
Abstract
This study reports a set of primeval marine incursions identified in two drill cores, 1PS-06-CE, and 1PS-10-CE, which recovered the Barbalha Formation, Araripe Basin, Brazil. Based on a multi-proxy approach involving stratigraphy, microbiofacies, ichnofossils, and microfossils, three short-lived marine incursions were identified, designated Araripe Marine Incursions (AMI) 1–3. AMI-1 and AMI-2, which occur within the shales of the Batateira Beds (lower part of the Barbalha Formation), were identified by the occurrence of benthonic foraminifera, calcareous nannofossils, dinocysts, and a mass mortality event of non-marine ostracods. AMI-3 was recognized in the upper part of the Barbalha Formation, based on the occurrence of ichnofossils and planktonic foraminifera. The observation of the planktonic foraminifera genus Leupoldina for the first time in the basin indicates early Aptian/early late Aptian age for these deposits, and the first opportunity of correlation with global foraminifera biozonation. Our findings have implications for the breakup of the Gondwana Supercontinent, as these incursions represent the earliest marine-derived flooding events in the inland basins of northeastern Brazil.
Similar content being viewed by others
Introduction
The Aptian (126.3 to 113.1 Ma1) is an important stage of the Early Cretaceous, characterized by significant paleogeographic and paleoceanographic events around the world. Oceanic gateways gradually opened during this stage, and rising sea levels allowed the dispersion of marine biota (e.g.2,3). During the Early Cretaceous, the Gondwana Supercontinent broke up into the South American and African continents, leading to the establishment of the South Atlantic Ocean (e.g.4).
The initial evolution of the South Atlantic Ocean has been widely debated in terms of relative motion and timing. Reversal-related magnetic anomalies indicate that onset of the opening occurred around Anomaly M13 (134 Ma5) in the Austral Atlantic Ocean, but the Cretaceous Normal Polarity Superchron (C34n; 121 to 83.65 Ma6) complicates age determination for its subsequent spread northwards, between the Barremian and Aptian interval.
Identification of the source and pathways taken by the first marine flooding episodes from the South Atlantic Ocean into the Brazilian intracratonic sedimentary basins is further complicated by conflicting results based on paleontological and sedimentary proxies (e.g.7,8,9). Paleontological data supports a clear connection with Tethyan sea waters (e.g.7), while a combination of stratigraphy and geometry of the deposits establishes a transgression path from south to north and further inland8,10,11.
The geological record of northeastern Brazilian marginal and interior basins is fundamental for obtaining temporal and geographical constraints for the first marine incursions in the region, including their provenance and relationship with the relative motions between the incipient African and South American plates. A combination of paleontological, sedimentological, stratigraphic, and geochemical approaches is essential to obtain a clearer image of these ingressions and the relationship between the Tethys and the proto-South Atlantic waters, as well as the development of a more accurate paleogeographical model for this time interval. The recognition of marine incursions within these basins provides essential information to help decipher the pathways of this major geological event.
In this study, we provide a multi-proxy analysis of two boreholes, 1PS-06-CE and 1PS-10-CE, from the eastern portion of the Araripe Basin, Northeast Brazil. The origin of this large interior basin is related to the breakup of the Gondwana11,12,13,14, and late Aptian–early Albian marine incursions have already been identified (e.g.7,8,15,16,17,18,19,20). Our work yielded new ichnological, micropaleontological, and microbiofaciological data for the Barbalha Formation, the lowermost stratigraphic unit of the post-rift sequence of the basin and allowed identification of the primeval marine incursions in northeastern Brazil.
Geological setting
The Araripe Basin is a large inland basin in northeastern Brazil, associated with the opening of the South Atlantic Ocean10. Its development, along several other rift basins in Brazil, was triggered by the breakup of the Gondwana Supercontinent during the Cretaceous11,12,13,14,21, and its deposits lay atop Precambrian terrains (Piancó-Alto Brígida and Granjeiro), in the transversal domain of the Borborema Province, to the south of the Patos Shear Zone.
The Santana Group (sensu Neumann and Assine22) is the stratigraphic record of the local Alagoas Stage (Aptian/lower Albian), equivalent to the post-rift I megasequence11. The precise age of the deposits of this group is the subject of considerable debate (e.g.7,15,18,19,23,24,25). Though typically assigned to the Aptian, several studies place the upper part of this unit (Romualdo Formation) in the Aptian–Albian interval (e.g.15,23).
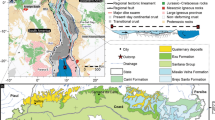
The sedimentary succession of the Santana Group is represented by the Barbalha, Crato, Ipubi, and Romualdo formations (Fig. 1), from base to top, with mixed carbonate-siliciclastic deposits8,11. The Barbalha Formation, focus of this study, is divided into two parts8. The lower one is essentially siliciclastic, passing from a fluvial to a lacustrine continental environment in a transgressive pattern that culminates in the Batateira Beds, a laterally extensive layer of mudstones and marls that are an important stratigraphic marker for the basin8. The type of preservation of the fossils in these beds corroborates an interpretation of continental environment setting, with dysoxic/anoxic intervals11,12,26,27,28. The upper part of the Barbalha Formation is composed of a siliciclastic succession of sandstones and mudstones with a fining upward pattern, that ends with the deposition of the laminated limestones of the Crato Formation reflecting a change from fluvial to hypersaline lacustrine environments (e.g.29).

Geographic location and stratigraphic position of the studied area. (a) Map of South America and studied area (red polygon). (b) Simplified geological map of the Araripe Basin with location of the boreholes used in this study, modified from Assine et al.8. (c) Stratigraphic framework of the Alagoas Post-rift Stage in the Araripe Basin, composed of three depositional sequences bounded by disconformities. DS-1 depositional sequence 1, DS-2 depositional sequence 2, DS-3 depositional sequence 3, modified from Assine et al.8. Graphic art made with CorelDRAW Graphics Suite × 6, version 16.0 (https://www.coreldraw.com/).
