The four experimental diets in the trial were formulated with low levels of fish meal and wheat gluten, compensated with the marine hydrolysate test products (calanus, salmon, sardine, and tuna fish) and additional plant proteins (Table 2). To make all the diets equivalent in terms of protein supply, the inclusion level of the hydrolysates ranged from 2.95% to 4.20% (dry basis) based on their different crude protein content (Table 1). Overall, the formulated diets were isonitrogenous (48% crude protein), isolipidic (16% crude fat) and isoenergetic (21.9 MJ/kg).
Manufactured diets
Diets were produced by extrusion at SPAROS. All powder ingredients were mixed according to the target formulation in a double-helix mixer (model RM90, MAINCA, Spain) and ground (below 250 µm) in a micropulverizer hammer mill (model SH1, Hosokawa-Alpine, Germany). Test hydrolysates from calanus and salmon, since presented in the liquid form, were incorporated in the feed mash after grinding. Diets (pellet sizes: 1.5 and 2.0 mm) were manufactured with a twin-screw extruder (model BC45, Clextral, France) with a screw diameter of 55.5 mm. Extrusion conditions were as follows: Feeder rate (44 kg/h), screw speed (142 rpm), water addition (approximately 290 ml/min), temperature barrel 1 (24–29 °C), temperature barrel 3 (105–107 °C). Extruded pellets were dried in a vibrating fluid bed dryer (model DR100, TGC Extrusion, France). After cooling, oils were added post-extrusion by vacuum coating (model PG-10VCLAB, Dinnissen, The Netherlands). Completed pellets were packed in sealed plastic buckets and shipped to the research site where they were stored at room temperature, in a cool and well-aerated environment. Samples of each diet were taken for analytical characterization (Table 3, Tables S1-S2 in supplementary data). Analysis of complete feeds were carried out with analytical duplicates, following the methodology described by AOAC49 for most analytics: Dry matter after drying at 105 °C for 24 h; total ash by combustion (550 °C, 6 h) in a muffle furnace (Nabertherm L9/11/B170, Germany); crude protein (N × 6.25) by a flash combustion technique followed by gas chromatographic separation and thermal conductivity detection with a Leco N analyser (Model FP-528, Leco Corporation, USA); crude lipid by petroleum ether extraction (40–60 °C) using a Soxtherm system (Multistat SX PC, Gerhardt, Germany); gross energy in an adiabatic bomb calorimeter (Werke C2000, IKA, Germany). Amino acids were determined after hydrolysis in 6 M HCL at 108 °C for 24 h in nitrogen-flushed vials, using a Waters Pico-Tag reversed-phase HPLC system with norleucine as an internal standard. The resulting chromatograms were analyzed with Breeze software (Waters, USA). Tryptophan was not analyzed due to its destruction by acid hydrolysis. Minerals and fatty acids were analyzed at a certified laboratory (Eurofins Anàlisis Alimentario SL, Spain), according to ISO 27,085:2009 by ICP-AES methodology50 for minerals, and by gas-chromatography and flame ionization detection (GC-FID) for fatty acids.
Ethical statement
The feeding trial was conducted at the experimental facilities of SPAROS (Olhão, Portugal), which are registered for experimentation with aquatic species by Direção-Geral de Alimentação e Veterinária (Ministry of Agriculture, Portugal). The experimental protocol was approved by the Animal Welfare Committee—Órgão Responsável pelo Bem-Estar Animal (ORBEA) of SPAROS (Trial reference: CALBASS). Experiments were conducted by FELASA certified scientists and technical staff, in full compliance with ARRIVE guidelines and the European (Directive 2010/63/EU) and Portuguese (Decreto-Lei nº. 113/2013, August 7th) legislation on the protection of animals for scientific purposes.
Feeding trial
The trial was performed at the experimental research facilities of SPAROS (Olhão, Portugal), with European sea bass (Dicentrarchus labrax) sourced at Estação Piloto de Piscicultura de Olhão (EPPO) from the Portuguese Institute of the Sea and Atmosphere (IPMA). Approximately 650 fish were transferred to the SPAROS experimental facilities by a duly authorized carrier and no significant mortality or pathological signs were observed in association with transport. The fish were kept in sanitary quarantine for three weeks, during which the fish were maintained and fed a standard commercial diet, before the start of the trial.
