
Abstract
Based on new fossil materials, a new species Toxorhina (Ceratocheilus) christelius sp. nov. has been described herein with complete documentation of drawings and photographs. Features such as wide spine on the gonocoxite differentiating the new species of Toxorhina were discussed. Finding new interesting fossil materials also allowed for providing an emended diagnosis and additional description of known Eocene species—Toxorhina (Ceratocheilus) eridanus. Comparison of chosen morphological features of fossil and recent representatives of the genus were given and key for fossil species of subgenus Ceratocheilus was introduced. Distribution of recent Toxorhina and evolutionary history of the genus were discussed. The results of research on fossil materials prove that the stratigraphic range of the subgenus Ceratocheilus and the genus Toxorhina goes back to the Eocene, there is no evidence of their existence on Earth before. Moreover, these insects were probably associated with a warm climate, they were found for example in Baltic amber, the deposits of which were formed mainly in a subtropical climate. The reach diversity of the genus Toxorhina of recent fauna is strictly observed in tropical zones around the world.
Similar content being viewed by others



Introduction
The Limoniidae genus Toxorhina Loew is represented in recent fauna by over 150 species and subspecies within three subgenera: Ceratocheilus Wesché, Eutoxorhina Alexander, Toxorhina Loew. Subgenus Ceratocheilus is currently the most speciose (85 species, vast majority in Old World), subgenus Toxorhina comprises 68 species (nearly 1/3 in New World) and subgenus Eutoxorhina is represented only in Australian and Oceanian region by three species (Table 1; Supplementary Table 1). Fossil record of Toxorhina is scarce and limited only to three species placed in subgenus Ceratocheilus and one unplaced to any subgenus. The Toxorhina (Ceratocheilus) eridanus Meunier was described based on inclusions in Eocene Baltic amber, the second one—Toxorhina (Ceratocheilus) caucasiensis Krzemiński and Freiwald is known from imprints in sediments of Miocene from Stavropol (Stavropol’ Kraï, Russia). Toxorhina (Ceratocheilus) mexicana was described from Miocene Mexican amber. Another species—Toxorhina madagascariensis Meunier was described from Holocene defaunation resin of Madagascar–, but it remains unplaced to any subgenus of Toxorhina (Table 2). Comparative studies of new fossil materials allowed to establish and describe a new species for science and provide a hypothesis about Eocene and post-Eocene stages of evolution of this fossil taxa rare in fossil record.
Results
Systematic palaeontology
Order: Diptera Linnaeus
Infraorder: Tipulomorpha Rohdendorf
Family: Limoniidae Speiser
Subfamily: Limoniinae Speiser
Genus: Toxorhina Loew
Type species: Toxorhina fragilis Loew, by subsequent designation by Osten Sacken
Subgenus: Ceratocheilus Wesché
1910 Ceratocheilus Wesché, p. 358 (as genus)
Type species: Ceratocheilus winnsampsoni Wesché [= Styringomyia cornigera Speiser], by subsequent designation of Brunetti)
Toxorhina (Ceratocheilus) eridanus Meunier

Toxorhina (Ceratocheilus) eridanus Meunier5 (Limoniidae), inclusions in Baltic amber, (A–E, G, H)—No. CCHH 832-1 (male) (coll. Ch. and H. W. Hoffeins), specimen from Senckenberg Deutsches Entomologisches Institut (SDEI) Müncheberg, Germany: (A) body, dorso-ventral view; (B) enlarged view of the tip of rostrum with palpus visible; (C) last tarsomeres; (D) drawing of head with part of rostrum, latero-dorsal view (reconstruction); (E) photograph of head with part of rostrum, latero-dorsal view; (F)—No. K2532 (male) (coll. Klebs), specimen from University of Göttingen, drawing of wing (reconstruction); (G) photograph of enlarged view of sc-r position; (H) photograph of enlarged view of base of wing; (I) relation between the length of rostrum (r), antenna (a) and wing (w). p—palpus.

Toxorhina (Ceratocheilus) eridanus Meunier4 (Limoniidae), inclusions in Baltic amber, No. K2532 (male) (coll. Klebs), specimen from University of Göttingen: (A) drawing of hypopygium, dorsal view (reconstruction); (B) drawing of tip of gonocoxite without spine and gonostyles, ventral view (reconstruction). gx—gonocoxite; ing—inner gonostylus; oug—outer gonostylus.

Toxorhina (Ceratocheilus) eridanus Meunier4 (Limoniidae), inclusion in Baltic amber, specimen from Natural History Museum Humboldt University, Berlin (NHMB) (female): (A) body, lateral view; (B) enlarged view of head, lateral view; (C) enlarged view of ovipositor; (D) enlarged view of the tip of rostrum with palpus visible; (E) amber piece with habitus in lateral view. fl—flagellum; scp—scape; ped—pedicel; p—palpus.
1917 Ceratocheilus eridanus—Meunier; p. 96–97, pl. 15, Figs. 67, 68, pl. 16, Fig. 72.
1931 Ceratocheilus eridanus—Alexander, p. 124, Figs. 168, 169.
1982 Ceratocheilus eridanus—Keilbach, p. 326.
1994 Toxorhina (Ceratocheilus) eridanus—Evenhuis, p. 87.
2015 Toxorhina (Ceratocheilus) eridanus—Kania, p. 85, 86, Figs. 26A–C, 34A–E.
Material examined. No. 925/det. Alexander, 1925 (female) Natural History Museum Humboldt University, Berlin (NHMB), No. K2532 (male) University of Gottingen (GMUG), coll. Klebs, No. CCHH 832-1 (male) Senckenberg Deutsches Entomologisches Institut (SDEI) Müncheberg, Germany, coll. Christel and Hans Werner Hoffeins; No. MP/4442 (male) Institute of Systematics and Evolution of Animals, Polish Academy of Sciences, Kraków, Poland (ISEA PAS).
