
Abstract
Obesity contributes to physical comorbidities and mental health consequences. We explored whether physical activity could influence more than metabolic regulation and result in psychological benefits through the brain-gut microbiome (BGM) system in a population with high BMI. Fecal samples were obtained for 16 s rRNA profiling and fecal metabolomics, along with psychological and physical activity questionnaires. Whole brain resting-state functional MRI was acquired, and brain connectivity metrics were calculated. Higher physical activity was significantly associated with increased connectivity in inhibitory appetite control brain regions, while lower physical activity was associated with increased emotional regulation network connections. Higher physical activity was also associated with microbiome and metabolite signatures protective towards mental health and metabolic derangements. The greater resilience and coping, and lower levels of food addiction seen with higher physical activity, may be explained by BGM system differences. These novel findings provide an emphasis on the psychological and resilience benefits of physical activity, beyond metabolic regulation and these influences seem to be related to BGM interactions.
Similar content being viewed by others

Introduction
In the past few decades, obesity rates have rapidly grown to epidemic proportions, with an estimated 650 million adults considered obese1. High BMI leads not only to the development of physical comorbidities, but also to mental health consequences, in a bidirectional relationship that likely explains the high comorbidity that is seen between obesity and the development of psychiatric disorders2. Individuals who have reported weight-related discrimination are more likely to engage in high-risk behaviors such as drug abuse and cigarette smoking, and also tend to experience greater weight gain over time3. Additionally, there is a greater likelihood of developing maladaptive eating behaviors, including food addiction, binge-eating, emotional eating, and increased consumption of calories4,5.
In individuals with higher BMI, modifiable factors such as resilience may serve a protective function against the predisposition of developing psychiatric disorders with obesity6. Resilience is defined as the ability to positively adapt in response to significant adversity or stressors, and develops via interplay between genetics, environmental factors, and social support systems7. Studies have shown that emotional resilience is protective against the development of obesity regardless of income, through positive associations with healthier dietary choices and moderating perceived stress and binge eating behavior8,9. In adults, physical activity (PA) is a well-recognized contributor to psychological resilience by blunting stress reactivity, protecting against the metabolic consequences of stress-inducing events, and promoting an anti-inflammatory state10,11,12. While some studies have been done on the individual physiological changes associated with PA in high BMI populations, there are a limited number of studies on the interactions between PA and various psychological variables in the context of the brain-gut microbiome (BGM) system.
A growing body of studies support the role of the BGM axis in the pathophysiology of obesity, mediated by alterations in metabolic, enteroendocrine, and neural signaling13. Having high BMI has been associated with changes in the diversity and composition of the gut microbiota, which lead to disruptions in the downstream metabolites and gut-endocrine signals that orchestrate energy homeostasis14. For example, obesity was found to be associated with increases in the Prevotella/Bacteroides ratio and decreases in fecal tryptophan levels, which is a metabolite related to the biosynthesis of serotonin15. Signals from the microbiome may thus also alter neural processes, with individuals with high BMI demonstrating alterations in reward and emotional regulation brain regions, which have also been linked to clinical measures such as food addiction, involving continued consumption of palatable foods despite meeting homeostatic energy requirements16,17. While the effects of PA on the brain, microbiome, and metabolites may have been examined independently, there is a lack of studies that utilize a systems-biology approach to study the effects of PA within the BGM as an integrated system, while incorporating psychosocial variables in the context of obesity. In this study, we predict that there are distinct brain, gut microbiome, and metabolite signatures based on PA, and that these BGM system associations modulate positive psychological changes in a population with high BMI (summarized in Fig. 1).

Study Overview, Workflow, and Results. Denotes the overview of workflow and analyses, and summarizes the findings from the study.
Methods
The sample recruited was comprised of 92 participants, with the absence of significant medical or psychiatric conditions. All procedures were performed in accordance with the relevant guidelines and regulations and were approved by the Institutional Review Board (16-000187, 16-000281) at the University of California, Los Angeles's Office of Protection for Research Subjects. All participants provided written informed consent.
Participants
Participants were selected as described in our previous studies15. Participants were excluded for the following: pregnant or lactating, substance use, abdominal surgery, tobacco dependence (half a pack or more daily), extreme strenuous exercise (> 8 h of continuous exercise per week such as marathon runners), current or past psychiatric illness and major medical or neurological conditions. Participants taking medications that interfere with the central nervous system or regular use of analgesic drugs were excluded. Because of the effect of handedness on fMRI activation, only right-handed participants were included to negate handedness as a cofounder. To avoid potential cofounders in microbiome analysis, included participants were also required to not have taken antibiotics or probiotics for at least 3 months before enrolling in the study. Only premenopausal females were enrolled and were scanned during the follicular phase of their menstrual cycles as determined by self-report of their last day of the cycle. Participants with hypertension, diabetes, metabolic syndrome or eating disorders were excluded to minimize confounding effects. We used body mass index (BMI) cutoffs to define our overweight (25 ≤ BMI < 30) and obese (BMI ≤ 30) groups. No participants exceeded 400lbs due to magnetic resonance imaging (MRI) scanning weight limits. Participants underwent MRI scans, anthropometrics (height, body weight, and body mass index), and fresh stool samples for 16 s ribosomal RNA gene sequencing and metabolite analysis were collected.
Questionnaires
Various questionnaires were utilized to assess participant’s physical activity levels and psychological well-being. Participants completed the validated International Physical Activity Questionnaire (IPAQ) long form, which comprises of 27 items that collected data in different domains (job-related, transport-related, domestic and leisure-time physical activity) and intensities (moderate, vigorous, walking) and includes sitting time18. The Guidelines for Data Processing and Analysis of the IPAQ categorical scoring were used to determine participants’ current level of physical activity and participants were grouped into low, moderate, or high physical activity level categories19.
Additionally, psychological resiliency was assessed using the Brief Resilience Scale (BRS)20. To assess stress, anxiety, and mood, the Hospital Anxiety and Depression Scale (HADS) was used. The HADS is a 14-item scale used to measure symptoms of anxiety and depression21. The questions are scored on a scale of 0–3, corresponding to how much the individual identifies with the question for the past week. In order to assess effective and ineffective coping strategies, participants also completed the Brief-COPE questionnaire, which is a 28 item self-report questionnaire that comprises of 14 two-item subscales including: (1) self-distraction, (2) active coping, (3) denial, (4) substance use, (5) use of emotional support, (6) use of instrumental support, (7) behavioral disengagement, (8) venting, (9) positive reframing, (10) planning, (11) humor, (12) acceptance, (13) religion, and (14) self-blame22.
