
Abstract
Salt caverns have been successfully used for natural gas storage globally since the 1940s and are now under consideration for hydrogen (H2) storage, which is needed in large quantities to decarbonize the economy to finally reach a net zero by 2050. Salt caverns are not sterile and H2 is a ubiquitous electron donor for microorganisms. This could entail that the injected H2 will be microbially consumed, leading to a volumetric loss and potential production of toxic H2S. However, the extent and rates of this microbial H2 consumption under high-saline cavern conditions are not yet understood. To investigate microbial consumption rates, we cultured the halophilic sulphate-reducing bacteria Desulfohalobium retbaense and the halophilic methanogen Methanocalculus halotolerans under different H2 partial pressures. Both strains consumed H2, but consumption rates slowed down significantly over time. The activity loss correlated with a significant pH increase (up to pH 9) in the media due to intense proton- and bicarbonate consumption. In the case of sulphate reduction, this pH increase led to dissolution of all produced H2S in the liquid phase. We compared these observations to a brine retrieved from a salt cavern located in Northern Germany, which was then incubated with 100% H2 over several months. We again observed a H2 loss (up to 12%) with a concurrent increase in pH of up to 8.5 especially when additional nutrients were added to the brine. Our results clearly show that sulphate-reducing microbes present in salt caverns consume H2, which will be accompanied by a significant pH increase, resulting in reduced activity over time. This potentially self-limiting process of pH increase during sulphate-reduction will be advantageous for H2 storage in low-buffering environments like salt caverns.
Similar content being viewed by others
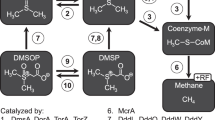

Introduction
Decarbonisation of industrial and societal activities is anticipated to hinge on “Power-to-Gas” technologies, which use electricity derived from solar or wind sources to produce hydrogen (H2), a universal energy carrier. H2 can be utilized, transported or stored for later use for several industrial sectors like chemical industry, heavy transport and steel production, “Gas-to-Power”. Storage of H2 is needed to buffer daily to seasonal variations in energy supply and demand1,2,3. Underground storage in salt caverns is suggested to be the ideal option for large-volume storage of H2 when the gas can be injected into a cavern and can be withdrawn flexibly for energy generation4,5,6. This process is very similar to the current storage of natural gas or oil. Salt caverns are solution mined large underground cavities inside a salt layer or salt dome by gradually dissolving the salt with freshwater or seawater. The resultant voids are several tens of meters in diameter and several hundreds of meters in length, with geometric cavern volumes of up to 106 cubic meters and maximum pressures of up to 300 bar. They are proposed to be ideal for short- to mid-term storage, with opportunities for rapid injection and withdrawal of gases for energy balancing. Major benefits of salt cavern storage are the available high volumes and relatively low operational costs5. As the demand for more storage sites increases, and upfront capital investments for salt caverns are high (~ 30 M€/onshore cavern7) there is an increasing need for improved understanding of possible microbiologically triggered subsurface reactions of H2 within the salt cavern. These reactions can potentially reduce the caloric value as well as leading to health/safety/environmental relevant generation of H2S, which constrain the operational window and requires purification measures. Currently there are only a hand full of salt cavern sites used for H2 storage worldwide5, without reported failures but operational data is not available.
Salt caverns are, like most subsurface environments, not sterile but harbour diverse microbial organisms8,9,10. Specially adapted extremophiles (halotolerant or halophilic) can live in or even require high-salt conditions for their survival11,12. Although high osmotic stress is suggested to cause energetic constraints13 by forcing the microorganisms to spend high amounts of energy for osmoregulation, i.e., production of compatible solutes; a higher salinity in salt cavern brines does not necessarily prevent the risks of microbial presence and/or activity. In case of H2 storage, the microorganisms will be in direct contact with the stored H2 for an extended period (up to several months for mid-/long-term storage). H2, being an excellent and ubiquitous electron donor, is an important driver for microbial activity in living environments14,15, which is in stark contrast to natural gas (CH4). Microbial activity in H2 filled salt caverns could induce a variety of processes and risks related to them: gas volumetric effects, gas composition changes and purity loss by e.g. sulphate-reducing microbes (SRM) forming the toxic gas H2S (see reaction 1), methanogenic archaea forming CH4 (see reaction 2), overall causing a reduced energetic value16,17.
In field trials for H2 storage in porous reservoirs and aquifers, microbial H2 consumption and conversion into CH4 has been described several times18,19. However, for salt caverns the overall microbial risks are not well studied and for cavern operators the extent of the microbial problem is unclear. For example, it is not known whether or how microbial H2 oxidation processes will occur in salt caverns and if so, at which speed and how pronounced. Some modelling approaches have indicated potential H2S formation20,21. However, these models are based on kinetic rates of sulphate reducers grown under standard laboratory conditions, and it can be assumed that a) growth and consumption is different with H2 as an electron donor, and b) that extremely halophilic strains show different rates due to their energy expenditures on osmoregulation. For better prediction of the microbial risks and the underlying economic risks, it is therefore necessary to study specific halophilic microbial rates to estimate microbial H2 oxidation under high-salt conditions.
Therefore, the aim of this study was to investigate known halophilic H2-consuming microbial strains to obtain not only H2 consumption rates but also to find some key parameters that influence and/or can be used as indicators for microbial H2 consumption. Furthermore, we compared our findings with microbial enrichments from a brine sampled in a salt cavern located in Northern Germany. These enrichments were incubated with H2 for over 150 days at the cavern specific temperature range. Our study clearly shows that microbial H2 consumption is a relevant topic for salt cavern storage and delivers important kinetic data on H2 consumption of both cultured and environmental samples.
