
Abstract
Understanding how weather conditions affect animal populations is essential to foresee population changes in times of global climate shifts. However, assessing year-round weather impacts on demographic parameters is hampered in migratory animals due to often unknown occurrence in space and time. We addressed this by coupling tracking and weather data to explain extensive variation in apparent survival across 19 years in a northern European population of little ringed plovers (Charadrius dubius). Over 90% (n = 21) of tracked individuals followed migration routes along the Indo-European flyway to south India. Building on capture–recapture histories of nearly 1400 individuals, we found that between-year variation in precipitation during post-breeding staging in northern South Asia explained 47% of variation in apparent adult survival. Overall, the intensity of the monsoon in South Asia explained 31–33% of variability in apparent survival. In contrast, weather conditions in breeding, final non-breeding and pre-breeding quarters appeared less important in this species. The integration of multi-source data seems essential for identifying key regions and periods limiting population growth, for forecasting future changes and targeting conservation efforts.
Similar content being viewed by others

Introduction
Weather conditions alter the fitness of animals1,2,3,4 hence between-year variation in weather conditions affect demographic rates of populations5,6,7,8. Weather conditions affect populations both during the reproductive period3,9 and the non-reproductive period6,7,10,11,12. Thus, a full annual perspective on these relationships is required to thoroughly understand population fluctuations13,14,15, especially, under current global climate shifts16,17,18.
Assessments of year-round relationships between weather conditions and migratory bird populations were long hampered by challenges to track movements of migratory birds19,20,21 limiting integration with weather datasets. These challenges have been overcome with recent advent in bio-logging that enable tracking of full annual cycles of migratory birds22. Combining population-specific tracking data, weather conditions experienced throughout the annual cycle, and long-term data on survival could help to understand the mechanisms behind the population changes in migratory species23. Despite the need for such knowledge, the responses of populations to year-round weather conditions are unknown in many terrestrial birds8,12,24.
Here, we use full annual tracking to acquire detailed weather conditions experienced by little ringed plovers (Charadrius dubius; Fig. 1) from a north European breeding population and link these conditions with demographic data. The little ringed plover is a small, long-distance migratory shorebird breeding across the Palearctic25. The European breeding populations can use both the Afro-Palearctic and the Indo-European migratory flyways26,27 but the population-specific migratory patterns are mostly unknown. Importantly, apparent adult survival varies largely between years in a north European little ringed plover population, but the source of variation remains unknown28. Building on the current knowledge of migratory patterns in the species and relationships between weather conditions and migratory populations, we predict the following:
-
1.
Population-specific tracking data will enable detailed assessment of the weather–apparent survival relationships despite highly variable and currently largely unknown migratory patterns in the little ringed plover26,27.
-
2.
Weather conditions (precipitation and temperature) will affect apparent survival of the little ringed plovers because they affect the availability of ephemeral wetland habitats and food abundance29,30.
-
3.
Weather conditions outside breeding grounds will play a crucial role for the apparent survival of the little ringed plover breeding in northern Europe31,32.

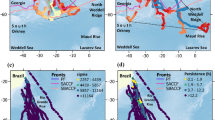
Distribution range of the little ringed plover (Charadrius dubius; light-coloured polygons27) and the main regions (kernel density estimates; polygons) where tracked individuals (n = 21; dots and triangles) occurred during the main annual cycle periods (Table 1). Coordinates of the geographic positions (dots and triangles) represent modus of daily positions derived from geolocators. Lines connect geographic positions and do not reflect migratory routes.
Results
Migratory pattern
Majority (19 out of 21) of the tracked little ringed plovers followed the Indo-European migratory flyway from Northern Europe to South Asia (Fig. 1). The migratory patterns were similar in these individuals that spent 53 ± 29 days (mean ± SD, n = 19) in the post-breeding staging region in northern South Asia, 164 ± 30 days (n = 15) in the non-breeding region in south India, and 16 ± 6 days (n = 15) in the pre-breeding staging region in northern South Asia (Fig. 1, Table 1). Beside these 19 individuals, one individual spent the main non-breeding period (236 days) close to the Caspian Sea without longer stopovers (> 6 days) during migration. A second individual followed the Afro-Palearctic migratory flyway through a post-breeding staging site in central Middle East (77 days) to East Africa (151 days) with a short stop (< 6 days) close to the Caspian Sea during northward migration (Fig. 1). We detected no statistically significant differences in the duration of the main annual cycle periods between females and males: the average differences (males–females) for post-, non- and pre-breeding periods were 0.5, 1.1 and − 0.6 days, respectively (Mann–Whitney non-parametric test P values 0.90, 0.68 and 0.95).