Widespread marine ingressions have been reported in the Romualdo Formation, during the late Aptian of the basin8,30. Its deposits of black shales with fossil-bearing carbonate concretions (e.g.16,31) are famous for their well-preserved fossils, and include continental (palynomorphs and ostracods15) and marine groups (dinoflagellates18, benthonic and planktonic foraminifera19,20, marine ostracods20, and calcareous nannofossils20). The distribution of these marine organisms, many related to the Tethyan Realm, suggests short marine incursions from the Central Atlantic Ocean (e.g.7,20,32,33), which could have occurred due to sufficient tectonic subsidence rates, accompanied by sea level rise episodes9,34 at different times during the Early Cretaceous.
Results
Sedimentology and ichnology
We recognized 16 sedimentary facies in the Barbalha Formation (Table 1). The lower part of this unit comprises sandstones and rare muddy deposits, overlain by laminites (Batateira Beds), with low ichnodiversity and low rates on the bioturbation scale (BS). The upper part is characterized by sandstone cosets (locally bioturbated), occasionally intercalated with fine-grained deposits (Fig. 2), overlaid by dominantly muddy deposits with interbedded sandstones (Fig. 3).

Trace fossils identified in the ichnofabrics of the studied section from Barbalha Formation (1PS-06-CE and 1PS-10-CE boreholes). (a–c) Ophiomorpha (Op), Scolicia (Sc), Palaeophycus (Pa), and Skolithos (Sk) (1PS-06-CE, 66.40 m). (d) Scolicia (Sc), Thalassinoides (Th), and Planolites (PL) (1PS-06-CE, 57.00 m). (e,f) Scolicia (Sc), Thalassinoides (Th), Diplocraterion (Di), Palaeophycus (Pa), Ophiomorpha (Op), and Lockeia (Lo) (1PS-06-CE, 68.50 m). (g) Thalassinoides (Th), Helminthopsis (He), Planolites (Pl), and Palaeophycus (Pa) (1PS-06-CE, 54.50 m). (h) Diplocraterion (Di), Thalassinoides (Th), and Scolicia (Sc) (1PS-10-CE, 33.50 m). Scale bars: 10 mm.

NW–SE stratigraphic section (location in Fig. 1b). Rio da Batateira sedimentary section adapted from Assine et al.8 and Varejão et al.34. Graphic art made with CorelDRAW Graphics Suite × 6 version, 16.0 (https://www.coreldraw.com/).
Ostracods
Eight species belonging to three genera of non-marine ostracods were identified in borehole 1PS-06-CE (Fig. 4a–h), with moderate to good preservation. Recovered species include Candonopsis? alagoensis, Candona? sp., Pattersoncypris alta, Pattersoncypris micropapillosa, Pattersoncypris salitrensis, Pattersoncypris angulata, Pattersoncypris sp. 1, and Pattersoncypris sp. 2.

Plate illustrating key microfossil species recovered from boreholes 1PS-06-CE and 1PS-10-CE. Ostracods: (a) Candonopsis? alagoensis (right lateral view; 1PS-06-CE, 99.30 m); (b) Cypridea araripensis (mold; 1PS-10-CE, 51.20 m); (c) Brasacypris subovatum (right lateral view; 1PS-10-CE, 95.76 m); (d) Pattersoncypris alta (right lateral view; 1PS-06-CE, 99.30 m); (e) Pattersoncypris salitrensis (right lateral view; 1PS-06-CE, 99.30 m); (f) Pattersoncypris angulata17 (1PS-06-CE, 99.30 m); (g) Pattersoncypris micropappilosa (right lateral view; 1PS-06-CE, 99.30 m); (h) Theriosynoecum silvai (right valve, 1PS-10-CE, 95.76 m). Foraminifera: (i–m) Bathysiphon sp. ((i,j) 1PS-06-CE, 117.90 m; (k–m) 1PS-10-CE, 103.69 m, and 102.90 m). Calcareous nannofossils: (n–s) Ascidian spicule (1PS-06-CE, 99.30 m). (t–u) Thoracosphaera spp. (1PS-06-CE, 114.70 m).
Ostracod abundance varies throughout this borehole. Between depths 112.30 and 92.40 m, which encompasses the Batateira Beds, there is a remarkably high abundance of well-preserved and articulated specimens (particularly at depths 99.80 and 99.30 m, with over 1000 individuals in each sample), including many younger ontogenetic stages. Between depths of 91.45 and 13.60 m, abundance and diversity of the ostracod fauna decreases considerably.
Comparatively, abundance and diversity are lower in borehole 1PS-10-CE, and fossil preservation is moderate to poor. The samples with the highest abundance are from the laminites of the Batateira Beds (101.54 m). Six species belonging to six genera of non-marine ostracods were recovered (Fig. 4a–h): Candonopsis? alagoensis, Cypridea araripensis?, Brasacypris subovatum, Pattersoncypris sp. 3, Theriosynoecum silvai, as well as one Gen. et sp. indet. ostracod. Ostracod distribution can be observed in Fig. 3.
Benthonic foraminifera
Abundant, monofaunistic associations of agglutinated (textularids) benthonic foraminifera, classified as Bathysiphon sp., were found near the base of the Batateira Beds in both boreholes (117.90 m in 1PS-06-CE and 105.90, 103.69, 103.20, and 102.90 m in 1PS-10-CE) (Fig. 4i–m). Specimens are well-preserved but usually fragmented. Their distribution can be observed in Fig. 3.
Calcareous nannofossils
Calcareous nannofossils were recovered only in borehole 1PS-06-CE, in the fine-grained facies of the Batateira Beds (Fig. 4n–u). We recorded poorly- to well-preserved ascidian spicules at 112.30 m and five specimens of calcareous dinocyst fragments (Thoracosphaera spp.) at 114.70 m.
We also observed 24 specimens of ascidian spicules at 99.30 m, the most abundant calcareous nannofossil assemblage of the borehole, and which are secreted by small (< 0.1 mm) soft-bodied tunicates that live in shallow water environments. They show diverse morphologies (rounded, spheric, and radial) as well as dissolution/overgrowth preservation effects. Four specimens of Thoracosphaera spp. found at 99.30 m have morphologies marked by fragments with large inner element sizes and low preservation. Calcareous nannofossil distribution can be observed in Fig. 3.