Material and methods
Source of organisms
The two halophilic cultures Desulfohalobium retbaense DSM569222 and Methanocalculus halotolerans DSM1409223 were purchased from the DSMZ (German Collection of Microorganisms and Cell Cultures GmbH). The original cavern brine was sampled at a cavern field located in Northern Germany. Samples were taken at the wellhead. Before taking samples the first brine was discarded (around of 10 min flushing) to avoid sampling the brine standing in the well. Afterwards the samples were filled into sterile and anoxic glass bottles under continuous nitrogen flush to preserve anoxic conditions and then immediately shipped to the lab. Brine properties are: salinity 27% (wt/wt), pH 7.4, sulphate 4190 + / − 57 mg/L, total inorganic carbon 84.9 + / − 0.4, total organic carbon 7.61 + / − 1.1 mg/L.
Standard culture conditions
Both reference strains were routinely cultured in their specific media given by the DSMZ. For D. retbaense DSM5692: 1 g/L NH4Cl, 0.3 g/L K2HPO4, 0.3 g/L KH2PO4, 20 g/L MgCl2 × 6 H2O, 100 g/L NaCl, 2.7 g/L CaCl2, 4 g/L KCl, 3 g/L Na2SO4, 1 mL/L trace element solution SL-10, 0.3 g/L Na2S × 9 H2O—pH 7.2. For M. halotolerans DSM14092: 1 g/L NH4Cl, 0.3 g/L K2HPO4, 0.3 g/L KH2PO4, 3.2 g/L MgCl2 × 6 H2O, 50 g/L NaCl, 0.6 g/L CaCl2, 0.17 g/L KCl, 3 g/L Na2SO4, 10 mL/L modified Wolins mineral solution, 0.3 g/L Na2S × 9 H2O, 2 g/L NaHCO3—pH 7.2. Carbon sources and yeast extract were added separately to the bottles depending on the experiment. Both microorganisms were incubated at their respective optimal temperature of 37 °C. Standard growth for D. retbaense was on 24 mM lactate with 0.1% yeast extract and 0.1% peptone. M. halotolerans was routinely cultivated with 20 mM acetate, 20 mM formate, 0.05% yeast extract and 80% H2, 20% CO2. Growth was confirmed by gas production (H2S or CH4).
H2 consumption experiments
Bottles (total volume 58.35 mL) were always filled with 25 mL medium. For growth on H2, D. retbaense cultures were amended with 24 mM acetate and 0.35 mL modified Wolins vitamin solution, inoculum 10% (2.5 mL) of a culture grown on lactate for 7 days. The incubations of M. halotolerans were amended with 20 mM acetate, 35 mM formate, 0.05% yeast extract, inoculum 10% (2.5 mL) of a culture grown on 80%/20% H2/CO2 for 4 days. Different amounts of H2 were added to headspace to obtain 10% (~ 4 mL), 40% (~ 11 mL) and 100% (~ 25 mL)(rest gas: N2) of the total headspace volume. Fluid and gas volume was always kept constant or defined volumes were withdrawn and considered in the calculations. Because M. halotolerans requires CO2 for methanogenesis, 5% of the gas phase was CO2, which was re-supplied after gas analysis at each sampling point. Incubation temperature was 37 °C for both strains. Each experiment was conducted twice independently. Sterile controls contained the media and all additions but were not inoculated. The original cavern brine was anoxically filled in sterile bottles and the headspace was flushed with 100% H2. Several different enrichments were set up either containing only 100% H2, , 20 mM acetate plus 0.04% yeast extract plus 100% H2 or 10% CO2 + 90% H2. All experimental conditions can be found in the supplement Table S1. Incubation temperatures was 30 °C. All bottles were stored upside down during incubation to minimize loss of H2 due to diffusion through the rubber stopper. Still, we observed diffusion through or reaction with the stoppers especially over longer incubation periods.
Sampling and calculations
Pressure measurements, gas analysis and liquid sampling were performed in regular intervals during the incubation: For D. retbaense every week, for M. halotolerans every day/every second day and for the original brine enrichment every 50 days. Pressure measurements of the experiments were conducted before and after each sampling session. When the pressure fell below 25 mbar (detection limit of the microGC), the bottles were re-pressurized with N2 (for D. retbaense) or N2/CO2 (for M. halotolerans). At some sampling points 1 mL liquid were withdrawn for pH, HPLC and cell number determination. To calculate the amount of H2 in the bottles, the ideal gas law (3) was used to correlate temperature (T in Kelvin), pressure (p) in the bottles with atmospheric pressure—water vapor pressure at the given temperature (in Pascal) and the gas constant 8.3144 (J/mol K). The measured composition of the gas phase (%) and the known volume of the bottle (58.3 mL—liquid medium + additions) was used to calculate gas volume of H2 the bottles containing the medium V (in m3).
The resulting mol of H2 was calculated into mL by assuming STP (Standard temperature and pressure) (1 mol = 22.4 L). For each sample we calculated the volume of H2 before and after the sampling procedure by measuring headspace pressure at the beginning and at the end of the sampling. The loss of H2 in between is related to withdrawal of gaseous and liquid samples. This loss through sampling was calculated and subtracted from the calculated first pressure value to obtain the volume of consumed H2. Absolute maximum rates were obtained by calculating the slope of H2 loss during days. Relative rates were obtained by normalizing the H2 values to the initial starting value, resulting in %-loss over time.
