
Abstract
In the last decade, a plethora of microRNAs (miRNAs) has been reported in a wide variety of physiological processes, including reproduction, in many aquatic organisms. However, miRNAome alterations occurred by environmental cues due to water temperature increment have not yet been elucidated. With the aim to identify epigenetic regulations mediated by miRNAs in the gonads in a climate change scenario, the animal model zebrafish (Danio rerio) were subjected to high temperatures during sex differentiation, a treatment that results in male-skewed sex ratios in the adulthood. Once the fish reached adulthood, gonads were sequenced by high-throughput technologies and a total of 23 and 1 differentially expressed miRNAs in ovaries and testes, respectively, were identified two months after the heat treatment. Most of these heat-recorder miRNAs were involved in human sex-related cancer and about 400 predicted-target genes were obtained, some with reproduction-related functions. Their synteny in the zebrafish genome was, for more than half of the predicted target genes, in the chromosomes 7, 2, 4, 3 and 11 in the ovaries, chromosome 4 being the place where the sex-associated-region (sar) is localized in wild zebrafish. Further, spatial localization in the gonads of two selected heat-recorder miRNAs (miR-122-5p and miR-146-5p) showed exclusive expression in the ovarian germ cells. The present study expands the catalog of sex-specific miRNAs and deciphers, for the first time, thermosensitive miRNAs in the zebrafish gonads that might be used as potential epimarkers to predict environmental past events.
Similar content being viewed by others

Introduction
Water sea temperature levels have been rising in the last 60 years1, with critical consequences for marine and aquatic life. Fish, thanks to their thermal plasticity, are able to survive the variations of water temperatures2. Nevertheless, sex determination in fish, unlike in mammals, is regulated by genetic and environmental factors3,4, and consequently, higher temperatures during sex differentiation skew the sex ratio towards males in many fish species5. Since the first study showing the crosslink between masculinization occurred by heat treatments and DNA methylation in the European sea bass (Dicentrarchus labrax) gonads6, in the last decade, studies describing the role of epigenetics in sexual development have emerged. In pufferfish (Takifugu rubripes), DNA methylation alterations were able to faithfully describe dimorphic differences in the gonadal epigenomes of fish subjected to different thermal regimes, identifying two genes (amhr2 gene and pfcyp19) as main actors of sex determination in this species7. Similarly, in half-smooth tongue sole (Cynoglossus semilaevis) differentially methylated regions (DMR) were observed in the gonads of sex-reversed fish indicating that high-temperature treatments override sexual fate determined by genetic factors through epigenetic pathways8. Recent transgenerational studies in zebrafish (Danio rerio) showed that temperature affected the testicular epigenome in the first generation but these effects were washed out in the second generation9.
miRNAs are small, non-coding RNAs, consisting of approximately 22 nucleotides, and are considered as epigenetic mechanisms responsible to regulate the post-transcriptional cellular machinery. These molecules regulate gene expression by preventing protein translation through binding to their target messenger RNAs (mRNA), serving as recruiters in the mRNA degradation pathways10. Over the past two decades, miRNA-related research has expanded considerably, as in only 2 years the miRNA submissions in public databases increased by fifty percent11. Over 3500 mature miRNAs have been identified in 16 teleostei species (www.mirbase.org), zebrafish being the first fish species with more miRNAs described in detail12. In fish, miRNAs play pleiotropic functions, for example, in the reproduction system, immune system, metabolism, and skeletal formation, among others (reviewed in13). In the last few years, studies in adult fish have revealed the presence of sexual dimorphism in the miRNA expression between ovary and testis in some species such as14, yellow catfish (Pelteobagrus fulvidraco)15, rainbow trout (Oncorhynchus mykiss)16, tilapia (Oreochromis niloticus) and zebrafish17,18.
Since the miRNome alterations can respond to environmental influences, they are foreseen as potential targets for improving productivity in aquaculture. Some studies have addressed the temperature effects on different target tissues. miRNA expression changed in response to increasing natural temperatures in zebrafish embryonic fibroblast cells19 and in rainbow trout liver20 and head kidney21. Cold tolerance has been tested by detailing miRNA expression in Emerald rockcod (Trematomus bernacchii) gills22, in turbot (Scophthalmus maximus) brain, head kidney and liver23, and in sole (Solea senegalensis) embryos24. To date, only few studies have addressed the miRNA alterations due to temperature increases in the gonads. Juvenile Atlantic cod (Gadus morhua) showed some, but few, DE miRNAs after heat during early development, although no differences between ovaries and testes were addressed25. Similarly, adult zebrafish gonads subjected to high temperature in combination with antidepressant compounds showed a variation of the miRNA abundance by a target miRNA approach 26. Thus, to our knowledge, no data regarding the long-term effects of the high temperatures in the miRNome of the ovaries and testes in fish have ever been reported. Therefore, the goal of this study was to characterize a set of miRNAs that could be used as epimarkers of the effects of heat-stress on fish gonads in a context of global warming.
Materials and methods
Experimental design
The AB zebrafish were reared at the experimental aquarium facilities of the Institute of Marine Sciences (ICM-CSIC) in Barcelona. Fish husbandry and thermal treatments were done as previously described in Ribas et al27. For this experiment, ~ 175 spawned eggs by a single pair mating were used. At 6 days post fertilization (dpf), 35 larvae were equally distributed into four tanks (two technical replicates for each group) of 2.8 L (Aquaneering, mod. ZT280) to avoid high-density masculinization effects28. Fish were exposed to high temperature (HT) at 34 ± 0.5 °C or to control temperatures (CT) at 28 ± 0.5 °C between 18 and 32 dpf. The temperature was changed at a rate of 1.5 °C/day to reach the desired temperatures. After the heat treatment, animals were grown until gonadal maturation, i.e., 90 dpf. The Chi-squared test with arcsine transformation was used to study differences in sex ratios. Biometry differences between CT and HT were determined by Student t-tests. Previously, for each group, homoscedasticity of variances and normality were checked by Levene’s test and Shapiro–Wilk test, respectively.
Sampling, sample selection and RNA extractions
Adult fish were sacrificed by cold thermal shock and the sex of the fish was visually assessed under the microscope. Gonads were isolated and flash-frozen into liquid nitrogen and kept at − 80 °C for further analyses. To unify the gonadal maturation, samples for sequencing analyses were selected based on two criteria. The first, based on macroscopical examination following Ribas et al27 by which all the selected samples were classified as Type 3 maturation, meaning fully mature gonads and, the second criteria, based on the highest gene expression levels of gonadal aromatase (cyp19a1a) and anti-Müllerian hormone (amh) in ovaries and testes, respectively, that worked as sex-markers (data not shown)29. miRNA of 16 gonads (four samples for each sex and treatment) was isolated by miRNAs isolation commercial kit (Qiagen® miRNA, 217004) and quality was assessed by BioAnalyzer (2100 Bioanalyzer, Agilent Technologies). On average, RNA Integrative Number (RIN) values for all the samples were ≥ 9 in all ovarian samples and between 7.6 and 9.6 in testes samples (Dataset 1, indicating high score RNA qualities.