Food addiction was assessed using the Yale Food Addiction Scale (YFAS) questionnaire, a 25-item scale developed to assess food addiction by assessing signs of substance-dependence symptoms in eating behavior23. This scale is based upon the substance dependence criteria as found in the DSM (e.g. tolerance [marked increase in amount; marked decrease in effect], withdrawal [agitation, anxiety, physical symptoms], and loss of control [eating to the point of feeling physical ill])23. The YFAS questionnaire is a 25-question survey that measures several aspects of food addiction behavior: food dependence, withdrawal, tolerance, continued use despite problems, time spent eating, loss of control, inability to cut down, and clinically significant impairment. Food addiction was defined as having a YFAS symptom count ≥ 3 with clinically significant impairment or distress. Clinically significant impairment or distress was defined as having a at least one positive response to the following two questions in the YFAS questionnaire: “My behavior with respect to food and eating causes significant distress” and “I experience significant problems in my ability to function effectively (daily routine, job/school, social activities, family activities, health difficulties) because of food and eating,” similar to previously published works24. The YFAS has displayed a good internal reliability (Kuder–Richardson α = 0.86)23.
All patients underwent the UCLA Diet Checklist, which is a questionnaire developed by our institution, intended to represent the diet that best reflects what participants consume on a regular basis, and has been used in our previously published works25. The specific diets incorporated into this checklist were: Standard American (characterized by high consumption of processed, frozen, and packaged foods, pasta and breads, and red meat; vegetables and fruits are not consumed in large quantities), Modified American (high consumption of whole grains including some processed, frozen, and packaged foods; red meat is consumed in limited quantities; vegetables and fruit are consumed in moderate to large quantities), Mediterranean (high consumption of fruits, vegetables, beans, nuts, and seeds; olive oil is the key monounsaturated fat source; dairy products, fish, and poultry are consumed in low to moderate amounts and little red meat is eaten), and all other diets that do not fit into the above categories (vegan, vegetarian, and gluten-free). This Diet Checklist was then internally validated against the standardized DHQ-III. For data analysis, we combined standard American and modified American diet as one category. Mediterranean, and all other diets were combined as “other” for analysis.
Microbiome: DNA extraction, 16S sequencing, alpha/beta diversity analyses, differential abundance testing
DNA from stool was extracted using the DNA Fecal Microbe Miniprep Kit (Zymo Research). The V4 region of 16S ribosomal RNA was amplified and underwent paired end sequencing on an Illumina HiSeq 2500 (San Diego, CA, USA) as previously described26. Sequences were processed through the DADA2 pipeline to generate exact amplicon sequence variants (ASVs) and taxonomy was assigned based upon the SILVA 138 database27. Microbial alpha diversity was assessed on data rarefied to equal sequencing depth applying metrics including the Chao1 index of richness and the Shannon index of evenness. Microbial composition (i.e. beta diversity) was compared across groups using robust Aitchison (a phylo-genetic distance metric) in QIIME2 and visualized with principal coordinates analysis28,29. The significance of beta diversity, adjusting for covariates, was assessed using multivariate PERMANOVA with significance determined by 100,000 permutations30. Predicted metagenomics was performed using PICRUSt2 in QIIME2 using the default settings to predict abundances of bacterial gene families annotated as KEGG orthologs (KO) based on nearest reference genomes to 16S sequences.
Differential abundance of microbes was analyzed using MaAslin2, which utilizes a generalized linear mixed model with total sum scaling normalization for microbiome data32. Predicted metagenome differences between groups was visualized through principal component analysis (PCA) and significance tested using PERMANOVA. Individual predicted genes were tested between groups using DESEq2 in R and corrected for multiple hypothesis testing using false discovery rate (FDR) correction (q < 0.05 for significance). The raw sequences can be accessed NIH NCBI BioProject (BioProject ID: PRJNA946906).
Metabolites
Using the same fecal samples as the 16S sequencing, samples were aliquoted under liquid nitrogen and then shipped to Metabolon. They were processed and analyzed as a single batch on Metabolon’s global metabolomics and bioinformatics platform. Using established protocols, data was curated by mass spectrometry as previously reported33. An untargeted metabolomics platform was used, and values were scaled and relative.
Brain: MRI acquisition
Whole brain structural and resting state functional connectivity data was collected using a 3.0 T Siemens Prisma MRI scanner (Siemens, Erlangen, Germany). Detailed information on the standardized acquisition protocols, quality control measures, and image preprocessing are provided in previously published studies15,25.
For the structural MRI acquisition, high resolution T1-weighted images were acquired: echo time/repetition time (TE/TR) = 3.26 ms/2200 ms, field of view = 220 × 220 mm, slice thickness = 1 mm, 176 slices, 256 × 256 voxel matrix, and voxel size = 0.86 × 0.86 × 1 mm25.
Whole brain resting state scans were acquired with eyes closed and an echo planar sequence with the following parameters: TE/TR = 28 ms/2000 ms, flip angle = 77°, scan duration = 10 m6s, FOV = 220 mm, slices = 40, and slice thickness = 4.0 mm25.
Brain: functional network construction
Functional brain networks were constructed as previously described15,34. To summarize, measures of region-to-region functional connectivity (Fisher transformed Pearson’s correlations) were computed using the CONN toolbox and the aCompCor method in Matlab Confounding factors such as white matter, cerebrospinal fluid, the six motion realignment parameters, and the root mean squared values of the detrended realignment estimates were regressed out for each voxel using ordinary least squares regression on the normalized, smoothed resting-state images35. Participants with RMS values over 0.25 were not included. Images were then filtered using a band-pass filter (0.008/s < f < 0.08/s) to reduce the low and high-frequency noises. Although the influence of head motion cannot be completely removed, CompCor has been shown to be particularly effective for dealing with residual motion relative to other methods31. Regions of interest were segmented with the Harvard–Oxford Subcortical atlases, the Schaefer 400 cortical atlas, and the Ascending Arousal Network brainstem atlas36,37. These atlases parceled into a total of 430 brain regions. The ROI-ROI functional connectivity between the brain regions was indexed by a matrix of Fisher Z transformed correlation coefficients reflecting the association between average temporal BOLD time series signals across all voxels in each brain region. The magnitude of the Z value represents the weights in the functional network. Permuted statistical values from ROI-to-ROI analyses were further corrected using the false discovery rate (FDR) to measure significance with p(FDR) < 0.05.