Between-year variation in weather conditions
The amount of precipitation varied largely between years in the four key regions visited by the tracked birds (coefficients of variation [CV] = 0.23–0.69, n = 18 for each region) and the temporal pattern differed between these regions (absolute Spearman correlation coefficients [|ρ|] ≤ 0.35; Fig. 2A). The pattern of between-year changes in near-surface mean air temperature differed between regions (|ρ|≤ 0.47), and the variation was extensive only in the breeding region (CV = 0.52; mean = 3 °C) and showed low variation outside breeding region with CV < 0.05 and mean > 20 °C (Fig. 2B). In contrast, the patterns of between-year changes in precipitation during the monsoon period showed strong correlation (|ρ|= 0.57–0.95) and high variation in the three non-breeding regions (CV = 0.18–0.46; Fig. 2C).

Between-year variation in amount of precipitation (A) and near-surface mean air temperature (B) during four main annual cycle periods of little ringed plovers (Charadrius dubius). (C) Between-year variation in the amount of precipitation during monsoon period (July–September; relative values presented). (D) Between-year variation in apparent survival estimates of adult little ringed plovers (mean ± SE) estimated from the time dependent model (Table 2).
Predictors of the apparent survival fluctuation
Apparent survival of the little ringed plovers varied substantially between years (ɸ = 0.35–0.85; ΔAICc = 6.04 compared to constant model, Table 2; Fig. 2D) and males exhibited higher survival than females, as previously found in 28 (Fig. 2D; Table 2). High temporal variation in annual survival was best explained (47% explained variation) by a positive relationship with amount of precipitation in the post-breeding staging region in northern South Asia (ΔAICc = 17.14 compared to constant model; Fig. 3; Table 2). Overall, the amount of precipitation during the monsoon period (July–September) in the staging regions (pre-breeding: ΔAICc = 11.17 and post-breeding ΔAICc = 10.76) and in the non-breeding region (ΔAICc = 10.46) received less support compared to the best model but remained more supported than the constant model and explained 31–33% of the variation in apparent survival (Table 2). Near-surface mean air temperature in the post-breeding and non-breeding regions was only weakly linked to survival and explained 8% and 11% of temporal variability in apparent survival (ΔAICc = 1.15 and 2.32; Table 2). Similarly, the amount of precipitation during the non-breeding, pre-breeding and breeding periods explained only little variation in apparent survival, as was the case also for near-surface mean air temperature in the pre-breeding and breeding regions (Table 2).

Apparent survival (with 95% confidence intervals) of little ringed plovers (Charadrius dubius) in relation to precipitation in the post-breeding staging region in South Asia (model ‘Precipitation post-breeding’ from Table 2). The annual estimates from the time dependent model are shown as dots.
Discussion
We show that the fluctuation in adult survival of a long-distance migratory species is associated with cross-continental weather conditions. By tracking, we uncovered the spatiotemporal distribution of the little ringed plover breeding in northern Europe, which was essential to narrow down the regions and periods of importance. By combining this information with weather and long-term breeding capture-recapture data, we pinpointed the weather conditions experienced throughout the annual cycle that are linked to high temporal variation in apparent survival. In particular, the amount of precipitation during the post-breeding staging period explained 47% of the temporal variation in survival. Our results show that the conditions during migratory periods and staging are important and have repercussions for individuals33,34 and illustrate the value of a full-annual perspective in such studies to provide essential knowledge on cross-continental relationships between weather conditions and survival rates.
The apparent survival of little ringed plovers breeding in northern Europe showed a strong positive link to precipitation during multiple non-breeding periods in South Asia. These findings are consistent with previous findings on positive relationships between survival and precipitation during the staging and other non-breeding periods10,34,35. Little ringed plovers migrated from Finnish breeding grounds to South Asian non-breeding quarters by crossing the vast Palearctic landmasses. Therefore, rains during the post-breeding period largely coinciding with the Asian summer monsoon form the only major climatic event they experience, which may explain the strong link between this event and their annual survival.