Small RNA library and sequencing
In total, 16 libraries were constructed individually from zebrafish gonads. Library preparation was performed by NEBNext® Small RNA Library Prep Set for Illumina® (Multiplex Compatible) kit following manufacturer’s instructions. Sequencing (1 × 50, v4, HiSeq) was performed at single-end mode with a read length of 50 bp at the Genomics Unit of the Centre for Genomic Regulation (CRG) in Barcelona.
miRNA validation
Validation of the miRNA sequencing data was done by qPCR of those eight selected sequenced miRNAs. cDNA was generated using the miRNA 1st-Strand cDNA Synthesis Kit (Agilent Technologies) following manufacturer’s instructions. Firstly, the polyadenylation reaction was performed after cDNA synthesis. qPCR was performed using the qPCRBio SyGreen blue mix low ROX (PCR Biosystems). A mix of 5 µL 2 × qPCRBIO SyGreen Blue mix, 0.4 µL forward primer, 0.4 µL universal reverse primer (Agilent Technologies), 100 ng cDNA and H2O up to 10 µL was made for each sample. The sequences of the forward primers for the selected miRNAs were as follows:
dre-miR-202-5p: TTCCTATGCATATACCTCTTT, dre-miR-92a-3p: TATTGCACTTGTCCCGGCCTGT, dre-miR-21-5p: TAGCTTATCAGACTGGTGTTGGC, dre-miR-146b-5p: TGAGAACTGAATTCCAAGGGTG, dre-miR122-5p: TGGAGTGTGACAATGGTGTTTG, dre-miR-726-3p: TTCACTACTAGCAGAACTCGG, dre-miR-726-5p: GGAATTCCGCTAGTTCTGAACT, dre-miR-143-3p: TGAGATGAAGCACTGTAGCTC. The dre-U6: ACTAAAATTGGAACGATACAGAGA, was used as the reference gene. A total of 9 comparisons for validations were performed as follows: dre-miR-146b-5p ovary control temperature (OCT) versus testis control temperature (TCT) and ovary high temperature (OHT) versus OCT, dre-miR-21-5p OHT versus OCT, dre-miR-726-3p OCT versus TCT, dre-miR-726-5p THT versus TCT, dre-miR-92a-3p OHT versus OCT, dre-miR-202-5p OHT versus OCT, dre-miR-143-3p THT versus TCT and, dre-miR-122-5p THT versus TCT.
Bioinformatics: miRNA mapping and annotations
Sequenced libraries were analyzed by Prost! as described by Desvignes et al18. Briefly, sequencing data were trimmed and reads were mapped on the reference genome version 11 of zebrafish (GRCz11) for annotations and to distinguish known and novel miRNAs. For novel miRNAs not fully annotated against the zebrafish genome the FishmiRNA database was consulted30. Expression of the sequenced miRNAs was determined by the raw count matrix used as input into the R package DESeq231 (version 3.15). Read data were normalized by DESeq functions and relative expression between groups was generated by base mean, log2 fold change and adjusted P-value (P < 0.05). To visualize the level of similarity of individual samples a Multi-Dimensional Scaling (MDS) plot was created with the R32 (version 4.2) package EdgeR33 from Bioconductor34 (version 3.15). Heatmaps of DE miRNAs in ovaries and testes (HT versus CT) were constructed using the R package pheatmap (version 1.0.12) (https://CRAN.R-proect.org/package=pheatmap).
Consistent gonadal miRNAs in zebrafish
To identify miRNAs in the zebrafish gonads, miRNA data from two available publications in zebrafish was used17,18 additionally to the data currently presented. The normalized read lists were used to identify significantly expressed miRNAs in testes or ovaries with an expression of 100 normalized reads or higher. Next, DE miRNAs between ovaries and testes in the three miRNA datasets were identified (expression of 100 normalized reads or higher and adjusted P ≤ 0.05). Venn Diagrams were created using the software from the Bioinformatics & Evolutionary Genomics group from Ghent University (http://bioinformatics.psb.ugent.be/webtools/Venn/).
Functional annotation of miRNA targets
To identify miRNA targets, 3′UTR regions and genome annotation for zebrafish were extracted from Ensembl (https://www.ensembl.org/) using the Biomart data mining tool. Putative miRNA targets were predicted with MiRanda35 with energy threshold-25 and other parameters left to their default value. Subsequent MiRanda output was pruned and processed to extract relevant information with a custom Perl script (Dataset 2). Seed region alignments were inspected by hand from MiRanda’s parsed output, all of them appeared to be solid enough. The portal (https://david.ncifcrf.gov/) was used to perform enrichment analyses and search for GO terms and KEGG pathways. Graphs of a representative summary of each GO term category for each gonad class were produced with Revigo36 using term frequency as the guiding parameter. A circular zebrafish genome graph was produced with Circos37.
Fluorescent in situ hybridization
For fluorescent in situ hybridization (FISH), ovaries dissected from a total of 12 zebrafish adult females were fixed overnight in 4% paraformaldehyde (PFA) at 4 °C, dehydrated in 100% methanol and stored at − 20 °C. Fixed ovaries from the control group (OCT) were paraffin-embedded and sections (9 μm thickness) were obtained with a microtome (HM355, Microm). The anti-sense miRCURY LNA miRNA detection probe dre-miR-146b-5p (YD00613622, QIAGEN) was used. The mmu-miR-122-5p miRCURY LNA probe (YD00615338, QIAGEN) was used to detect the dre-miR-122-5p mature form, since zebrafish and mouse miR-122-5p sequences are identical. Probe sequences were 5′-CAAACACCATTGTCACACTCC-3′ and 5′- CACCCTTGGAATTCAGTTCTC-3′ to detect dre-miR-122-5p and dre-miR-146b-5p, respectively. The scramble-miR miRCURY LNA Detection probe (5′-GTGTAACACGTCTATACGCCCA-3′, YD00699004, QIAGEN) was used as a negative control. All LNA probes were double-DIG labeled at both 5′ and 3′ ends. FISH was performed using the miRCURY LNA miRNA ISH kit (FFPE, 339450, QIAGEN) following the manufacturer’s instructions, Permeabilization was performed for 7 min at room temperature using Proteinase-K (10 μg/ml, P2308 Sigma). LNA probes were used at 40 nM at 53 °C (30 °C below the RNA Tm) for 2 h. Samples were then incubated overnight at 4 °C with a rabbit anti-DIG HRP-conjugate antibody (1:500, Roche). Then, the anti-DIG-HRP antibody was detected with the TSA-Cy3 substrate (1:50, TSATM PLUS Cy3 kit, NEL 745001KT, Perkin Elmer) for 10 min at room temperature. Nuclei were stained with 4% Methyl Green (MG, 323829-5G, Sigma-Aldrich) in PBS/0.1% triton for 15 min at room temperature. All pictures were taken with a Leica TCS SP8 laser scanning confocal microscope using 552 nm and 638 lasers for TSA-cy3 and MG detection, respectively.