Additional description. Body 6.48 long (male) (Figs. 1A–I; 2A,B)—7.43 long (excluding rostrum) (female) (Fig. 3A–E), head: head capsule 0.54 high (male), 0.72 long (male); rostrum 3.99 long (female)—4.38 long (male); antenna 0.67 long (1/0.09; 2/0.11; 3/0.06; 4/0.03; 5/0.03; 6/0.04; 7/0.04; 8/0.04; 9/0.04; 10/0.05; 11/0.06; 12/0.08) (male), 0.74 long (female), elongate setae of antenna 0.63 long (male); palpus very short; rostrum shorter than wing, antenna about 0.14 × of rostrum.
Thorax: wing 4.73 (female)—5.13 long, 0.85 (male)—1.35 (female) wide; Rs 0.44 mm (male) long; two last tarsomeres almost equal in length.
Abdomen: male terminalia—hypopygium 0.50 long, gonocoxite 0.37 long, ovipositor 1.46 long.
Remarks. Even though, Meunier3 provided a brief description of the male and female of Toxorhina (Ceratocheilus) eridanus and the species was reviewed by Alexander4 based on males specimens, specified clear diagnostic characters were given in 20155. New fossil materials allow to provide additional description of the species.
Toxorhina (Ceratocheilus) christelius sp. nov.

Toxorhina (Ceratocheilus) christelius sp. nov. (Limoniidae), inclusion in Baltic amber, (A) No. CCHH 832-2 (male), (coll. Ch. and H.W. Hoffeins), specimen from Senckenberg Deutsches Entomologisches Institut (SDEI) Müncheberg, Germany: (A) drawing of head with part of rostrum, lateral view (reconstruction); (B) relation between the length of rostrum (r), antenna (a) and wing (w); (C–F) No. MP/4441 (male), holotype, specimen from Institute of Systematics and Evolution of Animals, Polish Academy of Sciences, Kraków, Poland (ISEA PAS): (C) drawing of wing; (D) photograph of wing; (E) head, frontal view; (F) tip of rostrum with palpi well visible.

Toxorhina (Ceratocheilus) christelius sp. nov. (Limoniidae), inclusion in Baltic amber, (A–E) No. MP/4441 (male), holotype, specimen from Institute of Systematics and Evolution of Animals, Polish Academy of Sciences, Kraków, Poland (ISEA PAS): (A) body, ventral view; (B) photograph of hypopygium with spine on gonocoxite, ventral view; (C) enlarged view of spine on gonocoxite; (D) drawing of hypopygium—gonocoxite with spine, ventral view (reconstruction); (E, F) No. MP/4443 (male), holotype, specimen from Institute of Systematics and Evolution of Animals, Polish Academy of Sciences, Kraków, Poland (ISEA PAS): (F) photograph of hypopygium, latero-ventral view; (G) photograph of enlarged view of spine on gonocoxite, latero-ventral view. Abbreviations: the black arrow indicates spine.
Diagnosis. Crossvein sc-r situated before fork of Rb; Rs elongate, longer than basal section of R5 between its origin and point of contact with r-m; R2+3+4 about twice as long as Rs, reaching wing margin in approximately 0.3 × distance from tip of R1 to tip of R5; tip of R2+3+4 just beyond level of d-cell; d-cell almost rhomboidal; m-cu at or just before fork of Mb on M1+2 and M3+4; distance between M1+2 and M3 narrow, branch M3 and M4 about twice as wide as distance of M1+2 and M3 (in the widest area); gonocoxite with distinctly distinguished, wide, huge spine of internal part of hypopygium.
Etymology. The specific name is dedicated to Christel Hoffeins (Hamburg, Germany), the owner of amber inclusions collections and the expert of inclusions in Baltic amber.
Material examined. Holotype No. MP/4441 (male) Institute of Systematics and Evolution of Animals, Polish Academy of Sciences, Kraków, Poland (ISEA PAS). Additional material: No. MP/4443 (male) (ISEA PAS), No. CCHH 832-2 (male) Senckenberg Deutsches Entomologisches Institut (SDEI) Müncheberg, Germany (the collection of Christel and Hans Werner Hoffeins).
Description. Body (Figs. 4A–F; 5A–D) 4.41–6.03 long (excluding rostrum), brown, wings without color pattern, pterostigma absent.
Head: head capsule 0.47 high, 0.54 wide, rostrum 2.86–3.81 long; antenna about 0.55 long (1/0.06; 2/0.10; 3/0.06; 4/0.02; 5/0.02; 6/0.03; 7/0.03; 8/0.03; 9/0.03; 10/0.04; 11/0.05; 12/0.08); approximately as long as head, scape cylindrical, narrow, short, pedicel large, massive, widened in midlength, slightly wider than long, first flagellomere large, wider than long, much wider than other flagellomeres but distinctly smaller than pedicel, remaining flagellomeres cylindrical, longer than wide, flagellomeres become slender and more elongate to apex of antenna, last flagellomere narrow, on the last three flagellomeres occur very elongate setae according to pattern: eight flagellomere with one very elongate setae, the last two flagellomeres with three very elongate setae, longer than half length of antenna, approximately 0.32 long; additionally on flagellomeres are presented two not very elongate setae, much shorter than length of flagellomeres bearing them and numerous very short setae; palpus very short, 0.04 long, about 0.125 × of rostrum; rostrum shorter than wing, antenna about 0.16 × of rostrum.
Thorax: scutellum widened in midlength; wing 3.67–4.77 long, 1.06–1.28 wide; Rs 0.62 long; vein M3 1.5 × the length of d-cell; d-cell twice as long as wide, 0.33 long; haltere with narrow, elongate stem slightly longer than knob.
Abdomen: male terminalia—hypopygium 0.43–0.49 long, gonocoxite, approximately twice as long as wide with wide, massive spine on internal part of ventral side of gonocoxite; outer gonostylus elongate, inner gonostylus narrow, sharpened at their end, slightly widened at base, generally tiny and slender.
