
Abstract
Understanding large carnivore demography on human-dominated lands is a priority to inform conservation strategies, yet few studies examine long-term trends. Jaguars (Panthera onca) are one such species whose population trends and survival rates remain unknown across working lands. We integrated nine years of camera trap data and tourist photos to estimate jaguar density, survival, abundance, and probability of tourist sightings on a working ranch and tourism destination in Colombia. We found that abundance increased from five individuals in 2014 to 28 in 2022, and density increased from 1.88 ± 0.87 per 100 km2 in 2014 to 3.80 ± 1.08 jaguars per 100 km2 in 2022. The probability of a tourist viewing a jaguar increased from 0% in 2014 to 40% in 2020 before the Covid-19 pandemic. Our results are the first robust estimates of jaguar survival and abundance on working lands. Our findings highlight the importance of productive lands for jaguar conservation and suggest that a tourism destination and working ranch can host an abundant population of jaguars when accompanied by conservation agreements and conflict interventions. Our analytical model that combines conventional data collection with tourist sightings can be applied to other species that are observed during tourism activities.
Entender los patrones demográficos de los grandes carnívoros al interior de paisajes antrópicos es fundamental para el diseño de estrategias de conservación efectivas. En el Neotrópico, el jaguar (Panthera onca) es una de estas especies cuyas tendencias poblacionales y tasas de supervivencia en paisajes productivos son desconocidas. Para entender mejor estas dinámicas, integramos nueve años de fototrampeo junto a fotos de turistas para estimar la densidad, supervivencia, abundancia y probabilidad de avistamiento de esta especie en una finca ganadera y destino turístico en Colombia. Entre 2014 y 2022 encontramos que la abundancia incrementó de cinco a 28 individuos y la densidad de 1.88 ± 0.87 jaguares/ 100 km2 a 3.80 ± 1.08 jaguares/ 100 km2. La probabilidad de avistamiento por turistas aumentó de 0% en 2014 a 40% en 2020 antes de la pandemia del Covid-19. Nuestros resultados presentan las primeras estimaciones robustas de abundancia y supervivencia de este felino en paisajes antrópicos dónde el manejo de sistemas productivos combinados con turismo e intervenciones para la mitigación del conflicto puede albergar poblaciones abundantes de jaguares, demostrando su importancia para la conservación de esta especie. Nuestro modelo, al combinar datos convencionales con avistamientos, podría ser aplicado a otras especies observadas durante actividades turísticas.
Similar content being viewed by others

Introduction
Large carnivores can play crucial regulatory roles in the ecosystems they inhabit, yet populations of many species continue to decline despite conservation efforts1. A key strategy for conserving large carnivores is reducing pressures in human-dominated landscapes, given that most suitable habitat occurs outside of protected areas2,3. Working lands can provide vital, permeable habitat for carnivores4. However, interactions between large carnivores and humans on private lands may be negative, impacting both carnivores and people5. Conservation scientists and practitioners are interested in developing strategies to promote coexistence with large carnivores6, where carnivore populations are sustained and human livelihoods thrive with minimal conflict7.
Whereas local support for carnivore presence on working lands tends to be low due to livestock depredation and safety concerns, value for carnivores at the global scale, especially felids, tends to be high8. Successful coexistence with large carnivores requires converting global existence value into tangible financial benefits for local communities9. Conservationists have explored various funding mechanisms to create value from the presence of large carnivores for rural communities9,10. One such opportunity to promote coexistence locally and generate income for those living with carnivores is wildlife tourism, where tourists pay to observe fauna in their natural habitats.
Because attitudes towards large carnivores are a strong predictor of human behavior towards them11,12, proponents hope that tourism income can sway local communities towards positive perceptions of carnivores while benefiting wildlife populations13. Income from wildlife tourism can offset the costs of living with large carnivores, such as livestock depredation14. Furthermore, wildlife tourism can contribute to conservation through funding anti-poaching patrols, livestock compensation programs, and species recovery and restoration, as well as providing opportunities for scientific research and wildlife monitoring13,15. However, wildlife tourism initiatives can have both positive and negative outcomes for wildlife populations16,17,18. The success of coexistence strategies like tourism requires understanding and quantifying such impacts19, and these strategies promote coexistence more effectively when accompanied by efforts to reduce livestock losses and conservation agreements20. Moreover, the presence of tourists with high-quality photography capabilities can have important, albeit underutilized, contributions to wildlife monitoring21.
Developing effective coexistence strategies for jaguars (Panthera onca) is urgently needed. Jaguars are among the most emblematic large carnivores22. Over half (55%) of remaining populations exist outside of protected areas23, hence research and conservation strategies on working lands are of critical importance24. Jaguars have lost nearly half their range in the last 50 years25. Major threats to their persistence are habitat loss and fragmentation26, direct killings to prevent or retaliate for livestock losses, and prey depletion25.
Understanding jaguar survival rates is critical to their conservation, but acquiring accurate population metrics can be difficult and costly27. Jaguars are naturally elusive, and densities are low23. Longitudinal studies of jaguars are therefore challenging because of the financial and logistical difficulty of long-term monitoring. Across the jaguar’s extensive range, only four long-term jaguar population studies have been published27,28,29,30. In addition, no published long-term study has addressed survival and abundance on working lands, where densities tend to be lower23 and survival is likely more challenging.
