
Abstract
Abrupt onset of severe neuropsychiatric symptoms including obsessive–compulsive disorder, tics, anxiety, mood swings, irritability, and restricted eating is described in children with Pediatric Acute-Onset Neuropsychiatric Syndrome (PANS). Symptom onset is often temporally associated with infections, suggesting an underlying autoimmune/autoinflammatory etiology, although direct evidence is often lacking. The pathological mechanisms are likely heterogeneous, but we hypothesize convergence on one or more biological pathways. Consequently, we conducted whole exome sequencing (WES) on a U.S. cohort of 386 cases, and whole genome sequencing (WGS) on ten cases from the European Union who were selected because of severe PANS. We focused on identifying potentially deleterious genetic variants that were de novo or ultra-rare (MAF) < 0.001. Candidate mutations were found in 11 genes (PPM1D, SGCE, PLCG2, NLRC4, CACNA1B, SHANK3, CHK2, GRIN2A, RAG1, GABRG2, and SYNGAP1) in 21 cases, which included two or more unrelated subjects with ultra-rare variants in four genes. These genes converge into two broad functional categories. One regulates peripheral immune responses and microglia (PPM1D, CHK2, NLRC4, RAG1, PLCG2). The other is expressed primarily at neuronal synapses (SHANK3, SYNGAP1, GRIN2A, GABRG2, CACNA1B, SGCE). Mutations in these neuronal genes are also described in autism spectrum disorder and myoclonus-dystonia. In fact, 12/21 cases developed PANS superimposed on a preexisting neurodevelopmental disorder. Genes in both categories are also highly expressed in the enteric nervous system and the choroid plexus. Thus, genetic variation in PANS candidate genes may function by disrupting peripheral and central immune functions, neurotransmission, and/or the blood-CSF/brain barriers following stressors such as infection.
Similar content being viewed by others


Introduction
The acute onset of severe neuropsychiatric symptoms and abrupt loss of function in a child is an unusual and dramatic event. A clinical syndrome, Pediatric Acute Onset Neuropsychiatric Syndrome (PANS), includes a multitude of symptoms (Table 1)1. These fluctuate over time, and the onset, as well as “flares” are frequently in association with infections, which has led to speculations of autoimmune or autoinflammatory mechanisms2,3. Standard psychiatric care is often unable to sufficiently control the severe neuropsychiatric symptoms and, according to the Stanford PANS Clinic, the median Caregiver Burden Inventory score during a 1st PANS flare is higher than for those caring for someone with Alzheimer disease4. While there is a general recognition of the condition, a great deal of controversy remains concerning the validity of the PANS grouping with regards to common pathogenetic mechanisms and optimal treatment.
Complicating the matter, many patients with PANS have comorbid or a family history of neurodevelopmental and/or neuropsychiatric disorders. While some data suggests autoantibodies to neuronal antigens and elevated levels of inflammatory cytokines in PANS, there are inconsistencies5,6,7. Imaging studies in some PANS cases show local inflammation in the thalamus, basal ganglia, and amygdala, supporting a neuroinflammatory or autoimmune etiology8. Several reports indicate that patients may respond to immunomodulators, such as non-steroidal anti-inflammatory drugs (NSAIDs), intravenous immunoglobulin (IVIg), corticosteroids, and more recently, the B-cell inhibitor Rituximab5,9,10. However, there are few large, well-controlled clinical trials, with variable results9,10. Reluctance to administer immunomodulators remains due to these issues and the lack of consistent, objective biomarkers. Although most children with PANS show significant improvement at follow-up, according to a recently published longitudinal study, full remission was rare, and more than one-third were classified as having a chronic course11.
Heterogeneity on several fronts is a plausible explanation for the inconsistent findings regarding autoantibodies, inflammatory markers, and the relationship to infectious pathogens. Streptococcus pyogenes, Borrelia burgdorferi, and influenza virus are examples of potential autoimmune or proinflammatory triggers4,5,6,7. Non-infectious environmental factors that activate innate immune pathways (i.e., sterile or non-infectious inflammation), such as oxidative stress, toxin exposure, or emotional stress, could initiate abnormal inflammatory responses in genetically susceptible children. Finally, heterogeneity in genetic risk factors that cause dysregulation of peripheral immunity, and/or central neuronal/innate immune pathways, could be at play, all leading to a common clinical phenotype. In this model, some genetic subgroups will express classic markers of inflammation or autoantibodies, but others won’t. However, gene discovery in PANS that can test this model is still in its infancy.
Whole exome sequencing (WES) and whole genome sequencing (WGS) are gene discovery tools used by researchers, and increasingly by clinicians, to identify ultra-rare, biologically powerful genetic factors underlying disease states. However, such studies have not yet been reported in PANS. We now describe the discovery of ultra-rare variants in 11 genes in 21 PANS cases using these sequencing strategies.