Ethics
Experimental procedures agreed with the European regulations of animal welfare (ETS N8 123,01/01/91) and were approved by: (1) the Ethical Committee of Consejo Superior de Investigaciones Científicas (CSIC) that evaluates projects and procedures in which animals are used for experimentation and other scientific purposes –RD 53/2013 (Spanish Ministry of Science and Innovation), and (2) the Department of Territory and Sustainability of Catalan government regulations (34, 53/2013) with the approval number 9977. The study was carried out in compliance with the ARRIVE guidelines.
Results
Sex ratio and biometry
After heat treatment during sex differentiation, a 17% masculinization was observed in the high temperature (HT) group (Supplemental Figure S1), although differences were not significant. The accumulated thermal unit (ATU) that fish were subjected during the thermal treatment were 419.84 (28 °C) and 511.25 (34 °C) for control temperature (CT) and HT, respectively. The mean weight of the animals was as follows: female CT 0.38 ± 0.09 g, male CT 0.27 ± 0.08 g, female HT 0.26 ± 0.10 g and male HT 0.24 ± 0.09 g. The mean size of the animals was as follows: female CT 2.48 ± 0.26 cm, male CT 2.48 ± 0.05 cm, female HT 2.53 ± 0.15 cm, male HT 2.35 ± 0.10 cm. No significance was found in either female or male CT versus HT in weight or size. The weights, lengths and K-factor and statistical results of all 16 fish can be found in Supplemental Table S1.
miRNA sequencing overview and validation
On average, we obtained 25.3 million sequences per library and the total number of sequences exceeded 101 million, 71 and 30 million for testes and ovaries, respectively. The length distribution showed that over 57.9% of the obtained sequences were 22 nucleotides (nts), and the overall length distribution had a range between 18 and 25 nts (Supplemental Figure S2). A total of 359 mature miRNAs were identified after alignments against the zebrafish genome (Dataset 3). Eleven miRNAs were not fully annotated with Prost!, of which 9 miRNAs were identified using FishmiRNA30 and two miRNAs were aligned to the genome of Gasterosteus aculeatus with their Ensembl Gene ID. Only two miRNAs were not annotated and were defined as novel. Thus, 99.4% of the miRNA sequenced was annotated against the zebrafish genome. The datasets generated and analyzed during the current study are available in NCBI SRA repository with the accession number: PRJNA755482.
The MDS analyses clustered the ovarian samples based on the treatment while in testes the sample clustering was not as clear (Supplemental Figure S3A and B). The two MDS components explained 59% and 62% for ovaries and testes, respectively of the variance among the samples. There was one testicular control sample (i.e., TCT3) that was clustered individually from the other samples, but was not discarded from further analyses.
miRNA-seq data was validated by testing the expression of eight miRNAs in a total of nine different comparisons among groups (dre-miR-146b-5p, dre-miR-21-5p, dre-miR-726-3p, dre-miR-726-5p, dre-miR-92a-3p, dre-miR-202-5p, dre-miR-143-3p, dre-miR-122-5p) by qPCR analyses in the ovary and testis in three different comparisons based on their expression in sequencing data. Results showed a linear regression with R2 = 0.9287 and P = 2.9e−5, thus validating miRNA sequencing results (Supplemental Figure S4).
miRNAs in the zebrafish gonads
Data from two similar studies, one from the same zebrafish strain18 and one from a different zebrafish line (crossing nacre transparent, −/−, with zf45Tg17) were used in order to identify miRNAs that were consistently expressed in the zebrafish gonads within one given sex.
Comparing our miRNA data with the two available libraries, we found 32 and 50 miRNAs in ovary and testis, respectively, specific for our data. A total of 131 and 137 miRNAs in the ovary and the testis, respectively, were found between the results reported by Desvignes et al.18 and our present data while only 37 and 34 were common between our data and the results of Presslauer et al.17 (Fig. 1A,B). Between all three datasets, 35 and 32 common miRNAs in ovaries and testes were found, respectively (Fig. 1A,B and Supplemental Table S2). A total of 25, 14 and 20 DE miRNAs in ovary and 16, 3 and 26 in testis for Presslauer et al.17, Desvignes et al.18 and our data, respectively, were identified as unique for each of the three publications (Supplemental Figure S5A, B). Comparing DE miRNAs between ovary and testis in the three studied data, identified 1 common miRNA for each studied tissue (Supplemental Figure S5A, B), dre-miR-200b-3p in ovary and dre-miR-212-5p in testis. Since our and Desvignes et al.18 data used the same zebrafish AB strain, we selected those common DE miRNAs between ovary and testis (8 and 11, respectively) to plot a heatmap (Fig. 1C) that showed those miRNAs that were constitutively differentially expressed between both sexes.

The number of miRNAs that were significantly expressed (average normalized reads > 100) of two AB strains (current data and Desvignes et al.18) and zf45Tg hybrid zebrafish (Presslauer et al.47). (A) In ovaries, and (B) testes. (C) Heatmap of differentially expressed miRNAs between ovary and testis commonly found in Desvignes et al.18 and present data.
miRNAs sensitive to temperature in the gonads
One miRNA was found to be significantly upregulated (adjusted P-value ≤ 0.05) in testis between CT and HT groups, i.e., dre-miR-122-5p, and 23 miRNAs were differentially regulated in the ovary (adjusted P-value ≤ 0.05, Fig. 2) (Supplemental Table S3), giving that 24 DE miRNAs were altered due to the heat in the zebrafish gonads. The five top upregulated miRNAs in the ovary were dre-miR-499-5p, dre-miR-202-5p, dre-miR-92b-3p, dre-miR-454b-3p and, dre-miR-725b-5p. The most downregulated were dre-miR-726-5p, dre-miR-184-3p, dre-miR-146b-5p, dre-miR-34a-5p and, dre-miR-132-3p (Supplemental Figure S6). Overall, the temperature-strongly repressed the expression of those thermal sensitive miRNAs when compared to those miRNAs that were upregulated, for example, the highest fold change was over six-fold for the inhibition of the dre-miR-725b-5p expression whereas was over two-fold for the upregulation of the dre-miR-499.