Hato La Aurora, a working cattle ranch and private reserve, is the only place in Colombia where jaguar tourism is practiced in the species’ natural habitat. Hato La Aurora was formally established as a reserve in 2008, and hunting was prohibited on the ranch since 1979. Because of the killing of jaguars for their pelts (known locally as tigrilladas) in the mid-twentieth century31, populations were decimated, and no jaguars were observed in Hato La Aurora until 2002. In 2013, Hato La Aurora began working with Panthera Colombia, a felid conservation organization, to monitor the jaguar population and reduce human-jaguar conflict in and around the ranch. Panthera Colombia implemented conservation agreements and electric fences around calving pastures to reduce livestock depredations from jaguars with 19 ranches neighboring Hato La Aurora, totaling 8558 hectares contiguous to Hato La Aurora and 19,326 hectares nearby (Panthera Colombia, unpublished data).
Here, we present the first longitudinal study and demographic estimates for jaguars on working lands and in Colombia. We integrated nine years of camera trap data and tourist photos of jaguars collected between 2014 and 2022 in a Barker Robust Design Model (Barker/RD)32. Our approach estimated survival rates, probability of observation by tourists, and abundance of jaguars, and we derived finite rate of change in abundance and recruitment33. We complemented the robust design analysis by comparing our results with spatially explicit density estimates in 2014 and 2022 from the study area. We hypothesized that jaguar abundance, survival, density, and observation by tourists increased over time as a result of recently implemented conservation efforts. We discuss how our findings contribute to human-jaguar coexistence strategies, and how our modeling approach can contribute to demographic studies of species photographed by tourists.
Results
We recorded 50 individual jaguars from 659 identifiable camera trap records and 79 tourist sightings between 2014 and 2022 (Fig. 1). We detected 19 females and 31 males, although five individuals (four of which were females) were removed from the analysis because they were not detected within closed-capture time periods.

Photos: Fundación Jaguar & Panthera Colombia (left), S. Rincón (right).
Example of a camera trap detection (left) and a tourist sighting (right) of the same individual.
We ran 26 models in the Barker/RD to explore our hypotheses (Table A1), three of which summed to 0.76 of model weight (Table 1). The most parsimonious model had an Akaike Information Criteria (AIC)34 weight of 0.32 and constant survival rates for males and females, probability of observation by tourists variable by time, and constant detection over the study period. Other parameters estimated by the Barker/RD model are found in Table A5.
Our second most parsimonious model, with 0.24 model weight, differed only by constant probability of observation by tourists. Our third model, with 0.21 model weight, demonstrated evidence of transience, whereby a jaguar was detected once in the study area and never again. Below we present estimates from the top model and model-averaged estimates for our parameters of interest. Model-averaged estimates for all parameters can be found in Table A5.
Survival estimates
Our survival parameter was apparent survival because dead recoveries of individuals were not possible28. Apparent survival was constant throughout the study period (Fig. 2a), though the third most parsimonious model suggested transience, where an individual was detected once and did not return. Model-averaged estimates suggest a slight difference between males and females. The average survival rate was 0.783 (SE 0.075, 95% CI: 0.603–0.896) for males and 0.798 (SE 0.068 95% CI: 0.633–0.900) for females. Survival rates given transience were lower, 0.706 (SE 0.199, 95% CI: 0.318–0.934) for males and 0.720 (SE 0.200, 95% CI: 0.322–0.941) for females.

(a) Model weighted survival probability for male and female jaguars. (b) Model weighted derived estimates of male and female jaguar abundances. (c) Derived abundance estimates and Delta method results for recruitment. (d) Resighting probability by tourists. Error bars and shaded areas indicate 95% confidence intervals.
Tourist observation estimates
The probability of observation by tourists varied over time (Fig. 2d). From 2014 to 2017, the probability of observation by tourists of both sexes was effectively zero. In 2018, the probability of observation by tourists increased to 0.348 (SE 0.157, 95% CI: 0.121–0.674) for females and 0.342 (SE 0.158, 95% CI: 0.116–0.674) for males. Observation by tourists peaked in 2020 at 0.409 (SE 0.181, 95% CI: 0.137–0.750) for females and 0.403 (SE 0.182, 95% CI: 0.133–0.748) for males. The observation probability dropped to less than half of that value in 2021, at 0.186 (SE 0.083, 95% CI: 0.072–0.400) for males and 0.190 (SE 0.084, 95% CI: 0.074–0.407) for females, before rebounding to 0.273 (SE 0.114, 95% CI: 0.108–0.537) for males and 0.279 (SE 0.115, 95% CI: 0.111–0.542) for females in 2022.
Abundance estimates
Abundance increased more than five-fold over the nine-year period in Hato La Aurora (Fig. 2b and 2c). In 2014, there were estimated 3.00 males (SE 0.017, 95% CI: 2.97–3.03) and 2.63 females (SE 0.557, 95% CI: 0.12–3.36). The maximum estimated abundance in the nine-year period was in 2022, when males reached 14.54 individuals (SE 1.392, 95% CI: 11.81–17.27) and females reached 14.37 (SE 1.307, 95% CI: 11.81–16.93). The lowest recorded abundance was in 2016, when abundance was only four—1.00 female and 3.41 males (SE 0.0105, 95% CI: 0.98–1.02; SE 0.686, 95% CI: 0.2.06–4.74). Our derived abundance estimates differ from our raw numbers of jaguars, which suggests that we did not observe all jaguars that were present in Hato La Aurora (p* < 1) most years, with the exception of 2018. However, detection was high during the study period: 0.900 for mixture 1 of jaguars whose territory was mostly in the reserve and 0.412 for mixture 2 of jaguars whose territory partially overlapped in the top model.