Microbiofacies
Based on thin sections of boreholes 1PS-06-CE and 1PS-10-CE, we observed three horizons with marine microfossils (textularids, serpulids, and planktonic foraminifera) in the Barbalha Formation (Figs. 4l and 5a–j). The first horizon is a mudstone with abundant textularids, Bathysiphon sp. (at 103.69 m in borehole 1PS-10-CE). The second horizon consists of wackestone and packstone laminites (99.30 m in borehole 1PS-06-CE, and 100.63 m and 100.38 m in borehole 1PS-10-CE), with abundant ostracods, serpulid tubes (Fig. 5d–j), microbial pellets, organic matter, and calcareous concretions. Two distinct ostracod deposits were preserved in these laminites: a packstone with disarticulated (reworked) and compressed ostracod valves (Fig. 5i–j), and a wackestone with articulated, adult- to juvenile-sized ostracod carapaces, associated with serpulid tubes in an organic-rich matrix (Fig. 5j), interpreted as in situ. The serpulids observed are 50–250 µm long tubes with thin microcrystalline calcite walls (dark color in thin sections) and usually well-developed collars (Fig. 5d). Transverse sections are circular to ovoid, well-preserved, with partial dissolution and locally compressed. They are most abundant on the organic muddy microbial matrix among ostracod valves (Fig. 5g–j) but are absent in the ostracod packstone.

Key microfossil species recovered (1PS-06-CE and 1PS-10-CE boreholes): (a) Leupoldina sp. (1PS-06-CE, 39.20 m); (b) Globigerinelloides cf. barri (1PS-06-CE, 39.20 m); (c) Globigerinelloides cf. ferreolensis (1PS-06-CE, 39.20 m); (d) Serpulids (collar = white arrows) (1PS-10-CE, 100.45 m); (e,f) Serpulids (1PS-06-CE, 99.30 m); (g,h) Wackestone and packstone with ostracods (1PS-06-CE, 99.30 m); (i) Wackestone with ostracods and serpulids (red frame corresponds to image “(j)”) (1PS-06-CE, 99.30 m); (j) Detail view of wackestone with ostracod shells and serpulid tubes (red arrows) (1PS-06-CE, 99.30 m); (k) Sergipea variverrucata (1PS-10-CE, 104.2 m); (l) Botryococcus sp. (1PS-10-CE, 104.2 m); (m) Oligosphaeridium sp. (1PS-06-CE, 115.8 m); and (n) Cometodinium sp. (1PS-10-CE, 101.5 m; fluorescence mode).
The third horizon is represented by the reddish-brown, cemented mudstone of the F3 facies (39.20 m depth, borehole 1PS-06-CE). Planktonic foraminifera specimens are frequent (Fig. 5a–d) and include the genera Leupoldina and Globigerinelloides. Most specimens have undergone some degree of dissolution, and internal molds, partially dissolved tests, and ghosts are prevalent. Poor preservation hinders species-level identification for leupoldinids (described as Leupoldina sp.) and globigerinelloidids (Globigerinelloides cf. barri and G. cf. ferreolensis) (Fig. 5a–c).
Genus Leupoldina is characterized by pseudo-planispiral coiling, radially elongate chambers, and branched disaxial outer chambers, which makes the last whorl asymmetrical. In thin sections, early chambers are globular, and final chambers may be elongated, ovoid to circular, depending on the orientation of the cut, commonly with paired projections arranged symmetrically on each side of a median plane. Paired projections (branching) of final two chambers are typical for most species of Leupoldina (Fig. 5a). Globigerinelloides specimens were identified by their typical planispiral biumbilicate coiling in equatorial sections.
Organic-walled microfossils
Pollen grains and plant spores (e.g., Classopollis classoides, Inaperturopollenites spp., Araucariacites australis and Cicatricosisporites spp.) are responsible for the majority of palynomorph counts in samples from boreholes 1PS-06-CE and 1PS-10-CE, followed by freshwater algae (Botryococcus sp. and Pediastrum spp.), with punctual occurrences in the Batateira Beds of dinocysts, Oligosphaeridium sp. at 115.80 m on 1PS-06-CE, and Cometodinium sp. at 101.54 m on 1PS-10-CE (Fig. 5k–n).
Discussion
Biostratigraphic framework and age controls
The stratigraphic positions of the biomarkers discussed in this chapter are shown in the Supplementary Material (SM).
The absence of globally distributed marine microfossils in the Brazilian interior basins has historically prevented their correlation with global chronostratigraphic charts, therefore they have only been calibrated using local biozones so far. Here we report for the first time the occurrence of Leupoldina spp. in the Barbalha Formation, in thin sections (Fig. 5a).
Leupoldina has numerous occurrences in the Tethyan region35, and is used in global foraminiferal biozonation schemes36,37 due to its distinct morphology from other planktonic foraminifera of this interval. Poor preservation and cut orientation limited species-level identification of Leupoldina specimens, but their occurrence, associated with Globigerinelloides, does allow us to assign biozones L. cabri to G. algerianus36. corresponding to the early Aptian/early late Aptian stage1 (Fig. 6). This outcome challenges the local biostratigraphic framework15,38 showing that the chronostratigraphic framework for the post-rift sequence in the Araripe Basin must be reviewed.

Planktonic foraminifera zonation scheme for the late Barremian–Aptian interval. Planktonic foraminifera range is taken from Verga and Premoli-Silva35,37 (adapted from Ogg et al.1). Graphic art made with CorelDRAW Graphics Suite × 6, version 16.0 (https://www.coreldraw.com/).
Concerning the local zonation schemes, the presence of the age-indicative ostracods Pattersoncypris spp. and Theriosynoecum silvai at boreholes 1PS-06-CE and 1PS-10-CE indicates local Alagoas Stage (Aptian–early Albian) for both sites (Zone 011; also called the Pattersoncypris and Krommelbeincypris Zone39).
Furthermore, the occurrence of age-indicative pollen grains belonging to the species Sergipea variverrucata (local Zone P-27040) constrains the studied intervals to a late Aptian age.
The lack of representatives of typical lower Aptian palynomorph-based biozones is probably due to paleoclimate exclusion41. We contend that paleoclimate dynamics might have controlled the local appearance and disappearance of plant species, which directly affected the palynostratigraphic records.