Statistical analysis
General linear model (GLM) with linear contrasts were applied to examine group differences in baseline demographic and behavioral differences (High PA vs. Low PA, High PA vs. Moderate PA, Moderate PA vs. Low PA). Means were reported with their corresponding standard deviations.
We calculated beta diversity using DEICODE plugin in QIIME 2, which accounts for sparse compositional nature of microbiome data with a robust Aitchison analysis. This method has been shown to yield higher discriminatory power compared to other common metrics, such as UniFrac or Bray-Curtis28. Alpha diversity was calculated in QIIME 2 using data rarefied to 32,303 sequences and significance was determined using Chao1 and Shannon index by analysis of variance. Association of microbial genera were evaluated using MaAslin2 in R, which uses a generalized linear mixed model with total sum scaling normalization. Differential abundance p-values were converted to q-values to adjust for multiple hypothesis testing using a false discovery rate (FDR) correction (q < 0.05 for significance).
Sparse partial least squares discriminant analysis (sPLS-DA) was conducted using the R package mixOmics as a data reduction method for the resting-state brain connectivity and metabolites separately as previously described38,39. Prior to analyses, resting-state connectivity and the metabolite datasets were preprocessed. The identification of near zero variance predictors was determined on the metabolite data and then removed with the cutoff being 50% of the values must be distinct with respect to the number of subjects. The resting-state functional connectivity and metabolomics data were scaled and centered.
For integrated analyses, significant findings from fMRI, metabolite, 16S microbiome, and clinical data were combined into one dataset, and Spearman’s correlations between datapoints were performed using the Hmisc and corrplot packages in R. All p-values were adjusted for multiple hypothesis testing using (FDR) correction (q < 0.05 for significance). A summary of the workflow can be visualized in Fig. 1.
Results
Participant characteristics and psychosocial measures
Psychosocial and behavioral characteristics of the 92 individuals (males = 24, females = 68) who are overweight or obese (mean BMI = 33.22 kg/m2, mean age = 32.84 years) are summarized in Table 1. Based on the IPAQ scoring guidelines for determining PA levels, the average total PA in the high (n = 43, males = 15, females = 28), moderate (n = 32, males = 5, females = 27), and low (n = 17, males = 4, females = 13) groups were 13,432.84 MET minutes, 5,081.70 MET minutes, and 1822.953 MET minutes respectively (p < 0.001). There were no significant differences in education or income levels between the groups, except within the high versus moderate PA comparison for education level (p = 0.05).
The high PA group had greater average BRS resilience scores (p = 0.04) and ability to cope through acceptance of reality (p = 0.04), and a significant difference compared to the low PA group.
Based on PA, there were also significant differences in multiple food addiction measures, as assessed using the Yale Food Addiction Scale (YFAS), with food craving scores being lowest with high PA group. When comparing between high vs. low PA groups, significant differences were found with the following YFAS measures: continued use (p = 0.025), giving up (p = 0.005), time spent (p < 0.001), loss of control (p = 0.01), and symptom count (p < 0.001). Significant differences were also seen between moderate vs. low PA for the following YFAS measures: tolerance (p = 0.04), continued use (p = 0.04), time spent (p = 0.002), loss of control (p = 0.02), and symptom count (p = 0.003; Table 1). There were no differences in macronutrient intake, including energy (kcal), fat (grams), carbohydrate (grams), protein (grams), and cholesterol (mg) when comparing between the PA level groups (Supplemental Table 1).
PA differentiates brain functional connectivity
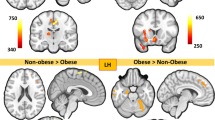
After adjusting for confounding variables such as age, sex, BMI, and diet, a sPLS-DA of brain functional connectivity displayed significant clustering based on PA level (Fig. 2a). Connectivity between 73 pairs of brain regions were associated with PA. The brain networks involved included the salience (SAL), central autonomic (CAN), central executive (CEN), emotional regulation (ERN), sensorimotor (SMN), default mode (DMN), and occipital (OCC) networks. The specific brain regions are summarized in Table 2.

Brain Connectivity Differences Based on Level of PA. (a) Clustering plot by SPLS-DA discriminating brain functional connectivity by PA groups. (b) Connectogram demonstrating q-value significant (< 0.05) brain connections derived from FDR correction between high vs. moderate PA individuals. Red lines denote increased connectivity in the high group versus green lines represent increased connectivity in the moderate group. (c) Connectogram demonstrating q-value significant (< 0.05) brain connections derived from FDR correction between moderate vs. low PA individuals. Green lines denote increased connectivity in the moderate group versus blue lines represent increased connectivity in the low group. (d) The q-value significant brain regions when comparing high versus moderate PA are displayed. (e) The q-value significant brain regions when comparing moderate versus low PA are displayed.
Compared to both moderate and low PA individuals, those with high PA have increased functional connectivity in 56 pairs of brain connections as summarized in Table 2, involving the DMN, CEN, SMN, OCC, CAN, ERN, and SAL networks (Fig. 2b,d). In contrast to high PA participants, those with moderate and low PA had significantly increased functional connectivity in 16 pairs of brain regions, including brain regions involving the DMN, CEN, OCC, ERN, and CAN networks.
When comparing moderate versus low PA, there were 55 pairs of brain connections that were increased in connectivity in the low PA group, involving the networks DMN, ERN, OCC, CEN, SAL, SMN, and CAN (Fig. 2c,e). Of these brain connections, 51 of these were the same regions that were increased in the high PA group when compared to the moderate and low groups, meaning that these regions were highest in functional connectivity in the high PA group, followed by the low PA individuals, and had the least functional connectivity in the moderate PA group (Table 2).