Comparison. T. (C.) christelius sp. nov. differ from T. (C.) eridanus especially by morphology of hypopygium. Gonocoxite of a new species comes with distinctly distinguished, wide, huge spine on internal part of ventral side of gonocoxite, clearly visible from ventral side of hypopygium. In T. (C.) eridanus spines on gonocoxite does not occur, the gonocoxite of T. (C.) eridanus is only slightly widened distally. Moreover, tip of vein R2+3+4 in T. (C.) christelius sp. nov. is situated at the level of fork of M1+2 or just beyond, in T. (C.) eridanus tip of R2+3+4 is situated far beyond level of d-cell, while for example in T. (C.) caucasiensis vein R2+3+4 is rather short and its tip is situated approximately at the level of half d-cell. In T. (C.) mexicana R2+3 is very short, equal in length to Rs, while in all known from fossil record species is longer. In T. (C.) eridanus and T. (C.) christelius sp. nov. sc-r occur before or at fork of Rb, in T. (C.) caucasiensis beyond this bifurcation. In addition, the distance between veins M1+2 and M3 in the T. (C.) christelius sp. nov. is narrow, what is well visible at the level of vein m-m, in T. (C.) caucasiensis, T. (C.) eridanus or T. (C.) mexicana veins M1+2 and M3 are widely separated. In contrast to T. (C.) madagascariensis, basal part of d-cell of T. (C.) madagascariensis is narrowed, in new species is distinctly wide.
Key to fossil species of genus Toxorhina
-
1.
Basal part of d-cell rectangular.......2.
–. D-cell narrowed at its base......T. madagascariensis Meunier
-
2.
2. Vein R2+3+4 longer than Rs; tip of R2+3+4 beyond the level of fork of Mb......3.
–. Vein R2+3+4 very short, equal in length of vein Rs; tip of R2+3+4 before or at the level of fork of Mb......T. (C.) mexicana Kopeć, Kania and Krzemiński (Fig. 6G,H)
Figure 6 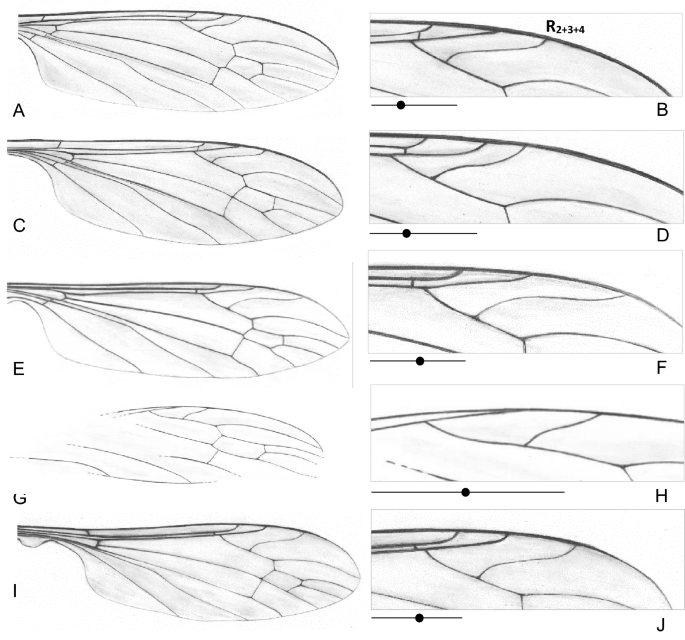
Wing venation of fossil Toxorhina (Ceratocheilus): (A–D) Toxorhina (Ceratocheilus) caucasiensis Krzemiński and Freiwald6; (E, F) Toxorhina (Ceratocheilus) eridanus Meunier5, (G, H) Toxorhina (Ceratocheilus) mexicana Kopeć, Kania, Krzemiński, 2016 (after Kopeć et al.7), (I, J) Toxorhina (Ceratocheilus) christelius sp. nov. (B, D, F, H, J)—enlarged view of part of wing with position of vein R2+3+4 with a diagram showing the position (black point) of the Rs bifurcation.
-
3.
3. Tip of R2+3+4 positioned at the level of d-cell; sc-r occurs at or before the level of fork of Mb; the distance between tips of R1 and R2+3+4 rather wide, about 0.3× the distance between tips of R2+3+4 and R5; tip of A1 just before of m-cu and of fork of Mb level......4.
–. Tip of R2+3+4 positioned before the level of d-cell; sc-r occurs beyond the level of fork of Mb; the distance between tip of R1 and R2+3+4 very short, about 0.2× the distance between tips of R2+3+4 and R5; tip of A1 at or just beyond of m-cu and of fork of Mb level......
T. (C.) caucasiensis Krzemiński and Freiwald (Fig. 6A–D)
-
4.
Wide spine present on gonocoxite; R2+3+4 separates Rs beyond half the length of the distance from fork of Rb to the point of connection of cross-vein r-m; the distance between M1+2 and M3 narrow, about twice as narrow as branch of M3 and M4; d-cell almost rhomboidal......
T. (C.) christelius sp. nov. (Figs. 4; 5; 6I,J)
–. Gonocoxite without spine; R2+3+4 separates Rs before half the length of the distance from fork of Rb to the point of connection of cross-vein r-m; the distance between M1+2 and M3 wide, about as wide as branch of M3 and M4 or slightly wider; d-cell almost rectangular;......