Analytical methods
Gas composition was measured with a micro gas chromatography (microGC) 490 (Agilent) by directly measuring the gas in the headspace of the serum bottles. Pressure in the bottles was measured using a pressure sensor from Sensortechnics 0-3 barg Press D/C 2916 with an individual set-up for direct measurement of the headspace of serum bottles. Liquid samples were analyzed by using liquid chromatography of Agilent 1260II UHPLC equipped with a Flexible pump, autosampler, 1260 RI, and 1260 DA HS detectors. All analytes were identified and quantified based on their respective reference standard calibration curves. For determining cell numbers, we isolated DNA from 1 mL of sample which was withdrawn using syringes and centrifuged for 20 min at 13,000 rpm. The pellet was frozen for several hours at − 80 °C and after that at − 20 °C. This procedure was necessary because the pellets did not freeze at − 20 °C due to the high salt content. Prior to DNA isolation, the pellet was ultrasonicated for 5 min, frozen for several hours at − 80 °C, and again ultrasonicated 5 min. DNA was isolated using the Blood&Tissue Kit (Qiagen) following manufacturer’s instructions. Copy numbers were measured via digital droplet PCR (ddPCR; BioRad) using Dsr1 primer for sulphate-reducing bacteria24 or standard 16S rRNA Archaea25. The ddPCR reactions were run with a total volume of 20 µL on a DX200 instrument (BioRad) using 1 × EvaGreen supermix (BioRad) and 250 nM (final concentration) of primers. Complete PCR reactions were emulsified with QX200 Droplet Generation Oil for EvaGreen using the QX200 Droplet Generator and then transferred to a 96-well plate. PCR reactions were performed in a C1000 Touch Thermocycler with deep-well module (BioRad) using the following program: 95 °C for 15 min, 40 cycles of 95 °C for 30 s, 57.1 °C (sulphate-reducing bacteria) or 63.1 °C (archaea) for 1 min, 4 °C for 5 min, 90 °C for 10 min and finally an infinite hold at 4 °C. Plates were equilibrated to room temperature for at least 10 min before being analysed on a QX200 Droplet Reader (BioRad). Thresholds for positive and negative droplets were manually set using positive (Bacteria or Archaea cultures) and negative (ultra-pure water) controls. The copy numbers were calculated to cell numbers based on the available genome of D.retbaense with 1 gene copy of dsr1/cell26. The genome of M. halotolerans is not available and therefore we assume 1.7 gene copy numbers/cell of 16S rRNA in general Archaea based on the rrnDB database (value taken on the 24.10.2022).
Results
Hydrogen consumption during halophilic sulphate reduction
The halophilic sulphate-reducing bacteria Desulfohalobium retbaense consumed H2 in the headspace over several weeks. In the 10% H2 bottles the cells consumed all available H2 (around 4 mL H2) after 20 days while the incubations with 40% H2 (initial 11 mL H2) and 100% H2 (initial 25 mL H2) H2-headspace were only partially consumed (Fig. 1A). We observed maximum rates of 0.31, 0.44 and 0.62 mL/day for 10, 40 and 100% H2 respectively. In the set-ups with 10% H2, we added 10% H2 at day 35 after the first consumption but subsequently consumption rate was significantly slower with a maximum rate of 0.09 mL/day. In total D. retbaense consumed 3.8 + 1.8 (initial 10% H2 + renewed addition of 10% H2), 7.5 (40% H2 set-up) and 14.3 (100% H2 set-up) mL of H2, which correspond to the relative amounts of 99% + 52%, 65% and 57% H2 (Fig. 1B). The experiment was repeated twice with a standard deviation variation of the maximum rates between 7 and 23% (see all data in suppl. Table 2). In sterile controls 0 mL for 10% H2 , 0.9 mL for 40% H2 and 1.5 mL for 100% H2 were lost through diffusion out of the rubber stopper or by other chemical reactions, which shows that some loss occurs abiotically. During the incubation, either no or only minor amounts of acetate were consumed (0 mM with 10% H2, 0.7 mM with 40% H2 and 1.1 mM with 100% H2) (Table 1), which shows that acetate was not limiting. From an initial 2.80E+10 cells/mL, cell numbers did only slightly increase with all H2 concentrations to around 4E+10 cells/mL indicating that the strain was not able to build up substantially more cell mass.

(a) Consumed hydrogen of Desulfohalobium retbaense in ml over time measured in the headspace of bottles incubated at near atmospheric pressure and 37 °C. Bottles were amended with 100% H2 (filled circle, solid line), 40% H2 (filled triangle, solid line) and 10% H2 (open square, solid line). The star * at 35 days indicates re-feeding the bottles with 10% H2. pH measured at the day are given above the line. (b) Values plotted in loss in % over time. Error bars indicate deviation from the mean of duplicates. At day 35, 10% of H2 was again added to the 10% H2 bottles (dashed line—2nd addition).
Only minor amounts of H2S were detected in the headspace with a maximum value of around 4000 ppm at day 32 in one duplicate of the 40% H2 bottles. However, at the next sampling stage the H2S concentration was again back to the background level (suppl. Fig. S1). This indicates a changing H2S/HS− dissolution in the liquid, which was confirmed by pH measurements, showing a significant increase in pH over time. Starting from pH 7.5 at day 0 and resulting in a final pH of 8.9 for 10% H2 and pH 9.1–9.2 for the 40% H2 and 100% H2 bottles (Fig. 1A; Table 1, all pH values over time can be found in supplemental Table 3). Sulphate was not quantified but was in excess (21 mM) in the media allowing for theoretical consumption of 47 mL H2. We therefore assume that sulphate was never limiting.