Results
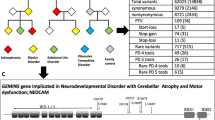
Ultra-rare variants were found in 11 genes: PPM1D (3 cases), NLRC4 (4 cases), RAG1 (3 cases), SGCE (2 cases), CACNA1B (2 cases), SHANK3 (3 cases), PLCG2, CHK2 (also known as CHEK2), GRIN2A, GABRG2, and SYNGAP1 in 21 patients with PANS. Among the NLRC4 and SHANK3 cases is one individual, case 19, in whom an ultra-rare variant was found in both genes (SHANK3, c.4622C>T, P1541L; NLRC4, c.928C>T, p.310X). The ultra-rare variants and brief descriptions of the cases are shown on Fig. 1 and Table 2, respectively. All of the cases were heterozygous for the variants. In the European cohort, two affected siblings (Cases 1 and 2) were found with an ultra-rare missense mutation in PPM1D (c.131C>G; p.S44W). Case 1 was treated with and responded to IVIg treatment. They inherited the variant from an asymptomatic parent who has a family history of autoimmune disorders, a common scenario in PANS families12,13. S44W is a known single nucleotide variant (SNV), rs373862041, with a minor allele frequency (MAF) of 0.00026, based on the TOPMed database of more than 125,000 samples14. The ultra-rare variant in CACNA1B, found in two affected siblings (Cases 4 and 5), is a 48 bp insert at the exon 2 splice donor site (c.390 + 1) that is predicted to disrupt splicing. It is part of a set of multiallelic insertion variants, rs370237172, that has an overall MAF of 0.017. However, the 48 bp insert is ultra-rare and has not been observed in control data sets. We have been unable to genotype the parents. Interestingly, the same 48 bp insert was also identified in a young woman who has been incapacitated with chronic fatigue syndrome (myalgic encephalomyelitis/chronic fatigue syndrome) and anorexia (unpublished observations). Known transmission from a parent in the European cohort was also found in Cases 10 and 11 (NLRC4: c.772 T>C; p.C258R; and SGCE: del150 Iso; c.450_452, respectively. Case 10 inherited the mutation from a mother who has a history of psoriasis and arthritis. She was diagnosed with focal epilepsy as an adult but has no history of PANS. A younger sibling with the same variant has a history suggestive of PANS but this has not yet been clinically ascertained. Two siblings of Case 11 (SGCE (del150 Iso; c.450_452) also inherited the variant. One has asthma, allergies, and contact dermatitis, but no neuropsychiatric problems. The other has similar atopic problems and debilitating cluster-type headaches.

Position of PANS ultra-rare variants within each candidate gene.
For the other ultra-rare variants, parental genotypes were not available. Remarkably, in the U.S. cohort, different ultra-rare variants were found in two of the same genes identified in the European cohort (NLRC4 and SGC2), and another, CHK2, codes for a well-established PPM1D substrate15.
In summary, all 21 cases (14 boys and 7 girls) described sudden onset of symptoms and in 20 cases these could be temporally associated to one or several infections (streptococcus (n = 16), intracellular bacteria (n = 7), or virus (n = 6) (Table 2). Mean age of PANS symptom onset was 7.15 years old (SD = 3.73). Their symptoms included OCD (n = 21), aggression or emotional dysregulation (n = 14), restrictive eating (n = 7), sensory or perception symptoms (n = 7), motor symptoms (n = 8), GI symptoms (n = 5), urinary urgency/incontinence (n = 3) and joint pain (n = 2). All seven cases who had received IVIg therapy reported partial or complete symptom reduction.
Cases 3 (PPM1D), 9 (SYNGAP1), 13 and 14 (NLRC4), 15 and 16 (RAG1), 19 and 20 (SHANK3), and 21 (CHK2) also had preexisting ASD or developmental delay prior to developing PANS. Case 3 had PANS characterized by severe OCD triggered by Streptococcus infections which improved with IVIg and plasmapheresis and was subsequently diagnosed with Jansen de Vries Syndrome (JdVS) based on a typical truncating mutation in PPM1D exon 6 (c.1547 C>G; p.S516X)16. Case 9 has a nonsense mutation in SYNGAP1 codon 1179 and was diagnosed with ASD as a child. Case 9 developed acute onset OCD and food restrictions following a culture positive Streptococcus pharyngitis. Case 18 (SHANK3: c.3637dupG, p.1213fsX68), who was diagnosed with Phelan-McDermid syndrome, which is usually caused by SHANK3 deletions, developed the sudden onset of severe OCD following bacterial infections. Case 20 (SHANK3: c.4776dupC, p.T1593fsX86) also had a history ASD with minimal verbal ability since early childhood. Case 20 developed the sudden onset of OCD and aggression as a child that improved dramatically with IVIg, although the baseline ASD and poor verbal ability did not improve.
Connectivity network
To assess potential functional connections between the PANS candidate genes, we generated a connectivity network to show the direct (solid lines) and indirect interactions (dotted lines) (Fig. 2). Central to the network is the NF-κB complex transcriptional regulator, which is activated by a variety of immune, infectious, and non-immune (e.g., oxidative stress; toxins) stressors17,18. Inappropriate activation of NF-κB has been associated with inflammatory diseases19. Directly connected to the NF-κB hub are the PANS candidate genes; PPM1D, PLCG2, NLRC4, RAG1, and CHK2. This hub represents the set of genes that likely function through a disruption of peripheral and central innate immunity. The PANS candidate genes not directly connected to NF-κB expression are those that are primarily expressed in the brain and cause neurodevelopmental disorders (CACNA1B, SYNGAP1, GRIN2A, SGCE, GABRG2, and SHANK3).

Connectivity network. A connectivity network was generated for each candidate gene using IPA software. Central to the network is the NF-κB complex transcriptional regulator hub, which connects to PPM1D, PLCG2, NLRC4, RAG1, BID, and CHK2. The PANS candidate genes not directly connected to NF-κB expression are those that are primarily expressed in the brain and cause neurodevelopmental disorders (CACNA1B, SYNGAP1, GRIN2A, SGCE, GABRG2, and SHANK3).
