
Abstract
Dietary supplementations with n-3 polyunsaturated fatty acid (PUFA) have been explored in autism spectrum disorder (ASD) but their efficiency and potential in ameliorating cardinal symptoms of the disease remain elusive. Here, we compared a n-3 long-chain (LC) PUFA dietary supplementation (n-3 supp) obtained from fatty fish with a n-3 PUFA precursor diet (n-3 bal) obtained from plant oils in the valproic acid (VPA, 450 mg/kg at E12.5) ASD mouse model starting from embryonic life, throughout lactation and until adulthood. Maternal and offspring behaviors were investigated as well as several VPA-induced ASD biological features: cerebellar Purkinje cell (PC) number, inflammatory markers, gut microbiota, and peripheral and brain PUFA composition. Developmental milestones were delayed in the n-3 supp group compared to the n-3 bal group in both sexes. Whatever the diet, VPA-exposed offspring did not show ASD characteristic alterations in social behavior, stereotypies, PC number, or gut microbiota dysbiosis while global activity, gait, peripheral and brain PUFA levels as well as cerebellar TNF-alpha levels were differentially altered by diet and treatment according to sex. The current study provides evidence of beneficial effects of n-3 PUFA based diets, including one without LCPUFAs, on preventing several behavioral and cellular symptoms related to ASD.
Similar content being viewed by others

Introduction
Autism Spectrum Disorder (ASD) is a neurodevelopmental disorder characterized by persistent deficits in communication and social interactions, and restricted repetitive behaviors, interests, or activities1. Imaging and post-mortem studies in ASD patients have identified the cerebellum as one of the major affected brain regions with reduction of its volume and decreased Purkinje cell (PC) number2. Similar findings were observed in both genetic and environmental animal models, where the cerebellar regions crus I and crus II involved in motor and cognitive functions were shown to be the most affected by PC loss, with sex differences3,4,5. Many comorbidities are also associated with ASD including inflammation, gastrointestinal and eating disorders6,7. There is currently no cure or preventive strategies for this disease other than symptomatologic. Polyunsaturated fatty acids (PUFAs) from the n-6 and n-3 families are found in large quantities in brain cell membranes and are crucial for brain function and development8,9. Linoleic acid (LA, n-6) and alpha-linolenic acid (ALA, n-3) are essential fatty acids (FA) that cannot be synthetized by mammals and need to be provided through the diet. LA and ALA are then metabolized into long-chain (LC) PUFA, arachidonic acid (AA, n-6) and eicosapentaenoic acid (EPA, n-3)-docosahexaenoic acid (DHA, n-3), respectively. However, as ALA is not sufficient to supply the brain in n-3 LCPUFAs, additional dietary source from fatty fish is necessary10. Low levels of n-3 PUFA have been reported in the blood and brain of patients diagnosed with ASD11,12,13,14,15. As PUFAs from maternal diet are crucial for the developing brain during the perinatal period9, insufficient n-3 PUFA dietary supply during this critical period leads to aberrant brain lipid composition, metabolism, and signaling pathways16,17. This, in turn may be associated with neurodevelopmental and psychiatric disorders8,18,19. Thus, n-3 PUFA dietary supplementation, during the perinatal period for prevention and at adulthood for correction of or prevention from ASD have been explored in clinical and animal settings20,21,22,23,24,25,26.
While several studies point to potential benefits of n-3 PUFA supplementation during the perinatal period, the exact contribution of n-3 PUFAs species (i.e., precursor versus long-chain) on neurodevelopmental disorders are not well defined. This, along with the conflicting results obtained in clinical ASD studies, prompt us to perform a large scale and a side-by-side comparison of two diets. The first is a balanced diet (n-3 bal) with ALA as the only n-3 PUFA source from plant oils. The second is a n-3 LCPUFA supplemented diet (n-3 supp) with DHA and EPA from fatty fish. Both diets were isocaloric and differed in their fatty acid composition. Comparisons were performed in the valproic acid (VPA) mouse model of ASD, in males and females analyzed separately. Indeed, environmental factors acting in utero are suspected to contribute to the etiology of ASD27. Among these, VPA exposure during pregnancy has been consistently shown to be a major factor associated with developmental defects increasing the risk to develop ASD28. The VPA model has proven to have strong construct, face, and more recently, predictive validity29,30,31. Injection of VPA to pregnant rodent females systematically induces ASD-related syndromes in the offspring such as impaired social interaction32,33,34, repetitive behavior34 and delayed motor development35 (see also our reviews on this subject2,36). In this line, and prior to the current study, we had recently performed a full-scale analysis of this ASD mouse model on a wide range of behaviors with molecular and cellular correlates from early postnatal age to adulthood. A special focus was made on motor behavior and cerebellar implications under a standard diet containing comparable LA/ALA ratio (5.3) than the diets used here (6.2)3. We showed major social deficits, stereotypies and cerebellar motor and gait disorders in the VPA ASD mouse model which were correlated with cerebellar regional PC loss in crus I and crus II. This allowed us to obtain a solid starting point to investigate n-3 LCPUFA supplementation effects on the ASD phenotype. Here, we extend our analysis to inflammation and gut microbiome, to determine their implication in dam and in male and female offspring behavioral, cellular, and metabolic responses to differential diets.
Results
Maternal behavior is not affected by either VPA treatment or diet
VPA-exposed juvenile mice produce aberrant patterns of isolation stress-induced ultrasonic vocalizations37. This may influence in turn maternal behavior, which is key to normal social and motor development of offspring38. Here, we assessed various parameters related to maternal behavior following pup separation in relation with treatment and diet (Suppl. Fig. S1). Two-way ANOVA analysis indicated no differences between experimental groups, regardless of the treatment or diet on maternal care, nesting and stress related behaviors. In summary, our results indicate that VPA treatment did not affect maternal behavior when mothers were fed either n-3 LCPUFA or its precursors.
Developmental milestones of offspring are delayed by n-3 LCPUFA supplementation
We have recently shown that prenatally VPA-exposed mice exhibit significant early postnatal behavioral impairments3. Here, we investigated righting reflex and eye-opening time-period from P9 to P16 in relation to treatment and diet (Fig. 1). SAL/n-3 supp males showed an aberrant righting reflex at P9 and P13 compared to both the SAL/n-3 bal group and compared to the VPA/n-3 supp group specifically at P13 only. Similar differences were found in females at P11 but not at P13 (Fig. 1a). In addition, eye opening was delayed in both male and female SAL/n-3 supp mice compared to SAL/n-3 bal mice at P13 and only in males at P14. Eye opening was also delayed in VPA/n-3 supp females compared to VPA/n-3 bal females at P13 and P14 (Fig. 1b).