Multi-proxy evidence of marine incursions
Three marine transgressive events, here named Araripe Marine Incursions (AMI), are clearly identifiable in the Barbalha Formation (Fig. 7). The Depositional Sequence 1 (sensu Assine et al.8) starts with very coarse- to coarse-grained sandstone facies (St2), superimposed by layers of fine- to very fine-grained sandstone with ripples (Sr) (Fig. 3; Table 1). Muddy levels (F6 and F1) are observed in lateral associations, sometimes interbedded with rippled sandstone (Sr), and as suggested previously, correspond to fluvial channels with laterally well-developed overbank42 (Fig. 3; Table 1).

Plots of marine proxies’ analyses, with salinity indicators and identification of marine incursions in (a) 1PS-10-CE borehole, and (b) 1PS-06-CE borehole. Ostracod, palynomorph, and foraminifera biozones according to Poropat and Colin39, Regali and Silva Santos40, and Ogg et al.1, respectively. Graphic art made with CorelDRAW Graphics Suite × 6, version 16.0 (https://www.coreldraw.com/).
Paleocurrent data reveal preferential flow towards SE8,10,27,42. The presence of agglutinating foraminifera in the sandstone deposits (1PS-10-CE) indicates brackish conditions possibly due to deltaic influence in this system.
The Batateira Beds43 overlie this fluvial system, with intercalating siliciclastic (F1 and F2) and heterolithic facies (Hsr). They have been largely classified as a lacustrine system (Fig. 3; Table 1) due to the occurrence of non-marine elements such as ostracods, fish, and continental palynomorphs (e.g.11,44). We have also observed an abundance of non-marine ostracods (Pattersoncypris micropapillosa, P. salitrensis, P. angulata, Pattersoncypris sp. 1, Pattersoncypris sp. 2, Candonopsis? alagoensis, and Candona? sp.) in these deposits (Fig. 4a–h). However, we present ichnological and micropaleontological data that supports input of marine waters in two distinct intervals, producing a salinity gradient between brackish to freshwater conditions in this lacustrine paleoenvironment (Fig. 7).
The first marine incursion (AMI-1) can be observed in both boreholes, in marls (facies M) just above transgressive surface TS-1 (Fig. 3 and 7). It is marked by abundant agglutinated foraminifera (Bathysiphon sp.) and the occurrence of calcareous and organic-walled dinocysts, marine elements that occur in association with an ichnofabric composed of Planolites, Palaeophycus, and Thalassinoides. The low ichnodiversity and occurrence restricted to facies F1 points to the establishment of opportunistic colonization due to the onset of brackish conditions45,46.
We identified the second marine incursion restricted to borehole 1PS-06-CE, AMI-2, in the middle part of the Batateira Beds, associated with its laminite deposits (C facies). It corresponds to a maximum flooding surface (MFS-1) (Figs. 3 and 7), recognized due to recovery of marine elements such as calcareous dinocysts (Thoracoshpaera spp.), calcareous nannofossils (ascidian spicules), and serpulid tubes. The thickness of these lacustrine deposits increases towards southeast, reaching 30 m in borehole 1PS-06-CE (Fig. 3). The presence of these marine elements, resting above the fluvial facies of both wells, reveals a generalized flooding event associated with an increase in the relative sea level.
Non-marine ostracod carapaces are particularly abundant in borehole 1PS-06-CE at the AMI-2 event (more than 1000 specimens recovered per sample at depths 99.80 and 99.30 m). The assemblage contains a large amount of well-preserved juvenile instars and adults, all with closed carapaces, indicating that they died at those life stages (Fig. 5g–j). Interestingly, carapaces of adult Pattersoncypris, a mixohaline genus, are smaller than usual47. All these features are likely result of development in a stressful environment and, eventually, a mass mortality event (e.g.48,49). Salinity changes in the depositional setting of the Batateira Beds could be responsible, as these ostracods did not tolerate marine conditions. This mortality event, associated with the presence of serpulids (Fig. 5i–j), foraminifera, calcareous nannofossils, and calcareous dinoflagellates, reinforces the hypothesis of establishment of fully marine conditions during this interval. Towards the top, the Batateira Beds record the reestablishment of lacustrine environments following a sea level drop.
The overlying coarse-grained facies record subaerial exposure, reactivation of the fluvial system, and the beginning of Depositional Sequence 2 (sensu Assine et al.8 and Scherer et al.42) generating a clear sequence boundary (SB; Fig. 3). Paleocurrent measurements from outcrops (e.g., Rio Batateira) show paleoflow direction mainly towards southeast8,10,27,42,44. Sandy deposits (Gt, Sgt, St1, and St2) of fluvial channels are predominant in proximal settings (borehole 1PS-10-CE), and facies F2 and F3 represent overbank deposits in an underdeveloped floodplain within the river system27,34,42. The blocky and slickensides structures, associated with rhizobioturbation in facies F3 also indicate eventual subaerial exposure.
Distally, in borehole 1PS-06-CE and above the SB, bioturbation is characterized by ichnofabrics composed of Planolites, Palaeophycus, Thalassinoides, and Cylindrichnus, as well as monospecific Thalassinoides ichnofabrics (Hsr, F1, and F3). The low ichnodiversity and low bioturbation scale values (BS = 1–2) reflect stressful conditions45,46 and are related to the colonization of softgrounds, possibly stressed by salinity fluctuations (e.g.50). They are an impoverished expression of the Cruziana ichnofacies due to the introduction of brackish conditions in the distal portions of the fluvial setting.
The beginning of AMI-3 is identified by the gradual increase of ichnodiversity and bioturbation scale values towards the top in distal settings. Bioturbation in facies Srd, Srw, and St2 is represented by ichnofabrics composed of Diplocraterion, Ophiomorpha, Palaeophycus, Planolites, Thalassinoides, Chondrites, Helmintopsis, Scolicia, Lockeia, and Skolithos (BS 1–5) (Table 1), indicating establishment of an impoverished expression of the mixed Skolithos-Cruziana ichnofacies (e.g.50). These ichnological characteristics might reflect stressful conditions caused by salinity changes. Moreover, the presence of Scolicia associated with the Srw lithofacies suggests that salinity was sufficient to sporadically support the establishment of a stenohaline fauna.