A total of 360 fish were distributed into 12 tanks (30 fish per tank), with three tanks for each of the experimental diets. Four experimental diets were formulated (Table 2), each with the inclusion of a marine hydrolysate (calanus, sardine, tuna, or salmon). The mean initial body weight of the fish was 8.9 ± 0.4 g, and they were fed the experimental diets for 84 days. The sub-square PVC tanks contained 90 L of water and were located indoors, supplied with flow-through saltwater (flow rate: 4.6 L/min), and subjected to photoperiod conditions of 12 h light and 12 h dark. The average water temperature during the trial was 23.0 ± 1.4 °C, salinity was 35.8 ± 0.5 ‰, dissolved oxygen levels in the water were kept above 5.3 mg/L, and ammonia levels were below the detection limit.
Fish were hand-fed to visual satiety in four daily meals at 09:00, 12:00, 15:00, and 18:00. Utmost care was taken to avoid feed wastage and allow precise quantification of intake. Lightly anesthetized (20 mL/L of AQUI-S, New Zealand) fish were group weighed at the start, day 30, day 67, and finally at day 84. Feed intake was calculated, survival was monitored daily, and growth performance was recorded on the abovementioned days. Feed conversion ratio (FCR), protein efficiency ratio (PER), and specific growth rate (SGR) were calculated (n = 3) upon completion of the trial using these formulas:
-
FCR: crude feed intake/weight gain.
-
PER: wet weight gain/crude protein intake.
-
SGR: (Ln final body weight – Ln initial body weight) × 100/days.
Clean-up and fractionation of calanus hydrolysates
Crude calanus hydrolysate (CCH, 46% dry matter) was freeze-dried to afford a light brown powder. To further refine peptides from the crude hydrolysate, CCH was diluted 1:1 in Milli-Q water, filtered using Millex syringe filter with a PVDF membrane (pore size 0.45 μm, Merk Millipore, Burlington, MA, USA), and desalted using semi-preparative chromatography. Fractionations were performed using a Dionex Ultimate 3000 instrument (Thermo Scientific, Waltham, MA, USA) equipped with a quaternary pump and thermometer auto-sample, and a variable wavelength UV-detector. Desalting was performed using a reversed-phase Thermo Betasil C18 column, 250 × 10 mm i.d., 10 μm particle size (Thermo Scientific). The mobile phase used was H2O (Solvent A) and acetonitrile (Solvent B), both acidified with 0.1% formic acid. The following elution profile was used at a flow rate of 2 mL/min: 0 min, 0% B; 10.0 min, 0% B; 35.0 min, 100% B; 41.0 min, 100% B; 51.0 min, 0% B. The desalted calanus hydrolysate (DCH) fractions were collected between 5.2 min and 35.0 min retention time and freeze-dried to afford light yellow powder. Both freeze-dried CCH and DCH were directly used for the initial screening in the cell culture assays. DCH was further fractionated using size exclusion chromatography (SEC). Fractionation based on molecular weight was performed using the same system described above and a preparative Yarra SEC-2000 column, 300 × 21.1 mm i.d., 5 µm particle size (Phenomenex, Torrence, CA, USA). The mobile phase consisted of a mixture of acetonitrile and ultrapure water in a proportion of 30:70 (v/v), containing 0.1% formic acid. CCH diluted 1:1 in Milli-Q water filtered using Millex syringe filter with a PVDF membrane was used as an injection solution. The chromatographic separation was performed using isocratic elution with 30% acetonitrile (0.1% formic acid) at a flow rate of 3 mL/min from 0 to 55 min. This was followed by washing for 3 min with NaH2PO4 (0.10 M) and subsequent column equilibration of 20 min with 30% acetonitrile. Fractions were collected from a total of 15 injections of 500 μL. A total of six fractions CH-F1 to CH-F6 were collected, pooled, and freeze-dried prior to analysis with cell cultures. The active fractions (F4 and F6) identified from the cell culture study were analysed for molecular weight distribution using the protocol by Wubshet et al.51.