Developmental milestones of offspring are delayed by n-3 LCPUFA supplementation. (a) Righting reflex time at P9 (top), P11 (middle) and P13 (bottom) in both males (left) and females (right). n = 27 (SAL/n-3 bal male), 24 (VPA/n-3 bal male), 34 (SAL/n-3 supp male), 24 (VPA/n-3 supp male), 38 (SAL/n-3 bal female), 25 (VPA/n-3 bal female), 26 (SAL/n-3 supp female) and 24 (VPA/n-3 supp female) mice. (b) Eye opening score from P12 to P16 in both males (left) and females (right). n = 29 (SAL/n-3 bal male), 27 (VPA/n-3 bal male), 36 (SAL/n-3 supp male), 26 (VPA/n-3 supp male), 39 (SAL/n-3 bal female), 28 (VPA/n-3 bal female), 27 (SAL/n-3 supp female) and 26 (VPA/n-3 supp female) mice. Data are expressed as mean ± SEM and were analyzed through a two-way ANOVA followed by Tukey post-hoc multiple analysis. *p < 0.05, **p < 0.01, ***p < 0.001.
These results indicate that n-3 supp diet affected sensorimotor development dependent on VPA treatment and sex of the animals.
No major behavioral or cellular alterations following VPA treatment regardless of dietary intervention
Previous evidence from our group as well as other groups have reported that mice exposed prenatally to VPA and that were under a standard diet, consistently show reduced social behavior in the 3-CT test3,29,30. Here, using the same experimental procedures in the same housing conditions as previously, we report that both diets prevented the VPA-induced social impairments in male and female young adult mice (Fig. 2a). Three-way ANOVA showed that all groups spent more time in the social chamber compared to the nonsocial one. A treatment effect was found in VPA/n-3 supp males that were slightly more social than the SAL/n-3 supp group. We also investigated the time spent close to the novel mouse as well as the distance traveled in the chambers during this test and found the same outcome, where all groups showed normal social abilities (data not shown).

Offspring social, and grooming behavior are not altered by VPA following both n-3 PUFA diets. (a) Phase II from the 3-CT in both males (left) and females (right females). n = 27 (SAL/n-3 bal male), 25 (VPA/n-3 bal male), 35 (SAL/n-3 supp male), 25 (VPA/n-3 supp male), 39 (SAL/n-3 bal female), 27 (VPA/n-3 bal female), 27 (SAL/n-3 supp female) and 26 (VPA/n-3 supp female) mice. Data are expressed as mean ± SEM and were analyzed through a three-way ANOVA followed by Tukey post-hoc multiple analysis. *p < 0.05, **p < 0.01, ***p < 0.001. SC social chamber, NSC non-social chamber. (b) Number of grooming episodes and cumulative grooming duration in both males (left) and females (right). (c) Number of rearing episodes and cumulative rearing duration in both males (left) and females (right). n = 28 (SAL/n-3 bal male), 27 (VPA/n-3 bal male), 35 (SAL/n-3 supp male), 24 (VPA/n-3 supp male), 37 (SAL/n-3 bal female), 27 (VPA/n-3 bal female), 26 (SAL/n-3 supp female) and 26 (VPA/n-3 supp female) mice. Data are expressed as mean ± SEM and were analyzed through a three-way ANOVA for 3-CT or two-way ANOVA for grooming and rearing behavior, followed by Tukey post-hoc multiple analysis. *p < 0.05, **p < 0.01, ***p < 0.001.
Stereotyped behavior has often been reported in various genetic and environmental animal models of ASD2,3,4,5. This behavior is reminiscent of repetitive behaviors in ASD patients and constitutes one of the major symptoms used in the diagnosis of ASD1. We have previously shown that prenatal exposure to VPA significantly increases stereotyped behavior in both males and females as compared to controls3. Here, we report no differences in grooming parameters between groups, (Fig. 2b). In addition, VPA/n-3 bal male and female mice showed an increased frequency and duration of rearing behavior compared to both SAL/n-3 bal and VPA/n-3 supp mice (Fig. 2c). However, VPA had no effect on rearing behavior in the supplemented groups as no differences were observed in VPA/n-3 supp versus SAL/n-3 supp groups or in SAL/n-3 bal versus SAL/n-3 supp groups.
Gait impairments have often been reported in both ASD patients and animal models, however they are not yet considered as a diagnostic criteria in ASD39,40. We have previously shown significant gait deficits in genetic and environmental ASD animal models, including the VPA model2,3,4,5. Here, we show moderate gait deficits, which were for the most part observed in the VPA/n-3 supp male group as compared to the SAL/n-3 supp group (Fig. 3). For instance, dynamic and temporal parameters such as stride length and swing time were affected in the supplemented groups under VPA. Morphological parameters such as paw width, paw length and paw area were also altered by both treatment and diet. In females, only a few parameters were affected, such as kinematic-related fore and hindlimbs base of support. VPA/n-3 bal females showed a decreased base of support compared to the VPA/n-3 supp group on both forelimbs and hindlimbs, and additionally to SAL/n-3 bal females for forelimbs only.

Offspring gait analysis exhibited sex differences in relation with diet. (a) Paw width in males. (b) Paw length in males. (c) Paw area in males. (d) Stride length in males. (e) Swing time in males. n = 27 (SAL/n-3 bal male), 26 (VPA/n-3 bal male), 36 (SAL/n-3 supp male), 24 (VPA/n-3 supp male) mice. (f) Fore and Hindlimbs Base of Support in females. n = 39 (SAL/n-3 bal female), 28 (VPA/n-3 bal female), 27 (SAL/n-3 supp female), 26 (VPA/n-3 supp female) mice. Data are expressed as mean ± SEM and were analyzed through a two-way ANOVA followed by Tukey post-hoc multiple analysis. *p < 0.05, **p < 0.01, ***p < 0.001.
Taken together, these results indicate that both diets were able to reduce if not compensate for ASD related social, motor and gait symptoms, whatever the treatment, diet, or sex.
We have previously shown significant PC number decreases in the crus I or crus II cerebellar areas in both environmental and genetic ASD animal models3,4,5. Here, we examined PC number in crus I and crus II in all experimental groups (Suppl. Fig. S2). Two-way ANOVA analysis showed no treatment or diet effect in either males or females These results indicate that both n-3 bal and n-3 supp diets protected from PC loss induced by VPA in males and females alike, contrary to previous observations in mice fed with a standard animal facility diet.
Diet, but not VPA treatment, differentially alters n-3 and n-6 LCPUFA levels in the liver, but not in the cerebellum
We assessed the FA profile of the liver and the cerebellum, a cerebral region with high concentrations of DHA, sensitive to changes in FA composition, and that is at the crossroad of many cellular and molecular alterations in ASD, including loss of PC 41. Analyses were performed longitudinally to compare dam and offspring FA profiles. Given that n-3 LCPUFA supplementation was provided through diet, we analyzed, among others, levels of n-3 and n-6 LCPUFAs, DHA, EPA and AA (Fig. 4 and Suppl. Fig. S3). EPA levels were significantly increased in both the cerebellum and the liver in dams, male and female offspring, as expected. In the liver, in all n-3 LCPUFA supplemented groups, whether dams, male or female offspring, a significant increase (up to threefold) in DHA levels was found compared to n-3 balanced groups. In the cerebellum, DHA levels also increased in the VPA/n-3 supp group compared to VPA/n-3 bal animals, but only moderately (+ 25% approximately) and only in female offspring. Interestingly, no group differences in DHA cerebellar levels were found in dams and male offspring. The AA levels decreased in SAL/n-3 supp and VPA/n-3 supp groups compared to SAL/n-3 bal and VPA/n-3 bal groups respectively both in the liver and the cerebellum and in males and females. However, dam AA levels in the liver increased in the VPA/n-3 bal compared to both SAL/n-3 bal and VPA/n-3 supp.