Scolicia has been mainly recorded in marine environments, from shoreface to deep-sea (e.g.45), and represents bioturbation by spatangoids echinoderms, which are truly marine organisms (e.g.51). The record of Scolicia in stressed substrates of distal, brackish, tidally-dominated, or -influenced channels is scarce (e.g.52). In addition, the higher ichnodiversity (marine elements) suggests increased marine conditions towards the top (Fig. 3).
These fluvial facies are overlain by muddy deposits (F3, F2, and F1) interbedded with sandstones (Sr, Srd, St2, and Sm), interpreted as bayhead delta deposits34 and suggesting flooding of the fluvial system (Fig. 3; Table 1). The ichnological assemblages and micropaleontological data support this interpretation of flooding in both boreholes.
In both boreholes, the bioturbation above the TS-2 is characterized by ichnofabrics composed of Diplocraterion, Thalassinoides, Palaeophycus, Planolites, Chondrites, Cylindrichnus, Helminthopsis, Scolicia, and Skolithos, with BS 3–4, and are restricted to the Srd lithofacies (Fig. 3). They represent the establishment of an impoverished expression of the mixed Skolithos-Cruziana ichnofacies (e.g.50) and continuous marine influence in the deposits at 1PS-06-CE.
The higher ichnodiversity, including Scolicia burrows in 1PS-06-CE, suggests that marine conditions were more stable in the southeastern area. In addition, the occurrence of planktonic foraminifera (Leupoldina sp. and Globigerinelloides spp.) in 1PS-06-CE, laterally associated with ichnological assemblages of the impoverished mixed Skolithos-Cruziana ichnofacies of 1PS-10-CE, shows that marine to brackish conditions followed the onset of the bayhead delta. The presence of Leupoldina sp. and Globigerinelloides spp. also points to the establishment of typical marine conditions in the upper Barbalha Formation, configuring the second maximum flooding surface (MFS-2) (Fig. 3).
Conclusions
We dated the deposits of the Barbalha Formation for the first time based on the recovery of the planktonic foraminifera genera Leupoldina and Globigerinelloides. These genera mark the L. cabri to G. algerianus zones, which corresponds to the early Aptian/early late Aptian interval.
Three marine incursions (AMI-1 to AMI-3) were identified in the Barbalha Formation based on a multi-proxy analysis (micropaleontological, ichnological, and sedimentary analyses). The lower two occur in the Batateira Beds (lower Barbalha Formation), recording the primeval marine incursions into the Araripe Basin related to the breakup of the Gondwana, and the opening of the South Atlantic Ocean.
AMI-1, near the base of the Batateira Beds, is characterized by an abundance of the agglutinated foraminifera, and minor occurrence of organic-walled dinocysts and calcareous dinocysts.
AMI-2, in the laminite deposits of the Batateira Beds of borehole 1PS-06-CE, is characterized by calcareous dinocysts, ascidian spicules, microbial peloids, and abundant serpulid tubes. The mass mortality event of non-marine ostracods reinforces this marine incursion event.
AMI-3, in the upper part of the Barbalha Formation, is characterized by the occurrence of bioturbation mainly represented by Scolicia, Diplocraterion, Thalassinoides, Ophiomorpha, Skolithos, and Chondrites, as well as the presence of planktonic foraminifera Leupoldina spp. and Globigerinelloides spp.
These incursions are the oldest recorded so far related to the breakup of the Gondwana Supercontinent and the opening of the South Atlantic Ocean. Future studies with fossil records, geochemistry, and paleomagnetism will contribute to the characterization of these marine deposits, as well as the routes taken by these marine incursions.
Materials and methods
Sedimentary sections
We described and analyzed core samples taken from boreholes 1PS-06-CE and 1PS-10-CE, drilled by the Santana II Project53 in the eastern portion of the Araripe Basin, northeastern Brazil. Lithological, ichnological, microbiofaciological, and micropaleontological analyses were completed, preferentially done in the fine siliciclastic and limestone lithologies (shales, mudstones, siltstones, marls, and laminites), but also in some sandstone levels. For further details, we provide the full dataset of microfossils/microbiofacies in the supplementary materials (SM).
Sedimentological, ichnological, and stratigraphic analysis
The characterization of the sedimentary facies of boreholes 1PS-06-CE and 1PS-10-CE followed the usual methods, with description of physical sedimentary structures and basic lithology, focusing on lithofacies and ichnology. Intervals with no recovery were interpreted based on the accompanying well drilling data, namely gamma-ray values (indirect data); a small portion was obtained from cut samples. For the purposes of stratigraphic correlation, the laminite deposits of the Batateira Beds, in the lower part of the Barbalha Formation, were chosen as our datum; specifically, the levels containing serpulids and the ostracods mass mortality event, which occur on both drill cores.
Direct macroscopic observation was used to describe trace fossils, following the main ichnotaxobases applicable to ichnofabrics: burrow limit, burrow infill, branching pattern, and presence/absence of spreiten. The guidelines used for ichnotaxonomy classification followed Bertling et al.54. In some cases, identification of ichnotaxa was hampered by the loss of ichnotaxonomical features, dark lithology, and/or core deformation, which erased this data, as well as the restrictions inherent to observations made using two-dimensional core surfaces.
The amount of bioturbation was quantified considering the average diameter of 4 cm for the cores and using the bioturbation scale (BS) proposed by Reineck55, which ranges from 0 (non-bioturbated) to 6 (completely bioturbated). Data from the Rio da Batateira outcrop8,33 was taken into consideration for the creation of the stratigraphic section, due to its geographical proximity and relevant correlation to the stratigraphic sequence.
Calcareous microfossils
Samples were prepared for the recovery of both ostracods and foraminifera using the methodology developed for the rocks of the Romualdo Formation used by Bom et al.49, and consisted of the immersion of 20 g of sediment in 200 mL of deionized water with 3 mL of Extran for 24 h. The sediments were then washed through 250, 180, 63, and 45 µm sieves, and dried for 48 h in a lab oven at 40 ºC. After hand picking, we imaged the most representative specimens in an EVO/MA15 Zeiss scanning electron microscope (SEM).