Identification of PANS candidate genes with differential regulation in microglia
PPM1D, PLCG2, and NLRC4 have well-established effects on microglia function20,21,22. To further assess the expression of these genes and other PANS candidates in microglia, we analyzed a gene expression dataset derived from LPS-treated mice (GSE102482)23. We found that Sgce and Plcg2 were the 2nd and 8th most downregulated transcripts, and Nlrc4 the 6th most upregulated transcript in microglia derived from control and LPS-treated mice upon an analysis of a subset of 148 autism and pediatric immune disorder genes (Fig. 3). Similarly, the expression of Sgce is induced in microglia following viral-mediated neuroinflammation in mice24, and we previously found that Plcg2 expression increased significantly in microglia derived from a mouse model of Rett syndrome, in which innate immune pathways were the most enriched differentially expressed genes25.

Heat map of LPS-mediated gene expression in microglia. Microarray gene expression data (GSE102482) from Greenhalgh et al. were analyzed to determine the expression pattern of a subset of 148 autism and pediatric immune disorder genes. The heat map shows the data obtained from RNA extracted from microglia grown on their own or co-cultured with peripheral macrophages. Arrows point to differentially expressed genes that are PANS candidates described in this report.
Expression of PANS candidate genes in PBMCs
To gain a more comprehensive understanding of the gene expression patterns in specific cell types and develop hypotheses for how PANS candidate gene variants lead to neuroinflammation, we examined several RNA expression resources, one of which was a single cell RNA sequence (scRNA-seq) database of peripheral blood mononuclear cells (PBMCs)26. This database contains scRNA-seq data from control samples (N = 6) and hospitalized COVID-19 patients (N = 7). The PANS candidate genes that have established effects on peripheral immune function, PPM1D, CHK2, and RAG1, were expressed in multiple PBMC types, while PLCG2 was primarily expressed in B-cells, and NLRC4 in monocytes and neutrophils (Supplementary Fig. S1). Interestingly, the expression of several of these genes was markedly altered in the COVID-19 samples. Most notably, a 170-fold increase in NLRC4 expression and a 100-fold increase in PLCG2 expression was found in developing neutrophils. In addition, 13 to 16-fold increases in PPM1D expression were found in IgG and IgM plasmablasts, interferon stimulated genes in T4 cells, and developing neutrophils in the COVID-19 samples, while a 12-fold decrease in expression in T.gd cells (γδ T cells) was observed. Finally, a 17-fold decrease in CHK2 expression was found in interferon-stimulated genes in T4 cells derived from the COVID-19 patients.
Although most of the PANS candidate genes that are highly expressed in neurons were expressed at very low levels in PBMCs, we identified three notable exceptions. Most striking is GABRG2, which showed only marginal expression in all PBMCs, but a conspicuous, 24-fold increase in γδ T cells derived from the COVID-19 samples. SYNGAP1 was expressed in many PBMCs, especially T cells. Like GABRG2, expression in γδ T cells increases approximately threefold in the COVID-19 samples. Finally, SHANK3 expression, while negligible in most PBMCs, was abundantly expressed in a cluster consisting of two groups of cells labeled as stem cells and eosinophils. In addition, a 64% increase was detected in the COVID-19 samples.
Expression of PANS candidate genes in the brain and other tissue
We next analyzed the cell and tissue expression pattern of the PANS candidate genes in normal human tissues using the GTEx dataset (Supplementary Fig. S2)27. Expression levels generally conformed to the functional groups described above and the connectivity network. The genes affecting peripheral immunity that are connected to the NF-κB hub were expressed primarily in blood and EB-transformed lymphoblasts, and less so in the brain, while the genes associated with neurodevelopmental disorders showed the opposite pattern. PPM1D, RAG1, and SGCE were highly expressed in both.
Expression of PANS candidate genes in adolescent and fetal mouse brain
The GTEx dataset was developed from bulk RNA-seq. To further characterize the expression of PANS candidate genes in individual neuronal brain cell types, we used published scRNA-seq data from the mouse (adolescent and developing brains)28,29. One of the more striking observations in the adolescent brain is the relatively high level of Chk2 expression in ependymal cells compared with all other brain cell types (Fig. 4; see Supplementary Table S1 for abbreviation key). Ependymal cells line the ventricles and spinal canal and play an important role in the production of CSF, and as part of the choroid plexus, regulation of the blood-CSF barrier30. The choroid plexus plays a key role in neuroinflammatory and neurodegenerative disorders31,32,33,34.

scRNA-seq expression pattern of each PANS candidate gene in mouse adolescent brains (open access article; CC BY license http://creativecommons.org/licenses/by/4.0/).
Ppm1d, Sgce, Plcg2, Syngap1, and Shank3 were also expressed in these cells, suggesting that their mutated versions could disturb the blood-CSF barrier, allowing cytokines, chemokines, autoantibodies, and immune cells to enter the brain.
Another interesting finding in the mouse adolescent brain database was the relatively high level of Ppm1d, Syngap1, Cacna1b, and Sgce expression throughout the mouse enteric nervous system. The potential implications of this expression pattern will be described in the discussion section.
Expression In the developing mouse brain is shown in Supplementary Fig. S3. One of the more interesting findings was Shank3, which, as expected, was diffusely expressed throughout the brain in both glutamatergic neurons and GABAergic interneurons. However, the highest levels were seen in the choroid plexus and cerebral vasculature. In fact, all of the PANS candidate genes, with the exception of RAG1 were expressed in fetal choroid plexus, cerebral vasculature, and pericytes, with Sgce, Chk2, and Ppm1d, along with Shank3, being most prominent. This was similar to the single cell expression pattern seen in adolescent brains (Fig. 4), reinforcing the idea that disruption of the blood-CSF barrier and/or the BBB is playing a role in the emergence of PANS.