Hydrogen consumption during methanogenesis
The halophilic methanogen Methanocalculus halotolerans consumed H2 much faster compared to D. retbaense but still slow compared to other reported methanogenic growth27,28. In all set-ups all H2 was consumed (Fig. 2A) with maximum rates of 1.1, 4.1 and 4.7 mL/day for the 10% H2, 40% H2 and 90% H2 set-ups. Re-addition of 10% H2 in the 10% H2 bottles led to an increased activity with a consumption rate of 1.7 mL/day (Fig. 2B). The experiment was conducted twice with a standard deviation between maximum rates between 15 and 37%. CH4 was produced accordingly in all set-ups with consumption of CO2 (suppl. Fig. S2). CO2 was always re-added when values dropped below 1.5%, to not limit growth. Formate was completely consumed after 1 day in all set-ups also the non-hydrogen controls. Acetate was consumed only in low concentrations for the 10% H2 (0.2 mM acetate consumed) and 40% H2 bottles (0.5 mM acetate consumed). 2.2 mM acetate were consumed in the 90% H2 bottles (Table 1). Cells numbers increased from 7.2E+09 to 4E+10 cells/mL for the 10% H2, to 4.5E+10 cells/mL for the 40% H2 and 5.1E+10 cells/mL and 90% H2 set-ups. Similar to D. retbaense, pH increased in all the M. calculus cultures. Starting from a pH of 7.2, it increased to pH 8.6, 8.5 and 8.8 (Table 1; Fig. 2A; Supplemental Table 3).

(a) Consumed hydrogen of Methanocalculus halotolerans in mL over time measurable in the headspace of bottles incubated at near atmospheric pressure and 37 °C. Bottles were amended with 90% H2 (filled circle, solid line), 40% H2 (filled triangle, solid line) and 10% H2 (open square , solid line). The star * at 35 days indicates re-feeding the bottles with 10% H2. pH measured at the day are given above the lines (10% H2 day 5 is under line). (b) Values plotted in loss in % over time. At day 3, 10% H2 was re-added in the 10% set-ups (dashed line—2nd addition). Error bars indicate deviation from the mean of duplicates.
Hydrogen consumption of a real salt cavern brine community
The sampled cavern is located in Northern Germany in the Permian Zechstein Group salt layer. It has been filled with brine since the initial leaching several decades ago. The brine has a salinity of 27%, pH 7.4 and a high sulphate content of 4190 ± 57 mg/L. The pure cavern brine was incubated in serum bottles with a headspace of 100% H2. We incubated for 176 days at 30 °C which is relevant for this specific cavern. We added acetate and yeast extract to one set of bottles to trigger microbial activity, the other set was pure brine without any additional supplements. Sterile water with 100% H2 lost 1.6 mL of H2 during the incubation of 211 days. The brine sample incubated with pure 100% H2, consumed 1.7 mL H2 on average (Fig. 3A). One bottle showed significantly lower total H2 loss compared to the duplicate, although initial consumption rate was similar. In contrast with addition of acetate and yeast, 5.3 mL H2 (corresponding to ~ 11%) (Fig. 3B) was consumed in both bottles with the maximum rate of 0.069 mL/day and a total of 0.9 mM acetate was consumed after 136 days. At day 100 no acetate was consumed although sulphate-reduction was already visible. This indicates that the first carbon from either the brine or the yeast extract was used before acetate was consumed. H2S was first detected after 94 days with a maximum of 1505 ppm at day 136 in the H2 + acetate + yeast bottles and declined afterwards due to the increase in pH. In the pure H2 bottles, H2S was not detected in the headspace, but black precipitates were visible on the glass wall, which indicates possible FeS formation caused by H2S reacting with ferrous iron ions. pH first decreased to around 7 and afterwards increased to 8.5 in the H2 + acetate + yeast bottles and to pH 7.8 in the pure H2 bottles. Interestingly at the last sampling point (after 176 days), trace amounts of CH4 (< 0.1%) were detected in one incubation of the acetate + yeast + H2 experiment. CO2 concentrations were always below 0.1%. It is possible that the CH4 production is attributed to acetogenic or methylotrophic methanogens or the detected trace amounts of CO2 are indeed sufficient. Further investigations are needed to confirm this observation.

(a) Consumed hydrogen of an original salt cavern brine in mL over time measurable in the headspace of bottles incubated at near atmospheric pressure and 30 °C. Bottles were amended with 100% hydrogen (emty triangle, solid line) or with addition of 20 mM acetate and 0.04% yeast extract (solid triangle, solid line). H2S measured in ppm in the headspace is given on the secondary axis for the acetate + yeast extract bottles (red line). Diffusion loss in an only sterile lab water bottle is shown with filled circle, dashed line. (b) Values plotted in % loss. pH values are given at the single sampling points in the graph. Error bars indicate deviation from the mean of duplicates.
Cultures containing 10% CO2 + 90% H2 were established to investigate activities of hydrogenotrophic methanogens and acetogens, but no CH4 or acetate production was observed at any time. It must be mentioned that the addition of CO2 led to a pH decrease to pH 6.5 because of the lack of buffering compounds in the brine, which might have affected the microbial community.