Both n-3 and n-6 LCPUFA profiles are modified in the liver, but not in the cerebellum, in relation with diet. Liver DHA and AA levels (top) in dams (left), offspring males (center) and females (right). n = 5 mice per group except 4 on SAL/n-3 supp male AA levels. Cerebellum DHA and AA levels (bottom) in dams (left), offspring males (center) and females (right). n = 5 mice per group except 4 on SAL/n-3 bal male and VPA/n-3 supp female AA levels. Data are expressed as median and min to max and were analyzed through a two-way ANOVA followed by Tukey post-hoc multiple analysis. *p < 0.05, **p < 0.01, ***p < 0.001.
These results indicate a major effect of n-3 LCPUFA supplementation on DHA and AA levels within the liver but not within the cerebellum where DHA levels were equivalent (saline and VPA males) or only slightly increased (in VPA females).
Moderate to no alterations in inflammatory markers in conjunction with diet and treatment
n-3 PUFAs are associated with anti-inflammatory processes and could influence behavior output9. Here, we investigated mRNA levels of several markers of inflammation in the offspring cerebellum: TNF-alpha, TGF-beta, and ARG. No differences were found between groups for TGF-beta and ARG, whatever the sex. However, TNF-alpha expression increased slightly but significantly in SAL/n-3 supp females compared to SAL/n-3 bal females. While VPA treatment increased TNF-alpha levels in males, no subsequent differences between groups were found (Fig. 5).

Moderate to no alteration in inflammatory markers in conjunction with diet and treatment. TNF-alpha cerebellar levels (top) in both males and females. n = 5 (SAL/n-3 bal male), 5 (VPA/n-3 bal male), 4 (SAL/n-3 supp male), 4 (VPA/n-3 supp male), 4 (SAL/n-3 bal female), 5 (VPA/n-3 bal female), 5 (SAL/n-3 supp female) and 5 (VPA/n-3 supp female) mice. TGF-beta cerebellar levels (middle) in both males and females. n = 5 (SAL/n-3 bal male), 5 (VPA/n-3 bal male), 5 (SAL/n-3 supp male), 4 (VPA/n-3 supp male), 5 (SAL/n-3 bal female), 5 (VPA/n-3 bal female), 5 (SAL/n-3 supp female), 4 (VPA/n-3 supp female) mice. ARG cerebellar levels (bottom) in both males and females. n = 5 (SAL/n-3 bal male), 5 (VPA/n-3 bal male), 5 (SAL/n-3 supp male), 4 (VPA/n-3 supp male), 5 (SAL/n-3 bal female), 5 (VPA/n-3 bal female), 5 (SAL/n-3 supp female), 4 (VPA/n-3 supp female) mice. Data are expressed as median and min to max and were analyzed through a two-way ANOVA followed by Tukey post-hoc multiple analysis. *p < 0.05, **p < 0.01, ***p < 0.001.
Microbiota inter- and intra-diversity and bacteria relative abundance are not affected by diet
Microbiota has been shown to be influenced by in utero VPA treatment on offspring and by n-3 PUFA supplementation or deficiency42,43,44. Using a n-3 balanced or n-3 LCPUFA supplemented diet, we found no differences in alpha-diversity in either dams or offspring. Interestingly, offspring beta-diversity was not different across groups, whereas dam beta-diversity was increased by VPA treatment, whatever the diet, with VPA n-3 suppl dams being the most dissimilar (Fig. 6a and Suppl. Figs. S4–S6). Additionally, in dams, but not in offspring, relative abundances of Bacteroidetes and Actinobacteria were increased in VPA/n-3 supp group compared to SAL/n-3 supp and VPA/n-3 bal group (Fig. 6b and Suppl. Fig. S7). No differences were found in female offspring phyla abundance, however, male offspring Bray–Curtis dissimilarity analysis showed that the VPA/n-3 supp group has a more diverse microbial composition compared to SAL/n-3 supp group (Fig. 6c and Suppl. Fig. S5). These results show no major differences between diets on the offspring microbiota, regardless of the treatment.

VPA does not alter offspring microbiota inter- and intra-diversity nor offspring microbiota relative abundance under both n-3 PUFA diets. Microbial diversity: (a) Chao1 index (first line) in dams (left), offspring males (center) and females (right). n = 5 mice per group. (b) Bray–Curtis index (third line) in dams (leftl), offspring males (center) and females (right). n = 10 (SAL/n-3 bal dam), 25 (VPA/n-3 bal dam), 25 (SAL/n-3 supp dam), 25 (VPA/n-3 supp dam), 10 (SAL/n-3 bal male), 25 (VPA/n-3 bal male), 20 (SAL/n-3 supp male), 25 (VPA/n-3 supp male), 10 (SAL/n-3 bal female), 25 (VPA/n-3 bal female), 25 (SAL/n-3 supp female) and 25 (VPA/n-3 supp female) mice. (c) Phyla abundance: Bacteroidetes abundance (first line) in dams (left), offspring males (center) and females (right). n = 5 mice per group except 4 in SAL/n-3 supp and VPA/n-3 supp male. Firmicutes abundance (second line) in dams (left), offspring males (center) and females (right). n = 5 mice per group except 4 in SAL/n-3 supp male and SAL/n-3 bal female. Data are expressed as median and min to max and were analyzed through a two-way ANOVA followed by Tukey post-hoc multiple analysis. *p < 0.05, **p < 0.01, ***p < 0.001.
Discussion
In this study, we investigated the potential benefits of adding n-3 LCPUFA as compared to an isocaloric balanced diet with a balanced LA (n-6)/ALA (n-3) ratio starting from the perinatal period to adulthood on a multiplicity of social, motor and gait behaviors as well as on cerebellar cellular, molecular, and metabolic correlates in both male and female VPA mice models of ASD. We found that adult VPA-exposed animals, regardless of the diet, were not showing social deficits, stereotypies or cerebellar PC loss, all of which are major hallmarks of ASD and were consistently reported in the VPA rodent model fed with a standard diet (see our reviews on the subject2,36). Developmental milestones, gait and inflammatory profiles were only slightly affected by diet in conjunction with VPA prenatal exposure. Gut microbiota composition was not altered either by diet or treatment. Additionally, cerebellar DHA levels were not affected by diet, while liver DHA levels dramatically increased with a n-3 LCPUFA supplementation. Using dams and their offspring, we were also able to perform longitudinal studies on several parameters. We found that while maternal behavior was not altered by either treatment or diet, dams exhibited a change in their microbiota composition and increased n-3 LCPUFA liver levels in conjunction with VPA treatment. Taken together, these results indicate that both diets with either n-3 PUFA precursors from plant oils or additional n-3 LCPUFAs from fatty fish provide protection against major behavioral, cellular, and molecular VPA-induced ASD symptoms.