PA and gut microbiome composition
When comparing the three PA groups after adjusting for covariates such as age, sex, BMI and diet, significant differences in beta diversity were seen. The low PA group had a significantly different beta diversity compared to the high and moderate group, which had similar beta diversity signatures (Fig. 3A). No differences were seen with alpha-diversity indices (Fig. 3B,C). Significant differences in relative abundance were also seen when comparing both the high versus low and moderate versus low PA groups with the MaAslin2 analysis, after adjusting for covariates. When comparing high and low PA participants as seen in Fig. 3D, three genera (Fournierella, Acidaminococcus, and Prevotella) were higher in abundance and two genera (Lachnospira, Riminococcus gnavus) were lower in abundance in the high PA group. Fournierella demonstrated the greatest positive fold change when comparing high versus low PA. In the moderate versus low comparison, one genus (Prevotella) showed a greater relative abundance and seven genera (Blautia, Faecalibacterium, Bacteroides, Fusicantenibacter, Lachnospiraceae, Lachnospira, and CAG-56) had a lower relative abundance in the moderate compared to low PA (Fig. 3E). Prevotella showed the greatest positive fold change in the moderate versus low comparison and is increased in relative abundance in a dose-dependent fashion, as it also demonstrated a positive fold change in the high PA group compared to low (Fig. 3D,E).

Microbial Taxa Associated with Physical Activity. (A) Principal coordinate analysis plot of the microbiome showing beta-diversity by PA level encircled by 95% confidence interval ellipses, adjusting for sex, age, BMI, and diet. (B, C) Box plot of microbial alpha-diversity by Shannon index and Chao index respectively across PA groups. (D) MaAslin2 analysis comparing high vs. low PA showing three genera elevated in high PA participants and two genera elevated in low PA participants. (E) MaAslin2 analysis showing one genus increased and seven decreased with moderate when compared to low PA individuals. (F) Boxplot depicting the differences in Prevotella to Bacteroides ratio across high, moderate, and low PA groups.
When comparing Prevotella to Bacteroides ratio, there was overall significant differences seen across all PA groups (p = 0.03). Specifically, there was a significant difference seen between the high vs. low PA groups (p = 0.05) and the moderate vs. low (p = 0.02), but not in the high vs. low (p = 0.48) comparison. Individuals in the moderate PA group had the highest Prevotella to Bacteroides ratio and those in the low PA group had the lowest (Fig. 3F).
Microbial function was assessed by bacterial transcript abundances, which were annotated by KEGG orthology (KO)), and differential abundance testing identified 12 bacterial transcripts that were increased in relative abundance in the low PA group when compared with both the high and low PA groups, which is summarized in Supplemental Fig. 1.
Fecal metabolites associated with PA
After adjusting for confounding variables such as age, sex, BMI, and diet, 32 metabolites were associated with PA, with 13 categorized as amino acids, seven as lipids, four as nucleotides, three as carbohydrates, two as peptides, two as cofactors, and one as belonging to the energy super pathway (Table 3).
The top three metabolites that were most associated with PA were cytosine, glycosyl ceramide (D18:2/24:1, D18:1/24:2), and histidine. Additionally, lignoceroylcarnitine (C24) levels were highest in the moderate PA group and also elevated in the high PA group when compared to the low PA group. Glycosyl-N-(2-hydroxynervonoyl)sphingosine (D18:1/24:1(2Oh)) and 1-methylnicotinamide showed a dose dependent negative trend with more PA, with highest levels in the low PA group and lowest in those with high PA. On the other hand, ribulose, phenylalanine, aspartate, thymine, hypoxanthine, and glycylisoleucine levels showed a positive trend with PA, with highest levels in the high PA group. A summary of the trends for each metabolite that were associated with PA is shown in Fig. 4.

Metabolites Associated with PA. Boxplots depicting the fecal metabolites significantly associated with PA across high, moderate, and low PA groups.
Significant associations were also identified between the significant psychosocial variables (coping, resilience score, food addiction measures, education), the metabolites, and pairs of connected brain regions across all physical activity group comparisons, which is summarized in Table 4.
Discussion
In this study, we demonstrated that there are significant alterations associated with PA seen in the functional connectivity of the brain, beta diversity and relative abundance of the gut microbiome, and metabolites produced. These BGM system alterations are associated with improved psychosocial measures in an overweight and obese population. Given that individuals with high BMI face additional weight-related stressors compared to normal-weight individuals, these findings explore the possible utility of PA in preventing and treating mental illnesses in the high BMI population and how PA possibly promotes health beyond solely metabolic regulation3,42.
In this study, an association was identified between higher PA and greater resilience, which is a known protective factor against the development of psychiatric disorders such as depression and post-traumatic stress disorder (PTSD)43. This finding is consistent with previous studies showing higher resilience scores in participants with more physical exercise in various populations44,45,46. Within many of the YFAS food addiction measures, low PA was associated with the highest food addiction scores while high PA participants had the lowest scores, and these findings were associated with altered connectivity within brain regions of the DMN. Specifically, the moderate PA participants when compared to those with high PA had increased connectivity between the angular gyrus and middle temporal gyrus regions, which a previous study demonstrated to be increased in activation when participants passively viewed visual food cues versus while they actively inhibited the urge to eat47. Previous studies on the chronic effects of exercise on appetite parameters have been largely conflicting, with some studies reporting an increase in subjective appetite in the fasted state after aerobic exercise, whereas others have reported a reduction or no change48,49,50,51. Our findings suggest that the subjective appetite responses to PA may be intensity-dependent, with a greater amount of PA associated with reduced appetite.