Discussion
Hydrogen consumption of halophilic microbes
With the increased interest in storing H2 in the subsurface and especially in salt caverns, it becomes important to understand the risks of microbial H2 consumption during storage within high-salt environments. Many Bacteria and Archaea can live and even thrive under high salt concentrations. Although high salinity environments are hostile for many groups of microorganisms12,29, diverse microbial communities can still be found in high-saline environments like salt lakes and also salt caverns30. However, a distinction between halophilic and halotolerant must be made. Halophiles range from slightly to moderate to extreme require a certain amount of salt to grow and show their optimum growth behavior typically at salinities above 100 g/L (Oren, 2011) and can tolerate up to extreme ranges (> 200 g/L). Halotolerant microbes do not need high salt concentrations but can tolerate it to a certain degree. Salinity is a common stress factor and microorganisms have different strategies to adapt to the osmotic pressure imposed by the ionic strength of the surrounding environment31. Both strategies, the “salt-in”- or the “osmotic solutes” strategy, require a certain amount of energy (ATP) to uphold cell activity. Either ATP is used directly in the production of compatible solutes (Oren, 2006), or ATP is required to uphold the correct sodium and potassium gradient between the inside and the outside of the cell. This is also the reason why many halophilic microbes living at the “energetic edge”, using much energy for upholding their cell stability, tend to grow slower compared to non-halophilic counterparts. Therefore, it is very important to specifically assess H2 oxidation by halophilic H2-oxidizers because the relative kinetic rates in such environments are largely unknown.
To estimate the H2 consumption rates and associated effects in high-salt subsurface environments, we investigated and cultured two halophilic microorganism which belong to the two most relevant metabolic groups regarding H2 consumption: (i) Sulphate reduction is a very widespread metabolism with many different microbial groups potentially using H2 as electron donor in the presence of sulphate as electron acceptor32, resulting in the production of the toxic and corrosive gas H2S; (ii) Methanogenesis from H2 and CO2 is another highly relevant H2 consumption process, which recently received major attention for potentially producing “green” methane (biomethanation) in the subsurface after injecting renewable H2 with captured CO233. The two investigated strains in our study are halophilic with Desulfohalobium retbaense at a salt optimum at 12% growing up to 24%22 and Methanocalculus halotolerans with the optimum of 5% growing up to 12.5% salinity23. Both strains were originally described to be able to use H2 for growth, both requiring acetate as an additional carbon source. To estimate the potential H2 consumption kinetics in the high-saline subsurface we carefully cultured the strains with different volumes of H2 in the headspace. We clearly observed H2 consumption over time with increasing consumption rates with increasing H2 concentrations. When comparing the maximum rates, we observe only a doubling of rate (0.6 mL/day) for 100% H2 compared to 10% H2 (0.3 mL/day). Interestingly, when looking at the H2 loss relative to the initial concentration, growing with initial 10% H2 instead of 40% or 100% has the highest relative rate (= loss in %). Providing all nutrients in excess and starting with a high cell number, we would have expected a much more pronounced consumption with 100% H2. This observation might be due to physiological constrains, like limitations of H2 transport to and into the cell. H2 is normally a very scarce electron donor in the environment, typically in the nmol range. So high amounts of H2 could have a negative effect on cellular H2 uptake or involved hydrogenase enzymes but details need to be further investigated. Overall, D. retbaense was not able to oxidize all the provided H2 in the 40% and 100% H2 set-ups although acetate and sulphate was provided in excess. This lack of continued activity or consumption can be explained by an intense pH increase during H2-oxidation. It can be seen in the reaction rate of H2-dependent sulphate reduction (1) that this reaction is a proton consuming process leading to an increase in surrounding pH34,35. This also explains the near complete absence of gaseous H2S throughout our study. At higher pH H2S will be in its highly soluble form HS− (HS− + H+ ⇋ H2S). HS− is a weak acid which should partly counteract the pH increase but clearly the intense H2 oxidation is the main driver and overrides the HS− effect. In biogas plants and biocorrosion studies it is commonly reported that intense sulphate-reduction can lead to a pH increase36,37. As biogas and corrosion involves also intermediate H2 release, the observed pH increase might be also a direct effect of microbial H2 oxidation. Although it has been long known that H2 is an electron donor for SRMs, it was never clearly described that the significant pH effect limits the microbial activity when growing on pure H2. As the pH approached pH 9, which is beyond the acceptable limit for D. retbaense, its activity ceased. Another inhibiting effect could be significant amounts of dissolved HS− (theoretically based on the amount of H2 consumed: 1.4, 1.9 and 3.6 mL for the 2 × 10% H2, 40% H2 and 100% H2 respectively), which might have an additional inhibitory effect on the activity although assume that this inhibition is rather small as sulphate reducers tend to have a high tolerance for HS−. When growing on lactate as an electron donor, pH did not rise significantly with an end value of pH 7.8 after 7 days with H2S concentrations far beyond 15% of the headspace (suppl. Table 4). We assume that this is due to the production of the organic acid acetate and CO238, which both will lead to a decrease in pH. So, the observed pH increase in culture growing on only H2 is purely associated to the enzymatic process of H2 oxidation.
We were however not able to correlate the overall volume of H2 consumed with the pH. In the 40% H2 set-up the strains consumed 6.6 mL (consumption minus the diffusion loss observed in the sterile controls) and in the 100% H2 set-up it consumed almost double with 12.4 mL and both reached a pH 9.1–9.2. One possible explanation could be that under 100% H2 atmosphere more H2 is dissolved in the media. H2 has a low solubility in water (0.0014 g gas per kg water at 37 °C) and even lower in saline brine. So, growth will be limited by H2 dissolution, which will be increased with higher H2 concentration or pressure. Given the optimal growing conditions in the beginning (optimal pH and nutrients) and a high initial cell number, 100% H2 gives the cells more electron donor to be active in the initial growth phase compared to 40% H2. Also, concurrent chemical reactions of the H2 gas with the slightly alkaline media, leading to additional abiotic H2 loss, could be factor. Another reason could be that the increasing alkaline stress will trigger expression of pH-homeostasis related genes in D.retbaense cells. For Desulfovibrio vulgaris it was shown that several genes for ATP synthase, Na+/H+ antiporters and amino acid metabolism were expressed after an alkaline shock, which is a strategy to retain the cellular redox state and avoid an inverse pH gradient over the cell membranes39,40. These stress adaptations might also play a role in our tested strain and higher H2 concentrations might trigger a stronger or more complex gene expression, keeping the pH more stable. Further careful investigations combined with chemical and biological kinetic modelling and even including transcriptomic studies will help to understand the H2- and proton consumption.