T. (C.) eridanus Meunier (Fig. 6E,F)

Discussion
The subgenus Ceratocheilus is well distinguishable from two other subgenera of Toxorhina by morphology of wing with two branches of Rs reaching the wing margin and the presence of corniculus (soft, pale, triangular sac on anterior vertex). In subgenera as Toxorhina and Eutoxorhina single branch of Rs reaches the wing margin9, therefore in these subgenera venation tends to be reduced and simplified (three medial veins reaching the margin in subgenus Toxorhina and two medial veins reaching margin in Eutoxorhina). The antennae of Toxorhina sensu lato are peculiar, in the male 12-segmented, with several basal antennomeres united into truncated, conical fusion-segment, the flagellar antennomeres, except the last two, glabrous, the outer pair with very long and conspicuous verticils. In the female the antenna is apparently 14 or l5 segmented, with the outer four or five flagellomeres provided with elongate verticils. Male hypopygium is peculiar and variable, in general basistyle (gonocoxite) bears or not an apical spine; dististyles (gonostyles) are variously shaped; aedeagus deeply to profoundly bifid, more or less resembling a tuning fork; ovipositor with the cerci long and slender, gently upcurved to the acute tips; hypogynal valvae shorter, compressed. All subgenera of Toxorhina are characterized by elongated rostrum, exceeding half of body length, or the entire body or wing length, bearing the reduced mouthparts at the tip. The biology of the group is known very little, adults use the long mouthparts to probe a variety of flowering plants for nectar10,11. The immature stages were found in slimy, well rotten wood and branches, under leaf litter, in the saturated soil of swamps and in the liquid between fibers in petioles of rotten palm fronds11,12. Adults found at the salt marshes13 also support such kind of habitat for immatures. The species of the genus Toxorhina are believed to represent strongly an ‘equatorial’ group2 (Table 1, Supplementary Table 1). Subgenus Ceratocheilus covers 18 species recorded in Afrotropical region, eight in the Madagascan region, 20 in Oriental, seven in Australian and 16 in Oceanian, two in Novozealandic, ten in Panamanian and three species in Neotropical regions. The subgenus Ceratocheilus is absent in Holarctic regions, also not recorded in Saharo-Arabian and reaching borders of Sino-Japanese regions. Subgenus Toxorhina presents a different pattern, with 24 species in the Panamanian region but with two species reaching further north to Nearctic region and a three more south-east to Neotropical region, 16 species recorded in Oriental region, 13 in Oceanian, ten in Afrotropical and two in Madagascan regions. Subgenus Eutoxorhina is restricted to single species in Australian region and two in Oceanian region. The data available suggest more restricted distributional patterns of Ceratocheilus in tropical-subtropical zones. Some species from northern parts of Oriental region—like e.g. Toxorhina (Ceratocheilus) bistyla Alexander, T. (C.) brevifrons (Brunetti), T. (C.) fulvicolor Alexander, T. (C.) huanglica Zhang, Li and Yang, T. (C.) omnifusca Zhang, Li and Yang are distributed in higher, montane habitats, between 1000 and 3500 m a.s.l.9,14,15, these from more southern parts of Oriental regions seem to present similar preferences. Montane habitats seems to be favorable also for Panamanian species of subgenus Ceratocheilus, the same could be inferred for Oceanian (mainly known from New Guinea) species. Afrotropical species seem to be present in various habitats from highlands to lowland marshlands, the latter seems to be also habitat for Neotropical species. Regarding the scarce fossil record of the subgenus Ceratocheilus—the oldest known species come from the Eocene from Baltic amber—T. (C.) eridanus Meunier and described above T. (C.) christelius sp. nov. These finding gave some additional data to discussion on reconstructions of the palaeohabitats of the Baltic amber forests. Some montane species of insects were reported from Baltic amber, as well as these with modern relatives present in marshlands and other humid habitats16.
Miocene findings of the fossils ascribed to the subgenus Ceratocheilus are limited. A female named Toxorhina (Ceratocheilus) mexicana Kopeć, Kania and Krzemiński was described from Lower Miocene (Burdigalian) amber of Chiapas (Mexico) (Fig. 7). The reconstruction of the Mexican amber forest shows it to be close to modern lowland tropical forest, especially to mangrove formation17,18,19,20.

Map of the distribution of know localities of the representatives of the genus Toxorhina.
Another species comes from Middle Miocene, ca. about 13 Ma (Late Chokrakian; Serravalian) deposits of Stavropol (Stavropol’ Region, Russia). The aquatic groups (dragonflies, caddisflies, mayflies, aquatic beetles, bugs, some dipterans) compose 30% of all found insect imprints (of a total some 3000 specimens)21. The depositional conditions reconstructed presents mixture of lentic and lotic habitats, possibly a lagoon, a coastal freshened area, with periodical inflow of fresh water from nearby rivers, the presence of oxbow lakes and creeks21. Such habitats correspond quite well with known requirements of recent species of the subgenus Ceratocheilus, at its immature stages. Former investigations22,23 indicated climatic conditions shifting from warm humid to tropical during the time of deposition, with several biocenosis from littoral and coast plants associations, submerged and waterside plant associations, floodplain forests, deciduous-evergreen forests, to light, dry pine forests and light, dry deciduous forests and steppe type shrubs. Such variable habitats seems to offer a good food-source for imagines of flies.