Discussion
In the current study, we used next generation sequencing to search for ultra-rare genetic variants in patients who met diagnostic criteria for PANS as established by the PANS Consensus Conference1. Mutations were identified in 21 patients in 11 genes that separate into two broad functional categories: those that affect peripheral innate and adaptive immune pathways, and those that are expressed primarily in cortical neurons, where they function as synaptic regulators and have all been implicated in other neurological and neurodevelopmental disorders. The findings additionally indicated that when mutated, the majority of genes could have unanticipated effects on immune cells, especially in response to infectious diseases. In addition, several genes are expressed in the choroid plexus and brain vasculature, suggesting they might contribute to a breach in the blood-CSF barrier and blood–brain barrier (BBB) that accompanies inflammation and infection. Although there is a clear ascertainment bias for the participants in the European sample, the finding of ultra-rare variants in the same genes and in an unselected modestly sized population in our United States cohort suggest that approximately 5% of PANS cases may be caused by biologically powerful ultra-rare genetic variants that affect immune and/or brain function.
Genetic contributions to disease can be divided into common variants that have relatively small biological effects, which are identified by large-scale GWAS, and high impact, biologically significant ultra-rare variants. So far, no reports have been published in which common variants associated with PANS have been identified, although, there is a small, unpublished study derived from a direct-to-consumer DNA testing service that reported several SNPs associated with PANS (https://ejournal2.undip.ac.id/index.php/jbtr/article/view/12082?utm_source=DSMN8&utm_medium=LinkedIn). However, the sample size is too small to assess their validity.
Several mutations were discovered in genes with well-established effects on peripheral innate and adaptive immunity: NLRC4, PPM1D, RAG1, CHK2, and PLCG2. NLRC4 codes for a component of the inflammasome, a cytosolic multiprotein complex that assembles in response to exogenous or endogenous stressors that plays a major role in autoinflammatory diseases, macrophage activation syndrome, and panoptosis35,36. A total of four patients with ultra-rare variants in NLRC4 were found in our two cohorts (cases 10, 13, 14, and 19; case 19 who also has an ultra-rare SHANK3 variant). Two variants, c.772T>C (p.C258R) and c.928C>T (p.R310X) map to the NACHT domain, which has intrinsic ATPase activity and facilitates self-oligomerization37. Gain-of-function mutations in this region lead to severe hyperinflammatory disorders38. Such mutations abrogate autoinhibition by the C-terminal leucine rich region39. p.R310X would eliminate this region from the protein. The p.R310X variant has been observed in six patients with multiple sclerosis from two independent families following exome sequence screening, although the segregation pattern was incomplete, indicating that other factors may contribute to MS development in these families40.
The other two ultra-rare variants, c.2309 T>C (p.M770T) and c.1799A>G (p.E600G), map to the leucine-rich repeat region (LRR). Mutations in the LRR affecting the oligomerization interface were recently described in two patients with early-onset macrophage activation syndrome41.
Mutations in PPM1D were identified in 3 cases. PPM1D codes for a serine/threonine phosphatase that negatively regulates p53 and other members of the DNA repair pathway, and somatic gain-of-function mutations act as tumor suppressor genes42. The most common are frameshift and nonsense mutations in exons 5 and 6 that lead to the production of a truncated protein that has a stabilizing effect on the retained catalytic portion. Germline exons 5 and 6 truncating mutations are found in children with JdVS, which is characterized by intellectual disabilities (ID), restricted eating, high pain threshold, autism spectrum disorder (ASD), and psychiatric symptoms (primarily severe anxiety), symptoms that overlap with PANS16. Case 1 had severe, chronic PANS who was found, along with an affected sibling (Case 2), to have an ultra-rare PPM1D mutation. This mutation is upstream of the phosphatase domain and its effect on PPM1D catalytic function is not immediately clear. However, the nearby serine amino acids S40 and S46 are substrates for the serine kinases DYRK1A and HIPK2, which cause ID, ASD, and abnormal feeding behavior when mutated43,44. DYRK1A also has important effects on innate immunity by regulating the balance between T regulatory (Treg) cells and Th17 cells, the latter of which mediates the effects of Streptococcus on neuroinflammation in a mouse model of PANS45,46,47,48. HIPK2 regulates PPM1D protein levels and knockdown attenuates expression of inflammatory cytokines in LPS-stimulated macrophages49,50. The effect of S44W on immune function and its effect on DYRK1A and HIPK2 awaits experimental validation. The finding of PANS patient with a JdVS mutation (Case 3) strongly supports the idea that some PPM1D genetic variants increase the risk of PANS. It is interesting to note that most children with JdVS have a history of recurrent infections16. Although the underlying molecular and cellular basis for this is not known, infections and PANS could be due to the effect of PPM1D on T- and B-lymphocyte differentiation, and cytokine production, as described in humans and in mouse knockout models51,52,53,54.
The rare variant we identified in CHK2, is a truncating mutation in the distal end of the kinase domain and is probably a loss-of-function mutation. Like PPM1D, CHK2 also codes for a regulator of the DNA repair response by phosphorylating p5315,55. It also regulates IL-2 expression in T-cells56, and Chk2 inactivation in mouse B cells leads to decreased Ig hypermutation, Ig class switching and immune dysregulation57.
RAG1 codes for recombination activating gene 1, which is involved in antibody and T-cell receptor V(D)J recombination and loss of expression leads to abnormalities in T- and B-cell tolerance and immune dysregulation, leading to both an increased risk of infection and autoimmune problems58,59. Ultra-rare mutations were found in three patients (cases 15, 16, and 17), one of which causes a frameshift in exon 2 (c.256_257delAA; p.K86VfsX33). This variant has previously been reported in a heterozygous patient with adult-onset lymphopenia, and in a homozygous patient with severe combined immunodeficiency (SCID)60.