Recruitment and population growth from derived parameters
The Delta method for estimating derived parameters revealed an increasing population during the study; finite rate of change in abundance (λ) averaged 1.389 for males and 1.822 for females (Table A5). Notably, the female population had the highest growth between 2016 and 2017, when it rose to 6.801 because of the arrival of five new females. Male population growth was highest in the intervals of 2014–2015 (2.215) and 2016–2017 (2.000).
Recruitment of both male and female jaguars was highest from 2021–2022, when it was 6.253 for males and 6.699 for females (Fig. 2c). The lowest recruitment for males (− 2.184) occurred between 2017 and 2018. The lowest recruitment for females (− 1.714) occurred between 2015 and 2016. Life histories of jaguars between 2014 and 2022 are shown in Fig. 3.

Life histories of jaguars detected in Hato La Aurora since 2014. Bolded names are jaguars that were born in Hato La Aurora. Jaguar icon credit: Gabriela Palomo-Muñoz.
SECR density
Trapping effort for spatially explicit capture-recapture (SECR) density in 2022 totaled 1985 camera trap nights. We recorded 17 individuals (99 capture events at 19 of 32 stations): nine females (49 capture events) and eight males (50 capture events). Seven females and seven males were captured at different stations. The best model was SECR.σ where σ varies between males and females (Table 1). SECR.σ produced a density estimate of 3.803 jaguars/100 km2 (SE 1.048; 95% CI: 2.238–6.464). The g0 parameter estimate was 0.018 (SE 0.003; 95% CI: 0.013–0.026). The σ estimate was 2.321 km (SE 0.274; 95% CI 1.842–2.924) for females and 5.091 (SE 1.031; 95% CI: 3.437–7.540) for males. Despite overlapping estimates confidence intervals (CIs), the estimated density in 2022 was approximately twice that from 2014 (1.88 ± 0.87 jaguars/100 km2; 95% CI: 0.79–4.48)24. Estimates for σ were also similar for females (females 2014: 2.327, SE 0.693 95% CI 1.315–4.119) and higher for males (males 2014: 1.426, SE 0.129 95% CI 1.195–1.701) than 2014 estimates indicating that the smaller grid size in 2022 did not bias density estimates.
Discussion
Conserving large carnivores requires strategies beyond protected areas2. Working lands with adequate conservation measures can provide sufficient habitat to sustain resident jaguar populations that are comparable to those of protected areas4. Promoting coexistence can be facilitated by economic mechanisms like tourism that can ease the livelihood impacts of living with large carnivores. To inform such initiatives, there is a need for rigorous, long-term data collection to understand impacts to carnivore demography35. However, few long-term demographic studies exist for jaguars, hindering the evaluation of conservation efforts30. Where such studies exist, they tend to focus on protected areas27,28,29. Our longitudinal study of jaguars in Hato La Aurora, a working ranch and tourism destination in the Colombian Llanos, suggests that private lands with low-intensity cattle ranching and tourism can sustain an abundant jaguar population if combined with conservation actions such as hunting prohibitions, abundant prey36, conservation agreements with adjacent ranches28 and depredation reduction strategies in the form of electric fencing of calving pastures.
Our application of the Barker/RD, which integrated tourist photos into nine years of camera trapping data, provided much-needed demographic estimates for jaguars, including the first survival estimates for jaguars on working lands and in Colombia. The inclusion of tourist photos increased the precision of survival estimates32 and also allowed for quantification of jaguar sightings. Our estimates indicate that Hato La Aurora supported 28 ± 2.70 individual jaguars on the 15,000-hectare ranch in 2022, which is comparable to that of small protected areas within their range. In the federally protected Cockscomb Basin Wildlife Sanctuary in Belize, for example, estimated jaguar abundance peaked at 31 ± 4.77 individuals in 49,000 hectares27, an area more than three times the size of Hato La Aurora. Jaguars in Hato La Aurora had a high survival rate (0.78 ± 0.075), again similar to the highest estimated survival rate from Cockscomb Basin Wildlife Sanctuary in Belize (0.78 ± 0.05)27.
Our 2022 density estimate of 3.80 ± 1.08 jaguars/100 km2 in Hato La Aurora is consistent with recent density studies of jaguars from other working lands. In the Brazilian Pantanal, Devlin et al.4 estimated 4.08 ± 0.73 jaguars/100 km2 on multi-use (ranching, conservation, and tourism) landscapes. On a state-run cattle ranch in the Venezuelan Llanos with a long history of conservation, Jędrzejewski et al.37 estimated a density of 7.67 jaguars/100 km2. The ecological similarity of the Venezuelan Llanos with Hato La Aurora suggests that the Colombian Llanos could host a higher density of jaguars if threats are sufficiently reduced.
The increase in density estimates from 1.88 ± 0.87 in 2014 to 3.80 ± 1.08 jaguars/100 km2 in 2022, and the concomitant increase in jaguar abundance from five to 28 individuals, is encouraging for range-wide conservation efforts like the Jaguar Corridor Initiative, which seeks to maintain genetic connectivity between source populations throughout Central and South America38. This population increase was likely due to the high survival rate of jaguars and the 1.82 annual population growth (λ) of females, which is an important determinant of demography for long-lived species with low reproduction rates39. However, the specific causal mechanisms driving the population increase require further investigation. We speculate that the status of Hato La Aurora as a private reserve and the conservation actions (e.g., electric fences for calving pastures and conservation agreements) implemented by Panthera Colombia on smaller, nearby ranches enhanced habitat suitability for jaguars, reduced human hunting of prey, and decreased livestock depredations and therefore retaliatory killings. The distribution of land ownership in the Colombian Llanos, whereby large ranches are often surrounded by smaller parcels, may necessitate a dual approach to coexistence strategies. Actions like electric fencing to reduce depredations on larger ranches like Hato La Aurora may be cost-prohibitive because of the extension of land and size of cattle herds. Similarly, tourism on smaller ranches surrounding Hato La Aurora is challenging due to smaller plots of land and limited infrastructure, but electric fencing of pastures is more feasible. In addition, it is possible that jaguar populations are recovering throughout the region since hunting and pelt exports were outlawed following the inclusion of jaguars in the CITES Appendix I in the 1970s31. Tourism is the economic mechanism that allows the ranch owners to coexist with jaguars in Hato La Aurora, and therefore is of critical importance to jaguar persistence on the landscape.