The existence of a warmer and more equable climate in the Eocene and Miocene is now an indisputable fact24. These times were probably suitable for ancestors of recent taxa of the subgenus Ceratocheilus to spread and migrate in search of appropriate habitats. During the Eocene Temperature Optimum an exchange processes between the faunas of North America and the Western Palearctic was possible due to existence of land bridges24,25. Subsequent cooling, a decrease in the CO2 and oxygen concentrations in the atmosphere, aridization, etc., during the Oligocene decreased availability of appropriate habitats and farther changes and biogeographic shifts were caused by the next Miocene Climate Optimum26. The data available for the genus Toxorhina and its subgenera are very scarce, however patterns observed among other insects, e.g. in thermophilic ants23 or Lophopidae planthoppers27,28,29 could be a good reference for reconstruction of biogeographic scenario of these flies. Ancestral forms of Toxorhina most probably diversified during the warming periods of the Eocene (Paleocne-Eocene Thermal Maximum and Eocene Climatic Optimum) in the warm, humid areas of Europe in Thulean and western part of Boreal regions30, as documented by fossils from Baltic amber. The Limoniidae genera Elephantomyia Osten Sacken and Helius Lepeletier and Serville have rostrum elongate as in Toxorhina and its subgenera, in the genus Geranomyia Haliday mouth parts of are strongly elongate, but rostrum short9,31,32,33. The phylogenetic position of the genus Toxorhina within Limoniinae remains unclear, but it could be assumed that it separated somewhere in Late Cretaceous with diversification and spread to various habitats of nectar producing angiosperms34,35. Tendency to elongation of rostrum is observed in Helius since the Cretaceous with wide diversity in the Eocene32,33, and similar disparity pattern could be assumed for ancestors of Toxorhina. In addition, the morphological disparity of the Toxorhina is exposed also in venation patterns resulting in taxonomic separation to subgenera. The plesiomorphic condition in Limoniinae is placement of veinlet m-cu behind fork of Mb, in the Eocene Ceratocheilus it is placed at fork of Mb and in the Mid-Miocene taxa it is place anterior of fork or at fork of Mb. Regarding distributional patterns it could be assumed that ancestors of modern subgenus Toxorhina as well as ancestors of modern Ceratocheilus rapidly spread in the northern Hemisphere, crossing Atlantic Ocean in the Paleocene to earliest Eocene through De Geer and Thulean land bridges existing at these times24,25. Eastward spreading of ancestors of all subgenera probably took place in the Palaeocene–Eocene as well, but with Oligocene and Miocene climatic changes, colling and drying and decline of megathermal forest36,37, ancestors of modern representatives of subgenera Toxorhina and Ceratocheilus became relic in montane megathermal localities2. These changes together with tectonic events27 in the area resulted in further diversification and ancestors of modern subgenus Eutoxorhina reached Australia, Fiji and New Caledonia2. In the New World ancestors of both subgenera Toxorhina and Ceratocheilus could be assumed to migrate to suitable habitats with tectonic and climatic changes37,38,39, to montane and swampy areas of Central and South America2. Here these forms diversified and adapted to local conditions resulting in modern diversity (Table 1, Supplementary Table 1).
These disparity events resulted in the differences of the relation between the length of rostrum and wing species of recent fauna of Ceratocheilus. In Toxorhina (Ceratocheilus) fulvicolor Alexander described from India rostrum is shorter than wing as in Toxorhina (Ceratocheilus) contractifrons (Edwards) known from Malaysia or Toxorhina (Ceratocheilus) omnifusca described from China. In fossil representatives of subgenus e.g. in T. (C.) eridanus and newly described species T. (C.) christelius sp. nov. wing is longer than rostrum. The most obvious features that differentiate of some fossil species of Thoxorhina are morphology of hypopygium, but especially shape, position and length of vein R2+3+4, length of Rs, and position of tips of Sc and R1 (Fig. 6A–E). In T. (C.) christelius sp. nov. wide, massive spine occur on gonocoxite. For example in T. (C.) caucasiensis Rs is very short and R2+3+4 separate sector radii before half the length of Rs and basal section of R5 combined, while in T. (C.) eridanus R2+3+4 separate Rs beyond half the length of this section. Both, in recent and fossil fauna of Ceratocheilus morphology of antenna is not very differentiated, 12-segmented antenna with very elongate setae on the last segments, usually longer than half the length of antenna.
The most characteristic feature for subgenus Ceratocheilus is elongate rostrum, similarly to a craneflies belonging to the genus Helius Lepeletier and Serville or Elephantomyia Osten Sacken. The group of Limoniinae Speiser which is characterized by elongate rostrum, distinctly differ by other morphological feature, for example last palpomere of the genus Helius is very elongate, sometimes reaching the length of the previous three, while in representatives of subgenus Ceratocheilus of the genus Toxorhina palpus is reduced, similarly to this of Elephantomyia. Moreover, relations of the length of head or body, antenna, palpus and rostrum distinctly differ in these three subgenera. Elongate rostrum of Helius perhaps was used for a specific food spectrum, it was probably the nectar of flowers33,40,41,42,43,44. These three groups of insects are probably closely related, as it was suggested based on molecular data and fossil analysis the genera as Helius, known since early Cretaceous period, and much younger Elephantomyia44,45. The discovery of new Ceratochelius in Eocene Baltic amber contributes to a better understanding of the diversity and disparity of the genus Toxorhina and also subfamily Limoniinae, and it is important for further research on the evolution of this group of insects. So far, only seven specimens of Toxorhina (Ceratocheilus) from Baltic amber were known, new materials (seven not known so far specimens) give us interesting information, especially about morphology of gonocoxite, for example in new species T. (C.) christelius sp. nov. characteristic wide spine on gonocoxite is present, while in T. (C.) eridanus this structure is absent.
Conclusions
The finding of more fossils of Toxorhina should make it possible to understand the patterns of morphological evolution and disparity associated with the adaptation of these flies to their mode of life, present conclusions about their (palaeo)biogeography, and evolution of biocoenoses inhabited by these insects.
Material and methods
Taxonomic treatments
To avoid a lengthy list of references dealing with scientific names detailed taxonomic treatment and chresonymy is given in the supplementary materials.
Geological context
The age range of all Baltic amber bearing strata possibly cover 48 to 23 million years, but it is still controversial though has been a matter of debate for many years46,47,48,49,50,51,52,53,54,55. Based on pollen, spores and phytoplankton of the amber embedding layer, the Blue Earth the most current state of knowledge is that it is of Priabonian age, estimated between 38 and 34 million years old47,52. The age of Baltic amber has also been determined to approximately 47–41 Ma4, but the reliability of the methods used in his study have been questioned due to contaminations that can lead to older age estimations48,49,50,51,53,54.