Heatmap of 23 differentially expressed (DE) miRNAs in mature ovaries after exposing zebrafish to high temperature during sex differentiation. The color scale ranges from blue to red, where blue shows relative overexpression and red is relative under expression. Both miRNAs and samples were grouped by hierarchical clustering.
miRNA target predictions and functional annotation
To inspect the biological roles of the identified 24 DE miRNAs after high-temperature treatments in the zebrafish gonads, target genes of the 24 miRNAs were predicted upon the zebrafish genome by 3′-UTRs. Most of the DE miRNAs had multiple target genes and many of them were regulated by more than one miRNA. We predicted 1205 and 101 target genes for ovary and testis, respectively, by the control groups. In ovary, 407 unique targets were found for the 23 DE miRNAs whereas in testis, 85 unique targets were found for dre-miR-122-5p. The full list of the predicted targets is shown in Supplemental Table S4.
To better understand the relationship between DE miRNAs and their function in the gonads after heat exposure, GO enrichment analyses of the putative target genes were performed (Supplemental Table S5). In ovary, 54 GO terms for Biological process (BP), 27 for Cellular component (CC), and 42 for Molecular function (MF) were predicted and 3 GO terms for BP, 4 for CC, and 3 for MF in testis. In ovary, some of the most enriched GO terms for BP were: regulation of transcription, transport and phosphorylation (Fig. 3); for CC were: membrane, integral component of membrane and extracellular region (Supplemental Figure S7A), and for MF were: metal ion binding, zinc ion binding, and transferase activity (Supplemental Figure S7B).

Visual representation of Gene Ontology (GO) terms related to Biological process obtained from predicted target genes of differentially expressed miRNAs in ovary. The most frequent terms were regulation of transcription, transport and phosphorylation. Color intensity represents the uniqueness of the GO term. Plot_size shows the frequency of the GO term in the UniProt database. The GO terms with a dispensability of < 0.25 are annotated in the plot.
Synteny of the target genes
A synteny map indicated the widespread distribution of the 24 DE miRNAs (23 in ovaries, 1 in testis) in the zebrafish genome (Fig. 4). In the ovary, the spatial distribution of the 407 target genes in which DE miRNAs interacted was mostly localized in chromosomes 7, 2, 4, 3 and 11 (Fig. 5A). These five chromosomes contained 54.5% of the predicted target genes. 16 DE miRNAs are targeting genes in chromosome 4 (Supplemental Table S6), some of which were related to the reproductive system (e.g. SRY-box transcription factor 5, sox5, RAS like estrogen-regulated growth inhibitor, rerg) or the immune system (interleukin 15 receptor subunit alpha, ilr15β, B-cell translocation gene 1, btg1). In testis, Fig. 5B shows the top 15 chromosomes in which the 85 predicted genes were localized. The chromosomes 14, 2, 7, 15, 1 contained 35.3% of the predicted genes.

Circular localization of predicted target genes from differentially expressed (DE) miRNAs in the zebrafish genome. 407 predicted target genes of DE miRNAs in the ovary (purple) and 85 predicted target genes in the testis (green) were distributed throughout the zebrafish genome, with the highest percentage present in chromosomes 7 and 14 for ovary and testis, respectively.

Chromosomal distribution in the zebrafish genome of the number of predicted target genes obtained from differentially expressed miRNAs after high temperature in the ovary (A) and testis (B) in adult zebrafish.
Spatial expression of selected miRNAs in the gonads
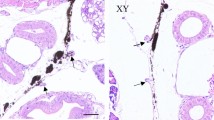
To better understand the functionality of the DE miRNAs in the gonads, the cellular localization of two DE miRNAs was performed by FISH. dre-miR-146b-5p was selected since it was DE in both OHT versus OCT and OCT versus TCT in the present data, as well as DE in ovary versus testis in the data from Presslauer et al17 Further, this miRNA presented one of the highest fold change (Supplementary Figure S6) and high abundance in the RNA sequencing. Moreover, to our knowledge, no available literature on its presence in the gonads have been described to date. dre-miR-122-5p was selected for being the only DE miRNA in testes after high temperature. In ovaries, dre-miR-146b-5p and dre-miR-122-5p were detected in the germ cells with an expression that was inversely proportional to oocyte maturation (Figs. 6, 7), detecting expression in small oocytes likely corresponding to pre-vitellogenic and early-vitellogenic oocytes, whereas no expression was detected in large oocytes, including late and post-vitellogenic oocytes. No signal was detected in any of the follicular cells of the ovary. In testis, the signal was not detected for any of the probes used, neither in germ nor follicular cells (data not shown). No signal was detected in negative controls, where scramble miRNA probes were used.

Fluorescent in situ hybridization (FISH) of dre-miR-146b-5p in the ovary of adult zebrafish. A total of 6 female fish were used to obtain the results. (A) Sections A and A’ showed scramble probe. Section B, B′ and B″ showed the localization of miR-146b in germ cells. Size bar = 100 µm.

Fluorescent in situ hybridization (FISH) of dre-miR-122-5p in the ovary of adult zebrafish. A total of 6 female fish were used to obtain the results. Sections A and A′ showed scramble probe. Section B, B′ and B″ showed the localization of dre-miR-122-5p in germ cells. (V) Vitellogenic oocyte, (LV) Late vitellogenic oocyte. A and B scale bar = 500 µm and A′, B′, B″ size bar = 100 µm.
Discussion
Temperature increase influences sexual development by skewing sex ratios towards males in fish5. The study of the underlying epigenetic mechanisms of this masculinization has relied on DNA methylation analyses in the gonads of some fish species, like European sea bass6, half-smooth tongue sole8, tilapia38, fugu7 and zebrafish9. However, other epigenetic mechanisms, and specifically translation interference by miRNAs, have not yet been elucidated. Here, gonadal data is described on miRNAs affected by changes in temperature during early development that likely play a role in the final sexual phenotype in zebrafish.
To date, available reports in zebrafish show high variability on the sex ratio changes in zebrafish subjected to high temperatures (from 22 to 60% masculinization)39, as zebrafish present interfamily variation due to the genetic and environmental influences on the final sexual phenotype40. In the current study, a 17% difference of masculinization was observed from 70% at low temperature to 87% at high temperature, although not significant. Non-significance might be explained by a low number of biological replicates or/and by the genetic factor of the skewed sex ratios towards males of the family used (i.e. 70%)41. To induce a significant masculinization, higher temperatures (i.e. 36ºC) can be performed but in contrast, few or no female samples would have been obtained9,27. Thus, the experimental approach confirmed the successful implementation of heat treatment as well as validating previously reported results.