While multiple studies have shown negative impacts of low temperatures on survival of short-distance migratory shorebirds36,37,38, our study is among the first to show an effect of precipitation on adult survival in a long-distance migratory shorebird. Previously, a link between adult survival of common sandpipers (Actitis hypoleuca) and the North Atlantic Oscillation was detected39, indicating that warm and wet conditions in the wintering areas result in higher survival but that precipitation has no effect on survival. However, we detected only very little between-year variation in temperature measures at the non-breeding sites far below the diurnal and annual changes experienced by the birds. Our results are consistent with studies suggesting that long-distance migratory shorebirds mainly incur mortality at the non-breeding sites31,40. The lack of previous studies reporting a relationship between precipitation and survival of long-distance migratory shorebirds may reflect the fact that shorebirds generally exhibit low levels of temporal variation in adult survival and high survival compared to other similar-sized birds41,42,43. Interestingly, plovers (genus Charadrius) have lower survival than expected from their body size43 and exhibit substantial annual variation in survival (e.g.,44 and this study). Pronounced annual variation in survival suggests that they are more vulnerable to environmental conditions than other shorebird species.
Susceptibility to environmental conditions may be linked to variation in the quality of non-breeding habitats. Similar to other long-distance shorebirds that migrate across continents and opportunistically stop at suitable sites45, the little ringed plovers are known to use staging sites such as sandbanks, rivers and lakes, rice fields, residual flood waters, short-vegetated areas near villages or water, airfields, and pastures30,46. Some of these habitats are highly dependent on water level and may thus make little ringed plovers susceptible to drying of the foraging habitats. Dry conditions affect the presence of water bodies, which reduces availability of invertebrate food in the staging areas47. Hence, drying of the staging habitats may have severe consequences for refueling performance, body condition, success of migration48,49,50, and ultimately survival, as documented here.
Precipitation during the post-breeding staging was more important than during the pre-breeding and non-breeding periods. Dry conditions during post-breeding staging may be more critical for survival because post-breeding staging occurs after an energetically-demanding migratory period51 and is accompanied by the moult of flight feathers that occurs mainly after August52. Importantly, it appears that there is a relatively strong impact of the summer monsoon (but see model supports) extending to the entire non-breeding period of the little ringed plovers and potentially affecting their survival throughout this period of the annual cycle. As the magnitude and direction of climatic changes differ between regions53, our results can be used to assess potential future impacts because the amount of precipitation is predicted to increase in South Asia54. This may benefit some long-distance migrants such as the little ringed plover via increased survival. However, marked temporal variation in weather conditions may also increase variation in survival potentially translating to population growth rates and consequently depress population size in the long-term (e.g.,8).
Most of the tracked little ringed plovers followed the Indo-European migratory flyway that has been previously described mainly for passerines (see55,56,57). Such a low spread within the non-breeding grounds was unexpected as European ringing recoveries show rather large variation in non-breeding directions and sites27,58,59,60 and contrasts with the Swedish little ringed plovers that cover almost an entire nonbreeding range of the species26. A potential limitation to our results is a spatiotemporal mismatch between survival and tracking datasets. However, we do not expect extensive mixing of the studied population with a population from southern Sweden with highly variable migratory patterns26 because of short natal and breeding dispersal distances28, and the presence of the Bothnian Bay likely forming a barrier between these populations. This explanation is also indirectly supported by the ring recoveries59.
In our study, the amount of precipitation explained up to 47% of the temporal variation in apparent survival of adults, more than reported in previous studies on terrestrial birds (< 18%; e.g.,15,61,62). Such a strong association likely reflects a precise match between the tracking and weather datasets advocating and further supporting our assumption that the breeding population largely follows the migration schedule and pattern of the tracked individuals.
Weather conditions in the staging region during post-breeding migration appears to limit survival in the population of this migratory shorebird and considerable amount of explained variation suggests susceptibility of the population to the weather conditions in this short period of the annual cycle. Our results pinpointed the region limiting the population for which the climatic models forecast long-term increase in the amount of precipitation that might prevent population declines in the species. We believe that long-term studies of populations and integration of their migratory patterns with environmental conditions will become a crucial step for better understanding large-scale population size dynamics, forecasting future changes and targeting conservation efforts in migratory birds.
Material and methods
We studied a north European breeding population of little ringed plovers to assess the potential impacts of between-year changes in weather conditions on adult survival. First, we gathered tracking data from two ‘tracking’ areas (62° 36′ N, 29° 48′ E and 65° 00′ N, 25° 30′ E) to identify migratory pattern in the north European population (Fig. 1). Second, we estimated apparent survival of adult individuals from data collected in a ‘ringing’ area (60° 17′ N, 25° 6′ E; Fig. 1).