In addition, we report that the n-3 LCPUFA supplemented groups exhibited a slight delay in both righting reflex and eye opening, in accordance with previous studies reporting adverse consequences in postnatal development after maternal dietary n-3 LCPUFA supplementation or deficiency during gestation and lactation45,46,47,48. Contrary to the common misconception that elevated levels of n-3 LCPUFAs are strictly beneficial, a change in the n-6/n-3 PUFA ratio, irrespective of its direction, was reported to be deleterious during development, with some consequences at adulthood, depending on the animal model22,23. Indeed, in both the BTBR and C57BL/6 J mouse strains, body weight was decreased following perinatal diet intervention whereas in the Fmr1 KO mouse, body weight was increased22,23. In addition, we report here that the n-3 balanced diet protected from VPA-induced developmental delays that we previously reported with a standard diet3, as both righting reflex and eye-opening scores in VPA-exposed animals were similar to the saline groups. Metabolic studies on this matter may help unravel the mechanisms behind this developmental delay.
Deficits in social preference or social novelty and increased grooming behavior are repeatedly found in several animal models of ASD including the VPA model under a standard diet, as we also reported recently3,4,5. Here, we found that both diets, n-3 PUFA balanced and n-3 LCPUFA supplemented, protected against social deficits and stereotypy anomalies, two major ASD-associated symptoms. To our knowledge, this study is the first to investigate n-3 LCPUFA and precursor effects on social behavior since the vast majority of studies compared n-3 LCPUFA supplementation and deficiency, where n-3 LCPUFA supplementation was shown to alleviate stereotypies and social behavior impairments49.
We have previously shown, in the same experimental settings, that rearing, which represents global activity, decreased in VPA-exposed animals fed with a regular animal facility diet3. Here, the n-3 balanced diet increased rearing behavior in VPA-exposed animals compared to controls and this was normalized with the n-3 LCPUFA supplemented diet. In physiological conditions, high n-3 LCPUFA supplementation for 3 weeks after weaning reduces rearing50, whereas in depression and anxiety models, which are ASD comorbidities, opposite results are found51.
Our previous work on several environmental and genetic ASD animal models fed with a regular diet consistently showed motor and gait impairments3,4,5. Gait is seldom explored in these models even though ASD patients exhibit an irregular walk and balance difficulties associated with cerebellar dysfunction52. Here, we showed that diet had a differential effect on ASD VPA male and female mice. The n-3 LCPUFA supplementation ameliorated gait parameters in females, with an increased hindlimb base of support, suggesting better stability, whereas VPA-exposed males with n-3 LCPUFA supplementation displayed dynamic, temporal and morphological impairments compared to controls, albeit of a lower magnitude than what we observed with animals under a regular diet3. These results suggest moderate sex-dependent gait deficiencies in conjunction with n-3 LCPUFA supplementation, possibly resulting from metabolic differences between males and females which are hypothesized to be estrogen-related53.
Crus I and crus II cerebellar regions are involved in both cognitive and motor functions, which make them a target of choice in ASD physiopathology2,54. A decrease in PC number has been widely reported in both ASD patients and in animal models, including the VPA mouse model under a standard diet3,5,55,56,57,58. Here, we showed that PC cell number in these regions was not altered between groups, regardless of the treatment, diet, or sex. These findings fit with our main hypothesis, which is that both diets, with n-3 PUFA precursors or with additional n-3 LCPUFAs, protect from VPA-induced ASD behavioral symptoms and cellular correlates.
We then investigated FA profiles in the liver and the cerebellum in order to determine whether n-3 PUFA dietary supplementation is associated with higher n-3 PUFA levels in these regions. The n-3 LCPUFA diet was highly supplemented in DHA and EPA, whereas the n-3 balanced diet contained n-3 precursors (ALA), with shared LA/ALA ratio of 6.2 in the two diets. Thus, our lipid analysis focused on n-3 and n-6 LCPUFAs, DHA, EPA and AA respectively. As expected, n-3 LCPUFA dietary supplementation resulted in a major decrease in liver and cerebellar AA levels and a significant increase in liver and cerebellar EPA levels in both males and females. However, liver DHA levels were increased in all groups. In the cerebellum, there was a diet effect with increased DHA levels in the VPA female group, but not in males. This indicates that a n-3 LCPUFA supplementation does not further increase cerebellar DHA levels in the male groups. These findings are in line with another study where a n-3 PUFA balanced diet with ALA as the only source of n-3 PUFA, increased DHA and decreased AA levels in the cortex and protected from deficits in emotional behavior in adult and old CD1 mice (2–5 months and 19–23 months), as compared to a n-3 deficient diet59. Previous studies have investigated the role of n-3 LCPUFA on inflammation and concluded that high DHA brain levels are linked to an anti-inflammatory profile whereas high AA brain levels are correlated with a pro-inflammatory profile60,61,62. We found that cerebellar TNF-alpha mRNA levels were increased in SAL/n-3 supp females only. This cytokine is one of the most studied and is known to increase levels of AMPA receptors and number of synapses, enhancing excitatory post-synaptic activity63. Through this mechanism, TNF-alpha has been found to improve learning and memory in rats64. Taken together, these results on PUFA liver and cerebellar profiles, and on cerebellar inflammation, highlight sex differences and female sensitization to n-3 LCPUFA supplementation.
ASD patients suffer from gastrointestinal issues (GI) hypothesized to result from a gut microbiota dysbiosis, i.e. a microbial composition imbalance65,66. Differences in alpha and beta-diversity and an imbalance in Bacteroidetes and Firmicutes have been consistently reported in the VPA ASD mouse model fed with a standard diet42,67. The n-3 LCPUFA supplementation was shown to contribute to microbiota diversity and homeostasis44,68, but studies on ALA effects are lacking. Here, we found no differences in alpha-diversity, beta-diversity, or phyla abundance in either male or female offspring, whatever the sex, treatment, or diet. This further consolidates our proposition that both diets, with or without n-3 LCPUFAs, bring behavioral and biological benefits in the VPA-induced ASD animal model.
One of the strengths of this study is the global and longitudinal approach where, in addition to male and female offspring analyzed separately, we also investigated treatment and diet influences on dam maternal behavior, liver and cerebellar FA profiles, as well as microbiota composition. Maternal care received by the pups during the first postnatal weeks can affect their behavior in adulthood69,70 and in our hands, we did not find any differences in maternal behavior between the dam groups, regardless of treatment or diet. These findings align with those of another study, where VPA treatment did not affect maternal behavior71. However, we found more drastic differences due to treatment and diet interaction in dams than in offspring pertaining to maternal FA profiles and gut microbiota composition. In fact, VPA-exposed dams with the n-3 balanced diet exhibited increased AA liver levels, which were normalized with the n-3 LCUPFA supplementation diet. In addition, Bacteroidetes and Actinobacteria proportion in VPA/n-3 supp dams increased, as did the beta-diversity in this experimental group. Dams may be more sensitive to diet changes as they were fed with a regular diet before gestation, whereas offspring were given the same diet, whether balanced or supplemented, from embryonic stage to sacrifice at adulthood.