Here, we reported a total of 24 DE miRNAs in the zebrafish gonads two months after heat exposure during fifteen days of early development when gonads are differentiating. In a similar study in Atlantic cod, embryos were incubated at high temperatures resulting in alterations of some, but few, miRNAs in juvenile animals in different tissues, including gonads, although the sexual dimorphic difference was not studied25. Thus, the alteration of the miRNA expression due to environmental cues indicates that they can be considered as heat recorders as their expression depends on past events. Only one out of the 24 miRNAs altered by elevated temperatures is testis specific. By using the same experimental approach in zebrafish in Ribas et al.27, testicular transcriptome presented no DE genes after the heat exposition during sex differentiation when compared to the control, revealing that in testes, of some certain neomales, no relevant transcriptomic differences after the heat treatment was presented27. Nevertheless, in the same study, another neomale population in the heated group showed a larger amount of DE genes (~ 700) when compared to the control. In the ovarian transcriptomes, only 20 DE genes were found when compared to the control but a larger number of DE genes were found (~ 9650) when compared to so-called pseudofemales (females with phenotypic ovaries and with a male-transcriptomic profile). Overall, when comparing the overviewed number of DE miRNAs and the DE genes obtained from both studies in the zebrafish gonads treated with high temperature during sex differentiation, the alterations in the miRNome and the transcriptomes were more severe in the ovaries, probably due to the fact that in the adult female fish, ovaries needed to resist the sex-reversal process while some adult males were already sex-reversed females.
Here, twelve of the miRNAs were upregulated in the adult ovaries of the heat-treated fish, among them, dre-miR-202-5p. Emerging evidence suggests that this miRNA is highly expressed in female gonads of many animals, e.g., fish25, frogs42 and goats43 and it has even been proposed as a marker of fish fecundity and fertility44. miRNA-202 was found in testes in both Sertoli cells and spermatogonia stem cells in mammals45 and in a similar manner than in rainbow trout16and medaka44. Another miRNA that was upregulated by heat treatment was dre-miR-92a, which has been found as the most abundant miRNA in zebrafish gonads17. This miRNA was responsible for cell cycle progression during the early stage of embryo development and metamorphosis in both Japanese flounder (Paralichthys olivaceus)46 and in zebrafish47.
The expression of eleven miRNAs was downregulated due to the temperature increase, for example, dre-miRNA-21-5p. This miRNA is highly conserved throughout evolution and abundantly distributed in many tissues in fish. This is the case of the heart48, kidney49, and ovary50. It was also linked to the fish immune response through the TLR28 signaling pathway51. Many functions have been related to miRNA-21 in humans as being found in different cancers, although in fish, fewer data of its biological role are available. Strikingly, most of the miRNAs here identified as heat recorders, are related to ovarian and prostate cancer in humans, either promoting or suppressing cancer progression and thus much literature related to these diseases is available. This is the case of, for example, miR-19b52, miR-15b53, and miR-45454 three upregulated miRNAs in the fish ovaries after the heat and; miR-27b55, miR-21256, miR-146b57, and miR-34a58, which were downregulated. The emerging research on miRNAs has flourished the utility of miRNAs as bioclinical markers in human cancers during the last decade59 but also as attractive drug targets for human diseases with no current effective treatments60. This has attracted the attention of many pharmaceutical companies which are developing clinical trials, such as, miRNA-21 and mRNA-92 which are in phase 2 and 1, respectively61 and interestingly, were found down- and upregulated, respectively, in the ovaries after the heat treatment in the present study. Thus, the exploration of the usefulness of miRNAs as heat markers becomes attractive as a potential method to predict animals with different susceptibilities to environmental cues and might be explored to be transfered as markers for aquaculture purposes.
Predicted target genes from the DE miRNAs due to exposure to elevated temperatures, showed functions related to reproduction and sex. This is the case of Polycomb Group RING Finger (pcgf6) gene-targeted by dre-miR-458-3p62; pcgf5a gene targeted by dre-miR-184-3p63, and Dickkopf-related protein (dkk1b) targeted by dre-miR-212-5p64. Similarly, those miRNAs downregulated in the ovaries after the heat targeted to reproduction-related genes such as sox5 targeted by dre-miR-15b-1-5p65, and Nuclear Receptor Subfamily 5 Group A Member 2 (nr5a2) targeted by dre-miR-19b-3p66. In the testes, only one miRNA was identified as heat recorder, miR-146b and targeted, for example, to BCL2 Binding Component 3 (bbc3) gene which is related to prostate cancer67, and Tet methylcytosine dioxygenase 2 (tet2), a demethylator of many genes, included the SRY, a key gene in the regulation of male sex determination in mammals68. Overall results confirmed that the miRNA machinery was active and essential to regulate the environmental cues that occur in the adult fish gonads.
The synteny of the predicted target genes of the heat recorders miRNAs on the zebrafish genome showed multiple regions in all the 25 chromosome pairs, but more abundantly in chromosomes 7, 2, 4, 3 and 11 in the ovary, accounting for 54.5% of the predicted target genes in the present data. To foster the identification of sex-determining gene(s) in this popular animal model, many sex genetic studies in the last decade have been performed by crossing natural and domesticated zebrafish strains. By single nucleotide polymorphisms (SNPs) and sequence-based polymorphic restriction site associated (RAD-tag) strategies, several sex-linked loci in the chromosomes 4 and 3 have been identified69,70 and in the chromosomes 5 and 1671. Strikingly, in chromosome 4, the sex-association region (sar) was localized in wild zebrafish strains72, a chromosome that from our data supported more than 10,5% of the predicted target genes of the miRNAs sensitive to heat. Furthermore, chromosome 16 accounted ~ 7.5% of our predicted target genes. Thus, although more research is required to understand the biological functions of the present data, we can ascertain some of the chromosomes that host genes regulated by miRNAs sensitive to heat.
Further, we identified sexual dimorphism in the expression of miRNAs in the fish gonads with a total of 45 and 54 up- and downregulated, respectively, in the ovary when compared to testis. To increase consistency, our data were compared with two available data of the same species resulting in common miRNAs. We found that miRNA-200b-3p was upregulated in the ovary in the three zebrafish comparisons. The role of this miRNA is not fully understood but it is known to be involved in many human cancers: kidney73, prostate74, and breast75. It is highly released in the serum of the anovulatory women diagnosed with polycystic ovary syndrome and suggested as a clinical marker76. miR-212-5p was DE in the testes versus ovaries in the three zebrafish gonadal miRNA datasets and inhibited after the heat treatment in the ovaries. This can indicate its role by dysregulation of ovarian functions during the masculinization event occurred by heat. The miRNA-212 function is not stated but in humans, it is related to cell proliferation and angiogenesis and is present in the brain and gonads77. In addition, miRNA-212 was found in tilapia gonads78, which, together with the current results, show its relevance presence in the reproduction system in fish.