The H2 electron chain towards sulphate is well understood and relatively conserved in members of the Deltaproteobacteria41, therefore it can be assumed that the observed effects of pH increase when grown on H2 is a general phenomenon and not strain specific.
Similarly, methanogenesis is a proton consuming process (reaction rate 2). Also, during methanogenesis the Archaea are consuming bicarbonate/CO2 from the media, which reduces buffer potential and increases pH as described previously42. We observed in our set-ups with M. halotolerans a pH increase, although not as intense as with D. retbaense. All H2 was consumed in all cases although activity rates decreased during the end of incubation for the 90% H2 bottles probably because pH increased and was close to the upper pH limit of this strain. Although the absolute rates were highest with the highest amount of H2, the relative consumption rates (= loss in %) with 10% H2 were the fastest and renewed addition of 10% H2 gave an even increased rate which is related to either increased cell numbers or increased activity. If this effect of higher relative rates with lower H2 concentrations will have an effect on the storage site, needs to be evaluated in the future.
Potential risks for souring of salt caverns
Halophilic archaea and bacteria have been isolated from pure halite crystals in salt mines43 and it is therefore expected that that also salt caverns will be habitat to a diverse set of microbes. Recent studies of salt cavern microbiology showed microbes in all studied caverns10. Also, our studied salt cavern brine was microbiologically active and after several months of incubation with and without nutrient addition we observed H2 consumption. With addition of the growth booster yeast extract (supplying microbes with a variety of vitamins and trace elements) and the carbon source acetate, a significant H2 consumption was measurable together with acetate consumption, H2S production and also a significant pH increase. At the end of the incubation the pH increased 1 log factor from an initial 7.5 to 8.5. This shows that H2 oxidation by halophilic SRMs does indeed lead to a significant pH increase similarly to what we observed with our type strain D. retbaense. In the pure brine bottles without addition of carbon source or yeast extract, H2 consumption was less pronounced but still the presence of black iron sulfide minerals and a pH increase suggest that sulphate-reduction was on-going. This shows that the natural community is limited by a certain growth factor, trace element, vitamin or similar. We hope to be able to identify and enrich the sulphate-reducing community in future studies to better understand the growth behavior.
If we assume that the studied sulphate reducers will also be active in the salt cavern when in contact with H2, there is indeed a real risk for H2 loss and souring. The sampled cavern brine has a very high sulphate content of over 4000 mg/L (~ 44 mM), which gives sulphate-reducing organisms sufficient electron acceptors. A back of the envelope calculation of a typical salt cavern with a volume of 600.000 cubic meters (most volume will be occupied with gas but roughly 3000 m3 will be brine in the sump together with insoluble minerals) contains 12.6 metric tons of sulphate in the cavern to be potentially converted by sulphate-reducing organisms. However, in case of 100% sulphate conversion without any additional input and no inhibition/limitation, the cavern could contain around 3E+06 L liters of H2S or only 0.6% of the total cavern volume. Based on the equilibrium (1), four times the amount of H2 would be needed as electron source (1.2E+07 L), a loss of around 2% of the total volume. These values will depend on many factors including the physical cavern properties, the brine chemistry and our observed limiting effects of nutrients and pH. The consumption rates will firstly be dependent on the surface area of the brine-gas interphase and therefore the cavern shape. A higher surface area will allow for a higher H2 availability for the microbes. An important limiting factor will be the amount of available carbon source and/or growth factors, which led to a H2 loss of maximum 5–13% under laboratory conditions. Total inorganic carbon has been detected in the cavern brine of around 85 mg/L and some organic carbon of around 7.7 mg/L which was most likely introduced by the leaching process using diesel oil as a leaching blanket (personal communication with the cavern operators). This method of using diesel oil during leaching is/was very common and some organic carbon can be expected in most if not all salt-caverns. Other sources of organic carbon might also include the originally used leaching water (often sea water) or the presence of small amounts of organic compounds in the salt rock. Still, additional growth enhancers were necessary for significant sulphate reduction to occur, which slowed down when pH approached 9. This means that during the first H2 storage cycles, some volumes of H2 could be converted by microbes until the pH value will be outside of the optimum for these microbes and we speculate that the overall %-loss of H2 in a salt cavern due to microbial activity will be relatively low. As a cautionary note: the derived laboratory enrichments can only give hints about the metabolic potential in extreme environments since most microbes do not grow under lab conditions. A much more complex and chemolithoautotrophic community with higher cell numbers might be present and active in the cavern leading to a) faster H2 consumption and b) longer and more intense H2 consumption. For example, it could be that alkali-tolerant strains will take over H2 oxidation as soon as neutrophilic strains reached their limit or a certain adaptation to higher pH will occur. Indeed, given the production of methane after 170 days when pH was seemingly too high for the sulphate-reducing community, methanogenesis is inferred. Until the end of the experiment, we were not able to stimulate methanogenesis or acetogenesis directly by adding CO2 in our lab enrichments. As the brine seems to be very sensitive to pH changes due to low buffering material, addition of CO2 led to a strong pH decrease. However, the fact that we observed methane in our H2 + acetate + yeast shows that the microbial metabolism is possible and might cause a continued H2-loss after sulphate reduction. Based on our results with M. halotolerans it also seems that methanogens might be able to better cope with the pH increase, as the strain was able to consume all provided H2 even at pH > 8.5. This complex but slow interplay of the cavern community when in contact with H2 needs to be further studied.