There are few limitations to this study that need to be highlighted. (i), As shown in Supplementary Figs. S8 and S9, SAL/n-3 supp groups tend to have a higher body weight than the SAL/n-3 bal group. (ii), Further studies are needed with a direct comparison of VPA effects in mice under a standard diet or a n-3 deficient diet. (iii), Diets used in this study differed mainly by the presence or not of LCPUFA, and therefore they have a different fatty acid composition and origin (from plant oils or fatty fish), which could have also a biological impact.
Taken together, our findings indicate that n-3 PUFA dietary supplementation, with or without LCPUFAs, prevent ASD-related disturbances in the VPA mouse model. These beneficial effects were evidenced at the behavioral, cellular, and molecular levels, in both sexes, although females seem to be somewhat more sensitive to n-3 LCPUFA supplementation. Additional investigations are warranted, aiming at deciphering the underlying biological mechanisms of dietary effects on ASD symptoms. They also need to consider sex and age differences, two parameters seldomly investigated at least in preclinical settings, where experiments are performed mostly in young adult males under a regular diet.
Methods
Animals and treatment
Animal housing and experimental procedures were performed in accordance with the European Union directive (2010/63/EU) and validated by the ethics committee (Approval # 202002051628899). C57BL/6J mice (Charles River Laboratories) were housed in ventilated cages with access to food and water ad libitum. Room temperature was maintained at 23 °C on a 12-h-light/-dark cycle (08:00–20:00).
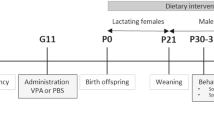
A total of 337 mice were used in this study: 55 females and 44 males were used for mating, resulting in 120 female and 118 male offspring. From the first gestational day (E0) and throughout gestation and nursing, pregnant females were fed with an isocaloric diet supplemented with n-3 LCPUFA (DHA and EPA) (n-3 supp) or not (n-3 bal) until weaning of their litter (SAFE, Augy, France). Both diets had a LA/ALA ratio of 6.2, and thus share the same amount of n-3 and n-6 precursors. As such, diets differed mostly by the presence of n-3 LCPUFA, DHA and EPA in the n-3 supp group (detailed description of diets are provided in Suppl. Tables S1 and S2). Animals under the n-3 supp diet received approximately 481.12 DHA and 703.99 EPA (mg/kg of body weight/day). Pregnant females received a single i.p. injection of either VPA (450 mg/kg, Sigma-Aldrich, P4543) or saline solution (NaCl 0.9%) at gestational day E12.5 as previously described3,30. After giving birth, dams were allocated to four experimental groups depending on prenatal treatment and diet: SAL fed with the n-3 bal diet (SAL/n-3 bal), VPA fed with n-3 bal diet (VPA/n-3 bal), SAL fed with the n-3 supp diet (SAL/n-3 supp) and VPA n-3 fed with the n-3 supp diet (VPA/n-3 supp) (timeline and experimental procedures are detailed in Suppl. Fig. S10). Male and female offspring were separated at weaning (postnatal day 28, P28) and housed by groups of 2 to 5 animals per cage, where they received ad libitum access to water and controlled access to the same diet as the respective dams. Dam body weight (postnatal day 14, P14) as well as male and female offspring body weights (weeks 4, 5, 6, 7 and 8) are presented in Supplementary Fig. S8. Male and female offspring food intakes are presented in Supplementary Fig. S9. Male and female offspring were affected into one of the following 8 groups for behavior analysis: 29 males and 39 females SAL/n-3 bal, 36 males and 27 females SAL/n-3 supp, 27 males and 28 females VPA/n-3 bal, and 26 males and 26 females VPA/n-3 supp. The same animals were used for gut microbiota, FA profile and inflammation markers. For this, 5 mice were randomly selected per group, with a maximum of 2 animals from the same litter and sex.
Maternal and offspring behavior procedures
Maternal behavior experiments were performed on SAL/n-3 bal (n = 15), SAL/n-3 supp (n = 14), VPA/n-3 bal (n = 13), and VPA/n-3 supp (n = 12) groups. Dam performances on pup retrieval were assessed at P9. For this, both the litter and the dam were removed and separated from the home cage for 5 min before being placed back. Dam behavior directed towards the pups, or the environment was recorded for 15 min and analyzed on Solomon Coder (András Péter, Keele, UK).
Developmental milestones of offspring were assessed by measuring righting reflex at P9, P11 and P13 as well as eye opening from P12 to P16. Spontaneous activity in the cylinder was recorded at P48 for grooming and rearing scoring using Solomon Coder (András Péter, Keele, UK) as previously described3,72. Spatial, temporal, and kinematic gait parameters were analyzed during spontaneous walk at P49 using an automated gait analysis system (Gaitlab, Viewpoint, France) as previously described3. Social interaction was assessed between P50 and P60 using the three-chamber test (3-CT) as previously described3,73.
Tissue processing, immunohistochemistry and stereology counting
At the end of behavioral experiments, mice were deeply anesthetized with pentobarbital (120 mg/kg) for tissue processing. Animal perfusion, brain retrieval, tissue sectioning and cresyl violet/neutral red staining were performed as previously described4. Stereological estimates of PC number within crus I and II cerebellar regions were obtained using the optical fractionator method (Mercator Software, Explora Nova, France) and systematic random sampling as previously described4 in the following groups: [Crus I: SAL/n-3 bal (males n = 11 and females n = 13), SAL/n-3 supp (males n = 11 and females n = 7), VPA/n-3 bal (males n = 10 and females n = 11) and VPA/n-3 supp (males n = 10 and females n = 9)] and [Crus II: SAL/n-3 bal (males n = 11 and females n = 12), SAL/n-3 supp (males n = 12 and females n = 9), VPA/n-3 bal (males n = 10 and females n = 11) and VPA/n-3 supp (males n = 10 and females n = 9].
Fecal microbiome analysis
Fecal samples were collected from both the dam through lactation period (P15, P20 and P22) and the litter after weaning (P36, P41 and P43). Samples were conserved at − 80 °C in nucleic acid conservative buffer until DNA extraction (RNA Protect, Qiagen, Venlo, The Netherlands). Wet-lab and bioinformatic analysis were performed as previously described74,75. After verification on rarefaction curve, rarefaction (subsampling to 5000 sequences per sample) was performed before determination of alpha diversity metrics (Chao1′s and Shannon’s indexes) and beta diversity metrics (Bray–Curtis dissimilarity and weighted UniFrac76). ANCOM was used to identify differentially abundant genera among the groups77.
Analysis of inflammation makers by qPCR
RNA expression of genes implicated in inflammatory process was investigated as previously described78. Briefly, RNA from hemi-cerebellum of dams and male and female offspring was extracted using TRIzol extraction kit (Invitrogen, Life Technologies, Saint-Quentin-Fallavier, France). Purity and concentration of RNA were determined using a Nanodrop 1000 spectrophotometer (Nanodrop technologies, Wilmington, DE, USA) and a bioanalyzer (Agilent, Les Ulis, France). Reverse transcription was performed on one or two micrograms of RNA by Superscript IV (Invitrogen, Life Technologies, Saint Aubin, France). TaqMan® specific primers were used to amplify genes of interest as previously described78 with a focus on tumor necrosis factor alpha (TNF-alpha, Mm00443258_m1), transforming growth factor-beta 1 (TGF-beta 1, Mm01178820_m1) and Arginase 1 (Arg1, Mm00475988_m1). The housekeeping gene was beta-2-microglobulin (B2M, Mm00437762_m1). Fluorescence was determined on a LightCycler® 480 instrument II (Roche, La Rochelle, France). Data were analyzed using the comparative threshold cycle (Ct) method and results were expressed as relative fold change to control target mRNA expression.