The gonadal localization of two miRNAs, dre-miR-122-5p and dre-miR-146b-5p, showed similar results. In ovary, their expressions were found in the germ cells but not in the granulosa or theca cells while fluorescent intensity was stronger in less mature oogonial cells, suggesting a potential role of these miRNAs in germ cell development. In testis, although miR-122 is involved in zebrafish sperm quality79 and male fertility in mammals80, its localization, together with that for miR-146b, was not possible in the zebrafish testicular cells, so more sensitive methods need to be readied. miR-122 is involved in humans in many cancer and has reached phase II in clinical trials for treating hepatitis60. In fish, much literature related to miR-122 is available certifying the role in the immune81 and in the metabolic systems being highly abundant in the zebrafish liver82. The presence of miR-122 was detailed in many fish species such as tilapia, medaka, carp, and in many fish tissues such as the spleen, head kidney 83. In the gonads, it was detected in mature sharpsnout seabream (Diplodus puntazzo) but not in the marine medaka84. Strikingly, miRNA122 was reported to be sensitive to cold temperatures in the Senegalese sole (Solea senegalensis)24, thus the role of this miRNA as a thermal recorder is worth further exploring. Regarding miR-146b in humans, it plays a role in the innate immune response85 and is involved in gliomas and ovarian cancers86. In fish, very little data is available but it was upregulated in response to infection in zebrafish embryos87 and spleen88 and the sperm of growth hormone (GH)-transgenic zebrafish79. Overall, to our knowledge, this is the first time that the cellular localization of these two miRNAs are described in the gonads.
Conclusions
Present data evidence that high temperature alters the miRNome in the fish gonads. The influence of heat treatment during gonadal development altered the expression of 23 miRNAs in the ovaries, by enhancing, for example, miR-92b-3p and miR-202-5p, or repressing, for example, miR-212-5p and miR-146b-3p expressions. In testes, miR-122-5p was the only miRNA sensitive to heat. These miRNAs act as heat recorders and might be potential targets for developing predictive tools of heat response, essential in a climate change scenario or to increase productivity from sustainability. In addition, as most of the 24 DE miRNA have been found to be involved in different diseases, mostly related to cancer, the data here might be helpful to enhance our knowledge on the functional roles of these miRNAs in fish-related diseases.
Data availability
The datasets generated and analyzed during the current study are available in NCBI SRA repository with the accession number: PRJNA755482.
Change history
28 February 2024
A Correction to this paper has been published: https://doi.org/10.1038/s41598-024-55491-x
References
Dangendorf, S. et al. Persistent acceleration in global sea-level rise since the 1960s. Nat. Clim. Change 9, 705–710 (2019).
Liu, H. et al. Sexual plasticity: A fishy tale. Mol. Reprod. Dev. 84, 171–194 (2017).
Devlin, R. H. & Nagahama, Y. Sex determination and sex differentiation in fish: An overview of genetic, physiological, and environmental influences. Aquaculture 208, 191–364 (2002).
Wang, H.-P. & Shen, Z.-G. Sex control in aquaculture. In Sex Control in Aquaculture 1–34 (Wiley, 2018). https://doi.org/10.1002/9781119127291.ch1.
Ospina-Alvarez, N. & Piferrer, F. Temperature-dependent sex determination in fish revisited: Prevalence, a single sex ratio response pattern, and possible effects of climate change. PLoS ONE 3, e2837 (2008).
Navarro-Martín, L. et al. DNA methylation of the gonadal aromatase (cyp19a) promoter is involved in temperature-dependent sex ratio shifts in the European sea bass. PLoS Genet. 7, e1002447 (2011).
Zhou, H. et al. Changes in DNA methylation during epigenetic-associated sex reversal under low temperature in Takifugu rubripes. PLoS ONE 14, e0221641 (2019).
Shao, C. et al. Epigenetic modification and inheritance in sexual reversal of fish. Genome Res. 24, 604–615 (2014).
Valdivieso, A., Ribas, L., Monleón-Getino, A., Orbán, L. & Piferrer, F. Exposure of zebrafish to elevated temperature induces sex ratio shifts and alterations in the testicular epigenome of unexposed offspring. Environ. Res. 186, 109601 (2020).
Iwakawa, H. O. & Tomari, Y. The functions of microRNAs: mRNA decay and translational repression. Trends Cell Biol. 25, 651–665 (2015).
Kozomara, A., Birgaoanu, M. & Griffiths-Jones, S. miRBase: From microRNA sequences to function. Nucleic Acids Res. 47, D155–D162 (2019).
Lim, L. P., Glasner, M. E., Yekta, S., Burge, C. B. & Bartel, D. P. Vertebrate microRNA genes. Science 299, 1540 (2003).
Herkenhoff, M. E. et al. Fishing into the MicroRNA transcriptome. Front. Genet. 9, 88 (2018).
Bizuayehu, T. T. et al. Sex-biased miRNA expression in atlantic halibut (Hippoglossus hippoglossus) brain and gonads. Sex. Dev. 6, 257–266 (2012).
Jing, J. et al. Sex-biased miRNAs in gonad and their potential roles for testis development in yellow catfish. PLoS ONE 9, 1–16 (2014).
Juanchich, A. et al. Characterization of an extensive rainbow trout miRNA transcriptome by next generation sequencing. BMC Genom. 17, 164 (2016).
Presslauer, C., Bizuayehu, T. T., Kopp, M., Fernandes, J. M. O. & Babiak, I. Dynamics of miRNA transcriptome during gonadal development of zebrafish. Sci. Rep. 7, 1–16 (2017).
Desvignes, T., Batzel, P., Sydes, J., Eames, B. F. & Postlethwait, J. H. miRNA analysis with Prost! reveals evolutionary conservation of organ-enriched expression and post-transcriptional modifications in three-spined stickleback and zebrafish. Sci. Rep. 9, 3913 (2019).
Ji, X. et al. Identification and characterization of miRNAs involved in cold acclimation of zebrafish ZF4 cells. PLoS ONE 15, e0226905 (2020).
Huang, J. et al. Identification and characterization of microRNAs in the liver of rainbow trout in response to heat stress by high-throughput sequencing. Gene 679, 274–281 (2018).
Zhou, C. Q., Zhou, P., Ren, Y. L., Cao, L. H. & Wang, J. L. Physiological response and miRNA-mRNA interaction analysis in the head kidney of rainbow trout exposed to acute heat stress. J. Therm. Biol. 83, 134–141 (2019).
Vasadia, D. J., Zippay, M. L. & Place, S. P. Characterization of thermally sensitive miRNAs reveals a central role of the FoxO signaling pathway in regulating the cellular stress response of an extreme stenotherm, Trematomus bernacchii. Mar. Genom. 48, 100698 (2019).
Nie, M. M. et al. Network of microRNA-transcriptional factor-mRNA in cold response of turbot Scophthalmus maximus. FISH Physiol. Biochem. 45, 583–597 (2019).
Campos, C. et al. Thermal plasticity of the miRNA transcriptome during Senegalese sole development. BMC Genom. 15, 1–13 (2014).
Bizuayehu, T. T., Johansen, S. D., Puvanendran, V., Toften, H. & Babiak, I. Temperature during early development has long-term effects on microRNA expression in Atlantic cod. BMC Genom. 16, 1–12 (2015).