PLCG2 is primarily expressed in B-cells. Gain-of-function mutations have previously been reported in patients with severe sterile inflammation, recurrent bacterial infections, autoimmune disorders, and humoral immunodeficiency61, and in autoinflammatory disease through an increase in calcium influx upon B-cell activation62. The PLCG2 Y206F variant in Case 8 maps to the EF hand domain that binds calcium. However, the effect of Y206F on calcium homeostasis awaits experimental validation. Interestingly, this same ultra-rare variant was found in a patient diagnosed with Familial Cold Autoinflammatory Syndrome-3 (ncbi.nlm.nih.gov/clinvar/, accession number VCV000574390.1).
Six PANS candidate genes identified in this study, SHANK3, GRIN2A, SYNGAP1, CACNA1B, GABRG2, and SGCE, are primarily expressed in neurons, especially at excitatory synapses, and variants in these genes are associated with ASD and other neurodevelopmental disorders. SHANK3, for example, codes for a scaffold protein that regulates the assembly of the postsynaptic density (PSD) at glutamatergic excitatory synapses and is among the most commonly mutated genes in ASD and other neurodevelopmental disorders63,64,65. Of the 1003 genes in the Simons Foundation Autism Research Initiative (SFARI) database (gene.sfari.org/database/human-gene), SHANK3 has the second highest number of reports. Ultra-rare variants were found in three PANS patients. Although SHANK3 loss-of-function mutations in ASD are scattered throughout the gene, the majority, including our cases, are found in exon 21, which codes for the proline-rich region that binds to and attracts other synaptic proteins to the PSD.
Our finding of comorbid ASD and PANS in the three SHANK3 cases is similar to observations made by Bey et al., who identified de novo SHANK3 variants in four girls with lifelong, stable developmental delay (DD) who developed subacute, severe psychiatric symptoms resembling PANS, that responded, with varying degrees of success, to immunotherapy66. Similarly, in an analysis of 38 individuals with Phelan-McDermid Syndrome, acute regressions triggered by infections, and other life events were common, and several patients improved with IVIg67. These findings show that a subgroup of patients with SHANK3-associated ASD and developmental disorders have an underlying susceptibility to develop acute onset neuropsychiatric problems that have a neuroinflammatory component.
In four of the other PANS-associated neuronal candidate genes we identified, SYNGAP1, GRIN2A, GABRG2, and CACNA1B, variants have been found in ASD, epilepsy, and ID68,69,70,71. According to the SFARI database, SYNGAP1 has the ninth highest number of reports.
GRIN2A codes for the glutamate ionotropic receptor NMDA type subunit 2A. The ultra-rare variant found in Case 7 maps to a conserved region of the CTD domain, which interacts with the MAGUK (membrane-associated guanylate kinase) family of proteins that bind to PSD and function as important modulators of synaptic plasticity72. This subject has a history of dyslexia and ADHD.
CACNA1B codes for the voltage-dependent N-type calcium channel subunit alpha-1B, a constituent of the Cav2.2 channel. It too is a regulator of synaptic function that acts at the presynaptic terminal to increase neurotransmitter release, which then influences postsynaptic dendritic spine function73. In addition to having PANS, Case 4 was diagnosed with Hodgkin lymphoma that ultimately required an autologous hematological stem cell transplant (HSCT). It is interesting to note that Hodgkin lymphoma is often found in patients with autoimmune problems and is associated with abnormalities in IL-13 signaling, a cytokine produced by mast cells, eosinophils, nuocytes, and Th2 cells74,75. This patient also has psoriasis, a Th17 associated autoimmune disorder76,77. This suggests the possibility that CACNA1B could have unrecognized effects on the immune system by disrupting IL-13 and Th17 signaling leading to PANS and an increased risk of hematological malignancy. It is interesting to note that autologous HSCT is an emerging clinical tool for treating severe autoimmune disorders78, However, case 4 was in remission from PANS at the time HSCT was carried out.
GABRG2 codes for the gamma subunit of the GABA-A receptor, a ligand-gated chloride channel. It is one of the most mutated genes in febrile seizures and other forms of epilepsy79. We found a nonsense mutation in a child with severe PANS (Case 6) that responded to IVIg who also has a history of febrile seizures. This is the same mutation previously found in sleep-related epilepsy80.
Ultra-rare mutations in SGCE were found in two unrelated subjects, Cases 11 and 12. SGCE is a member of the sarcoglycan family of transmembrane proteins, a component of the dystrophin-associated glycoprotein complex (DGC). Case 11 responded to IVIg treatment. Studies in knockout mice show that SGCE disrupts excitatory synapse formation81. The DGC is involved in the formation of the glymphatic system, which is impaired in neuroinflammation and dementia82,83.
Remarkably, SGCE, CACNA1B, and GRIN2A are well-known for causing Myoclonus-Dystonia (M-D), a hyperkinetic movement disorder that resembles tics, which is found in a subgroup of PANS. SGCE, in fact, is the most commonly mutated gene in M-D, with more than one-third of cases having truncating mutations in exon 3, which contains the del150Iso in-frame mutation found in Case 1184,85,86. Interestingly, patients with myoclonus caused by SGCE mutations frequently have comorbid psychiatric symptoms, including depression, anxiety, bipolar disorder, and OCD85, symptoms that overlap with PANS. Similarly, GRIN2A and CACNA1B mutations have been found in dystonia and other movement disorders87,88.