Tagging and individual spatiotemporal distribution
We tagged 91 breeding adults (39 females, 43 males, 9 unidentified) in two tracking areas during 2015 and 2016 with light-level geolocators to determine their spatiotemporal distribution throughout the annual cycle. The mean body mass of tagged individuals was 38.3 ± 2.4 g (SD; n = 74), corresponding to relative tag load of 2.3% (mean; range 1.8; 3.3%), and we assume negligible effects of tagging on obtained migratory behaviour63,64. In the years following tag deployment, we recovered tags with at least 6 months of ambient light recording from 21 individuals (Electronic Supplementary Material 1).
We determined the geographic locations outside the breeding areas and the timing of annual cycle events using the ambient light intensity recordings from the geolocators. We estimated times of sunrises and sunsets from the log-transformed data (twGeos R package)65; and for further analysis used functions from GeoLight package version 2.0.066. We identified stationary periods (changeLight function; quantile = 0.9; days = 2) and calculated daily geographic positions using sun elevation angles (SEA) derived by Hill-Ekstrom calibration on the longest non-breeding stationary period67. We merged consecutive periods with overlapping locations and applied the Hill-Ekstrom calibration to derive final SEAs (mean = −6.7°; SD = 1.3, n = 21). In two individuals, the final SEA resulted in locations in the ocean, and we thus adjusted the SEAs (by −0.5° and −0.9°) to move the locations to the nearest land.
Population spatiotemporal distribution
Out of 21 tracked individuals, 19 spent the non-breeding period in southern India (Fig. 1). Only one individual spent the non-breeding period in Iran and one individual in Africa (Fig. 1) preventing us to robustly specify their space and time use. Hence, we considered the spatiotemporal distribution of 19 individuals following migratory routes to South Asia as the prevalent migratory strategy in the studied population (more details in “Discussion”).
To describe the population-specific space use outside the breeding areas, we extracted individual timing and daily position estimates during the following main periods: (1) the longest stationary period during southward migration (hereafter ‘post-breeding staging’), (2) the stationary period when the individual was farthest from the tracking areas (hereafter ‘non-breeding’), and (3) the longest stationary period during the northward migration (hereafter ‘pre-breeding staging’). The individual migration schedules are presented in Electronic Supplementary Material 2. We calculated the population timing of migration as the average of the first (arrivals) and the last (departures) days of the main periods (Table 1). In addition, we considered ‘breeding period’ (1–30 April) as the period around breeding site arrival in the tracked birds (mean = 18 April; n = 14). Sample sizes for timing estimates differ between main periods due to premature battery failures in some geolocators (Table 1). The breeding region is defined as the area surrounding the ringing area (described below; 60° 08′–60° 35′ N; 24° 49′–25° 49′E). We estimated the population space use outside the breeding areas during the main periods (hereafter ‘main regions’ together with the breeding region) from bi-daily position estimates using kernel density estimates. We applied kde.points function (GISTools R package)68 with a bandwidth of 10, and omitted the lowest 2% of density estimates and clipped out areas over seawater (Fig. 1).
Weather conditions during the annual cycle
In total, we extracted four precipitation and four near-surface mean air temperature measures matching spatiotemporal distribution, and three monsoon measures matching only spatial distribution. Weather variables were extracted from TerraClimate69 using the spatiotemporal distribution of the tracked population (Fig. 1; Table 1) by calculating weighted average of monthly values using the number of days in each month as a weight. We obtained these measures for seasons preceding the breeding periods 1981–1998 (see below). In addition, we collected information on the amount of precipitation in three main regions outside breeding areas (Fig. 1) during summer monsoon period in July–September; a major climatic event in South Asia70. We used R version 4.1.2 for the analysis71 and Google Earth Engine to gather weather data72.