Ikert, H. & Craig, P. M. Chronic exposure to venlafaxine and increased water temperature reversibly alters microRNA in zebrafish gonads (Danio rerio). Comp. Biochem. Physiol. Part D Genom. Proteom. 33, 100634 (2020).
Ribas, L. et al. Heat-induced masculinization in domesticated zebrafish is family-specific and yields a set of different gonadal transcriptomes. Proc. Natl. Acad. Sci. U. S. A. 114, E941–E950 (2017).
Ribas, L., Valdivieso, A., Díaz, N. & Piferrer, F. Appropriate rearing density in domesticated zebrafish to avoid masculinization: Links with the stress response. J. Exp. Biol. 220, 1056–1064 (2017).
Ribas, L. et al. Comprehensive transcriptomic analysis of the process of gonadal sex differentiation in the turbot (Scophthalmus maximus). Mol. Cell. Endocrinol. 422, 132–149 (2016).
Desvignes, T. et al. FishmiRNA: An evolutionarily supported microRNA annotation and expression database for ray-finned fishes. Mol. Biol. Evol. 39, msac004 (2022).
Love, M. I., Huber, W. & Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 15, 1–21 (2014).
R Core Team (2021) R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna. https://www.R-project.org
Robinson, M. D., McCarthy, D. J. & Smyth, G. K. edgeR: A bioconductor package for differential expression analysis of digital gene expression data. Bioinformatics 26, 139–140 (2009).
Gentleman, R. C. et al. Bioconductor: Open software development for computational biology and bioinformatics. Genome Biol. 5, 1–16 (2004).
Enright, A. J. et al. MicroRNA targets in drosophila. Genome Biol. 5, 1–14 (2003).
Supek, F., Bošnjak, M., Škunca, N. & Šmuc, T. Revigo summarizes and visualizes long lists of gene ontology terms. PLoS ONE 6, e21800 (2011).
Krzywinski, M. et al. Circos: An information aesthetic for comparative genomics. Genome Res. 19, 1639–1645 (2009).
Wang, J. et al. Transcriptomic and epigenomic alterations of Nile tilapia gonads sexually reversed by high temperature. Aquaculture 508, 167–177 (2019).
Abozaid, H., Wessels, S. & Hörstgen-Schwark, G. Effect of rearing temperatures during embryonic development on the phenotypic sex in zebrafish (Danio rerio). Sex. Dev. 5, 259–265 (2011).
Ribas, L., Valdivieso, A., Diaz, N. & Piferrer, F. Response to ‘the importance of controlling genetic variation—remarks on “appropriate rearing density in domesticated zebrafish to avoid masculinization: Links with the stress response”’. J. Exp. Biol. 220, 4079–4080 (2017).
Liew, W. C. et al. Polygenic sex determination system in zebrafish. PLoS ONE 7, e34397 (2012).
Michalak, P. & Malone, J. H. Testis-derived microRNA profiles of African clawed frogs (Xenopus) and their sterile hybrids. Genomics 91, 158–164 (2008).
Ding, Q. et al. Transactivation of miR-202–5p by steroidogenic factor 1 (SF1) induces apoptosis in goat granulosa cells by targeting TGF beta R2. Cells 9, 445 (2020).
Gay, S. et al. MiR-202 controls female fecundity by regulating medaka oogenesis. PLOS Genet. 14, e1007593 (2018).
Gu, W. et al. MicroRNA-22 regulates inflammation and angiogenesis via targeting VE-cadherin. FEBS Lett. 591, 513–526 (2017).
Li, X. M. et al. The role of miR-92 in regulating early development and metamorphosis of Japanese flounder Paralichthys olivaceus. Genes Genet. Syst. 95, 1–10 (2020).
Presslauer, C., Bizuayehu, T., Fernandes, J. & Babiak, I. miR-92a-3p controls cell cycle progression in zebrafish. Zebrafish https://doi.org/10.1101/680991 (2019).
Kolpa, H. J. et al. miR-21 represses Pdcd4 during cardiac valvulogenesis. Development 140, 2172–2180 (2013).
Hoppe, B. et al. MiR-21 is required for efficient kidney regeneration in fish. BMC Dev. Biol. 15, 1–10 (2015).
Wong, Q. W. L. et al. Identification and characterization of a specific 13-miRNA expression signature during follicle activation in the zebrafish ovary. Biol. Reprod. 98, 42–53 (2018).
Bi, D., Cui, J., Chu, Q. & Xu, T. MicroRNA-21 contributes to suppress cytokines production by targeting TLR28 in teleost fish. Mol. Immunol. 83, 107–114 (2017).
Zhong, Z. et al. Inhibition of microRNA-19b promotes ovarian granulosa cell proliferation by targeting IGF-1 in polycystic ovary syndrome. Mol. Med. Rep. 17, 4889–4898 (2018).
MacLean, J. A., King, M. L., Okuda, H. & Hayashi, K. WNT7A Regulation by miR-15b in ovarian cancer. PLoS ONE 11, e0156109 (2016).
An, Y. et al. MiR-454 suppresses the proliferation and invasion of ovarian cancer by targeting E2F6. Cancer Cell Int. 20, 237 (2020).
Liu, W., Lv, C., Zhang, B., Zhou, Q. & Cao, Z. MicroRNA-27b functions as a new inhibitor of ovarian cancer-mediated vasculogenic mimicry through suppression of VE-cadherin expression. RNA 23, 1019–1027 (2017).
Zhang, L. et al. MiR-212-3p suppresses high-grade serous ovarian cancer progression by directly targeting MAP3K3. Am. J. Transl. Res. 12, 875–888 (2020).
Yan, M. et al. miR-146b promotes cell proliferation and increases chemosensitivity, but attenuates cell migration and invasion via FBXL10 in ovarian cancer. Cell Death Dis. 9, 1–17 (2018).
Schmid, G. et al. Expression and promotor hypermethylation of miR-34a in the various histological subtypes of ovarian cancer. BMC Cancer 16, 102 (2016).
Rabinowits, G., Gercel-Taylor, C., Day, J. M., Taylor, D. D. & Kloecker, G. H. Exosomal MicroRNA: A diagnostic marker for lung cancer. Clin. Lung Cancer 10, 42–46 (2009).
Rupaimoole, R. & Slack, F. J. MicroRNA therapeutics: Towards a new era for the management of cancer and other diseases. Nat. Rev. Drug Discov. 16, 203–222 (2017).
Konoshenko, M. Y., Bryzgunova, O. E. & Laktionov, P. P. miRNAs and androgen deprivation therapy for prostate cancer. Biochim. Biophys. ACTA-Rev. Cancer 1876, 188625 (2021).
Endoh, M. et al. PCGF6-PRC1 suppresses premature differentiation of mouse embryonic stem cells by regulating germ cell-related genes. Elife 6, e21064 (2017).