These findings suggest that deleterious variants in these neuronal genes can result in a range of clinical phenotypes; M-D, ASD, developmental disabilities, epilepsy, and PANS, or PANS comorbid with these neurodevelopmental problems. It is important to add that cases 3 (PPM1D), 9 (SYNGAP1), 13 and 14 (NLRC4), 15 and 16 (RAG1), 19 and 20 (SHANK3), and 21 (CHK2) also had preexisting ASD or developmental delay prior to developing PANS.
Evidence that penetrance is not 100% is seen in several families in whom transmission occurred from a carrier parent who does not have PANS (Cases 1 and 2; 4 and 5), although other conditions such as asthma, allergies, and autoimmune disorders were reported. The lack of complete penetrance is consistent with a small monozygotic twin study in PANS that showed a range of different phenotypes, including complete discordance89. Similarly, the MZ twin concordance rate in autoimmune disorders ranges from approximately 20%-70% depending on the condition90. This suggests that environmental factors are playing a role in PANS. In the case of the generation divide between carrier parents and their affected children, changes in the prevalence of certain strains of bacteria and antigenic shifts over the decades in viruses implicated as PANS triggers could account for their different clinical outcomes. In addition, the current generation of youth might be exposed to non-infectious environmental triggers that were not as prevalent in past generations. Although unaffected carrier parents might be exposed to the same infectious agents or non-infectious stressors that are currently triggering PANS flareups in their children, age-related differences in the blood-CSF barrier or BBB could make the adult brain less susceptible91. Genetic background and stochastic T cell receptor and IgG gene rearrangement could also explain reduced penetrance in families and discordance in MZ twins.
The question remains if the PANS syndrome should be regarded as one or several clinical entities. The two functional groups into which our candidate genes can basically be divided; those that have established effects on peripheral innate and adaptive immunity, and those that affect synaptic function, in particular the PSD complex, indicates a peripheral versus central dichotomy. However, our results suggest this may be an oversimplification. First, as noted earlier, several PANS candidate genes that have effects on peripheral innate immunity, such as PPM1D, PLCG2, and NLRC4, also affect microglia function and are differentially expressed in those cells following an immune challenge (Fig. 3). Second, patients with ASD and mutations in genes that function as synaptic regulators, such as SHANK3, can respond to IVIg, as shown by Bey et al.66 and Case 20 in this report. Third, as presented in Supplementary Fig. S1, neuronal genes like SHANK3, GABRG2 and SYNGAP1 show expression patterns that suggest an effect on immune function, in particular γδ T-cells, based on their markedly altered expression during an infectious disease challenge. γδ T cells form a minor population of PBMCs, but they increase in number dramatically during infections and play a key role in autoimmunity and immune surveillance92. γδ T cells are also found in meninges where they secrete the proinflammatory cytokine IL-17a and participate in the development of anxiety-like behavior in mice93. Fourth, the neuronal PANS candidate genes that have effects on synaptic function are also expressed in the choroid plexus and brain vascular endothelium, in particular SHANK3, which could potentially connect peripheral inflammation with neuroinflammation through disruption of the brain/CSF barrier and BBB. SHANK3 is a cytoskeletal protein that regulates glutamatergic synaptogenesis, but it can also function as a scaffolding protein in epithelial cells94. In addition, PPM1D has been found to regulate BBB function and neuroinflammation in a co-culture of human brain-microvascular endothelial cells and human astrocytes treated with LPS95.
Expression of PANS candidate genes in the choroid plexus and vascular endothelium during fetal development is also interesting when considering the phenomenon known as maternal immune activation (MIA), a proinflammatory state in pregnancy triggered by infection, maternal autoimmune disorders, and non-infectious peripheral inflammation96. The fetal brain is vulnerable to changes in the maternal/fetal environment, such as MIA, which has been shown in animal models to adversely affect brain development, leading to behaviors and learning difficulties similar to those seen in ASD and schizophrenia97,98. Consequently, it is conceivable that the pathophysiological process that leads to the development of PANS and comorbid neurodevelopmental disorders could begin during fetal life in some patients and genetic subgroups. An effect on the choroid plexus and vasculature endothelium (and γδ T cells) could also explain the response to IVIg in those patients harboring mutations in the “neuronal” subgroup of candidate genes. IVIg can reduce neuroinflammation by affecting T-cell/microglia crosstalk, reducing levels of proinflammatory cytokines, blocking Fcγ receptors, inhibiting complement, and repairing disrupted brain-CSF barriers99,100.
It is also interesting to consider the implications of the finding that several PANS candidate genes are also mutated in D-M, ASD, and other neurodevelopmental disorders, as noted above. PANS is occurring as an independent phenotype in some of our cases, and as a comorbid trait in others (Table 2). These comorbid cases are like the SHANK3-mutated patients reported by Bey et al.66. Similarly, Jones et al., described eight children with ASD and other neurodevelopmental disorders, with a strong family history of maternal autoimmune thyroid disorders, who presented with infection-induced, abrupt onset of neuropsychiatric symptoms, primarily OCD and tics, along with autistic or global regression101. Numerous genetic and molecular studies also indicate an inflammatory component in the pathogenesis of subgroups of patients with schizophrenia and ASD102,103,104,105. In addition, it has been suggested that a subgroup of regressive autism, which is characterized by the sudden loss of previously acquired traits in early childhood, could have an autoimmune or infectious disease etiology106,107,108. Interestingly, deleterious GRIN2A mutations were found in ~ 2% of cases in a recent study by Yin et al.107. At the very least, an inflammatory component leading to the abrupt onset of PANS superimposed on a chronic neurodevelopmental disorder appears to be a real phenomenon and tools to improve early recognition of these cases may be used to motivate immunological therapies like IVIg that empirical evidence suggests could be very helpful.