No planktonic foraminifera species were recovered in washed samples, probably due to the strong cementation and poor preservation of the material in the sampled interval. Thus, their analysis was only possible through the thin sections made for microbiofacies studies.
All the studied material is currently stored in the micropaleontologic collection of the Museu de História Geológica do Rio Grande do Sul (MHGRS), Unisinos University, Brazil, under the curatorial numbers ULVG 13482 to ULVG 13491.
Organic-walled microfossils
We processed approximately 40 g of each sediment sample for palynology analysis, following standard techniques56. Minimums of 200 palynomorphs were counted in each sample for the palynological method. Species scanning, identification and counting were carried out with a Zeiss Imager.A2 microscope, using bright field illumination and incident blue light (fluorescence mode) at 200× , 400× , and 1000× (oil) magnifications. Photomicrographs were taken using a Zeiss AxioCam MRc (Micropaleontology Reference Center) digital camera. Our palynological analysis recorded pollen grains, spores, dinocysts, and freshwater green algae. The slides are stored in the collection of the MHGRS, Unisinos University, Brazil, under the curatorial numbers ULVG 13595 to ULVG 13682.
Calcareous nannofossils
Sample preparation for calcareous nannofossil analysis followed the decantation methodology described in Bown and Young57. Each sediment sample was fragmented in an agate mortar and placed in a Falcon tube with 40 mL of deionized water. The solution was stirred for 30 s and then set to decant for five minutes. The supernatant (approximately 0.2 mL) was then collected, poured onto a coverslip, and placed in a hotplate (60 °C) to dry. After dried, the coverslip was placed on a slide with Norland optical adhesive 61 and cured under UV light.
The slides were examined using a Zeiss Axio Imager.A2 microscope, at 1000× magnification. Data was processed using the software Zen 3.0 (blue edition) for micrometric measurements. The slides are stored in the collection of the MHGRS, Unisinos University, Brazil, under the curatorial numbers ULVG 13492 to ULVG 13594.
Microbiofacies
Lithology and preservation degree throughout the cored sections controlled the sampling interval. Select intervals of alternated mudstone-packstone and shale-siltstone required higher sampling density, while coarser-grained intervals were strategically undersampled.
We used a ZEISS Axioscope 5 petrographic microscope for microfossil identification and lithologic analysis, with transmitted and polarized light, objectives of 2.5× , 5× , 10× , and 20× magnification, and an attached digital camera. Data was processed using the software Zen 3.0 (blue edition) for micrometric measurements.
Calcareous rocks were described according to the nomenclatures of Dunham58. The taxonomic classification of planktonic foraminifera used was based on Verga and Premoli-Silva35,37, as well as the online Mesozoic Planktonic Foraminifera database located at https://www.mikrotax.org (pforams@mikrotax36). Planktonic foraminifera zones were defined based on the biostratigraphic schemes of Ogg1. We defined six semi-quantitative categories representing relative abundance, based on the number of specimens of foraminifera counted: very abundant (> 40), abundant (20–40), common (10–20), few (5–10), rare (3–5), and very rare (1–3) (SM). The slides are stored in the collection of the MHGRS, Unisinos University, under the curatorial numbers ULVG 13685 to ULVG 13722.
Data availability
The authors confirm that the data supporting the findings of this study are available within the article and its supplementary materials.
References
Ogg, J. G., Ogg, G. M. & Gradstein, F. M. Cretaceous. in A Concise Geologic Time Scale. 167–186. https://doi.org/10.1016/b978-0-444-59467-9.00013-3 (Elsevier, 2016).
Leckie, R. M., Bralower, T. J. & Cashman, R. Oceanic anoxic events and plankton evolution: Biotic response to tectonic forcing during the mid-Cretaceous. Paleoceanography 17, 13-1-13–29 (2002).
Hay, W. W. Evolving ideas about the Cretaceous climate and ocean circulation. Cretac. Res. 29, 725–753 (2008).
Moulin, M., Aslanian, D. & Unternehr, P. A new starting point for the South and Equatorial Atlantic Ocean. Earth Sci. Rev. 98, 1–37 (2010).
Channell, J. E. T., Erba, E., Nakanishi, M. & Tamaki, K. Late Jurassic-Early Cretaceous time scales and oceanic magnetic anomaly block models. in Geochronology, Time Scales and Global Stratigraphic Correlation. https://doi.org/10.2110/pec.95.04.0051 (SEPM Society for Sedimentary Geology, 1995).
Ogg, J. G., Hinnov, L. A. & Huang, C. Cretaceous. in The Geologic Time Scale 2012. 793–853. https://doi.org/10.1016/B978-0-444-59425-9.00027-5 (Elsevier B.V., 2012).
Arai, M. Aptian/Albian (Early Cretaceous) paleogeography of the South Atlantic: A paleontological perspective. Braz. J. Geol. 44, 339–350. https://doi.org/10.5327/Z2317-4889201400020012 (2014).
Assine, M. L. et al. Sequências deposicionais do Andar Alagoas da Bacia do Araripe, Nordeste do Brasil. Boletim Geoci. Petrobras 22, 3–28 (2014).
Luft-Souza, F. et al. Sergipe-Alagoas Basin, Northeast Brazil: A reference basin for studies on the early history of the South Atlantic Ocean. Earth-Sci. Rev. https://doi.org/10.1016/J.EARSCIREV.2022.104034 (2022).
Assine, M. L. Paleocorrentes e paleogeografia na Bacia do Araripe, Nordeste do Brasil. Rev. Brasil. Geoci. 24, 223–232 (1994).
Assine, M. L. Bacia do Araripe. Boletim Geoci. Petrobras 15, 371–389 (2007).
Ponte, F. C. & Appi, C. J. Proposta de revisão da coluna litoestratigráfica da Bacia do Araripe. in 36° Congresso Brasileiro de Geologia. Vol. 36. 211–226 (1990).