Le Faou, P., Völkel, P. & Angrand, P.-O. The zebrafish genes encoding the polycomb repressive complex (PRC) 1. Gene 475, 10–21 (2011).
Sreenivasan, R. et al. Gonad differentiation in zebrafish is regulated by the canonical Wnt signaling pathway. Biol. Reprod. 90, 45–46 (2014).
Schartl, M. et al. Sox5 is involved in germ-cell regulation and sex determination in medaka following co-option of nested transposable elements. BMC Biol. 16, 1–17 (2018).
Wang, L. et al. Molecular cloning and sexually dimorphic expression patterns of nr0b1 and nr5a2 in olive flounder, Paralichthys olivaceus. Dev. Genes Evol. 225, 95–104 (2015).
Dey, P., Ström, A. & Gustafsson, J. A. Estrogen receptor β upregulates FOXO3a and causes induction of apoptosis through PUMA in prostate cancer. Oncogene 33, 4213–4225 (2014).
Okashita, N., Kuroki, S., Maeda, R. & Tachibana, M. TET2 catalyzes active DNA demethylation of the Sry promoter and enhances its expression. Sci. Rep. 9, 1–11 (2019).
Anderson, J. L. et al. Multiple sex-associated regions and a putative sex chromosome in Zebrafish revealed by RAD mapping and population genomics. PLoS ONE 7, 40701 (2012).
Bradley, K. M. et al. An SNP-based linkage map for Zebrafish reveals sex determination loci. G3 Genes Genom. Genet. 1, 3 (2011).
Howe, K. et al. The zebrafish reference genome sequence and its relationship to the human genome. Nature 496, 498–503 (2013).
Wilson, C. A. et al. Wild sex in Zebrafish: Loss of the natural sex determinant in domesticated strains. Genetics 198, 1291 (2014).
Tang, O., Chen, X.-M., Shen, S., Hahn, M. & Pollock, C. A. MiRNA-200b represses transforming growth factor-beta 1-induced EMT and fibronectin expression in kidney proximal tubular cells. Am. J. Physiol. Physiol. 304, F1266–F1273 (2013).
Brase, J. C. et al. Circulating miRNAs are correlated with tumor progression in prostate cancer. Int. J. Cancer 128, 608–616 (2011).
Wang, Z., Humphries, B., Li, Y. & Yang, C. Abstract P6-01-09: MiRNA-200b suppresses triple negative breast cancer metastasis by targeting ARHGAP18 and causing sustained Rho A activation. in Poster Session Abstracts vols 77 MA-P P6-01-09-P6-01-09 (American Association for Cancer Research, 2017).
Eisenberg, I. et al. Elevated circulating micro-ribonucleic acid (miRNA)-200b and miRNA-429 levels in anovulatory women. Fertil. Steril. 107, 269–275 (2017).
Lei, Z. et al. Control of angiogenesis via a VHL/miR-212/132 axis. Cells 9, 1017 (2020).
Tao, W. et al. Integrated analysis of miRNA and mRNA expression profiles in tilapia gonads at an early stage of sex differentiation. BMC Genom. 17, 328 (2016).
Domingues, W. B. et al. GH overexpression alters spermatic cells microRNAome profile in transgenic Zebrafish. Front. Genet. 12, 704778 (2021).
Yu, Z., Raabe, T. & Hecht, N. B. MicroRNA Mirn122a reduces expression of the posttranscriptionally regulated germ cell transition protein 2 (Tnp2) messenger RNA (mRNA) by mRNA cleavage. Biol. Reprod. 73, 427–433 (2005).
Cui, J. X., Chu, Q. & Xu, T. J. miR-122 involved in the regulation of toll-like receptor signaling pathway after Vibrio anguillarum infection by targeting TLR14 in miiuy croaker. Fish Shellfish Immunol. 58, 67–72 (2016).
Cohen, A. & Smith, Y. Estrogen regulation of microRNAs, target genes, and microRNA expression associated with vitellogenesis in the Zebrafish. Zebrafish 11, 462–478 (2014).
Chen, J. Q. et al. Whole transcriptome-based miRNA-mRNA network analysis revealed the mechanism of inflammation-immunosuppressive damage caused by cadmium in common carp spleens. Sci. Total Environ. 717, 137081 (2020).
Papadaki, M., Kaitetzidou, E., Mylonas, C. C. & Sarropoulou, E. Non-coding RNA expression patterns of two different teleost gonad maturation stages. Mar. Biotechnol. 22, 683–695 (2020).
Taganov, K. D., Boldin, M. P., Chang, K.-J. & Baltimore, D. NF-kappa B-dependent induction of microRNA miR-146, an inhibitor targeted to signaling proteins of innate immune responses. Proc. Natl. Acad. Sci. U. S. A. 103, 12481–12486 (2006).
Katakowski, M. et al. Exosomes from marrow stromal cells expressing miR-146b inhibit glioma growth. Cancer Lett. 335, 201–204 (2013).
Ordas, A. et al. MicroRNA-146 function in the innate immune transcriptome response of zebrafish embryos to Salmonella typhimurium infection. BMC Genom. 14, 696 (2013).
Liyanage, T. D., Nikapitiya, C., Lee, J. & De Zoysa, M. Molecular insight into regulation of miRNAs in the spleen of zebrafish (Danio rerio) upon pathogenic Streptococcus parauberis infection. FISH Shellfish Immunol. 106, 898–909 (2020).
Acknowledgements
This study was supported by the Spanish Ministry of Science and Innovation grant AGL2015-73864-JIN “Ambisex” and 2PID2020-113781RB-I00 “MicroMet” and by the Consejo Superior de Investigaciones Científicas (CSIC) grant 02030E004 “Interomics” to LR, by grant AEI grant PID2019-108888RB-I00 to FP and with funding from the Spanish government through the ‘Severo Ochoa Centre of Excellence’ accreditation (CEX2019-000928-S). We thank the lab technician Sílvia Joly for her essential assistance in our team, Alejandro Valdivieso for the treatments of the zebrafish, the master student Irene Santisteban for helping with some miRNA analyses and Gemma Fusté for her assistance in fish facilities.
Author information
Authors and Affiliations
Contributions
T.G. performed the data analysis, interpreted the results and wrote the article. J.M., J.A. and J.B. performed the data analysis. V.T. performed FISH analysis. F.P. conceived the study. L.R. conceived the study, interpreted the results and wrote the article. All authors read and approved the submitted article.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
The original online version of this Article was revised: The original version of this Article omitted an affiliation for Tosca A. van Gelderen. Their correct affiliations are listed in the correction notice.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
van Gelderen, T.A., Montfort, J., Álvarez-Dios, J.A. et al. Deciphering sex-specific miRNAs as heat-recorders in zebrafish. Sci Rep 12, 18722 (2022). https://doi.org/10.1038/s41598-022-21864-3
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-022-21864-3
This article is cited by
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