Specimen repository
The study was based on seven inclusions of the subgenus Ceratocheilus (Limoniidae: Toxorhina) preserved in Baltic amber. All the studied specimens are deposited in public institutions. The material examined is deposited in Senckenberg Deutsches Entomologisches Institut (SDEI) Müncheberg, Germany (the collection of Christel and Hans Werner Hoffeins) (two inclusions), Natural History Museum Humboldt University, Berlin (NHMB) (one specimen), University of Göttingen (GMUG) (one specimen), Institute of Systematics and Evolution of Animals, Polish Academy of Sciences, Kraków, Poland (ISEA PAS) (three specimens).
Methods
The pieces of amber were examined using a Nikon SMZ 1500 stereomicroscope equipped with a Nikon DS-Fi1 camera, and the measurements were taken with NIS-Elements D 3.0 software in the lab of the Department of Biology, University of Rzeszów. Measurements of individual parts of the body were given only when the measured morphological structures were not distorted. All measurements are given in mm. Measurements of the vein M3 were taken from the point of connection of vein m-m with vein M3 to the margin of wing, measurements of the discal cell were taken from the proximal to distal ends of the d-cell. The length of hypopygium was measured from the posterior margin of tergite IX to the apex of gonocoxite. Basal section of R5 was mentioned as the length from fork of Rs to the point of connection R5 with crossvein r-m. Drawings were made based on specimens and the photographs by Iwona Kania-Kłosok. The map was built using the map Maps-For-Free (https://maps-forfree.com) and modified with the software packages Corel Draw and Corel PhotoPaint X7. The wing venation nomenclature is follows Alexander5, Krzemiński and Krzemińska56, the term “d-cell base” is used after Krzemiński and Freiwald3, the designation of the hypopygium were introduced after McAlpine et al.57 and Podenas58.
Nomenclatural acts
The electronic edition of this article conforms to the requirements of the amended International Code of Zoological Nomenclature, and hence the new names contained herein are available under that Code from the electronic edition of this article. This published work and the nomenclatural acts it contains have been registered in ZooBank, the online registration system for the ICZN. The ZooBank LSIDs (Life Science Identifiers) can be resolved and the associated information viewed through any standard web browser by appending the LSID to the prefix ‘http://zoobank.org/’. The LSID for this publication is: urn:lsid:zoobank.org:pub: 89897A75-CCFF-45D3-B0DB-0D5B0B73C6C0, for a new species is: urn:lsid:zoobank.org:act: CC518ED7-39ED-4BD9-A0EE-00D05E7F8FD4.
Data availability
All data generated or analyzed during this study are included in this published article.
Abbreviations
- GMUG:
-
University of Gottingen
- ISEA PAS:
-
Institute of Systematic and Evolution of Animals, Polish Academy of Sciences
- NHMB:
-
Natural History Museum Humboldt University, Berlin
- NHM:
-
Natural History Museum, London, UK
- PIN RAS:
-
Paleontological Institute, Russian Academy of Sciences, Moscow
- SDEI:
-
Senckenberg Deutsches Entomologisches Institut
- ZMB:
-
Zoological Museum Berlin
References
Holt, B. et al. An update of Wallace’s zoogeographic regions of the World. Science 339, 74–78 (2013).
Oosterbroek, P. CCW: Catalogue of the Crane-flies of the World (Diptera, Tipuloidea: Pediciidae, Limoniidae, Cylindrotomidae, Tipulidae) (version May 2021). https://ccw.naturalis.nl/stats.phphttp://nlbif.eti.uva.nl/ccw/index.php. Last update 15 March 2022 (2022).
Meunier, F. Ueber einige Mycetophiliden und Tipuliden des Bernsteins nebst Beschreibung der Gattung Palaeotanypeza (Tanypezinae) derselben Formation. Neues Jahrbuch für Mineralogie Geologie und Palaontologie 1917(3), 73–106 (1917).
Alexander, C. P. Crane flies of the Baltic Amber (Diptera). Bernstein-Forschungen 2, 1–135 (1931).
Kania, I. The taxonomy of the selected genera of the subfamily Limoniinae (Diptera: Limoniidae) from Baltic amber (Eocene), with notes on their phylogeny. Ann. Zool. 65(1), 71–100 (2015).
Zhang, X., Li, Y. & Yang, D. A review of the genus Toxorhina Loew from China, with descriptions of three new species (Diptera, Limoniidae, Limoniinae). ZooKeys 480, 59–80 (2015).
Alexander, C. P. & McAtee, W. L. Diptera of the superfamily Tipuloidea found in the District of Columbia. Proc. U.S. Natl. Mus. 58, 385–435 (1920).
Alexander, C. P. Diptera (Nematocera): Tanyderidae, Ptychopteridae, Tipulidae. S. Afr. Anim. Life 10, 229–441 (1964).
Hynes, C. D. The immature stages and biology of the craneflies Toxorhina caledonica and Elephantomyia garrigouana (Diptera: Limoniidae). Pan Pac. Entomol. 73, 93–99 (1997).
Rodrigues, L., Ortega, I., Vieira, R., Carrasco, D. & Proietti, M. Crane flies (Diptera, Tipuloidea) from southern Neotropical salt marshes: Survey with DNA barcoding. Iheringia Série Zoologia 109(e2019013), 1–9. https://doi.org/10.1590/1678-4766e2019013 (2019).
Alexander, C. P. Undescribed species of crane flies from the Himalaya mountains (Diptera: Tipulidae). XV. J. N. Y. Entomol. Soc. 75, 183–187 (1967).
Brunetti, E. Revision of the Oriental Tipulidae with descriptions of new species, part 2. Rec. Indian Mus. 15, 255–344 (1918).