Isolation and culturing of primary Atlantic salmon skeletal muscle cells
Atlantic salmon primary skeletal muscle cells were isolated according to Østbye and Ytteborg 28. The isolated muscle cells were seeded in T25 cell flasks (muscle tissue from eight 5 cm salmon per T25 cell flask) with growth media containing 10% fetal bovine serum (FBS), 0.01 M HEPES (Sigma-Aldrich, St. Louis, USA), 10 mL/L antibiotic–antimycotic (Sigma-Aldrich, St. Louis, USA), in L-15 medium GlutaMAX (Invitrogen, Carlsbad, USA). The cells were incubated at 13 °C without CO2 for 2 days. The cells were then trypsinated and transferred to 96-well plates for viability or proliferation assays or to 6-well plates for gene expression analysis. After one day incubation at 13 °C without CO2, the cells were added growth medium containing 2% FBS and the different concentrations of calanus hydrolysate. The cells were incubated with hydrolysates for 72 h before harvesting to the different analyses.
Two separate cell trials were conducted to investigate the effect of calanus hydrolysate on Atlantic salmon skeletal muscle growth. In the first cell trial, three concentrations (1.00, 0.10 and 0.01 mg/mL, pH7) of crude and desalted calanus hydrolysates were tested on viability, toxicity, and proliferation of cells. In the second cell trial, six different size separated fractions (F1-F6) of the desalted hydrolysate (0.10 mg/mL) were investigated by viability and proliferation assays. Only F4 and F6 were included for the gene expression analysis.
Cell viability, toxicity, and proliferation
Viability of the cells was evaluated by measuring ATP production with CelltiterGlo (Promega Corporation, Madison, WI, USA), potential toxicity of the hydrolysates using Cytotoxicity detection kit (LDH) (Roche Diagnostics GmbH, Mannheim, Germany), and proliferation of the cells using CyQUANT Cell Proliferation Assay (Molecular Probes, Oregon, USA), following the manufacturer’s protocols.
Gene expression analysis
RNeasy kit (Qiagen, Valencia, CA, USA) was used to isolate total RNA from the muscle cells following the producer’s protocol. All samples were treated with DNase I (Invitrogen, Carlsbad, USA) to remove genomic DNA. The concentration and purity of the RNA were evaluated using NanoDrop 1000 Spectrophotometer (NanoDrop Technologies, USA). Taqman reverse transcriptase reagents (Applied Biosystems, Foster City, CA, USA) were used to synthesize cDNA from 500 ng RNA in a 20 μL reaction volume according to the following conditions: 25 °C for 10 min, 37 °C for 30 min, and 95 °C for 5 min. The qPCR reaction mixture consisted of 4 μl diluted (1:10) cDNA, 1 μl forward and reverse primer at a final concentration of 0.5 μM (Table S8 in supplementary data), and 5 μl PowerUp SYBR Green Master Mix (Applied Biosystems, Foster City, California, United States). A standard curve was included for each primer pair to evaluate the primer efficiency, and a melting curve analysis was included to verify the amplification of only one product. All samples were analyzed in parallel, and non-template and non-enzyme controls were included. The qPCR reaction was run on a QuantStudio 5 instrument (Thermo Fisher Scientific, MA, USA) under the following conditions: 50 °C for 20 s, 95 °C for 20 s, 40 cycles with 95 °C for 1 s and 60 °C for 20 s, 95 °C for 1 s and 60 °C for 20 s, 95 °C for 1 s. RefFinder52 was used to evaluate the most stable reference gene of ef1a, rpol2 and etif3. The relative gene expression level was calculated according to the ΔΔCt method with efficiency correction53 using rpol2 as a reference gene.
NMR experiment
1H NMR analysis of the promising fraction (F6) was performed using a Bruker Ascend system (1H operating frequency of 400 MHz) equipped with a sample changer and a gradient inverse triple-resonance 5 mm probe-head (Bruker Biospin). NMR sample was prepared by dissolving appx. 5 mg of CH-F6 in 0.7 mL D2O containing 0.05 wt. % TMSP-d4 sodium salt as an internal standard. IconNMR (version 4.2, Bruker Biospin) was used for controlling automated acquisition of NMR data (temperature equilibration to 300 K, optimization of lock parameters, gradient shimming, and setting of receiver gain), and processing of NMR data was performed using Topspin (version 3.0, Bruker Biospin). The one-dimensional 1H NMR spectra were acquired using 30° pulses, 4 s acquisition time, 1 s relaxation delay and 32 K data points. The data points were zero-filled to 64 K and multiplied with an exponential function (line-broadening = 0.3 Hz) prior to Fourier transformation.