Our data shows that artificially leached salt cavern can contain H2-consuming microbes, especially sulphate-reducers which seem to be nutrient limited and additionally will significantly increase pH. Field tests are now required to understand if the observed effects regarding H2 consumption and pH will also occur in the cavern itself.
Conclusions
-
Halophilic hydrogen-consuming sulphate reducers and methanogens are able to consume significant volumes of H2 over time.
-
H2 oxidation leads to a significant pH increase in both cases. In case of the tested pure strain SRM, pH exceeds growth limits and H2 is not completely consumed.
-
Original salt cavern brine contains active SRMs which are able to consume parts of the available H2 especially when carbon and nutrients were added. H2S gas was measurable in the headspace but decreased again when brine pH increased. Some produced H2S also precipitated with trace amounts of iron as iron-sulfide minerals
-
The pH increase caused by H2 oxidizing-SRMs might limit microbial H2 consumption over long-term and might therefore be a self-limiting process in low-buffered environments like salt cavern brines.
Data availability
All main data generated or analysed during this study are included in this published article (and its Supplementary Information files).
References
Lott, M. C., Kim, S.-I., Tam, C., Houssin, D. & Gagné, J. Technology Roadmap: Energy Storage (International Energy Agency: Paris, 2014).
Yue, M. et al. Hydrogen energy systems: A critical review of technologies, applications, trends and challenges. Renew. Sustain. Energy Rev. 146, 111180 (2021).
Krevor, S. et al. Subsurface carbon dioxide and hydrogen storage for a sustainable energy future. Nat. Rev. Earth Environ. 66, 1–17 (2023).
Ball, M., Basile, A. & Veziroglu, T. N. Compendium of Hydrogen Energy: Hydrogen Use, Safety and the Hydrogen Economy (Woodhead Publishing, 2015).
Tarkowski, R. Underground hydrogen storage: Characteristics and prospects. Renew. Sustain. Energy Rev. 105, 86–94 (2019).
Raza, A. et al. A holistic overview of underground hydrogen storage: Influencing factors, current understanding, and outlook. Fuel 330, 125636 (2022).
Kruck, O., Crotogino, F., Prelicz, R. & Rudolph, T. Assessment of the potential, the actors and relevant business cases for large scale and seasonal storage of renewable electricity by hydrogen underground storage in Europe. HyUnder Deliv. 6, 66 (2013).
Bordenave, S., Chatterjee, I. & Voordouw, G. Microbial community structure and microbial activities related to CO2 storage capacities of a salt cavern. Int. Biodeterior. Biodegrad. 81, 82–87 (2013).
Laban, M. Hydrogen Storage in Salt Caverns: Chemical Modelling and Analysis of Large-Scale Hydrogen Storage in Underground Salt Caverns (Master thesis Delft University of Technology, 2020).
Schwab, L. et al. Structural analysis of microbiomes from salt caverns used for underground gas storage. Int. J. Hydrog. Energy 6, 66 (2022).
Jaakkola, S. T., Ravantti, J. J., Oksanen, H. M. & Bamford, D. H. Buried alive: Microbes from ancient halite. Trends Microbiol. 24, 148–160 (2016).
Oren, A. Microbial life at high salt concentrations: Phylogenetic and metabolic diversity. Saline Syst. 4, 2 (2008).
Oren, A. Thermodynamic limits to microbial life at high salt concentrations. Environ. Microbiol. 13, 1908–1923 (2011).
Gregory, S. P., Barnett, M. J., Field, L. P. & Milodowski, A. E. Subsurface microbial hydrogen cycling: Natural occurrence and implications for industry. Microorganisms 7, 53 (2019).
Greening, C. et al. Genomic and metagenomic surveys of hydrogenase distribution indicate H2 is a widely utilised energy source for microbial growth and survival. ISME J. 10, 761–777 (2016).
Dopffel, N., Jansen, S. & Gerritse, J. Microbial side effects of underground hydrogen storage–Knowledge gaps, risks and opportunities for successful implementation. Int. J. Hydrog. Energy 6, 66 (2021).
Panfilov, M. in Compendium of Hydrogen Energy 91–115 (Elsevier, 2016).
RAG. UNDERGROUND SUN STORAGE—Chemical Storage of Renewable Energy in Porous Subsurface Reservoirs with Exemplary Testbed. www.underground-sun-storage.at (2017).
Haddad, P. et al. Geological storage of hydrogen in deep aquifers—An experimental multidisciplinary study. Energy Environ. Sci. 15, 3400–3415 (2022).
Hemme, C. & van Berk, W. Potential risk of H2S generation and release in salt cavern gas storage. J. Nat. Gas Sci. Eng. 47, 114–123 (2017).
Veshareh, M. J., Thaysen, E. M. & Nick, H. M. Feasibility of hydrogen storage in depleted hydrocarbon chalk reservoirs: Assessment of biochemical and chemical effects. Appl. Energy 323, 119575 (2022).