Analysis of fatty acids in cerebellum and liver
FAs from the liver and the cerebellum were analyzed as previously described79,80,81. Briefly, liver and cerebellum lipids were extracted according to the Folch’s method82, FAs were transmethylated according to Morrison and Smith’s method83 and FA methyl esters (FAMEs) were analyzed on a FOCUS GC gas chromatograph (Thermo Electron Corporation) equipped with a split injector and a flame ionization detector, and a CPSil88-silica capillary column (100 m × 0.25 mm i.d., film thickness 0.20 μm, Varian, Les Ulis, France). The injector and the detector were maintained at 250 °C and 280 °C, respectively. Hydrogen was used as a carrier gas (inlet pressure 210 kPa). The oven temperature was fixed at 60 °C for 5 min, increased to 165 °C at a rate of 15 °C/min and then to 225 °C at a rate of 2 °C/min, and maintained at this temperature for 17 min. FAMEs were identified by making a comparison with commercial standards. FA composition is expressed as the percentage of total FAs.
Statistical analysis
For experiments related to behavior, data are expressed as mean ± Standard Error of the Mean while cellular and metabolic data are expressed as median ± min to max. Data were analyzed using GraphPad Prism-9 software (La Jolla, California, USA). Outliers were identified by Grubbs’ test and were removed from subsequent statistical analysis (to a maximum of 1 outlier removed per outcome). Two-way or three-way analysis of variance (ANOVAs) were performed followed by Tukey post-hoc multiple comparisons test when appropriate. For all analyses, a p value < 0.05 was considered significant. Detailed statistical analysis on all groups is available in Supplementary Tables S3–S15. Other detailed statistical analysis and raw data are available upon reasonable request.
ARRIVE guidelines
This study is in accordance with ARRIVE guidelines.
Data availability
Raw data for gut microbiota analysis are available on the following site (https://www.ncbi.nlm.nih.gov/bioproject/PRJNA854167). Accession SRA# are from SRR 19906240 to SRR19906299. Detailed statistical analysis is included in the supplementary material (Suppl. Tables S3–S15). Other details and raw data of all other experiments can be obtained upon reasonable request to the authors.
References
American Psychiatric Association. Diagnostic and Statistical Manual of Mental Disorders (DSM-5®) (American Psychiatric Pub, 2013).
Thabault, M. et al. Cerebellar and striatal implications in autism spectrum disorders: From clinical observations to animal models. Int. J. Mol. Sci. 23, 2294 (2022).
Al Sagheer, T. et al. Motor impairments correlate with social deficits and restricted neuronal loss in an environmental model of autism. Int. J. Neuropsychopharmacol. 21, 871–882 (2018).
Haida, O. et al. Sex-dependent behavioral deficits and neuropathology in a maternal immune activation model of autism. Transl. Psychiatry 9, 124 (2019).
Matas, E. et al. Major motor and gait deficits with sexual dimorphism in a Shank3 mutant mouse model. Mol. Autism 12, 2 (2021).
Davoli-Ferreira, M., Thomson, C. A. & McCoy, K. D. Microbiota and microglia interactions in ASD. Front. Immunol. 12, 676255 (2021).
Baraskewich, J., von Ranson, K. M., McCrimmon, A. & McMorris, C. A. Feeding and eating problems in children and adolescents with autism: A scoping review. Autism 25, 1505–1519 (2021).
Bazinet, R. P. & Layé, S. Polyunsaturated fatty acids and their metabolites in brain function and disease. Nat. Rev. Neurosci. 15, 771–785 (2014).
Martinat, M., Rossitto, M., Di Miceli, M. & Layé, S. Perinatal dietary polyunsaturated fatty acids in brain development. Role Neurodev. Disord. Nutr. 13, 1185 (2021).
Harris, W. S. Encyclopedia of dietary supplements. In Omega-3 Fatty Acids 2nd edn, (eds Coates, P. M. et al.) (Informa Healthcare, 2015).
Bell, J. G. et al. Essential fatty acids and phospholipase A2 in autistic spectrum disorders. Prostaglandins Leukot. Essent. Fatty Acids 71, 201–204 (2004).
Vancassel, S. et al. Plasma fatty acid levels in autistic children. Prostaglandins Leukot. Essent. Fatty Acids 65, 1–7 (2001).
Mazahery, H. et al. Relationship between long chain n-3 polyunsaturated fatty acids and autism spectrum disorder: Systematic review and meta-analysis of case-control and randomised controlled trials. Nutrients 9, 155 (2017).
Parletta, N., Niyonsenga, T. & Duff, J. Omega-3 and omega-6 polyunsaturated fatty acid levels and correlations with symptoms in children with attention deficit hyperactivity disorder, autistic spectrum disorder and typically developing controls. PLoS ONE 11, e0156432 (2016).
Yui, K., Imataka, G., Kawasak, Y. & Yamada, H. Increased ω-3 polyunsaturated fatty acid/arachidonic acid ratios and upregulation of signaling mediator in individuals with autism spectrum disorders. Life Sci. 145, 205–212 (2016).
Simopoulos, A. P. Evolutionary aspects of diet: The omega-6/omega-3 ratio and the brain. Mol. Neurobiol. 44, 203–215 (2011).
Blasbalg, T. L., Hibbeln, J. R., Ramsden, C. E., Majchrzak, S. F. & Rawlings, R. R. Changes in consumption of omega-3 and omega-6 fatty acids in the United States during the 20th century. Am. J. Clin. Nutr. 93, 950–962 (2011).
Shahidi, F. & Ambigaipalan, P. Omega-3 polyunsaturated fatty acids and their health benefits. Annu. Rev. Food Sci. Technol. 9, 345–381 (2018).
Madore, C. et al. Essential omega-3 fatty acids tune microglial phagocytosis of synaptic elements in the mouse developing brain. Nat. Commun. 11, 6133 (2020).
Alfawaz, H. et al. The independent and combined effects of omega-3 and vitamin b12 in ameliorating propionic acid induced biochemical features in juvenile rats as rodent model of autism. J. Mol. Neurosci. 66, 403–413 (2018).
Yadav, S. et al. Comparative efficacy of alpha-linolenic acid and gamma-linolenic acid to attenuate valproic acid-induced autism-like features. J. Physiol. Biochem. 73, 187–198 (2017).
van Elst, K. et al. Chronic dietary changes in n-6/n-3 polyunsaturated fatty acid ratios cause developmental delay and reduce social interest in mice. Eur. Neuropsychopharmacol. 29, 16–31 (2019).
Pietropaolo, S. et al. Dietary supplementation of omega-3 fatty acids rescues fragile X phenotypes in Fmr1-Ko mice. Psychoneuroendocrinology 49, 119–129 (2014).