Sustaining large carnivores on ranchlands requires minimizing livelihood impacts7. Tortato et al.14 found that in the Brazilian Pantanal income was over 50 times higher from tourism than the estimated cost of livestock depredation on cattle ranches. In Hato La Aurora, wildlife tourism is the economic vehicle that permits coexistence between livestock systems and jaguars, though the differential between tourism and livestock depredation is lower than that of the Brazilian Pantanal. Hato La Aurora loses on average 100 head of cattle, and a similar number of foals and pigs, to jaguars and pumas annually. Accurate data on income from tourism and the number of tourists visiting Hato La Aurora were unavailable, but ranch owners report that tourism income is crucial to offset the cost of living with carnivores. The allure of observing a jaguar in the wild raises the attractiveness of tourism in Hato La Aurora—further contributing to the viability of this coexistence strategy.
A challenge to large carnivore tourism, however, is reconciling their elusive nature with tourists’ desire for predictable, high-quality sightings40. Hunting prohibitions can make it easier to see species in tourist areas41, and savannahs are ideal observation sites because of visibility42. This appears to be the case in Hato La Aurora where our models showed higher probability of observation by tourists in recent years compared to 2014–2017. The probability peaked at 0.409 ± 0.182 in 2020 in the dry season months before the Covid-19 lockdown in March of 2020. The next year, in 2021, tourist sightings declined, which we attribute to lingering Covid-19 travel restrictions. We posit that, since jaguars in Hato La Aurora are not hunted or hazed, they may perceive a safer setting and be less likely to avoid human activity. Alternatively, a contributing factor to increased sightings may also be the knowledge of ranch owners and guides of daily patterns of jaguars. As tour guides came to understand where jaguars may be at peak times, they may have frequented those sites, leading to an increase in sighting probability.
Community science data collected by tourists, such as the photographs analyzed in our study, can have important and low-cost contributions to wildlife monitoring efforts21. Our application of the Barker/RD using tourist photos as auxiliary data can be applied to other monitoring programs of large, terrestrial mammals, especially to understand the probability of tourists observing them in the wild. When using tourist data, it would be useful to collect information on “sampling effort” of tourists by recording the number of people in each tour and the time spent observing wildlife and locations21. If using camera traps for robust design modeling, moving stations within and between survey periods may increase total detection probability of individual animals and reduce sex-specific and individual heterogeneity27,43.
Replicable and landscape-scale coexistence strategies are necessary for large carnivore conservation. In the case of jaguar tourism, understanding the causal mechanisms behind increased sightings is important to strengthen tourism in Hato La Aurora and beyond. Accompanying studies to assess tourism’s impact on attitudes and tolerance44,45 and local livelihoods46 are necessary for adaptive management of the industry and to optimize long-term benefits for jaguars. For jaguars in Colombia, prey depletion and the introduction of bovines into jaguar habitat are likely exacerbating conflict. Studies on jaguar diets, quantifying retaliatory killings, evaluating nonlethal strategies to reduce livestock depredation, and the human dimensions of living with jaguars are needed to understand barriers and enablers for human-jaguar coexistence at scale.
Methods
Study area
Colombia is the most diverse country by land area47, harboring 1941 species of birds48, 520 mammals49, and the third largest population of jaguars23. The department of Casanare, located in the country’s eastern Llanos (plains), is part of the transition zone between the tropical rainforest of the Amazon and the Eastern Andes. The landscape is dominated by seasonally flooded savannahs of the Orinoco basin, which form the largest wetland complex in the country. Casanare is a top destination for wildlife viewing due to open savannahs and abundant populations of mammals and birds. The area has been dominated by extensive cattle ranches since bovine introduction in the 1600s50. However, petroleum exploitation, oil palm plantations, and rice cultivation have increased in the last few decades51.
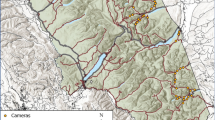
Hato La Aurora (5° 57′ 18.8″ N, 71° 29′ 0.1″ E to 6° 4′ 52.6″ N, 71° 17′ 51.4″ E) is a private reserve consisting of 15000 hectares of tropical savannahs and gallery forests in the department of Casanare (Fig. 4). The area receives 1000–3000 mm of rainfall each year, with marked dry (December–May) and wet (June–November) seasons52. The principal land use and economic activity is cattle ranching, with some introduced grasses for cattle forage. The surrounding area is comprised principally of extensively managed cattle ranches with riparian forest cover, though rice plantations are increasing in the region.

Sources: Natural Earth, U.S. Geological Survey, Panthera Colombia, Fundación Jaguar Colombia.
Location of Hato La Aurora, Colombia, and camera trap stations between 2014 and 2022.
Camera trapping
We installed a total of 296 camera-traps (models Panthera V3, V4, V5, V7, and Cuddeback 1279 and G-5048) between 2014 and 2022 at a distance of 1.5 ± 0.5 km for the study of medium and large vertebrates53 for a total of 16790 trap-nights (Table A1). Camera-trap grids used the same 24-h configuration and a quiet period of (30 s) between trigger events. No baits were used in any of the studies.