Capture–recapture and apparent survival analysis
We used capture–recapture histories of 1386 individuals (747 females, 639 males) collected in the ringing area (Fig. 1) to estimate apparent survival of the little ringed plover in 1980–1998. The dataset is detailed in28 and summarised in Electronic Supplementary Material 3. We employed Cormack-Jolly-Seber (CJS) models adapted for open populations73 in program MARK version 9.074. Our initial model included sex, time (t) and their interaction (*) for survival (Φ), and recapture probabilities (p) included also immediate trap-dependence (m; see75) on capture probabilities [Φ(sex*t) p(sex*m*t)]. We assessed goodness-of-fit with U-CARE 2.3.276 and found that this model fit the data28. In this study, we continue with the best-fit structure from28, i.e. [Φ(sex + t) p(sex + trend)] which includes sex and time for survival and trap response and temporal trend for recapture probabilities. In other models, we only varied the structure for time dependence in survival. In addition to the starting model in which survival was time dependent, we fit models where survival was constant in time or constrained by one of the 11 weather variables. Sex was included for survival in all models because females were found to have lower apparent survival than males in the previous study28. This sex difference in survival is likely caused by differences in permanent emigration as females disperse further than males28. This potential permanent emigration can reduce the apparent survival estimates. As breeding dispersal decisions are mostly linked to breeding success in shorebirds (e.g.,77) and the changing quality of the ephemeral breeding sites (mostly man-made habitats28) of the little ringed plover in this study population, it is not likely that permanent emigration would be linked to the weather conditions experienced in the non-breeding sites.
We compared models using the Akaike information criterion adjusted for small sample size (AICc)78 and considered a difference of at least 2 AICc units to infer a difference in model support. We assessed support for time dependence and weather variables by comparing those models to the constant model. In addition, we examined the percentage of temporal variation explained by the weather covariates by comparing deviances the weather covariate models to constant and time dependent models. We calculated the percentage of deviance explained by the covariate model following79 as
where Dev(c) is the deviance from the constant model, Dev(cov) is deviance from the covariate model and Dev(t) is the deviance from the time dependent model.
Ethical approval
This study complied with national law and the reporting in the manuscript follows the recommendations in the ARRIVE guidelines. The experimental protocols and methods were approved by the Centre for Economic Development, Transport and the Environment in Finland (permit number VARELY/1088/2015) and the Ringing Centre of the Finnish Museum of Natural History. All methods were carried out in accordance with relevant guidelines and regulations.
Data availability
Data are available from Zenodo data repository80 and MoveBank (ID 2279403362).
References
Marra, P., Hobson, K. A. & Holmes, R. T. Linking winter and summer events in a migratory bird by using stable-carbon isotopes. Science 282, 1884–1886 (1998).
Korslund, L. & Steen, H. Small rodent winter survival: Snow conditions limit access to food resources. J. Anim. Ecol. 75, 423–436 (2009).
Both, C., Bouwhuis, S., Lessells, C. M. & Visser, M. E. Climate change and population declines in a long-distance migratory bird. Nature 441, 81–83 (2006).
Rughetti, M. & Festa-Bianchet, M. Effects of spring–summer temperature on body mass of chamois. J. Mammal. 93, 1301–1307 (2012).
Davidson, J. & Andrewartha, H. The influence of rainfall, evaporation and atmospheric temperature on fluctuations in the size of a natural population of Thrips imaginis (Thysanoptera). J. Anim. Ecol. 17, 200–222 (1948).
Sillett, T. S., Holmes, R. T. & Sherry, T. W. Impacts of a global climate cycle on population dynamics of a migratory songbird. Science 288, 2040–2043 (2000).
SÆther, B. E., Sutherland, W. J. & Engen, S. Climate influences on avian population dynamics. Adv. Ecol. Res. 35, 185–209 (2004).
Frederiksen, M., Daunt, F., Harris, M. & Wanless, S. The demographic impact of extreme events: Stochastic weather drives survival and population dynamics in a long-lived seabird. J. Anim. Ecol. 77, 1020–1029 (2008).
Cox, A. R., Robertson, R. J., Rendell, W. B. & Bonier, F. Population decline in tree swallows (Tachycineta bicolor) linked to climate change and inclement weather on the breeding ground. Oecologia 192, 713–722 (2020).
Peach, W., Baillie, S. & Underhill, L. Survival of British Sedge Warblers in relation to west African rainfall. Ibis 133, 300–305 (1991).
Altwegg, R., Dummermuth, S., Anholt, B. R. & Flatt, T. Winter weather affects asp viper Vipera aspis population dynamics through susceptible juveniles. Oikos 110, 55–66 (2005).
Woodworth, B. K., Wheelwright, N. T., Newman, A. E., Schaub, M. & Norris, D. R. Winter temperatures limit population growth rate of a migratory songbird. Nat. Commun. 8, 14812 (2017).