Statistical analysis
Evaluation criteria data of the feeding trial are presented as means ± standard deviation, with n = 3 replicates. Data from the bioactivity studies with skeletal muscle cells are presented with box-plots (n = 3–5 replicates), to better represent the distribution of these specific data. All data were normally distributed with equal variances (Table S7.1–S7.3 in supplementary data). Starting weight of the fish in the different diet groups were similar, allowing a one-way analysis of variance (ANOVA) for comparison of growth parameters at different time points. One-way ANOVA was also performed for all the bioactivity studies with skeletal muscle cells. When appropriate, means were compared with the Tukey method. Statistical significance was tested at a 0.05 probability level. All statistical tests were performed using GraphPad Prism (version 9.2.0).
Data availability
The gene expression datasets generated and/or analyzed during the current study are available in the DataverseNO repository for open research data (https://doi.org/10.18710/MRVDNS), all other data can be found in the supplementary material or from the author upon request.
Abbreviations
- BCAA:
-
Branched-chain amino acid
- CH:
-
Calanus hydrolysate
- CCH:
-
Crude calanus hydrolysate
- CTL:
-
Control
- DCH:
-
Desalted calanus hydrolysate
- DMH:
-
Dimethyl glycine
- EPH:
-
Enzymatic protein hydrolysis
- F1–F6:
-
Fraction 1–6 of calanus hydrolysate
- FCR:
-
Feed conversion ratio
- MHC:
-
Myosin heavy chain
- Myf5:
-
Myogenic factor 5
- MyoD1a:
-
Myoblast determination protein 1a
- NMR:
-
Nuclear resonance magnetic spectroscopy
- PER:
-
Protein efficiency ratio
- RFU:
-
Relative fluorescence units
- SH:
-
Salmon hydrolysate
- SDH:
-
Sardine hydrolysate
- SEC:
-
Size-exclusion chromatography
- SGR:
-
Specific growth rate
- TH:
-
Tuna fish hydrolysate
References
United Nations, Department of economic and social affairs, population division. World population prospects 2019: Highlights. Report No. ST/ESA/SER.A/423, (2019).
Hoegh-Guldberg, O. et al. The Ocean as a solution to climate change: Five opportunities for action (World Resources Institute, 2019).
FAO. The state of world fisheries and aquaculture. Sustainability in action. (2020).
Henchion, M., Hayes, M., Mullen, A. M., Fenelon, M. & Tiwari, B. Future protein supply and demand: strategies and factors influencing a sustainable equilibrium. Foods 6, 53. https://doi.org/10.3390/foods6070053 (2017).
Costello, C. et al. The future of food from the sea. Nature 588, 95–100. https://doi.org/10.1038/s41586-020-2616-y (2020).
Hua, K. et al. The future of aquatic protein: Implications for protein sources in aquaculture diets. One Earth 1, 316–329. https://doi.org/10.1016/j.oneear.2019.10.018 (2019).
Egerton, S. et al. Replacing fishmeal with plant protein in Atlantic salmon (Salmo salar) diets by supplementation with fish protein hydrolysate. Sci. Rep. 10, 4194. https://doi.org/10.1038/s41598-020-60325-7 (2020).
Jingting, Y., Danting, G., Chun, K., Min, J. & Xueming, H. Effect of soybean antigenic protein on feed palatability of fishmeal replaced diets for obscure puffer (Takifugu fasciatus) and the alternation of diet preference by domestication. Aquaculture Rep. 17, 100332. https://doi.org/10.1016/j.aqrep.2020.100332 (2020).
Oliveira, M. & Vasconcelos, V. Occurrence of mycotoxins in fish feed and its effects: A review. Toxins (Basel) 12, 160. https://doi.org/10.3390/toxins12030160 (2020).
Skjoldal, H. R. The Norwegian Sea ecosystem (Tapir Academic Press, 2004).
Fjeld, K., Tiller, R., Grimaldo, E., Grimsmo, L. & Standal, I.-B. Mesopelagics-New gold rush or castle in the sky?. Marine Policy 147, 105359. https://doi.org/10.1016/j.marpol.2022.105359 (2023).
Eysteinsson, S. T., Gudjónsdóttir, M., Jónasdóttir, S. H. & Arason, S. Review of the composition and current utilization of Calanus finmarchicus – Possibilities for human consumption. Trends Food Sci. Technol. 79, 10–18. https://doi.org/10.1016/j.tifs.2018.06.019 (2018).
Albrektsen, S. et al. Future feed resources in sustainable salmonid production: A review. Rev. Aquac. 14, 1790–1812. https://doi.org/10.1111/raq.12673 (2022).