Camera trapping for density estimation (2014 and 2022 surveys) followed standardized recommendations53 and complied with the capture-recapture model assumptions: the population is closed, and all individuals have a possibility of being captured54,55. In April–May of 2014, we installed 52 double camera stations in a grid covering 152 km2 at an average distance of 1.6 ± 0.2 km (see Boron et al.24 for detailed survey information). In March–May of 2022, we placed 32 single camera stations (Table A6) in a grid covering 102 km2 (minimum convex polygon). We installed Cuddeback models 1279 and G-5048, and Panthera series 6 and 7 cameras at a height of 35–40 cm. Cameras remained active for 24 h per day. We used single stations because long-term monitoring of individuals provided photographic evidence of both sides of most jaguars, enabling individual identification when only one side was photographed. Like in 2014, the average distance between stations was 1.6 ± 0.2 km, which is consistent with recommendations for jaguar density studies53 and is appropriate when considering jaguar home ranges estimates, since it ensures all individuals can be photo-captured56. According to Colombian regulation, non-invasive camera trap studies do not require permits or approval from an Institutional Animal Care and Use Committee or equivalent.
Auxiliary tourist photographs
Trained guides accompanied groups and ensured compliance with the regulations of the reserve. Jaguar viewing occurred in open vehicles and tour guides were required to maintain a minimum distance of 100 m, avoid any noises, and not leave the vehicle. We collected tourist photos of jaguars that were observed on an opportunistic basis during the study period. Tourists and guides reported sightings and delivered photos to co-author J. Barragan for identification. Jaguars were uniquely identified by rosettes and spot patterns24. Between 2014 and 2022, we collected 79 direct observations where individuals were identifiable. These sightings occurred primarily during the dry season (December–May) due to access issues in the rainy season and prey species like capybaras (Hydrochoerus hydrochaeris) being constricted to available surface water. However, a small number of sightings (n = 9) occurred during the rainy season. Fifty-six additional sightings were discarded due to a lack of distinguishable photographic evidence.
Barker/Robust design
We used a Barker/RD32 to estimate survival (S), detection probability by camera trap (p), availability to camera traps given previously inside the study area (a′′) and previously outside the study area (a′), probability of observation by tourists given alive (R) and probability of being dead but not recovered (R′), and abundance (N) of jaguars in Hato La Aurora. We extracted nine annual primary periods from camera trapping studies, eight of which occurred during the dry season and onset of the rainy season (between February and June) and one which occurred during the rainy season (July–October). We shortened camera trap studies to four-month secondary periods for closure28, as required by the model32. Camera trap records from outside of the closed period were removed from analysis. Details of the study periods can be found in Table A2.
Detection histories were compiled for each individual. Camera trap data were used for the secondary periods. Tourist photos, which could be collected at any time, were included as auxiliary resightings of jaguars at the end of each primary period in the detection history. The inclusion of tourist photos allowed for measurement of probability of viewing by tourists and increased the precision of survival estimates. We then constructed models using the Barker/RD model in Program Mark57, allowing for a mixture of detection probabilities58 and conditioning on at least one capture59 for a primary period. Dead recovery information was not available in our study. We therefore fixed parameters of the probability of dead recoveries (r) to 0 and fidelity (F) to 1 since all observations were within the study area. We also accounted for the variable time interval lengths between primary periods, rescaling all survival parameters to an annual basis. Hypotheses and parameter definitions can be found in Table A1.
We used a stepwise approach to model selection because of the large number of potential models associated with Barker/RD28,60. We began by fitting models of probability of detection (p) to test hypotheses of time and sex variation. We did not test for trap response (c) because of the non-invasive nature of remote cameras and the possibility of the model mistaking trap response for heterogeneity. We tested all models for heterogeneity with two mixtures because of jaguars’ territorial nature and the assumption that some individuals’ home ranges will only partially overlap with the study area, while others will be completely within it. We hypothesized that p increased by primary periods because researchers improved camera site identification and placement techniques, and that p varied by sex61. We tested for variation in p by primary period, secondary period, sex, and constant detection.
We modeled the availability parameters, which describe whether a jaguar was previously in the study area (a′′) or previously outside the study area (a′). We hypothesized that the probability of being previously outside the study area (a′) and inside the study area (a′′) would be Markovian and be the result of sex and time interactions, whereby males were more likely to emigrate and immigrate than females because of dispersal and territoriality62. We compared models with no movement, random movement, and Markovian movement.
The resighting parameter in this version of the Barker/RD calculates the probability that each jaguar is observed directly by a tourist that year. We fit models to the probability of resighting (R), testing sex, time, sex and time interaction, linear trend, and constant models. Given the increase in jaguar sightings reported by tourists in the last 5 years, we hypothesized that resightings increased with time since 2017 and varied by sex, given that males are more likely to use higher risk areas63. Resighting effort is an unaccounted covariate in our study, since tour operators did not record the number of tours, hours of effort, or number of viewers.
Finally, we tested models for survival based on our hypothesis that survival would increase with time. We evaluated models with time variation, time and sex interaction, sex, and constant survival probabilities. Previous studies have applied prey density covariates to the survival parameter of robust design frameworks64. However, prey data were not available for all years of the study, thus could not be applied to survival. We selected the most parsimonious models using AIC for each hypothesized model34. We used model averaging for the estimated parameters above and the derived parameter of abundance (N).
Derived parameters from the Delta method
We used the Delta method to calculate population changes over time27,33 and identify the source of abundance increases. We used derived parameter estimates of N to calculate the finite rate of change in abundance between sampling periods (λ), as Nt+1/Nt. We calculated the number of new recruits in the population in Hato La Aurora as Nt+1−Nt φ where φ is survival at time t + 133.