Finally, an interesting aspect of our scRNA-seq analysis was the finding that many of the PANS candidate genes are expressed in enteric neurons. Enteric neurons have many connections with immune cells in the intestinal mucosa, and gut-associated lymphoid tissue, which could affect, or be affected by gut bacteria109. More specific to the candidate genes we identified, several have established effects on the gut. PPM1D, for example, has been found to have a protective effect on oxidation stress-induced gut permeability110. In addition, patients with PANS have been found to have differences in the gut microbiome compared with controls111, and have an increase in gut derived LPS112. In addition, Shank3 knockout mice have an altered microbiota composition, an increase in LPS levels in the liver, and altered gut permeability113. Finally, mutations in NLRC4 have been implicated in inflammatory bowel disease114,115. Thus, expression of PANS candidate genes in the enteric nervous system fits into the emerging idea that disruption of the gut-brain connection and the gut microbiome are involved in the pathogenesis of ASD, neurodegeneration, neuropsychiatric disorders, autoimmune disorders, and PANS.
In conclusion, we identified ultra-rare genetic variants in PANS patients that appear to function at multiple levels of the neuroinflammatory circuit, including peripheral and central innate immunity, synaptogenesis, the blood-CSF barrier, and perhaps the enteric nervous system. Dissecting the molecular and cellular pathogenesis of the PANS candidate variants will require an analysis in mouse models, as well as patient-specific induced pluripotent stem cells, from which cells such as neurons, microglia, astrocytes, vascular endothelium, and gut organoids can be derived to assess their importance in the development of the devastating symptoms that comprise the PANS syndrome.
Subjects and methods
Subjects
The European subjects were identified through a call for patients with severe symptoms from a PANS group called EXPAND, a European advocacy organization for families of children and adolescents with immune-mediated neuropsychiatric disorders, after a teenager with chronic PANS was found to have an ultra-rare variant in the PPM1D gene by WGS. Parents, caretakers, or patients signed informed consents approved by the Ethical Committee at Erasmus MC (MEC-2011–253 for control samples and MEC-2021–0359 for PANS patients; IRB/ Human Subject Assurance number/Federal Wide Assurance, FWA00001336) in accordance with the Declaration of Helsinki – ethical principles for medical research involving human subjects. Histories from this cohort were obtained by the participating physicians and collated by one of the co-authors (O.M). Cases from the U.S. were obtained from a large private practice run by one of the authors (R.T) devoted to PANS (PANDAS/PANS Institute, Ramsey, NJ). 383 PANS cases and 263 controls that included 133 triads had WES carried out by Centogene (see below) (Rostock, Germany). The subjects signed an informed consent with Centogene. Each case in the U.S. cohort was personally treated by one physician (R.T.) who obtained a detailed personal and family history, and permission to present genetic and clinical information relevant to the diagnosis of PANS. Each case met diagnostic criteria for PANS as established by the PANS Consensus Conference1. For cases and family members diagnosed with autoimmune disorders, no formal assessments were carried out for this study for most of the subjects. We relied on detailed family histories obtained by the personal physicians.
WGS and WES data analysis
The EU cases (1, 2, 4–11) were sequenced using DAB nanoball sequencing for WGS and Illumina for WES. The variants shown in Table 2 were found in both sequencing efforts. We implemented ANNOVAR, an efficient software tool that utilizes up-to-date information to functionally annotate genetic variants and allele frequencies detected in each patient’s genome. In addition to gene-based annotation, ANNOVAR is used for region-based annotations to identify variants in specific genomic regions, such as ultra-conserved regions across species (Vista database), predicted transcription factor binding sites (Transfac), segmental duplication regions, genome wide association study (GWAS) hits, database of genomic variants, DNAse I hypersensitivity sites, ENCODE H3K4Me1/H3K4Me3/H3K27Ac/CTCF sites, ChIP-Seq peaks, RNA-Seq peaks, and other annotations in genomic intervals. DNA nanoball sequencing was applied for WGS to study non-coding regions and in particular variants within the ultra-conserved regulatory regions. This provided highly reliable variant lists that were compared with our selected variants and filtered based on allele frequency in GNOMAD ClinVar (public NCBI repository derived data). Only variants that occurred in patients and were never detected in 597 healthy elderly (Wellderly) sample genomes were considered116. DNA Nanoball sequencing protocol is detailed in the classic paper by Drmanac et al.117. The ultra-rare variant assessment was confirmed using additional cohorts that have been sequenced by the Erasmus MC team headed by one of us (P.S.).
The U.S. samples were sequenced by Centogene using double stranded DNA capture baits against approximately 36.5 Mb of the human coding exome (targeting > 98% of the coding elements). RefSeq from the human genome build GRCh37/hg19) are used to enrich target regions from fragmented genomic DNA with the Twist Human Core Exome Plus kit. The generated library was sequenced on an Illumina platform to obtain at least 20 × coverage depth for > 98% of the targeted bases. The investigation for relevant variants focused on coding exons and flanking ± 20 intronic nucleotides of genes with clear gene-phenotype evidence. The resulting variant call file (VCF) file was then subjected to a custom pipeline developed by one of the co-authors (R.T). Variants were filtered as follows: 1. Those with annotated mean allele frequencies of > 0.1% were removed, 2. Variants marked as "benign" or "likely benign" on EITHER ClinVar or ACMG/InterVar database were removed, 3. Variants annotated as “low severity” were removed.