Mazahery, H. et al. A randomised-controlled trial of vitamin d and omega-3 long chain polyunsaturated fatty acids in the treatment of core symptoms of autism spectrum disorder in children. J. Autism Dev. Disord. 49, 1778–1794 (2019).
Nevins, J. E. H. et al. Omega-3 fatty acid dietary supplements consumed during pregnancy and lactation and child neurodevelopment: A systematic review. J. Nutr. 151, 3483–3494 (2021).
Basil, P. et al. Prenatal immune activation alters the adult neural epigenome but can be partly stabilised by a n-3 polyunsaturated fatty acid diet. Transl. Psychiatry 8, 125 (2018).
Arndt, T. L., Stodgell, C. J. & Rodier, P. M. The teratology of autism. Int. J. Dev. Neurosci. 23, 189–199 (2005).
Christensen, J. et al. Prenatal valproate exposure and risk of autism spectrum disorders and childhood autism. JAMA 309, 1696 (2013).
Mabunga, D. F. N., Gonzales, E. L. T., Kim, J., Kim, K. C. & Shin, C. Y. Exploring the validity of valproic acid animal model of autism. Exp. Neurobiol. 24, 285–300 (2015).
Nicolini, C. & Fahnestock, M. The valproic acid-induced rodent model of autism. Exp. Neurol. 299, 217–227 (2018).
Bossu, J.-L. & Roux, S. The valproate model of autism. Med. Sci. 35, 236–243 (2019).
Kim, Y. S. et al. Prevalence of autism spectrum disorders in a total population sample. Am. J. Psychiatry 168, 904–912 (2011).
Kataoka, S. et al. Autism-like behaviours with transient histone hyperacetylation in mice treated prenatally with valproic acid. Int. J. Neuropsychopharmacol. 16, 91–103 (2013).
Kim, J.-W. et al. Subchronic treatment of donepezil rescues impaired social, hyperactive, and stereotypic behavior in valproic acid-induced animal model of autism. PLoS ONE 9, e104927 (2014).
Main, S. L. & Kulesza, R. J. Repeated prenatal exposure to valproic acid results in cerebellar hypoplasia and ataxia. Neuroscience 340, 34–47 (2017).
Jaber, M. Genetic and environmental mouse models of autism reproduce the spectrum of the disease. J. Neural Transm. 130, 425–432 (2023).
Kuo, H.-Y. & Liu, F.-C. Valproic acid induces aberrant development of striatal compartments and corticostriatal pathways in a mouse model of autism spectrum disorder. FASEB J. 31, 4458–4471 (2017).
Zhang, T. Y., Chrétien, P., Meaney, M. J. & Gratton, A. Influence of naturally occurring variations in maternal care on prepulse inhibition of acoustic startle and the medial prefrontal cortical dopamine response to stress in adult rats. J. Neurosci. 25, 1493–1502 (2005).
Fatemi, S. H. et al. Consensus paper: Pathological role of the cerebellum in autism. Cerebellum 11, 777–807 (2012).
Yirmiya, N. & Charman, T. The prodrome of autism: Early behavioral and biological signs, regression, peri- and post-natal development and genetics. J. Child. Psychol. Psychiatry 51, 432–458 (2010).
Joffre, C. et al. Modulation of brain PUFA content in different experimental models of mice. Prostaglandins Leukot. Essent. Fatty Acids 114, 1–10 (2016).
de Theije, C. G. M. et al. Altered gut microbiota and activity in a murine model of autism spectrum disorders. Brain Behav. Immun. 37, 197–206 (2014).
Leyrolle, Q. et al. Maternal dietary omega-3 deficiency worsens the deleterious effects of prenatal inflammation on the gut-brain axis in the offspring across lifetime. Neuropsychopharmacology 46, 579–602 (2021).
Robertson, R. C. et al. Omega-3 polyunsaturated fatty acids critically regulate behaviour and gut microbiota development in adolescence and adulthood. Brain Behav. Immun. 59, 21–37 (2017).
Wainwright, P. E., Jalali, E., Mutsaers, L. M., Bell, R. & Cvitkovic, S. An imbalance of dietary essential fatty acids retards behavioral development in mice. Physiol. Behav. 66, 833–839 (1999).
Church, M. W., Jen, K.-L.C., Dowhan, L. M., Adams, B. R. & Hotra, J. W. Excess and deficient omega-3 fatty acid during pregnancy and lactation cause impaired neural transmission in rat pups. Neurotoxicol. Teratol. 30, 107–117 (2008).
Bianconi, S. et al. Effects of dietary omega-3 PUFAs on growth and development: Somatic, neurobiological and reproductive functions in a murine model. J. Nutr. Biochem. 61, 82–90 (2018).
Yang, R. et al. Effects of maternal deprivation stress and maternal dietary of n-3 polyunsaturated fatty acids on the neurobehavioral development of male offspring. Nutr. Neurosci. 24, 865–872 (2021).
Fortunato, J. J. et al. Effects of ω-3 fatty acids on stereotypical behavior and social interactions in Wistar rats prenatally exposed to lipopolysaccarides. Nutrition 35, 119–127 (2017).
Rockett, B. D., Harris, M. & Raza, S. S. High dose of an n-3 polyunsaturated fatty acid diet lowers activity of C57BL/6 mice. Prostaglandins Leukot. Essent. Fatty Acids 86, 137–140 (2012).
Wu, Y.-Q. et al. Long chain omega-3 polyunsaturated fatty acid supplementation alleviates doxorubicin-induced depressive-like behaviors and neurotoxicity in rats: involvement of oxidative stress and neuroinflammation. Nutrients 8, 243 (2016).
Mosconi, M. W. et al. Feedforward and feedback motor control abnormalities implicate cerebellar dysfunctions in autism spectrum disorder. J. Neurosci. 35, 2015–2025 (2015).
Giltay, E. J., Gooren, L. J. G., Toorians, A. W. F. T., Katan, M. B. & Zock, P. L. Docosahexaenoic acid concentrations are higher in women than in men because of estrogenic effects. Am. J. Clin. Nutr. 80, 1167–1174 (2004).
D’Mello, A. M. & Stoodley, C. J. Cerebro-cerebellar circuits in autism spectrum disorder. Front. Neurosci. 9, 408 (2015).
Fatemi, S. H. et al. Purkinje cell size is reduced in cerebellum of patients with autism. Cell Mol. Neurobiol. 22, 171–175 (2002).
Bauman, M. L. & Kemper, T. L. Neuroanatomic observations of the brain in autism: A review and future directions. Int. J. Dev. Neurosci. 23, 183–187 (2005).
Bailey, A. et al. A clinicopathological study of autism. Brain 121(Pt 5), 889–905 (1998).
Ellegood, J., Pacey, L. K., Hampson, D. R., Lerch, J. P. & Henkelman, R. M. Anatomical phenotyping in a mouse model of fragile X syndrome with magnetic resonance imaging. Neuroimage 53, 1023–1029 (2010).
Moranis, A. et al. Long term adequate n-3 polyunsaturated fatty acid diet protects from depressive-like behavior but not from working memory disruption and brain cytokine expression in aged mice. Brain Behav. Immun. 26, 721–731 (2012).