Spatially explicit capture recapture
We ran density analysis with 2022 data to complement robust design estimates and compare with a density survey in Hato La Aurora in 201424. We conducted density analysis fitting SECR models in a maximum likelihood framework65,66 in the R package “secr”67. SECR models identify individuals home range centers based on their spatial locations, then estimate density of these centers across an area that includes the camera grid6869. In addition to the standard capture-recapture assumptions, SECR models assume circular and constant home ranges during the survey, randomly distributed home range centers, and that the encounter rate of an individual with a trap decreases with increasing distance from the home range center following a predefined function68. We used the half-normal detection function where the probability of capture (p) of an individual (i) at a trap (j) decreases with distance (d) from the activity center as: Pij = g0 exp(− dij2/2σ2). The parameter g0 is the probability of capture when the trap is located exactly at the center of the home range, and sigma (σ) is parameter of the spatial scale over which detection declines away from the home range center68. As appropriate for camera trap data, we deployed the binomial encounter model (or Bernoulli model), enabling individuals to be captured at different camera stations during one sampling occasion (i.e., 24-h period) but only once at each station70,71. Like other felids, jaguars have different behavior and home ranges between sexes, hence we allowed both parameters g0 and σ to vary with sex of the individuals53,61 and compared four models using the AIC: the null model (SECR.0), a model where g0 varies between males and females (SECR.g0), a model where σ varies between males and females (SECR.σ), and a model where both g0 and σ vary between sexes (SECR.sex).
Data availability
The datasets for the SECR analysis are available in the appendices and all other data are available from the corresponding author on reasonable request.
References
Ripple, W. J. et al. Status and ecological effects of the world’s largest carnivores. Science (1979) 343, 6167 (2014).
di Minin, E. et al. Global priorities for national carnivore conservation under land use change. Sci. Rep. 6, 1–9 (2016).
Crooks, K. R., Burdett, C. L., Theobald, D. M., Rondinini, C. & Boitani, L. Global patterns of fragmentation and connectivity of mammalian carnivore habitat. Philos. Trans. R. Soc. B: Biol. Sci. 366, 2642–2651 (2011).
Devlin, A. L. et al. Drivers of large carnivore density in non-hunted, multi-use landscapes. Conserv. Sci. Pract. 5, 1 (2023).
Nyhus, P. J. Human-wildlife conflict and coexistence. Annu. Rev. Environ. Resour. 41, 143–171 (2016).
Lute, M. L., Carter, N. H., López-Bao, J. V. & Linnell, J. D. C. Conservation professionals agree on challenges to coexisting with large carnivores but not on solutions. Biol. Conserv. 218, 223–232 (2018).
Venumière-Lefebvre, C. C., Breck, S. W. & Crooks, K. R. A systematic map of human-carnivore coexistence. Biol. Conserv. 268, 109515 (2022).
Macdonald, D. W. et al. Understanding nuanced preferences for carnivore conservation: To know them is not always to love them. Glob. Ecol. Conserv. 37, e02150 (2022).
Dickman, A. J., Macdonald, E. A. & Macdonald, D. W. A review of financial instruments to pay for predator conservation and encourage human-carnivore coexistence. Proc. Natl. Acad. Sci. U. S. A. 108, 13937–13944 (2011).
Hyde, M. et al. Refining carbon credits to contribute to large carnivore conservation: The jaguar as a case study. Conserv. Lett. 15, 3 (2022).
Hazzah, L., Bath, A., Dolrenry, S., Dickman, A. & Frank, L. From attitudes to actions: Predictors of lion killing by Maasai Warriors. PLoS ONE 12, 258 (2017).
Marchini, S. & Macdonald, D. W. Predicting ranchers’ intention to kill jaguars: Case studies in Amazonia and Pantanal. Biol. Conserv. 147, 213–221 (2012).
Macdonald, C. et al. Conservation potential of apex predator tourism. Biol. Conserv. 215, 132–141 (2017).
Tortato, F., Izzo, T., Hoogesteijn, R. & Peres, C. The numbers of the beast: Valuation of jaguar (Panthera onca) tourism and cattle depredation in the Brazilian Pantanal. Glob. Ecol. Conserv. 11, 106–114 (2017).
Buckley, R. C., Morrison, C. & Castley, J. G. Net effects of ecotourism on threatened species survival. PLoS ONE 11, e0147988 (2016).
Bateman, P. W. & Fleming, P. A. Are negative effects of tourist activities on wildlife over-reported? A review of assessment methods and empirical results. Biol. Conserv. 211, 10–19 (2017).
Broekhuis, F. Natural and anthropogenic drivers of cub recruitment in a large carnivore. Ecol. Evol. 8, 6748–6755 (2018).
Mossaz, A., Buckley, R. C. & Castley, J. G. Ecotourism contributions to conservation of African big cats. J. Nat. Conserv. 28, 112–118 (2015).
Pereira, K. S., Gibson, L., Biggs, D., Samarasinghe, D. & Braczkowski, A. R. Individual identification of large felids in field studies: Common methods, challenges, and implications for conservation science. Front. Ecol. Evol. 10, 350 (2022).
Miller, J. R. B. et al. Effectiveness of contemporary techniques for reducing livestock depredations by large carnivores. Wildl. Soc. Bull. 40, 806–815 (2016).
Rafiq, K. et al. Tourist photographs as a scalable framework for wildlife monitoring in protected areas. Curr. Biol. 29, R681–R682 (2019).