Szadziewski, R., Szwedo, J. & Sontag, E. Fauna lasu bursztynowego/Fauna of the amber forest. In Bursztyn Bałtycki—Skarb Zatoki Gdańskiej/Baltic Amber—Treasure of the Bay of Gdańsk (eds Szadziewski, R. et al.) 216–217 (Związek Miast i Gmin Morskich, 2018).
Poinar, G. Jr. & Brown, A. Hymenaea mexicana sp. nov. (Leguminosae: Caesalpinioideae) from Mexican amber indicates Old World connections. Bot. J. Linnean Soc. 139(2), 125–132 (2002).
Langenheim, J. H. Preliminary investigations of Hymenaea courbaril as resin producer. J. Arnold Arborboretum 48, 203–230 (1967).
Solórzano-Kraemer, M. M. Systematic, palaeoecology, and palaeobiogeography of the insect fauna from Mexican amber. Palaeontogr. Abt. A 282, 1–133 (2007).
Solórzano-Kraemer, M. M. Mexican amber. In Biodiversity of Fossils in Amber from the Major World Deposits (ed. Penney, D.) 42–56 (Siri Scientific Press, 2010).
Sukatsheva, I. D., Ivanov, V. B., Melnitsky, S. I. & Aristov, D. S. Caddisflies (Insecta: Trichoptera) from the Middle Miocene of the Stavropol Region, Russia. Paleontol. J. 52(13), 1593–1609 (2018).
Becker-Migdisova, E. E. Tertiary Homoptera of Stavropol and a method of reconstruction of continental palaeobioceonoses. Palaeontology 10(4), 542–553 (1967).
Perfilieva, K. S. Distribution and differentiation of fossil Oecophylla (Hymenoptera: Formicidae) species by wing imprints. Paleontol. J. 55(1), 76–89 (2021).
Sanmartín, I., Enghoff, H. & Ronquist, F. Patterns of animal dispersal, vicariance and diversification in the Holarctic. Biol. J. Lin. Soc. 73, 345–390 (2001).
Brikiatis, L. The De Geer, Thulean and Beringia routes: Key concepts for understanding early Cenozoic biogeography. J. Biogeogr. 41, 1036–1054 (2014).
Steinthorsdottir, M. et al. The Miocene: The future of the past. Paleoceanogr. Paleoclimatol. 36, e2020PA004037 (2021).
Soulier-Perkins, A. A phylogenetic and geotectonic scenario to explain the biogeography of the Lophopidae. Palaeogeogr. Palaeoclimatol. Palaeoecol. 160, 239–254 (2000).
Soulier-Perkins, A., Ouvrard, D., Attié, M. & Bourgoin, T. Evolutionary patterns in biogeography and host plant association: ‘Taxonomic conservatism’ in Lophopidae (Hemiptera, Fulgoromorpha). Syst. Entomol. 32, 305–311 (2007).
Szwedo, J., Stroiński, A. & Lin, Q. B. Tip of the clade on the top of the World—The first fossil Lophopidae (Hemiptera: Fulgoromorpha) from the Palaeocene of Tibet. Sci. Nat. 102(28), 1–5 (2015).
Brysz, A. M., Bourgoin, T. & Szwedo, J. Spotted beauty—Gedanochila museisucini gen. nov. et sp. nov.—A new Achilidae from Eocene Baltic amber (Hemiptera: Fulgoromorpha) and its relation to Achilini. Zootaxa 5125(3), 295–312 (2022).
Podenas, S. & Gelhaus, J. Identification keys for Limoniinae (Diptera, Limoniidae) of Mongolia and adjacent territories. Vilnius, Lithuania, 85 (2007).
Kania, I. Subfamily Limoniinae Speiser, 1909 (Diptera, Limoniidae) from Baltic amber (Eocene): The genus Helius Lepeletier & Serville, 1828. Zootaxa 3814(3), 333–352 (2014).
Krzemiński, W., Kania, I. & Azar, D. The early Cretaceous evidence of rapid evolution of the genus Helius Lepeletier and Serville, 1828 (Limoniidae, Diptera). Cretac. Res. 48, 96–101 (2014).
Folk, R. A., Siniscalchi, C. M. & Soltis, D. E. Angiosperms at the edge: Extremity, diversity, and phylogeny. Plant Cell Environ. 43, 2871–2893 (2020).
López-Martínez, A. M. et al. Integrating fossil flowers into the angiosperm phylogeny using a total evidence approach. Biorxiv https://doi.org/10.1101/2022.02.17.480913 (2022).
Westerhold, T. et al. An astronomically dated record of Earth’s climate and its predictability over the last 66 million years. Science 369, 1383–1387 (2020).
Lim, J. Y. et al. The Cenozoic history of palms: Global diversification, biogeography and the decline of megathermal forests. Glob. Ecol. Biogeogr. 31, 425–439 (2022).
Iturralde-Vinent, M. A. & Macphee, R. Paleogeography of the Caribbean region: Implications for Cenozoic biogeography. Bull. Am. Mus. Nat. Hist. 238(238), 1–95 (1999).
Zachos, J., Pagani, M., Sloan, L., Thomas, E. & Billups, K. Trends, rhythms, and aberrations in global climate 65 Ma to present. Science 292(5517), 686–693 (2001).
Kania, I., Krzemiński, W. & Azar, D. The oldest representative of Helius Lepeletier & Serville 1828 (Diptera: Limoniidae) from Lebanese amber (Early Cretaceous). Insect Syst. Evol. 44, 231–238 (2013).
Kania, I., Krzemiński, W. & Arillo, A. First representative of the genus Helius Lepeletier and Serville, 1828 (Diptera, Limoniidae) from the Lower Cretaceous Álava amber (Spain). Cretac. Res. 63, 33–38 (2016).
Kania, I., Krzemiński, W. & Arillo, A. A new peculiar species of the genus Helius Lepeletier & Serville, 1828 (Diptera, Limoniidae) from Cretaceous Álava amber (Spain). Earth Environ. Sci. Trans. R. Soc. Edinb. 107, 1–7 (2017).