Several of the significant microbiome genera found to be associated with PA have previously been studied in the context of psychiatric illnesses. Specifically, individuals with lower PA were seen to have increased relative abundance of Blautia and Bacteroides, which have both been shown to be increased in patients with major depressive disorder (MDD) and bipolar disorder52,53. Prevotella has also been shown to be decreased in patients with MDD, and we saw that overall higher PA was associated with a positive fold change with Prevotella in both the high versus low and moderate versus low comparisons52. When looking at past studies involving patients with general anxiety disorder, higher levels of Bacteroides and lower levels of Prevotella correlated with severity of anxiety54,55. Therefore, the significantly increased Prevotella to Bacteroides ratio that we observed in both the moderate and high PA groups in comparison to the low PA group may suggest that increased physical activity is associated with microbiome signatures protective against depression and anxiety. In addition, we found in our study that low PA was associated with Ruminococcus gnavus, which can degrade mucins and lead to gut permeability56. There is an extensive body of data that has shown that depression is associated with a low-grade intestinal inflammation, which may allow invasive bacteria to translocation into the systemic circulation57,58. This can then trigger an increase in plasma immunoglobulins targeting these bacteria and could explain why clinical depression is accompanied by increases in IgA and/or IgM59. Overall, these findings suggest that with more PA, individuals with higher BMI can encourage a microbiome signature that is protective against developing certain psychiatric illnesses such as depression and anxiety.
In addition to Prevotella’s association with psychiatric disorders, it is known to predict increased weight loss in overweight individuals and is linked with dietary fiber induced improvements in glucose metabolism through increasing fasting plasma insulin60,61. As consistent with previous findings, higher Prevotella was associated with more PA when comparing high versus low PA individuals, further demonstrating the crucial role of PA in metabolic and weight regulation through alterations in the microbiome62. We also observed that Fournierella had the greatest increase in relative abundance in the high PA versus the low PA group. In a population of participants with abdominal obesity, Fournierella has been found to be positively associated with a green-Mediterranean diet as well as reduced intrahepatic fat overtime63. The increased abundance of these genera associated with lean-phenotypes and reduced intrahepatic fat further illustrate a widely accepted finding that more PA promotes a metabolically healthy microbiome that may prevent further weight gain in individuals who already have higher BMI.
Previous studies have also studied some of the metabolites that we found associated with PA in the context of cognitive health. In our study, we observed a negative trend between the metabolite glycosyl-N-(2-hydroxynervonoyl)-sphingosine (d18: 1/24: 1(2OH)) and increased PA. In a Puerto Rican study evaluating metabolites associated with cognitive function in a non-diabetic population, glycosyl-N-(2-hydroxynervonoyl)-sphingosine (d18: 1/24: 1(2OH)) was found to be related to poor cognition, with participants that scored higher on cognitive function having lower levels of glycosyl-N-(2-hydroxynervonoyl)-sphingosine (d18: 1/24: 1(2OH))64. Additionally, we found that histidine levels were higher in the high and moderate PA groups when compared to the low PA group. Studies have shown that histidine intake improves cognitive function, potentially via the metabolism of histidine to histamine, and the histamine receptors (H1 and H3) in the brain are involved in functions related to anxiety, stress, appetite, and sleep65. This suggests that increased PA potentially could contribute to promoting better cognitive function and mental health via influence on the metabolites.
PA -associated differences were also seen in brain functional connectivity, most notably when comparing the high versus moderate PA groups. The superior frontal gyrus (SupFG) and middle frontal gyrus (MFG), which are both frontal lobe regions implicated in general inhibitory control but also appetite control, were increased in connectivity in the high PA group compared to moderate PA, which is consistent with the clinical findings of lowest food cravings with high PA and highest food cravings with moderate PA66. A possible explanation is that the MFG has also been proposed to act as a circuit-breaker between the ventral and dorsal attention networks, and thus allows for top-down reorientation of attention from endogenous stimuli such as hunger cues to exogenous stimuli in the environment67. Similar to our findings, other studies have demonstrated that in comparison to obese individuals, previously obese individuals who successfully maintained weight loss as well as lean individuals have greater activation in the SupFG in response to food cues and during tasks involving response inhibition66,68. SupFG has also been negatively correlated to self-reported impulsivity, with ADHD individuals showing hypoactivity in SupFG and MFG69. The finding of increased functional connectivity in the brain between these two regions was also correlated to the increased histidine we observed with PA, which supports the hypothesis that the effects of PA on appetite may be through connections within the gut-brain axis.
In addition, more emotional regulation network regions were increased in connectivity in moderate PA participants, and were linked to central autonomic versus central executive as seen in the high PA group, suggesting more cognitive control over emotional food cravings with more PA. There were also overall more CEN regions increased in connectivity with high PA linked to the somatosensory and default mode networks, in comparison to the increased connectivity between CEN and occipital regions seen in moderate PA individuals. This suggests more cognitive modulation and evaluation of sensory stimuli with high PA, that may contribute to more restraint and less impulsivity in uncontrolled eating.
Our study had several strengths, including the integration of a comprehensive dataset including brain, gut microbiome, fecal metabolite, and psychosocial variables to determine associations with physical activity level. We also utilized consistent sample processing and OTU clustering and considered major covariates in our analyses. However, the directionality and causality between physical activity and alterations in the BGM system cannot be parsed through this study, but cross-sectional studies such as the one we presented here allow for further understanding the role of physical activity in preventing mental illnesses. Future studies including larger and longitudinal and more evenly distributed sample sizes within each physical activity level group are warranted and would allow for more statistical power in the analyses. Including additional serum metabolome markers can also provide further clarity on the metabolites studied. In addition, despite previous studies showing the validity of self-reported physical activity data, the additional use of objective data obtained through accelerometers would enhance the accuracy of the data, which could be explored in future projects70.
With the COVID-19 pandemic creating short and long-term mental health consequences in as much as 30% of the general population and individuals already suffering from a psychiatric disease, it is even more crucial to identify evidence-based methods to promote psychological resilience amidst this ongoing global health crisis71. We have identified novel targets within the BGM system that may be explored for the prevention and treatment of various psychiatric conditions, which individuals with high BMI are already at higher risk for Ref.72. This study will inform the design of future longitudinal studies that will elucidate the directionality of these associations.
Data availability
The datasets generated during and/or analyzed during the current study are available from the corresponding author on reasonable request. The raw microbiome sequences can be accessed NIH NCBI BioProject (BioProject ID: PRJNA946906).
References
Organization, W. H. Obesity and overweight. (2020).
Weiss, F. et al. Psychiatric aspects of obesity: A narrative review of pathophysiology and psychopathology. J. Clin. Med. 9, 2344 (2020).