Thornton, D. et al. Assessing the umbrella value of a range-wide conservation network for jaguars (Panthera onca). Ecol. Appl. 26, 1112–1124 (2016).
Jędrzejewski, W. et al. Estimating large carnivore populations at global scale based on spatial predictions of density and distribution—application to the jaguar (Panthera onca). PLoS ONE 13, e0194719 (2018).
Boron, V. et al. Jaguar densities across human-dominated landscapes in Colombia: The contribution of unprotected areas to long term conservation. PLoS ONE 11, 1–14 (2016).
Quigley, H. et al. Panthera onca, Jaguar. IUCN Red List 8235, 1–29 (2017).
Petracca, L. S. et al. Agricultural encroachment and lack of enforcement threaten connectivity of range-wide jaguar (Panthera onca) corridor. J. Nat. Conserv. 22, 436–444 (2014).
Harmsen, B. J. et al. Long term monitoring of jaguars in the Cockscomb Basin Wildlife Sanctuary, Belize; implications for camera trap studies of carnivores. PLoS ONE 12, e0179505 (2017).
Gutiérrez-González, C. E., Gómez-Ramírez, M. A., López-González, C. A. & Doherty, P. F. Are private reserves effective for jaguar conservation?. PLoS ONE 10, e0137541 (2015).
Srbek-Araujo, A. C. & Chiarello, A. G. Population status of the jaguar Panthera onca in one of its last strongholds in the Atlantic Forest. Oryx 51, 246–253 (2017).
Fragoso, C. E. et al. Unveiling demographic and mating strategies of Panthera onca in the Pantanal, Brazil. J. Mammal. https://doi.org/10.1093/jmammal/gyac123 (2023).
Payán, E. & Trujillo, L. A. The Tigrilladas in Colombia. CAT News 44 (2006).
Kendall, W. L. et al. Combining dead recovery, auxiliary observations and robust design data to estimate demographic parameters from marked individuals. Methods Ecol. Evol. 4, 828–835 (2013).
Karanth, K. U., Nichols, J. D., Kumar, N. S. & Hines, J. E. Assessing tiger population dynamics using photographic capture-recapture sampling. Ecology 87, 2925–2937 (2006).
Burnham, K. P. & Anderson, D. R. Model Selection and Multimodel Inference. Model Selection and Multimodel Inference—A Practical Information-Theoretic Approach (Springer Science, 2002).
Balme, G. A., Slotow, R. & Hunter, L. T. B. Impact of conservation interventions on the dynamics and persistence of a persecuted leopard (Panthera pardus) population. Biol. Conserv. 142, 2681–2690 (2009).
Soofi, M. et al. Assessing the relationship between illegal hunting of ungulates, wild prey occurrence and livestock depredation rate by large carnivores. J. Appl. Ecol. 56, 365–374 (2019).
Jędrzejewski, W. et al. Density and population structure of the jaguar (Panthera onca) in a protected area of Los Llanos, Venezuela, from 1 year of camera trap monitoring. Mamm. Res. 62, 9–19 (2017).
Rabinowitz, A. & Zeller, K. A. A range-wide model of landscape connectivity and conservation for the jaguar, Panthera onca. Biol. Conserv. 143, 939–945 (2010).
Robinson, H. S., Goodrich, J. M., Miquelle, D. G., Miller, C. S. & Seryodkin, I. V. Mortality of Amur tigers: The more things change, the more they stay the same. Integr. Zool. 10, 344–353 (2015).
Knight, J. Making wildlife viewable: Habituation and attraction. Soc. Anim. 17, 167–184 (2009).
Eshoo, P. F., Johnson, A., Duangdala, S. & Hansel, T. Design, monitoring and evaluation of a direct payments approach for an ecotourism strategy to reduce illegal hunting and trade of wildlife in Lao PDR. PLoS ONE 13, 2 (2018).
Grünewald, C., Schleuning, M. & Böhning-Gaese, K. Biodiversity, scenery and infrastructure: Factors driving wildlife tourism in an African savannah national park. Biol. Conserv. 201, 60–68 (2016).
Gerber, B. D., Ivan, J. S. & Burnham, K. P. Estimating the abundance of rare and elusive carnivores from photographic-sampling data when the population size is very small. Popul. Ecol. 56, 463–470 (2014).
Ohrens, O. et al. Predator tourism improves tolerance for pumas, but may increase future conflict among ranchers in Chile. Biol. Conserv. 258, 109150 (2021).
Van Der Meer, E. & Dullemont, H. Human-carnivore coexistence: Factors influencing stakeholder attitudes towards large carnivores and conservation in Zimbabwe. Environ. Conserv. 48, 48–57 (2021).
Salerno, J., Borgerhoff-Mulder, M., Grote, M. N., Ghiselli, M. & Packer, C. Household livelihoods and conflict with wildlife in community-based conservation areas across northern Tanzania. Oryx 50, 702–712 (2016).
Clerici, N. et al. Peace in Colombia is a critical moment for Neotropical connectivity and conservation: Save the northern Andes-Amazon biodiversity bridge. Conserv. Lett. 12, e12594 (2019).
Vélez, D. et al. Distribution of birds in Colombia. Biodivers. Data J. https://doi.org/10.3897/BDJ.9.e59202 (2021).
Suárez-Castro, A. F. et al. Vacíos de información espacial sobre la riqueza de mamíferos terrestres continentales de Colombia. Caldasia 43, 247–260 (2021).
Huertas-Ramirez, H. & Huertas-Herrera, A. Historiografía de la ganadería en la Orinoquía. AICA 6, 300–307 (2015).