Siddik, M. A. B., Howieson, J., Fotedar, R. & Partridge, G. J. Enzymatic fish protein hydrolysates in finfish aquaculture: A review. Rev. Aquac. 13, 406–430. https://doi.org/10.1111/raq.12481 (2021).
Sánchez, A. & Vázquez, A. Bioactive peptides: A review. Food Quality Safety 1, 29–46. https://doi.org/10.1093/fqsafe/fyx006 (2017).
Martínez-Alvarez, O., Chamorro, S. & Brenes, A. Protein hydrolysates from animal processing by-products as a source of bioactive molecules with interest in animal feeding: A review. Food Res. Int. 73, 204–212. https://doi.org/10.1016/j.foodres.2015.04.005 (2015).
Vang, B., Pedersen, A. M. & Olsen, R. L. Oil extraction from the copepod Calanus Finmarchicus using proteolytic enzymes. J. Aquat. Food Prod. Technol. 22, 619–628. https://doi.org/10.1080/10498850.2012.686008 (2013).
Klompong, V., Benjakul, S., Kantachote, D. & Shahidi, F. Antioxidative activity and functional properties of protein hydrolysate of yellow stripe trevally (Selaroides leptolepis) as influenced by the degree of hydrolysis and enzyme type. Food Chem. 102, 1317–1327. https://doi.org/10.1016/j.foodchem.2006.07.016 (2007).
Berge, G. M. & Storebakken, T. Fish protein hydrolyzate in starter diets for Atlantic salmon (Salmo salar) fry. Aquaculture 145, 205–212. https://doi.org/10.1016/S0044-8486(96)01355-5 (1996).
Resende, D. et al. Innovative swine blood hydrolysates as promising ingredients for European seabass diets: Impact on growth performance and resistance to Tenacibaculum maritimum infection. Aquaculture 561, 738657. https://doi.org/10.1016/j.aquaculture.2022.738657 (2022).
Colombo-Hixson, S. M., Olsen, R. E., Tibbetts, S. M. & Lall, S. P. Evaluation of Calanus finmarchicus copepod meal in practical diets for juvenile Atlantic halibut (Hippoglossus hippoglossus). Aquac. Nutr. 19, 687–700. https://doi.org/10.1111/anu.12016 (2013).
Torrecillas, S., Montero, D., Carvalho, M., Benitez-Santana, T. & Izquierdo, M. Replacement of fish meal by Antarctic krill meal in diets for European sea bass Dicentrarchus labrax: Growth performance, feed utilization and liver lipid metabolism. Aquaculture 545, 737166. https://doi.org/10.1016/j.aquaculture.2021.737166 (2021).
Bakke, S. et al. Dietary protein hydrolysates and free amino acids affect the spatial expression of peptide transporter PepT1 in the digestive tract of Atlantic cod (Gadus morhua). Comp. Biochem. Physiol. B: Biochem. Mol. Biol. 156, 48–55. https://doi.org/10.1016/j.cbpb.2010.02.002 (2010).
Zheng, K., Liang, M., Yao, H., Wang, J. & Chang, Q. Effect of size-fractionated fish protein hydrolysate on growth and feed utilization of turbot (Scophthalmus maximus L.). Aquaculture Res. 44, 895–902. https://doi.org/10.1111/j.1365-2109.2012.03094.x (2013).
Johnston, I. A. Environment and plasticity of myogenesis in teleost fish. J. Exp. Biol. 209, 2249–2264. https://doi.org/10.1242/jeb.02153 (2006).
Østbye, T.-K.K. et al. Molecular cloning of the Atlantic salmon activin receptor IIB cDNA – Localization of the receptor and myostatin in vivo and in vitro in muscle cells. Comp. Biochem. Physiol. D: Genomics Proteomics 2, 101–111. https://doi.org/10.1016/j.cbd.2006.12.003 (2007).
Østbye, T.-K.K. et al. Functional amino acids stimulate muscle development and improve fillet texture of Atlantic salmon. Aquac. Nutr. 24, 14–26. https://doi.org/10.1111/anu.12528 (2018).
Østbye, T.-K.K. & Ytteborg, E. Preparation and culturing of Atlantic salmon muscle cells for in vitro studies. Methods Mol. Biol. 319–330, 2019. https://doi.org/10.1007/978-1-4939-8897-6_19 (1889).