Kania-Kłosok, I., Krzemiński, W. & Arillo, A. Two new long-rostrum cranefly species from the Cretaceous Iberian amber (Diptera, Limoniidae, Helius). Sci. Rep. 11, 12851 (2021).
Kania, I. Subfamily Limoniinae Speiser 1909 (Diptera, Limoniidae) from Baltic amber (Eocene): The genus Elephantomyia Osten Sacken 1860. PLoS ONE 10(2), 117434 (2015).
Bertone, M. A., Courtney, G. W. & Wiegmann, B. M. Phylogenetics and temporal diversification of the earliest true flies (Insecta: Diptera) based on multiple nuclear genes. Syst. Entomol. 33, 668–687 (2008).
Podenas, S. & Poinar, G. O. New crane flies (Diptera: Tipulidae, Limoniidae) from Dominican and Mexican amber. Proc. Entomol. Soc. Wash. 103(4), 863–878 (2001).
Clauer, N., Huggett, J. M. & Hillier, S. How reliable is the K–Ar glauconite chronometer? A case study of Eocene sediments from the Isle of wight. Clay Miner. 40, 167–176 (2005).
Grimaldi, D. & Ross, A. J. Extraordinary Lagerstätten in amber, with particular reference to the Cretaceous of Burma. In Terrestrial Conservation Lagerstätten: Windows into the Evolution of Life on Land (eds Fraser, N. C. & Sues, H.-D.) 303 (Dunedin Academic Press, 2017).
Kasiński, J. R. & Kramarska, R. Sedimentary environment of amber-bearing association along the Polish-Russian Baltic coastline. Exkursionsfuhrer und Veroffentlichungen der Deutschen Gesellschaft fur Geowissenschaften 236, 46–57 (2008).
Kasiński, J. R., Kramarska, R., Sodkowska, B., Sivkov, V. & Piwocki, M. Paleocene and Eocene deposits on the eastern margin of the Gulf of Gdansk (Yantarny P-1 bore hole, Kaliningrad region, Russia). Geol. Quart. 64(1), 29–53 (2020).
Kosmowska-Ceranowicz, B., Kohlman-Adamska, A. & Grabowska, I. Erste Ergebnisse zur Lithologie und Palynologie der bernsteinfuhrenden Sedimente im Tagebau Primorskoje. Metalla Sonderheft 1, 5–17 (1997).
Sadowski, E.-M., Seyfullah, L. J., Schmidt, A. R. & Kunzmann, L. Conifers of the ‘Baltic amber forest’ and their palaeoecological significance. Stapfia 106, 1–73 (2017).
Standke, G. Die Tertiärprofile der samländischen Bernsteinküste bei Rauschen. Schriftenreihe für Geowissenschaften 7, 93–133 (1998).
Standke, G. Bitterfelder Bernstein gleich Baltischer Bernstein?—Eine geologische Raum- Zeit- Betrachtung und genetische Schlussfolgerungen. Exkursionsführer und Veröffentlichungen der Deutschen Gesellschaft für Geowissenschaften 236, 11–33 (2008).
Ritzkowski, S. K-Ar—Altersbestimmungen der bernsteinführenden Sedimente des Samlands (Paläogen, Bezirk Kaliningrad). Metalla Sonderheft. Neue Erkenntnisse zum Bernstein 66, 19–23 (1997).
Krzemiński, W. & Krzemińska, E. Triassic Diptera: Descriptions, revisions and phylogenetic relations. Acta Zoologica Cracoviensia 46, 153–184 (2003).
Krzemiński, W. & Freiwald, A. Toxorhina (Ceratocheilus) caucasiensis, a new species from the Middle Miocene of Stavropol (northern Caucasus, USSR) (Diptera, Limoniidae). Palaontologische Zeitschrift 65(1/2), 153–155 (1991).
McAlpine, J. F. B., Peterson, B., Shewell, G., Teskey, H., Vockeroth, J. & Wood, D. Manual of Nearctic Diptera, Vol. 1. Agriculture Canada Research Branch Monograph 27, 674 (1981).
Podenas, S. Rhabdomastix crane flies (Diptera, Limoniidae) in Baltic amber (Eocene). Proc. Entomol. Soc. Wash. 155, 41–66 (2006).
Kopeć, K., Kania, I. & Krzemiński, W. New and little known crane-fly species of the genera Helius, Elephantomyia and Toxorhina (Diptera, Limoniidae) from Dominican and Mexican amber. Palaeontol. Electron. 19.2.26A, 1–14 (2016).
Meunier, F. Sur une nouvelle espèce deToxorrhina du copal récent de Madagascar. Bulletin Société d'étude des sciences naturelles et du Musée d’Elbeuf 24, 97–100 (1906).
Acknowledgements
We would like to thank two Anonymous Reviewers for their valuable comments and remarks. We are deeply grateful to Christel and Hans Werner Hoffeins from Hamburg, Germany for making the Ceratocheilus specimens available for our study. This research was realized within the project of the National Science Centre, Poland, Grant No. 2020/37/B/NZ8/03042.
Funding
Open Access funding enabled and organized by University of Rzeszów.
Author information
Authors and Affiliations
Contributions
I.K.-K. conceived and designed the study, lead and performed the data analysis, interpretations and analysis, prepared photographs and drawings, wrote first draft and corrected the manuscript. W.K. analyzed the data and corrected the manuscript. J.S. performed paleobiogeographical and distributional analyses and corrected the manuscript. All authors edited and reviewed final version of the manuscript and agreed to the submitted version.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Kania-Kłosok, I., Krzemiński, W. & Szwedo, J. New finding of Toxorhina (Ceratocheilus) limoniid fly in Eocene Baltic amber and the biogeographical context of the genus. Sci Rep 12, 19382 (2022). https://doi.org/10.1038/s41598-022-23866-7
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-022-23866-7
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