Romero-Ruiz, M. H., Flantua, S. G. A., Tansey, K. & Berrio, J. C. Landscape transformations in savannas of northern South America: Land use/cover changes since 1987 in the Llanos Orientales of Colombia. Appl. Geogr. 32, 766–776 (2012).
IDEAM. Distribucion de la temperatura media anual (C) Promedio Multianual 1981–2010. http://atlas.ideam.gov.co/basefiles/Temp_Med_Anual.pdf (2014).
Tobler, M. W. & Powell, G. V. N. Estimating jaguar densities with camera traps: Problems with current designs and recommendations for future studies. Biol. Conserv. 159, 109–118 (2013).
Otis, D. L., Burnham, K. P., White, G. C. & Anderson, D. R. Statistical inference from capture data on closed animal populations. Wildl. Monogr. 1978, 3–135. https://doi.org/10.2307/3830650 (1978).
White, G. C. Capture-Recapture and Removal Methods for Sampling Closed Populations (Los Alamos National Laboratory, 1982).
Morato, R. G. et al. Space use and movement of a neotropical top predator: The endangered jaguar. PLoS ONE 11, 168 (2016).
White, G. C. & Burnham, K. P. Program MARK: Survival estimation from populations of marked animals. Bird Study 46, S120–S139 (1999).
Pledger, S. Unified maximum likelihood estimates for closed capture-recapture models using mixtures. Biometrics 56, 434–442 (2000).
Huggins, R. M. Some practical aspects of a conditional likelihood approach to capture experiments. Biometrics 47, 725 (1991).
Doherty, P. F., White, G. C. & Burnham, K. P. Comparison of model building and selection strategies. J. Ornithol. 152, 317–323 (2012).
Sollmann, R. et al. Improving density estimates for elusive carnivores: Accounting for sex-specific detection and movements using spatial capture-recapture models for jaguars in central Brazil. Biol. Conserv. 144, 1017–1024 (2011).
Kantek, D. L. Z. et al. Jaguars from the Brazilian Pantanal: Low genetic structure, male-biased dispersal, and implications for long-term conservation. Biol. Conserv. 259, 109153 (2021).
Conde, D. A. et al. Sex matters: Modeling male and female habitat differences for jaguar conservation. Biol. Conserv. 143, 1980–1988 (2010).
M’soka, J., Creel, S., Becker, M. S. & Droge, E. Spotted hyaena survival and density in a lion depleted ecosystem: The effects of prey availability, humans and competition between large carnivores in African savannahs. Biol. Conserv. 201, 348–355 (2016).
Borchers, D. L. & Efford, M. G. Spatially explicit maximum likelihood methods for capture-recapture studies. Biometrics 64, 377–385 (2008).
Efford, M. G., Dawson, D. K. & Borchers, D. L. Population density estimated from locations of individuals on a passive detector array. Ecology 90, 2676–2682 (2009).
Efford M. Secr: Spatially explicit capture-recapture models. R package version 4.6.0, https://CRAN.R-project.org/package=secr (2023).
Efford, M. G. Density estimation in live-trapping studies. Oikos 106, 598–610 (2004).
Royle, J. A. & Young, K. V. A hierarchical model for spatial capture recapture data. Ecology 89, 2281–2289 (2008).
Royle, J. A., Nichols, J. D., Karanth, K. U. & Gopalaswamy, A. M. A hierarchical model for estimating density in camera-trap studies. J. Appl. Ecol. 46, 118–127 (2009).
Noss, A. J., Polisar, J., Maffei, L., Garcia, R. & Silver, S. Evaluating jaguar densities with camera traps. Wildlife Conserv. Soc. 2013, 1–78 (2013).
Acknowledgements
We thank J. Gallo, J. Gonzalez, R. Hoogesteijn, A. Mejía, J. Morgan, R. Ortiz, L. Pérez, S. Quintero, A. Pizarro, G. Schaller, C. Trujillo, C. Valderrama and all the field guides for their help in camera trapping studies. We thank J.K. Young, T.L. Teel, G. Wittemyer, and J. Goodrich for comments to improve the manuscript. We also thank L.L. Bailey, and C. Venumière-Lefebvre for orientation and suggestions to improve the analysis. Finally, we thank the Barragan family for their commitment to conservation and allowing us to carry out these studies on their ranch. Any use of trade, firm, or product names is for descriptive purposes only and does not imply endorsement by the U.S. Government. Questions about the data used in the manuscript can be directed to the first author.
Funding
We are grateful for the support of the Bobolink Foundation, National Geographic Society, US Fish and Wildlife Service, and USAID’s Natural Wealth Program for funding camera trap studies and conservation interventions, the Center for Human-Carnivore Coexistence for support for M. Hyde, Panthera for support for E. Payán, D. Stasiukynas, S. Rincón, J. Rodríguez and V. Boron, and the US Department of Agriculture for support of S.W. Breck.
Author information
Authors and Affiliations
Contributions
M.H., V.B., J.B., E.P., and D.S. designed the study. E.P., J.B., M.H., V.B., K.R.C., S.W.B. acquired funding. J.B., V.B., E.P., D.S., S.R., M.H., and J.R. collected data. M.H., V.B., W.L.K, D.S. and S.R. curated and analyzed the data. M.H. wrote the main manuscript text. V.B., K.R.C., S.W.B., D.S., S.R., W.L.K., J.R. edited the manuscript. All authors reviewed and approved the manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Hyde, M., Payán, E., Barragan, J. et al. Tourism-supported working lands sustain a growing jaguar population in the Colombian Llanos. Sci Rep 13, 10408 (2023). https://doi.org/10.1038/s41598-023-36935-2
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-023-36935-2
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