Ådahl, E., Lundberg, P. & Jonzén, N. From climate change to population change: The need to consider annual life cycles. Glob. Change Biol. 12, 1627–1633 (2006).
Marra, P. P., Cohen, E. B., Loss, S. R., Rutter, J. E. & Tonra, C. M. A call for full annual cycle research in animal ecology. Biol. Lett. 11, 20150552 (2015).
Telenský, T., Klvaňa, P., Jelínek, M., Cepák, J. & Reif, J. The influence of climate variability on demographic rates of avian Afro-palearctic migrants. Sci. Rep. 10, 17592 (2020).
Dybala, K. E., Eadie, J. M., Gardali, T., Seavy, N. E. & Herzog, M. P. Projecting demographic responses to climate change: Adult and juvenile survival respond differently to direct and indirect effects of weather in a passerine population. Glob. Chang. Biol. 19, 2688–2697 (2013).
Gullett, P., Evans, K. L., Robinson, R. A. & Hatchwell, B. J. Climate change and annual survival in a temperate passerine: Partitioning seasonal effects and predicting future patterns. Oikos 123, 389–400 (2014).
Selwood, K. E., McGeoch, M. A. & Mac Nally, R. The effects of climate change and land-use change on demographic rates and population viability. Biol. Rev. 90, 837–853 (2015).
Bridge, E. S. et al. Technology on the move: Recent and forthcoming innovations for tracking migratory birds. Bioscience 61, 689–698 (2011).
van Bemmelen, R. S. A. et al. Red-necked phalaropes in the Western Palearctic reveals contrasting migration and wintering movement strategies. Front. Ecol. Evol. 7, 86 (2019).
Jiguet, F. et al. Unravelling migration connectivity reveals unsustainable hunting of the declining ortolan bunting. Sci. Adv. 5, eaau2642 (2019).
Stutchbury, B. J. M. et al. Tracking long-distance songbird migration by using geolocators. Science 323, 896 (2009).
Sanderson, F. J., Donald, P. F., Pain, D. J., Burfield, I. J. & Van Bommel, F. P. J. Long-term population declines in Afro-Palearctic migrant birds. Biol. Conserv. 131, 93–105 (2006).
Sandvik, H., Erikstad, K. E., Barrett, R. T. & Yoccoz, N. G. The effect of climate on adult survival in five species of North Atlantic seabirds. J. Anim. Ecol. 74, 817–831 (2005).
BirdLife International and NatureServe. Bird species distribution maps of the world. (2014).
Hedenström, A., Klaassen, R. H. G. & Åkesson, S. Migration of the little ringed plover Charadrius dubius breeding in South Sweden tracked by geolocators. Bird Study 60, 466–474 (2013).
Fransson, T., Österblom, H. & Hall-Karlsson, S. Svensk ringmärkningsatlas. (2008).
Pakanen, V., Lampila, S., Arppe, H. & Valkama, J. Estimating sex specific apparent survival and dispersal of Little Ringed Plovers (Charadrius dubius). Ornis Fenn. 92, 52 (2015).
Jarošík, V., Honěk, A., Magarey, R. & Skuhrovec, J. Developmental database for phenology models: Related insect and mite species have similar thermal requirements. J. Econ. Entomol. 104, 1870–1876 (2011).
Cramp, J. Handbook of the Birds of Europe, the Middle East and North Africa (Oxford University Press, 1992).
Leyrer, J. et al. Mortality within the annual cycle: Seasonal survival patterns in Afro-Siberian Red Knots Calidris canutus canutus. J. Ornithol. 154, 933–943 (2013).
Norris, R. D. & Marra, P. P. Seasonal interactions, habitat quality, an population dynamics in migratory birds. Condor 109, 535–547 (2007).
Schmaljohann, H., Eikenaar, C. & Sapir, N. Understanding the ecological and evolutionary function of stopover in migrating birds. Biol. Rev. 97, 1231–1252 (2022).
Doyle, S. et al. Temperature and precipitation at migratory grounds influence demographic trends of an Arctic-breeding bird. Glob. Change Biol. 26, 5447–5458 (2020).
Rockwell, S. M. et al. Seasonal survival estimation for a long-distance migratory bird and the influence of winter precipitation. Oecologia 183, 715–726 (2017).
Insley, H., Peach, W., Swann, B. & Etheridge, B. Survival rates of Redshank Tringa totanus wintering on the Moray Firth. Bird Study 44, 277–289 (1997).