Sánchez-Gurmaches, J. et al. In vivo and in vitro insulin and fasting control of the transmembrane fatty acid transport proteins in Atlantic salmon (Salmo salar). Am. J. Phys.. Regul. Integr. Comp. Phys. 301, R947–957. https://doi.org/10.1152/ajpregu.00289.2011 (2011).
Vegusdal, A., Ostbye, T. K., Tran, T. N., Gjøen, T. & Ruyter, B. Beta-oxidation, esterification, and secretion of radiolabeled fatty acids in cultivated Atlantic salmon skeletal muscle cells. Lipids 39, 649–658. https://doi.org/10.1007/s11745-004-1278-3 (2004).
Argyropoulos, C. et al. Hypertonicity: Pathophysiologic concept and experimental studies. Cureus 8, e596. https://doi.org/10.7759/cureus.596 (2016).
Davis, D. A. & Gatlin, D. M. Dietary mineral requirements of fish and marine crustaceans. Rev. Fish. Sci. 4, 75–99. https://doi.org/10.1080/10641269609388579 (1996).
Ishak, N. H. & Sarbon, N. M. A review of protein hydrolysates and bioactive peptides deriving from wastes generated by fish processing. Food Bioprocess Technol. 11, 2–16. https://doi.org/10.1007/s11947-017-1940-1 (2018).
Rashidian, G., Abedian Kenari, A. & Nikkhah, M. Dietary effects of a low-molecular weight fraction (10 kDa) from shrimp waste hydrolysate on growth performance and immunity of rainbow trout (Oncorhynchus mykiss) Employing nanodelivery systems. Fish Shellfish Immunol. 118, 294–302. https://doi.org/10.1016/j.fsi.2021.09.014 (2021).
Lima, R. D. C. L. et al. Peptides from chicken processing by-product inhibit DPP-IV and promote cellular glucose uptake: Potential ingredients for T2D management. Food Funct. 10, 1619–1628. https://doi.org/10.1039/C8FO02450B (2019).
Andreassen, R. C., Pedersen, M. E., Kristoffersen, K. A. & Beate Rønning, S. Screening of by-products from the food industry as growth promoting agents in serum-free media for skeletal muscle cell culture. Food Funct. 11, 2477–2488. https://doi.org/10.1039/C9FO02690H (2020).
Braun, T., Buschhausen-Denker, G., Bober, E., Tannich, E. & Arnold, H. H. A novel human muscle factor related to but distinct from MyoD1 induces myogenic conversion in 10T1/2 fibroblasts. Embo. J. 8, 701–709. https://doi.org/10.1002/j.1460-2075.1989.tb03429.x (1989).
Davis, R. L., Weintraub, H. & Lassar, A. B. Expression of a single transfected cDNA converts fibroblasts to myoblasts. Cell 51, 987–1000. https://doi.org/10.1016/0092-8674(87)90585-x (1987).
Wang, J., Yan, X., Lu, R., Meng, X. & Nie, G. Peptide transporter 1 (PepT1) in fish: A review. Aquac. Fish. 2, 193–206. https://doi.org/10.1016/j.aaf.2017.06.007 (2017).
Hansen, B. H., Degnes, K., Øverjordet, I. B., Altin, D. & Størseth, T. R. Metabolic fingerprinting of arctic copepods Calanus finmarchicus, Calanus glacialis and Calanus hyperboreus. Polar Biol. 36, 1577–1586. https://doi.org/10.1007/s00300-013-1375-8 (2013).
Steinsholm, S. et al. Sensory assessment of fish and chicken protein hydrolysates. Evaluation of NMR metabolomics profiling as a new prediction tool. J. Agric. Food Chem. 68, 3881–3890. https://doi.org/10.1021/acs.jafc.9b07828 (2020).
Sundekilde, U. K., Jarno, L., Eggers, N. & Bertram, H. C. Real-time monitoring of enzyme-assisted animal protein hydrolysis by NMR spectroscopy – An NMR reactomics concept. LWT 95, 9–16. https://doi.org/10.1016/j.lwt.2018.04.055 (2018).
Bai, K. et al. Dietary dimethylglycine sodium salt supplementation improves growth performance, redox status, and skeletal muscle function of intrauterine growth-restricted weaned piglets. J. Animal Sci. https://doi.org/10.1093/jas/skab186 (2021).