Ollivier, B., Hatchikian, C., Prensier, G., Guezennec, J. & Garcia, J.-L. Desulfohalobium retbaense gen. nov., sp. nov., a halophilic sulfate-reducing bacterium from sediments of a hypersaline lake in Senegal. Int. J. Syst. Evol. Microbiol. 41, 74–81 (1991).
Ollivier, B. et al. Methanocalculus halotolerans gen. nov., sp. nov., isolated from an oil-producing well. Int. J. Syst. Evol. Microbiol. 48, 821–828 (1998).
Kondo, R., Nedwell, D. B., Purdy, K. J. & Silva, S. Q. Detection and enumeration of sulphate-reducing bacteria in estuarine sediments by competitive PCR. Geomicrobiol. J. 21, 145–157 (2004).
Ovreås, L., Forney, L., Daae, F. L. & Torsvik, V. Distribution of bacterioplankton in meromictic Lake Saelenvannet, as determined by denaturing gradient gel electrophoresis of PCR-amplified gene fragments coding for 16S rRNA. Appl. Environ. Microbiol. 63, 3367–3373 (1997).
Spring, S. et al. Complete genome sequence of Desulfohalobium retbaense type strain (HR100T). Stand Genomic Sci. 2, 38–48 (2010).
Mickol, R. L., Laird, S. K. & Kral, T. A. Non-psychrophilic methanogens capable of growth following long-term extreme temperature changes, with application to Mars. Microorganisms 6, 34 (2018).
Han, W., He, P., Lin, Y., Shao, L. & Lü, F. A methanogenic consortium was active and exhibited long-term survival in an extremely acidified thermophilic bioreactor. Front. Microbiol. 10, 2757 (2019).
Oren, A. Life at high salt concentrations. The Prokaryotes 3, 263–282 (2006).
Oren, A. Diversity of halophilic microorganisms: Environments, phylogeny, physiology, and applications. J. Ind. Microbiol. Biotechnol. 28, 56–63 (2002).
Galinski, E. A. & Trüper, H. G. Microbial behaviour in salt-stressed ecosystems. FEMS Microbiol. Rev. 15, 95–108 (1994).
Matias, P. M., Pereira, I. s. A., Soares, C. M. & Carrondo, M. A. Sulphate respiration from hydrogen in Desulfovibrio bacteria: A structural biology overview. Prog. Biophys. Mol. Biol. 89, 292–329 (2005).
Strobel, G., Hagemann, B., Huppertz, T. M. & Ganzer, L. Underground bio-methanation: Concept and potential. Renew. Sustain. Energy Rev. 123, 109747 (2020).
Lens, P. & Kuenen, J. G. The biological sulfur cycle: Novel opportunities for environmental biotechnology. Water Sci. Technol. 44, 57–66 (2001).
Liamleam, W. & Annachhatre, A. P. Electron donors for biological sulfate reduction. Biotechnol. Adv. 25, 452–463 (2007).
Shan, L., Zhang, Z., Yu, Y., Ambuchi, J. J. & Feng, Y. Performance of CSTR–EGSB–SBR system for treating sulfate-rich cellulosic ethanol wastewater and microbial community analysis. Environ. Sci. Pollut. Res. 24, 14387–14395 (2017).
Pankhania, I. P. Hydrogen metabolism in sulphate-reducing bacteria and its role in anaerobic corrosion. Biofouling 1, 27–47 (1988).
Keller, K. L. et al. New model for electron flow for sulfate reduction in Desulfovibrio alaskensis G20. Appl. Environ. Microbiol. 80, 855–868 (2014).
Stolyar, S. et al. Response of Desulfovibrio vulgaris to alkaline stress. J. Bacteriol. 189, 8944–8952 (2007).
Sydow, U., Wohland, P., Wolke, I. & Cypionka, H. Bioenergetics of the alkaliphilic sulfate-reducing bacterium Desulfonatronovibrio hydrogenovorans. Microbiology 148, 853–860 (2002).
Pereira, I. A. C. et al. A comparative genomic analysis of energy metabolism in sulfate reducing bacteria and archaea. Front. Microbiol. 2, 69 (2011).
Delgado, A. G., Parameswaran, P., Fajardo-Williams, D., Halden, R. U. & Krajmalnik-Brown, R. Role of bicarbonate as a pH buffer and electron sink in microbial dechlorination of chloroethenes. Microb. Cell Fact. 11, 1–11 (2012).
Vreeland, R. H., Rosenzweig, W. D. & Powers, D. W. Isolation of a 250 million-year-old halotolerant bacterium from a primary salt crystal. Nature 407, 897–900 (2000).
Acknowledgements
We want to thank Equinor for funding this project and providing salt cavern brine for the experimental studies. We also want to thank Carsten Reekers, Nils Kruse and Michael Mollenhauer for their help in the project and the good discussions.
Author information
Authors and Affiliations
Contributions
ND: conceptualization, methodology, experiments, writing. KM: cell number analysis. AK: water and HPLC analysis. DB: sampling. EA: writing and editing. BAS: writing and editing. KD: lab support. JB: conceptualization. SH: conceptualization, writing, acquiring internal grant.
Corresponding author
Ethics declarations
Competing interests
The authors D. Boldt, J. Beeder and S. Hoth are employed by Equinor, which is a company working on hydrogen storage. The other authors declare no competing interests.
Additional information
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Dopffel, N., Mayers, K., Kedir, A. et al. Microbial hydrogen consumption leads to a significant pH increase under high-saline-conditions: implications for hydrogen storage in salt caverns. Sci Rep 13, 10564 (2023). https://doi.org/10.1038/s41598-023-37630-y
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-023-37630-y
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