Duriez, O., Ens, B. J., Choquet, R., Pradel, R. & Klaassen, M. Comparing the seasonal survival of resident and migratory oystercatchers: Carry-over effects of habitat quality and weather conditions. Oikos 121, 862–873 (2012).
Cook, A. S. C. P. et al. Temperature and density influence survival in a rapidly declining migratory shorebird. Biol. Conserv. 260, 109198 (2021).
Pearce-Higgins, J. W., Yalden, D., Dougall, T. & Beale, C. M. Does climate change explain the decline of a trans-Saharan Afro-Palaearctic migrant?. Oecologia 159, 649–659 (2009).
Weiser, E. L. et al. Environmental and ecological conditions at Arctic breeding sites have limited effects on true survival rates of adult shorebirds. Auk 135, 29–43 (2018).
Piersma, T. & Baker, A. Life history characteristics and the conservation of migratory shorebirds. In Behaviour and Conservation (eds Gosling, L. & Sutherland, W.) 105–124 (Cambridge University Press, 2000).
Conklin, J. R., Senner, N. R., Battley, P. F. & Piersma, T. Extreme migration and the individual quality spectrum. J. Avian Biol. 48, 19–36 (2017).
Méndez, V., Alves, J. A., Gill, J. A. & Gunnarsson, T. G. Patterns and processes in shorebird survival rates: A global review. Ibis (Lond.) 160, 723–741 (2018).
Roche, E. A. et al. Range-wide piping plover survival: Correlated patterns and temporal declines. J. Wildl. Manage. 74, 1784–1791 (2010).
Skagen, S. K. & Knopf, F. L. Toward conservation of midcontinental shorebird migrations. Conserv. Biol. 7, 533–541 (1993).
Kasahara, S., Moritomo, G., Kitamura, W., Imanishi, S. & Azuma, N. Rice fields along the East Asian-Australasian flyway are important habitats for an inland wader’s migration. Sci. Rep. 10, 4118 (2020).
Studds, C. E. & Marra, P. P. Linking fluctuations in rainfall to nonbreeding season performance in a long-distance migratory bird, Setophaga ruticilla. Clim. Res. 35, 115–122 (2007).
Newton, I. Can conditions experienced during migration limit the population levels of birds?. J. Ornithol. 147, 146–166 (2006).
Anderson, A. M. et al. Drought at a coastal wetland affects refuelling and migration strategies of shorebirds. Oecologia 197, 661–674 (2021).
Rakhimberdiev, E. et al. Fuelling conditions at staging sites can mitigate Arctic warming effects in a migratory bird. Nat. Commun. 9, 4263 (2018).
Wikelski, M. et al. Costs of migration in free-flying songbirds. Nature 423, 704 (2003).
Meissner, W. Ageing and sexing the curonicus subspecies of the Little Ringed Plover Charadrius dubius. Wader Study Gr. Bull. 113, 28–31 (2007).
Giorgi, F. Climate change hot-spots. Geophys. Res. Lett. 33, L08707 (2006).
Almazroui, M., Saeed, S., Saeed, F., Islam, M. N. & Ismail, M. Projections of precipitation and temperature over the South Asian countries in CMIP6. Earth Syst. Environ. 4, 297–320 (2020).
Lisovski, S. et al. The Indo-European flyway: Opportunities and constraints reflected by Common Rosefinches breeding across Europe. J. Biogeogr. 48, 1255–1266 (2021).
Lislevand, T. et al. Red-spotted Bluethroats Luscinia s. svecica migrate along the Indo-European flyway: A geolocator study. Bird Study 62, 508–515 (2015).
Brlík, V., Ilieva, M., Lisovski, S., Voigt, C. C. & Procházka, P. First insights into the migration route and migratory connectivity of the Paddyfield Warbler using geolocator tagging and stable isotope analysis. J. Ornithol. 159, 879–882 (2018).
Wernham, C. et al. The Migration Atlas: Movements of the Birds of Britain and Ireland (Poyser, 2002).
Saurola, P., Valkama, J. & Velmala, W. The Finnish Bird Ringing Atlas (Finnish Museum of Natural History and the Ministry of Environment, 2013).
Bairlein, F. et al. Atlas des Vogelzugs—Ringfunde Deutscher Brut- und Gastvögel (AULA-Verlag GmbH, 2014).