Chalvatzi, S. et al. Dimethylglycine supplementation in reduced energy broilers’ diets restores performance by improving nutrient digestibility. Animals 10, 789 (2020).
Ahmad, I., Ahmed, I., Fatma, S. & Peres, H. Role of branched-chain amino acids on growth, physiology and metabolism of different fish species: A review. Aquac. Nutr. 27, 1270–1289. https://doi.org/10.1111/anu.13267 (2021).
Godfrey, R. J., Whyte, G. P., Buckley, J. & Quinlivan, R. The role of lactate in the exercise-induced human growth hormone response: Evidence from McArdle disease. Br. J. Sports Med. 43, 521–525. https://doi.org/10.1136/bjsm.2007.041970 (2009).
Ohno, Y. et al. Lactate stimulates a potential for hypertrophy and regeneration of mouse skeletal muscle. Nutrients https://doi.org/10.3390/nu11040869 (2019).
Ringø, E. Effects of dietary lactate and propionate on growth and digesta in Arctic charr, Salvelinus alpinus (L.). Aquaculture 96, 321–333. https://doi.org/10.1016/0044-8486(91)90161-Y (1991).
AOAC. Official methods of analysis. 18th ed., Rev 1 edn, (Association of Official Analytical Chemists International, Gaithersburg, Maryland, US., 2006).
ISO. ISO 27085:2009 Animal feeding stuffs. Determination of calcium, sodium, phosphorus, magnesium, potassium, iron, zinc, copper, manganese, cobalt, molybdenum, arsenic, lead and cadmium by ICP-AES. 24 pp (International Organization for Standardization, Geneva, 24pp., 2009).
Wubshet, S. G. et al. FTIR as a rapid tool for monitoring molecular weight distribution during enzymatic protein hydrolysis of food processing by-products. Anal. Methods 9, 4247–4254. https://doi.org/10.1039/C7AY00865A (2017).
Xie, F., Xiao, P., Chen, D., Xu, L. & Zhang, B. miRDeepFinder: A miRNA analysis tool for deep sequencing of plant small RNAs. Plant. Mol. Biol. 80, 75–84. https://doi.org/10.1007/s11103-012-9885-2 (2012).
Pfaffl, M. W. in A–Z of Quantitative PCR, pp. 87–120. (ed Stephen A. Bustin) Ch. 87–120, (IUL Biotechnology Series, International University Line, 2004).
Acknowledgements
This study was supported by the Norwegian Research Council project “Benefits of calanus protein products in feed and food” (grant number 312136), TailoTides (grant number 320086), and by the MABIT project “Identification and characterization of growth-promoting peptides from Calanus® Hydrolysate” (grant number UB0077). The publication charges for this article have been funded by a grant from the publication fund of UiT The Arctic University of Norway. Special thanks to Vibeke Voldvik and Nina Solberg at Nofima and engineers at SPAROS for their contribution to lab work, fish handling, sampling, and analyses.
Funding
Open access funding provided by UiT The Arctic University of Norway (incl University Hospital of North Norway).
Author information
Authors and Affiliations
Contributions
Every author each contributed to various parts of the conception and design of the study; IB/JD/AMP/KEE for the feeding trial, and IB/SW/SBR/TKØ/AMP/KEE for the bioactivity studies. JD with engineers at SPAROS performed the feeding trial, and TKØ/SW/SBR with engineers at Nofima performed the bioactivity laboratory studies. IB/SW/TKØ wrote the first draft of the manuscript, and all authors contributed with reviewing, editing, and finalizing the manuscript.
Corresponding author
Ethics declarations
Competing interests
Isak Bøgwald is an industrial PhD student at UiT and Calanus AS, and Alice Marie Pedersen holds a position at Calanus AS. Tone-Kari K. Østbye, Sissel Beate Rønning, Jorge Dias, Karl-Erik Eilertsen, and Sileshi Gizachew Wubshet declare no commercial or financial relationships that could be construed as a potential conflict of interest. Calanus AS acquired funding, and SPAROS and Nofima performed practical executions of the feeding trial and bioactivity laboratory studies respectively.
Additional information
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Bøgwald, I., Østbye, TK.K., Pedersen, A.M. et al. Calanus finmarchicus hydrolysate improves growth performance in feeding trial with European sea bass juveniles and increases skeletal muscle growth in cell studies. Sci Rep 13, 12295 (2023). https://doi.org/10.1038/s41598-023-38970-5
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-023-38970-5
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