Brito Neves, B. B., Santos, E. J. & van Schmus, W. R. Tectonic history of the Borborema Province. in Tectonic Evolution of the South America. 31st International Geological Congress (eds. Cordani, U. G., Milani, E. J., Thomaz Filho, A. & Campos, D. A.). 151–182 (IUGS, 2000).
Fambrini, G. L. et al. Caracterização dos Sistemas Deposicionais da Formação Barbalha, Bacia do Araripe, Nordeste do Brasil. Vol. 103. http://www.lneg.pt/iedt/unidades/16/paginas/26/30/209 (2016).
Coimbra, J. C., Arai, M. & Carreño, A. L. Biostratigraphy of Lower Cretaceous microfossils from the Araripe Basin, northeastern Brazil. Geobios 35, 687–698 (2002).
Martill, D. M. The age of the Cretaceous Santana Formation fossil Konservat Lagerstätte of north–east Brazil: A historical review and an appraisal of the biochronostratigraphic utility of its palaeobiota. Cretac. Res. 28, 895–920 (2007).
Tomé, M. E. T. R., Lima Filho, M. F. & Neumann, V. H. M. L. Taxonomic studies of non-marine ostracods in the Lower Cretaceous (Aptian-lower Albian) of post-rift sequence from Jatobá and Araripe basins (Northeast Brazil): Stratigraphic implications. Cretaceous Res. 48, 153–176. https://doi.org/10.1016/j.cretres.2013.12.007 (2014).
Arai, M. & Assine, M. L. Chronostratigraphic constraints and paleoenvironmental interpretation of the Romualdo Formation (Santana Group, Araripe Basin, Northeastern Brazil) based on palynology. Cretac. Res. 116, 104610 (2020).
Melo, R. M. et al. New marine data and age accuracy of the Romualdo Formation, Araripe Basin, Brazil. Sci. Rep. 10, 10 (2020).
Araripe, R. C. et al. Upper Aptian–Lower Albian of the southern-central Araripe Basin, Brazil: Microbiostratigraphic and paleoecological inferences. J. South Am. Earth Sci. https://doi.org/10.1016/J.JSAMES.2022.103814 (2022).
Assine, M. L. Sedimentação e Tectônica da Bacia do Araripe, Nordeste do Brasil. in Dissertation (MSc). 1–124. (1990).
Neumann, V. H. & Assine, M. L. Stratigraphical propose to the post-rift-I tectonic-sedimentary sequence of Araripe Basin, northeastern Brazil. in 2° International Congress on Stratigraphy. 274–274 (2015).
Benigno, A. P. A., Saraiva, A. Á. F., Sial, A. N. & Lacerda, L. D. Mercury chemostratigraphy as a proxy of volcanic-driven environmental changes in the Aptian-Albian transition, Araripe Basin, northeastern Brazil. J. S. Am. Earth Sci. 107, 103020 (2021).
Lúcio, T., Souza Neto, J. A. & Selby, D. Late Barremian/Early Aptian Re–Os age of the Ipubi Formation black shales: Stratigraphic and paleoenvironmental implications for Araripe Basin, northeastern Brazil. J. S. Am. Earth Sci. 102, 102699 (2020).
Barreto, A. M. F. et al. U/Pb geochronology of fossil fish dentine from Romualdo Formation, Araripe Basin, northeast of Brazil. J. S. Am. Earth Sci. 116, 103774 (2022).
Assine, M. L. Análise estratigráfica da Bacia do Araripe, Nordeste do Brasil. Rev. Brasil. Geoci. 22, 289–300 (1992).
Fambrini, G. L. et al. Análise tectonossedimentar das fases início de rifte e clímax de rifte da Bacia do Araripe, Nordeste do Brasil. Geol. USP. Sér. Científ. 19, 205–236 (2019).
Chagas, D.B. Litoestratigrafia da Bacia do Araripe: Reavaliação e propostas para revisão. in Dissertation (MSc). 1–112. http://hdl.handle.net/11449/92893 (2006).
Varejão, F. G. et al. Exceptional preservation of soft tissues by microbial entombment: Insights into the taphonomy of the Crato Konservat-Lagerstätte. Palaios 34, 331–348 (2019).
Custódio, M. A. et al. The transgressive-regressive cycle of the Romualdo Formation (Araripe Basin): Sedimentary archive of the Early Cretaceous marine ingression in the interior of Northeast Brazil. Sed. Geol. 359, 1–15 (2017).
Maisey, J. G. Santana Fossils—An Illustrated Atlas. (Contributions to IUGS-IGCP Project 242, The Cretaceous of South America, 1991).
Goldberg, K., Premaor, E., Bardola, T. & Souza, P. A. Aptian marine ingression in the Araripe Basin: Implications for paleogeographic reconstruction and evaporite accumulation. Mar. Pet. Geol. 107, 214–221 (2019).
Araripe, R. C., Oliveira, D. H., Tomé, M. E., Moura de Mello, R. & Barreto, A. M. F. Foraminifera and Ostracoda from the Lower Cretaceous (Aptian–lower Albian) Romualdo Formation, Araripe Basin, northeast Brazil: Paleoenvironmental inferences. Cretaceous Res. 122, 104766 (2021).
Varejão, F. G. et al. Mixed siliciclastic-carbonate sedimentation in an evolving epicontinental sea: Aptian record of marginal marine settings in the interior basins of north-eastern Brazil. Sedimentology 68, 2125–2164 (2021).
Verga, D. & Premoli-Silva, I. Early Cretaceous planktonic foraminifera from the Tethys: The genus Leupoldina. Cretac. Res. 23, 189–212 (2002).
Huber, B. T. et al. Pforams@microtax: A new online taxonomic database for planktonic foraminifera. Micropaleontology 62, 429–438 (2016).
Verga, D. & Premoli-Silva, I. Early Cretaceous planktonic foraminifera from the Tethys: The large, many-chambered representatives of the genus Globigerinelloides. Cretac. Res. 24, 661–690 (2003).
Rios-Netto, A. D. M., Regali, M. D. S. P., Carvalho, I. D. S. & de Freitas, F. I. Palinoestratigrafia do intervalo Alagoas da Bacia do Araripe, Nordeste do Brasil. Rev. Brasil. Geoci. 42, 331–342 (2012).