Salewski, V., Hochachka, W. M. & Fiedler, W. Multiple weather factors affect apparent survival of European Passerine birds. PLoS One 8, e59110 (2013).
Schaub, M., Jakober, H. & Stauber, W. Demographic response to environmental variation in breeding, stopover and non-breeding areas in a migratory passerine. Oecologia 167, 445–459 (2011).
Brlík, V. et al. Weak effects of geolocators on small birds: A meta-analysis controlled for phylogeny and publication bias. J. Anim. Ecol. 89, 207–220 (2020).
Weiser, E. L. et al. Effects of geolocators on hatching success, return rates, breeding movements, and change in body mass in 16 species of Arctic-breeding shorebirds. Mov. Ecol. 4, 12 (2016).
Lisovski, S., Sumner, M. D., & Wotherspoon, S. J. TwGeos: Basic data processing for light based geolocation archival tags. 2015. https://github.com/slisovski/TwGeos
Lisovski, S. & Hahn, S. GeoLight—processing and analysing light-based geolocator data in R. Methods Ecol. Evol. 3, 1055–1059 (2012).
Ekstrom, P. A. An advance in geolocation by light. Mem. Natl Inst. Polar Res. 58, 210–226 (2004).
Brunsdon, C. & Chen, H. GISTools: Some further GIS capabilities for R. (2014).
Abatzoglou, J. T., Dobrowski, S. Z., Parks, S. A. & Hegewisch, K. C. TerraClimate, a high-resolution global dataset of monthly climate and climatic water balance from 1958–2015. Sci. Data 5, 170191 (2018).
Wang, B. The Asian Monsoon (Springer, 2006).
R Core Team. A Language and Environment for Statistical Computing (2021).
Gorelick, N. et al. Google Earth Engine: Planetary-scale geospatial analysis for everyone. Remote Sens. Environ. 202, 18–27 (2017).
Lebreton, J., Burnham, K. P., Clobert, J. & Anderson, D. R. Modeling survival and testing biological hypotheses using marked animals: A unified approach with case studies. Ecol. Monogr. 62, 67–118 (1992).
White, G. C. & Burnham, K. P. Program MARK: Survival estimation from populations of marked animals. Bird Study 46, S120–S139 (1999).
Pradel, R. Flexibility in survival analysis from recapture data: Handling trap-dependence. In Marked Individuals in the Study of Bird Population (eds Lebreton, J.-D. & North, P.) (Birkhäuser-Verlag, 1993).
Choquet, R., Lebreton, J. D., Gimenez, O., Reboulet, A. M. & Pradel, R. U-CARE: Utilities for performing goodness of fit tests and manipulating CApture-REcapture data. Ecography (Cop.) 32, 1071–1074 (2009).
Pakanen, V. M. et al. Natal dispersal does not entail survival costs but is linked to breeding dispersal in a migratory shorebird, the southern dunlin Calidris alpina schinzii. Oikos 2022, ee08951 (2022).
Burnham, K. & Anderson, D. Model Selection and Multimodel Inference: A Practical in-Formation-Theoretic Approach (Springer, 2002).
Grosbois, V. et al. Assessing the impact of climate variation on survival in vertebrate populations. Biol. Rev. 83, 357–399 (2008).
Brlík, V. et al. Survival fluctuations linked to variation in the South Asian monsoon in a Palearctic migratory shorebird. Zenodo https://doi.org/10.5281/zenodo.7026440 (2022).
Acknowledgements
TJ. was supported by the Finnish Cultural foundation. V.B. was supported by the Czech Science Foundation (Grant no. 20-00648S) and D.B. by Stiftelsen Olle Engkvist Byggmästare. We thank two anonymous reviewers for their critical comments. We thank Biodiversity Anthropocenes for funding article processing charges.
Author information
Authors and Affiliations
Contributions
V.B. and V.-M.P. designed the study and wrote the manuscript; S.H. provided part of the tracking devices; H.A. collected capture-recapture data; T.J. and D.B. acquired funding; T.J., J.L., V.-M.P. and K.K. collected geolocator data; V.B. analysed the geolocator data and collected the weather data; V.-M.P. and J.J. analysed the capture-recapture data. All authors commented and approved the manuscript.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Brlík, V., Pakanen, VM., Jaakkonen, T. et al. Survival fluctuation is linked to precipitation variation during staging in a migratory shorebird. Sci Rep 12, 19830 (2022). https://doi.org/10.1038/s41598-022-24141-5
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-022-24141-5
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