Sutin, A. R. & Terracciano, A. Perceived weight discrimination and high-risk health-related behaviors. Obesity (Silver Spring) 25, 1183–1186. https://doi.org/10.1002/oby.21845 (2017).
Durso, L. E., Latner, J. D. & Hayashi, K. Perceived discrimination is associated with binge eating in a community sample of non-overweight, overweight, and obese adults. Obes. Facts 5, 869–880. https://doi.org/10.1159/000345931 (2012).
Schvey, N. A., Puhl, R. M. & Brownell, K. D. The impact of weight stigma on caloric consumption. Obesity (Silver Spring) 19, 1957–1962. https://doi.org/10.1038/oby.2011.204 (2011).
Borinsky, S., Gaughan, J. P. & Feldman-Winter, L. Perceived overweight/obesity, low resilience, and body size dissatisfaction among adolescents. Obes. Res. Clin. Pract. 13, 448–452. https://doi.org/10.1016/j.orcp.2019.08.002 (2019).
Lutha, S. S. & Cicchetti, D. The construct of resilience: Implications for interventions and social policies. Dev. Psychopathol. 12, 857–885. https://doi.org/10.1017/s0954579400004156 (2000).
Guazzelli Williamson, V. et al. Psychological resilience, experimentally manipulated social status, and dietary intake among adolescents. Nutrients 13, 806. https://doi.org/10.3390/nu13030806 (2021).
Thurston, I. B., Hardin, R., Kamody, R. C., Herbozo, S. & Kaufman, C. The moderating role of resilience on the relationship between perceived stress and binge eating symptoms among young adult women. Eat. Behav. 29, 114–119. https://doi.org/10.1016/j.eatbeh.2018.03.009 (2018).
Silverman, M. N. & Deuster, P. A. Biological mechanisms underlying the role of physical fitness in health and resilience. Interface Focus 4, 20140040. https://doi.org/10.1098/rsfs.2014.0040 (2014).
Huang, C. J., Webb, H. E., Zourdos, M. C. & Acevedo, E. O. Cardiovascular reactivity, stress, and physical activity. Front. Physiol. 4, 314. https://doi.org/10.3389/fphys.2013.00314 (2013).
Gleeson, M. et al. The anti-inflammatory effects of exercise: Mechanisms and implications for the prevention and treatment of disease. Nat. Rev. Immunol. 11, 607–615. https://doi.org/10.1038/nri3041 (2011).
Gupta, A., Osadchiy, V. & Mayer, E. A. Brain-gut-microbiome interactions in obesity and food addiction. Nat. Rev. Gastroenterol. Hepatol. 17, 655–672. https://doi.org/10.1038/s41575-020-0341-5 (2020).
Agustí, A. et al. Interplay between the gut-brain axis, obesity and cognitive function. Front. Neurosci. 12, 155. https://doi.org/10.3389/fnins.2018.00155 (2018).
Dong, T. S. et al. Obesity is associated with a distinct brain-gut microbiome signature that connects Prevotella and Bacteroides to the brain’s reward center. Gut Microbes 14, 2051999. https://doi.org/10.1080/19490976.2022.2051999 (2022).
Berthoud, H. R., Münzberg, H. & Morrison, C. D. Blaming the brain for obesity: Integration of hedonic and homeostatic mechanisms. Gastroenterology 152, 1728–1738. https://doi.org/10.1053/j.gastro.2016.12.050 (2017).
Ravichandran, S. et al. Alterations in reward network functional connectivity are associated with increased food addiction in obese individuals. Sci. Rep. 11, 3386. https://doi.org/10.1038/s41598-021-83116-0 (2021).
Craig, C. L. et al. International physical activity questionnaire: 12-country reliability and validity. Med. Sci. Sports Exerc. 35, 1381–1395. https://doi.org/10.1249/01.Mss.0000078924.61453.Fb (2003).
Questionnaire, I. P. A. Guidelines for Data Processing and Analysis of the International Physical Activity Questionnaire (IPAQ) - Short and Long Forms. (2005).
Smith, B. W. et al. The brief resilience scale: assessing the ability to bounce back. Int. J. Behav. Med. 15, 194–200. https://doi.org/10.1080/10705500802222972 (2008).
Zigmond, A. S. & Snaith, R. P. The hospital anxiety and depression scale. Acta Psychiatr. Scand. 67, 361–370. https://doi.org/10.1111/j.1600-0447.1983.tb09716.x (1983).
Carver, C. S. You want to measure coping but your protocol’s too long: Consider the brief COPE. Int. J. Behav. Med. 4, 92–100. https://doi.org/10.1207/s15327558ijbm0401_6 (1997).
Gearhardt, A. N., Corbin, W. R. & Brownell, K. D. Preliminary validation of the yale food addiction scale. Appetite 52, 430–436. https://doi.org/10.1016/j.appet.2008.12.003 (2009).
Gordon, E. L., Ariel-Donges, A. H., Bauman, V. & Merlo, L. J. What is the evidence for “food addiction?” A systematic review. Nutrients 10, 477 (2018).
Coley, E. J. L. et al. Early life adversity predicts brain-gut alterations associated with increased stress and mood. Neurobiol. Stress 15, 100348–100348. https://doi.org/10.1016/j.ynstr.2021.100348 (2021).
Tong, M., Jacobs, J. P., McHardy, I. H. & Braun, J. Sampling of intestinal microbiota and targeted amplification of bacterial 16S rRNA genes for microbial ecologic analysis. Curr. Protoc. Immunol. 107, 41–11. https://doi.org/10.1002/0471142735.im0741s107 (2014).
Callahan, B. J. et al. DADA2: High-resolution sample inference from Illumina amplicon data. Nat. Methods 13, 581–583. https://doi.org/10.1038/nmeth.3869 (2016).
Martino, C. et al. A novel sparse compositional technique reveals microbial perturbations. mSystems https://doi.org/10.1128/mSystems.00016-19 (2019).
Lozupone, C. & Knight, R. UniFrac: A new phylogenetic method for comparing microbial communities. Appl. Environ. Microbiol. 71, 8228–8235. https://doi.org/10.1128/AEM.71.12.8228-8235.2005 (2005).
Anderson, M. J. A new method for non-parametric multivariate analysis of variance. Austral. Ecol. 26, 32–46. https://doi.org/10.1046/j.1442-9993.2001.01070.x (2001).