Poropat, S. F. & Colin, J.-P. Early Cretaceous ostracod biostratigraphy of eastern Brazil and western Africa: An overview. Gondwana Res. 22, 772–798 (2012).
Regali, M. S. P. & Silva Santos, P. R. Palinoestratigrafia e geocronologia dos sedimentos Albo-Aptianos de Sergipe-Alagoas. in Boletim do 5° Simpósio Sobre o Cretáceo do Brasil. 411–419 (UNESP-Campus de Rio Claro/SP, 1999).
Santos, A. et al. Earlier onset of the Early Cretaceous Equatorial humidity belt. Global Planet. Change 208, 103724 (2022).
Scherer, C. M. S., Goldberg, K. & Bardola, T. Facies architecture and sequence stratigraphy of an early post-rift fluvial succession, Aptian Barbalha Formation, Araripe Basin, northeastern Brazil. Sed. Geol. 322, 43–62 (2015).
Hashimoto, A. T., Appi, C. J., Soldan, A. L. & Cerqueira, J. R. O Neo-Alagoas nas bacias do Ceará, Araripe e Potiguar (Brasil): Caracterização estratigráfica e paleoambiental. Rev. Brasil. Geoci. 17, 118–122 (1987).
Chagas, D. B., Assine, M. L. & Freitas, F. I. Facies sedimentares e ambientes deposicionais da Formação Barbalha no Vale do Cariri, Bacia do Araripe, Nordeste do Brasil. Geoci. UNESP 26, 313–322 (2007).
Uchman, A. An opportunistic trace fossil assemblage from the flysch of the Inoceramian beds (Campanian-Palaeocene), Bystrica Zone of the Magura Nappe, Carpathians, Poland. Cretaceous Res. 13, 539–547 (1992).
Buatois, L. A. & Mángano, M. G. Ichnology: Organism–Substrate Interactions in Space and Time (Cambridge University Press, 2011).
Krömmelbein, K. & Weber, R. Ostracoden des Nordost-Brasilianischen Wealden. Beihefte Geol. Jahrbuch 115, 1–93 (1971).
Whatley, R. C. Population structure of ostracods: some general principles for the recognition of palaeoenvironments. in Ostracoda in the Earth Sciences (eds. De Deckker, P., Colin, J. P., Peypouquet, J. P. & Townsend, A.). 231–244 (Elsevier, 1988).
Bom, M. H. H. et al. Paleoenvironmental evolution of the Aptian Romualdo Formation, Araripe Basin, Northeastern Brazil. Glob. Planet. Change 203, 103528 (2021).
Buatois, L. A. et al. Colonization of brackish-water systems through time: Evidence from the trace-fossil record. Palaios 20, 321–347 (2005).
Smith, A. B. & Crimes, P. T. Trace fossils formed by heart urchins—A study of Scolicia and related traces. Lethaia 16, 79–92 (1983).
Díez-Canseco, D., Buatois, L. A., Mángano, M. G., Rodriguez, W. & Solorzano, E. The ichnology of the fluvial–tidal transition: Interplay of ecologic and evolutionary controls. in Developments in Sedimentology. Vol. 68. 283–321 (Elsevier, 2015).
Scheid, C., Munis, M. B. & Paulino, J. Projeto Santana—Relatório Final Etapa II. Recife: CPRM (1978).
Bertling, M. et al. Names for trace fossils: A uniform approach. Lethaia 39, 265–286 (2006).
Reineck, H. E. Sedimentgefüge im Bereich der Südliche Nordsee Vol. 505 (Abhandlungen der Senckenbergischen Naturforschenden Gesellschaft, 1963).
Wood, G. D. Palynological techniques-processing and microscopy. in Palynology: Principles and Application (eds. Jasonius, J. & McGregor, D. C.) Vol. 1. 29–50 (American Association of Stratigraphic Palynologists Foundation, 1996).
Bown, P. & Young, J. R. Calcareous Nannofossil Biostratigraphy (Chapman and Hall/Kluwer Academic, 1998).
Dunham, R. J. Classification of Carbonate Rocks According to Depositional Textures (AAPG, 1962).
Acknowledgements
The authors acknowledge the research and development project entitled “Mar Interior: Incursões marinhas e a bioestratigrafia do Cretáceo Inferior nas Bacias Interiores do Nordeste do Brasil,” sponsored by Petrobras S.A., for funding this study; the National Mining Agency (Agência Nacional de Mineração-ANM/Brazil) for providing the studied samples; and itt Oceaneon/Unisinos for providing the facilities during the preparation, processing, and analytical steps. We are also grateful to Daniela Reháková, Gerson José Salamoni Terra, Guilherme Krahl, and Renata Guimarães Netto for their helpful taxonomic and paleoecological discussions; and to Victória Herder Sander, Bruna Poatskievick Pierezan, Geovani Goulart Freitas, Lucas Vinicius Oliveira, Luciana Rigon Carneiro Duarte, and Valeska Meirelles, for all laboratory support during sample preparation and processing techniques. GF was sponsored by Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq/Brazil: grant number 308087/2019-4), and MLA is a research fellow of CNPq (grant number 310955/2021-1). Finally, we’d like to thank the two anonymous reviewers for their contributions to the manuscript.
Author information
Authors and Affiliations
Contributions
G.F, F.H.O.L., M.L.A., and O.S.J. designed the project. H.P.K., A.S.C., and J.V.M. conducted the stratigraphical, sedimentological, and ichnological studies. M.A.L.M., A.S., and L.M.L. performed the descriptive research of palynomorphs. M.A.B.S.F. and D.C. performed the descriptive research of ostracods. F.L.S. and O.S.J. performed the descriptive research of microbiofacies and foraminifera. A.N.A., E.J.F.T., and M.D.R.B. performed the descriptive research of calcareous nannofossils. S.B.F. and M.H.H.B. gave a critical review to the work. All authors reviewed the manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Fauth, G., Kern, H.P., Villegas-Martín, J. et al. Early Aptian marine incursions in the interior of northeastern Brazil following the Gondwana breakup. Sci Rep 13, 6728 (2023). https://doi.org/10.1038/s41598-023-32967-w
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-023-32967-w
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