Calder, P. C. Dietary modification of inflammation with lipids. Proc. Nutr. Soc. 61, 345–358 (2002).
Wainwright, P. E., Xing, H. C., Mutsaers, L., McCutcheon, D. & Kyle, D. Arachidonic acid offsets the effects on mouse brain and behavior of a diet with a low (n-6):(n-3) ratio and very high levels of docosahexaenoic acid. J. Nutr. 127, 184–193 (1997).
Palacios-Pelaez, R., Lukiw, W. J. & Bazan, N. G. Omega-3 essential fatty acids modulate initiation and progression of neurodegenerative disease. Mol. Neurobiol. 41, 367–374 (2010).
Beattie, E. C. et al. Control of synaptic strength by glial TNFalpha. Science 295, 2282–2285 (2002).
Brennan, E. M., Martin, L. J., Johnston, M. V. & Blue, M. E. Ontogeny of non-NMDA glutamate receptors in rat barrel field cortex: II. Alpha-AMPA and kainate receptors. J. Comp. Neurol. 386, 29–45 (1997).
de Theije, C. G. M. et al. Pathways underlying the gut-to-brain connection in autism spectrum disorders as future targets for disease management. Eur. J. Pharmacol. 668(Suppl 1), S70-80 (2011).
Bolte, E. R. Autism and Clostridium tetani. Med. Hypotheses 51, 133–144 (1998).
Lim, J. S., Lim, M. Y., Choi, Y. & Ko, G. Modeling environmental risk factors of autism in mice induces IBD-related gut microbial dysbiosis and hyperserotonemia. Mol. Brain 10, 14 (2017).
Kaliannan, K., Wang, B., Li, X.-Y., Kim, K.-J. & Kang, J. X. A host-microbiome interaction mediates the opposing effects of omega-6 and omega-3 fatty acids on metabolic endotoxemia. Sci. Rep. 5, 11276 (2015).
Weaver, I. C. G., Meaney, M. J. & Szyf, M. Maternal care effects on the hippocampal transcriptome and anxiety-mediated behaviors in the offspring that are reversible in adulthood. Proc. Natl. Acad. Sci. USA 103, 3480–3485 (2006).
Lucchina, L., Carola, V., Pitossi, F. & Depino, A. M. Evaluating the interaction between early postnatal inflammation and maternal care in the programming of adult anxiety and depression-related behaviors. Behav. Brain Res. 213, 56–65 (2010).
Kazlauskas, N., Campolongo, M., Lucchina, L., Zappala, C. & Depino, A. M. Postnatal behavioral and inflammatory alterations in female pups prenatally exposed to valproic acid. Psychoneuroendocrinology 72, 11–21 (2016).
Fleming, S. M., Ekhator, O. R. & Ghisays, V. Assessment of sensorimotor function in mouse models of Parkinson’s disease. J. Vis. Exp. 76, 50303 (2013).
Moy, S. S. et al. Sociability and preference for social novelty in five inbred strains: An approach to assess autistic-like behavior in mice. Genes Brain Behav. 3, 287–302 (2004).
Langevin, S. et al. Early nasopharyngeal microbial signature associated with severe influenza in children: A retrospective pilot study. J. Gen. Virol. 98, 2425–2437 (2017).
Gachet, C., Prat, M., Burucoa, C., Grivard, P. & Pichon, M. Spermatic microbiome characteristics in infertile patients: Impact on sperm count, mobility, and morphology. JCM 11, 1505 (2022).
Lozupone, C. A., Hamady, M., Kelley, S. T. & Knight, R. Quantitative and qualitative beta diversity measures lead to different insights into factors that structure microbial communities. Appl. Environ. Microbiol. 73, 1576–1585 (2007).
Mandal, S. et al. Analysis of composition of microbiomes: A novel method for studying microbial composition. Microb. Ecol. Health Dis. 26, 27663 (2015).
Chataigner, M. et al. Dietary fish hydrolysate supplementation containing n-3 LC-PUFAs and peptides prevents short-term memory and stress response deficits in aged mice. Brain Behav. Immun. 91, 716–730 (2021).
Labrousse, V. F. et al. Short-term long chain omega3 diet protects from neuroinflammatory processes and memory impairment in aged mice. PLoS ONE 7, e36861 (2012).
Madore, C. et al. Nutritional n-3 PUFAs deficiency during perinatal periods alters brain innate immune system and neuronal plasticity-associated genes. Brain Behav. Immun. 41, 22–31 (2014).
Delpech, J.-C. et al. Transgenic increase in n-3/n-6 fatty acid ratio protects against cognitive deficits induced by an immune challenge through decrease of neuroinflammation. Neuropsychopharmacol 40, 525–536 (2015).
Folch, J., Lees, M. & Sloane Stanley, G. H. A simple method for the isolation and purification of total lipides from animal tissues. J. Biol. Chem. 226, 497–509 (1957).
Morrison, W. R. & Smith, L. M. Prepation of fatty acid methyl esters and dimethylacetals from lipids with boron fluoride-methanol. J. Lipid Res. 5, 600–608 (1964).
Acknowledgements
The research leading to the results detailed in this manuscript received funding from the Fondation pour la Recherche Médicale (MJ) and the CPER-FEDER program (MJ), FRC CONNECT (SL) and Région Nouvelle Aquitaine (SL and MJ, 2019-1R3M08). VT is a recipient of a joint PhD fellowship from Nouvelle-Aquitaine Region/Fondation pour la Recherche Médicale. MT is a recipient of a joint PhD fellowship from Nouvelle-Aquitaine Region/Inserm. This work has benefited from the facilities and expertise of the PREBIOS platform (Université de Poitiers). Authors are grateful for the research staff at the Eye and Nutrition Research Group for lipidomic analyses (UMR1324 INRAE, Dijon, France). Authors are thankful to Jeffrey Arsham and Nathaniel L. Ritz for careful language editing. Some of these results were recently presented in a poster format at the FENS meeting in Paris (July 2022).
Author information
Authors and Affiliations
Contributions
Methodology, Data Curation and Acquisition, Investigation, Writing and Editing: V.T. Methodology, Data Acquisition: M.S., E.B., M.T. Methodology: J.E.L., A.A., M.F., C.B. Methodology, Data Curation and Acquisition, Investigation, Editing: M.P. Conceptualization, Project administration, Validation and Editing: S.L., C.J. Conceptualization, Funding Acquisition, Validation, Original Draft Preparation, Writing and Editing, Project administration, Supervision: M.J. All authors have approved the submitted manuscript prior to submission. All authors agree to be personally accountable for their own contributions and for ensuring that questions related to the accuracy or integrity of any part of the work, even ones in which they were not personally involved, are appropriately investigated, resolved, and documented in the literature.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Turpin, V., Schaffhauser, M., Thabault, M. et al. Mice prenatally exposed to valproic acid do not show autism-related disorders when fed with polyunsaturated fatty acid-enriched diets. Sci Rep 13, 11235 (2023). https://doi.org/10.1038/s41598-023-38423-z
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-023-38423-z
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