Ciric, R. et al. Benchmarking of participant-level confound regression strategies for the control of motion artifact in studies of functional connectivity. Neuroimage 154, 174–187. https://doi.org/10.1016/j.neuroimage.2017.03.020 (2017).
Mallick, H. et al. Multivariable association discovery in population-scale meta-omics studies. PLoS Comput. Biol. 17, e1009442. https://doi.org/10.1371/journal.pcbi.1009442 (2021).
Dong, T. S. et al. A distinct brain-gut-microbiome profile exists for females with obesity and food addiction. Obesity (Silver Spring) 28, 1477–1486. https://doi.org/10.1002/oby.22870 (2020).
Gupta, A. et al. Early adverse life events are associated with altered brain network architecture in a sex- dependent manner. Neurobiol. Stress 7, 16–26. https://doi.org/10.1016/j.ynstr.2017.02.003 (2017).
Power, J. D. et al. Methods to detect, characterize, and remove motion artifact in resting state fMRI. Neuroimage 84, 320–341. https://doi.org/10.1016/j.neuroimage.2013.08.048 (2014).
Irimia, A., Chambers, M. C., Torgerson, C. M. & Van Horn, J. D. Circular representation of human cortical networks for subject and population-level connectomic visualization. Neuroimage 60, 1340–1351. https://doi.org/10.1016/j.neuroimage.2012.01.107 (2012).
Schaefer, A. et al. Local-global parcellation of the human cerebral cortex from intrinsic functional connectivity MRI. Cereb. Cortex 28, 3095–3114. https://doi.org/10.1093/cercor/bhx179 (2018).
Osadchiy, V. et al. Analysis of brain networks and fecal metabolites reveals brain-gut alterations in premenopausal females with irritable bowel syndrome. Transl. Psychiatry 10, 367. https://doi.org/10.1038/s41398-020-01071-2 (2020).
Coley, E. J. L. et al. Early life adversity predicts brain-gut alterations associated with increased stress and mood. Neurobiol. Stress 15, 100348. https://doi.org/10.1016/j.ynstr.2021.100348 (2021).
Division of Nutrition, P. A., and Obesity, National Center for Chronic Disease. Healthy Weight, Nutrition, and Physical Activity. Centers for Disease Control and Prevention (2021).
Min, Y. et al. Sex-specific association between gut microbiome and fat distribution. Nat. Commun. 10, 2408. https://doi.org/10.1038/s41467-019-10440-5 (2019).
Reilly, J. J. & Kelly, J. Long-term impact of overweight and obesity in childhood and adolescence on morbidity and premature mortality in adulthood: Systematic review. Int. J. Obes. (Lond.) 35, 891–898. https://doi.org/10.1038/ijo.2010.222 (2011).
Pietrzak, R. H. et al. Psychosocial buffers of traumatic stress, depressive symptoms, and psychosocial difficulties in veterans of operations enduring freedom and Iraqi freedom: The role of resilience, unit support, and postdeployment social support. J. Affect. Disord. 120, 188–192. https://doi.org/10.1016/j.jad.2009.04.015 (2010).
Yoshikawa, E., Nishi, D. & Matsuoka, Y. J. Association between regular physical exercise and depressive symptoms mediated through social support and resilience in Japanese company workers: A cross-sectional study. BMC Public Health 16, 553. https://doi.org/10.1186/s12889-016-3251-2 (2016).
Fields, A. J., Hoyt, R. E., Linnville, S. E. & Moore, J. L. Physical activity, sleep, and C-reactive protein as markers of positive health in resilient older men. J. Health Psychol. 21, 1928–1938. https://doi.org/10.1177/1359105314568578 (2016).
Matzka, M. et al. Relationship between resilience, psychological distress and physical activity in cancer patients: A cross-sectional observation study. PLOS ONE 11, e0154496. https://doi.org/10.1371/journal.pone.0154496 (2016).
Tuulari, J. J. et al. Neural circuits for cognitive appetite control in healthy and obese individuals: An fMRI study. PLoS One 10, e0116640. https://doi.org/10.1371/journal.pone.0116640 (2015).
King, N. A. et al. Dual-process action of exercise on appetite control: Increase in orexigenic drive but improvement in meal-induced satiety. Am. J. Clin. Nutr. 90, 921–927. https://doi.org/10.3945/ajcn.2009.27706 (2009).
Martins, C., Kulseng, B., King, N. A., Holst, J. J. & Blundell, J. E. The effects of exercise-induced weight loss on appetite-related peptides and motivation to eat. J. Clin. Endocrinol. Metab. 95, 1609–1616. https://doi.org/10.1210/jc.2009-2082 (2010).
Guelfi, K. J., Donges, C. E. & Duffield, R. Beneficial effects of 12 weeks of aerobic compared with resistance exercise training on perceived appetite in previously sedentary overweight and obese men. Metabolism 62, 235–243. https://doi.org/10.1016/j.metabol.2012.08.002 (2013).
Martins, C., Kulseng, B., Rehfeld, J. F., King, N. A. & Blundell, J. E. Effect of chronic exercise on appetite control in overweight and obese individuals. Med. Sci. Sports Exerc. 45, 805–812. https://doi.org/10.1249/MSS.0b013e31827d1618 (2013).
Chung, Y. E. et al. Exploration of microbiota targets for major depressive disorder and mood related traits. J. Psychiatr. Res. 111, 74–82. https://doi.org/10.1016/j.jpsychires.2019.01.016 (2019).
Rong, H. et al. Similarly in depression, nuances of gut microbiota: Evidences from a shotgun metagenomics sequencing study on major depressive disorder versus bipolar disorder with current major depressive episode patients. J. Psychiatr. Res. 113, 90–99. https://doi.org/10.1016/j.jpsychires.2019.03.017 (2019).
Coupland, N. J. et al. Decreased prefrontal Myo-inositol in major depressive disorder. Biol. Psychiatry 57, 1526–1534. https://doi.org/10.1016/j.biopsych.2005.02.027 (2005).
Chen, Y. H. et al. Association between fecal microbiota and generalized anxiety disorder: Severity and early treatment response. J. Affect. Disord. 259, 56–66. https://doi.org/10.1016/j.jad.2019.08.014 (2019).
Jiang, H. Y. et al. Altered gut microbiota profile in patients with generalized anxiety disorder. J. Psychiatr. Res. 104, 130–136. https://doi.org/10.1016/j.jpsychires.2018.07.007 (2018).
O’Malley, D., Julio-Pieper, M., Gibney, S. M., Dinan, T. G. & Cryan, J. F. Distinct alterations in colonic morphology and physiology in two rat models of enhanced stress-induced anxiety and depression-like behaviour. Stress 13, 114–122. https://doi.org/10.3109/10253890903067418 (2010).
Jiang, H. et al. Altered fecal microbiota composition in patients with major depressive disorder. Brain Behav. Immun. 48, 186–194. https://doi.org/10.1016/j.bbi.2015.03.016 (2015).
Maes, M., Kubera, M., Leunis, J.-C. & Berk, M. Increased IgA and IgM responses against gut commensals in chronic depression: Further evidence for increased bacterial translocation or leaky gut. J. Affect. Disord. 141, 55–62 (2012).
Christensen, L. et al. Prevotella abundance predicts weight loss success in healthy, overweight adults consuming a whole-grain diet ad libitum: A post hoc analysis of a 6-Wk randomized controlled trial. J. Nutr. 149, 2174–2181. https://doi.org/10.1093/jn/nxz198 (2019).
Stanislawski, M. A., Dabelea, D., Lange, L. A., Wagner, B. D. & Lozupone, C. A. Gut microbiota phenotypes of obesity. npj Biofilms Microbiomes 5, 18. https://doi.org/10.1038/s41522-019-0091-8 (2019).
Petersen, L. M. et al. Community characteristics of the gut microbiomes of competitive cyclists. Microbiome 5, 98. https://doi.org/10.1186/s40168-017-0320-4 (2017).
Yaskolka Meir, A. et al. Effect of green-Mediterranean diet on intrahepatic fat: The DIRECT PLUS randomised controlled trial. Gut 70, 2085–2095. https://doi.org/10.1136/gutjnl-2020-323106 (2021).
Palacios, N. et al. Circulating plasma metabolites and cognitive function in a puerto rican cohort. J. Alzheimers Dis. 76, 1267–1280. https://doi.org/10.3233/jad-200040 (2020).
Thalacker-Mercer, A. E. & Gheller, M. E. Benefits and adverse effects of histidine supplementation. J. Nutr. 150, 2588S-2592S. https://doi.org/10.1093/jn/nxaa229 (2020).
Batterink, L., Yokum, S. & Stice, E. Body mass correlates inversely with inhibitory control in response to food among adolescent girls: An fMRI study. Neuroimage 52, 1696–1703. https://doi.org/10.1016/j.neuroimage.2010.05.059 (2010).
Corbetta, M., Patel, G. & Shulman, G. L. The reorienting system of the human brain: From environment to theory of mind. Neuron 58, 306–324. https://doi.org/10.1016/j.neuron.2008.04.017 (2008).
McCaffery, J. M. et al. Differential functional magnetic resonance imaging response to food pictures in successful weight-loss maintainers relative to normal-weight and obese controls. Am. J. Clin. Nutr. 90, 928–934. https://doi.org/10.3945/ajcn.2009.27924 (2009).
Booth, J. R. et al. Larger deficits in brain networks for response inhibition than for visual selective attention in attention deficit hyperactivity disorder (ADHD). J. Child Psychol. Psychiatry 46, 94–111. https://doi.org/10.1111/j.1469-7610.2004.00337.x (2005).
Cleland, C., Ferguson, S., Ellis, G. & Hunter, R. F. Validity of the International Physical Activity Questionnaire (IPAQ) for assessing moderate-to-vigorous physical activity and sedentary behaviour of older adults in the United Kingdom. BMC Med. Res. Methodol. 18, 176. https://doi.org/10.1186/s12874-018-0642-3 (2018).
Verdolini, N. et al. Resilience and mental health during the COVID-19 pandemic. J. Affect. Disord. 283, 156–164. https://doi.org/10.1016/j.jad.2021.01.055 (2021).
Simon, G. E. et al. Association between obesity and psychiatric disorders in the US adult population. Arch. Gen. Psychiatry 63, 824–830. https://doi.org/10.1001/archpsyc.63.7.824 (2006).
Funding
This research was supported by grants from the National Institutes of Health including R01 MD015904 (AG), K23 DK106528 (AG), R03 DK121025 (AG), T32 DK07180 (TD), ULTR001881/DK041301 (UCLA CURE/CTSI Pilot and Feasibility Study (AG); and pilot funds provided for brain scanning by the Ahmanson-Lovelace Brain Mapping Center. These funders played no role in study design, or the collection, analysis, and interpretation of the data. We acknowledge the analytical and data curation efforts provided by the neuroimaging core, the database and clinical core of the Goodman-Luskin Microbiome Center at UCLA, and Jennifer Yang. Provisional Patent filed: METHODS AND COMPOSITIONS FOR IMPROVED PSYCHOLOGICAL AND RESILIENCE MEASURES Inventor(s): Arpana Gupta et al.FH Reference No.: UCH-33625 (32246.33625) International Patent Application Serial No. PCT/US2023/067223.
Author information
Authors and Affiliations
Contributions
Conceptualization: M.G., A.G.; Methodology: T.S.D., A.G.; Formal analysis: V.S., Y.G., T.S.D., M.G., A.G.; Resources: A.G.; Data curation: A.V.; Original draft preparation: M.G.; Review and editing: M.G., T.S.D., R.L.B., A.G; visualization: V.S., Y.G., T.S.D, R.B.; supervision: M.G., A.G.; funding acquisition: A.G. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information

Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Guan, M., Dong, T.S., Subramanyam, V. et al. Improved psychosocial measures associated with physical activity may be explained by alterations in brain-gut microbiome signatures. Sci Rep 13, 10332 (2023). https://doi.org/10.1038/s41598-023-37009-z
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-023-37009-z
This article is cited by
-
Stress-resilience impacts psychological wellbeing as evidenced by brain–gut microbiome interactions
Nature Mental Health (2024)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.

{kind=link}