Network analysis
Functional analysis was performed within Ingenuity Pathway Analysis (IPA) (QIAGEN Inc.). All candidate genes were imported in IPA to assess the pathways involved. Using the network tool within IPA, a connectivity network was constructed based on the IPA/QIAGEN Knowledge Base. Both direct and indirect relationships were used to construct the biological network.
RNA expression analysis
The gene expression profiles of the candidate genes were analyzed using several datasets. First, we hypothesized that microglia might play a critical role in the PANS phenotype especially after an inflammatory stimulus. Consequently, a subset of 159 autism and pediatric immune disorder genes was analyzed based on a public dataset investigating the effect of lipopolysaccharide (LPS) on murine microglia and macrophages (GSE102482)23. After Robust Multichip Average normalization (RMA), a statistical analysis of microarray (SAM) was performed between non-stimulated microglia and LPS-stimulated microglia within OmniViz version 6.1.13.0 (Instem Scientific). The top 15 up-regulated and down-regulated genes were selected for visualization and further analysis.
Next, we investigated the gene expression profiles of our 11 PANS-associated candidates in peripheral blood mononuclear cells (PBMCs) under baseline conditions and after infection as many patients develop PANS symptoms following infections. Given the rising reporting of neuropsychiatric symptoms after COVID-19 infection, we used a recently published sc-RNA-seq COVID-19 dataset to investigate the gene expression profiles26. This database contains gene expression patterns in control samples (N = 6) and hospitalized severe COVID-19 patients (N = 7).
To investigate the expression of the candidate genes in normal human tissue we used the Genotype-Tissue Expression (GTEx) dataset (https://www.gtexportal.org/home)27. The Multi Gene Query available on the GTEx portal was used to construct the RNA profiles in normal human tissue lineages. Finally, as the GTEx portal uses bulk RNA-seq data, we wanted to further investigate the gene expression profiles in brain tissue using scRNA-seq. Since scRNA-seq data is not yet available for human brains, we analyzed the expression pattern in two datasets that were recently published containing scRNA-seq data for adolescent and fetal mouse brains28,29. Each candidate gene was investigated in both datasets on the Mouse Brain Atlas portal (http://mousebrain.org).
References
Chang, K. et al. Clinical evaluation of youth with pediatric acute-onset neuropsychiatric syndrome (PANS): Recommendations from the 2013 PANS consensus conference. J. Child Adolesc. Psychopharmacol. 25, 3–13 (2015).
Swedo, S. E. et al. Identification of children with pediatric autoimmune neuropsychiatric disorders associated with streptococcal infections by a marker associated with rheumatic fever. Am. J. Psychiatry 154, 110–112 (1997).
Calaprice, D., Tona, J., Parker-Athill, E. C. & Murphy, T. K. A survey of pediatric acute-onset neuropsychiatric syndrome characteristics and course. J. Child Adolesc. Psychopharmacol. 27, 607–618 (2017).
Frankovich, J. et al. The Burden of caring for a child or adolescent with pediatric acute-onset neuropsychiatric syndrome (PANS): An observational longitudinal study. J. Clin. Psychiatry 80, 17m12091. https://doi.org/10.4088/JCP.17m12091 (2018).
Wilbur, C. et al. PANDAS/PANS in childhood: Controversies and evidence. Paediatr. Child. Health. 24, 85–91 (2019).
highlights and controversies. Chiarello, F., Spitoni, S., Hollander, E., Matucci Cerinic, M. & Pallanti, S. An expert opinion on PANDAS/PANS. Int. J. Psychiatry Clin. Pract. 21, 91–98 (2017).
Gilbert, D. L. Inflammation in tic disorders and obsessive-compulsive disorder: Are PANS and PANDAS a path forward?. J. Child Neurol. 34, 598–611 (2019).
Zheng, J. et al. Association of pediatric acute-onset neuropsychiatric syndrome with microstructural differences in brain regions detected via diffusion-weighted magnetic resonance imaging. JAMA Netw. Open 3, e204063 (2020).
Vitaliti, G. et al. The usefulness of immunotherapy in pediatric neurodegenerative disorders: A systematic review of literature data. Hum. Vaccin Immunother. 11, 2749–2763 (2015).
Melamed, I. et al. Evaluation of intravenous immunoglobulin in pediatric acute-onset neuropsychiatric syndrome. J. Child Adolesc. Psychopharmacol. 31, 118–128 (2021).
Gromark, C. et al. A Two-to-five year follow-up of a pediatric acute-onset neuropsychiatric syndrome cohort. Child Psychiatry Hum. Dev. 1–11 (2021).
Gagliano, A. et al. Pediatric acute-onset neuropsychiatric syndrome: A data mining approach to a very specific constellation of clinical variables. J. Child Adolesc. Psychopharmacol. 30, 495–511 (2020).
Fernell, E. et al. Paediatric Acute onset Neuropsychiatric Syndrome: Exploratory study finds no evidence of HLA class II association but high rate of autoimmunity in first-degree relatives. Acta Paediatr. (2021).
Burgess, D. J. The TOPMed genomic resource for human health. Nat. Rev. Genet. (2021).
Yoda, A. et al. Intrinsic kinase activity and SQ/TQ domain of Chk2 kinase as well as N-terminal domain of Wip1 phosphatase are required for regulation of Chk2 by Wip1. J. Biol. Chem. 281, 24847–24862 (2006).
Jansen, S. et al. De novo truncating mutations in the last and penultimate exons of PPM1D cause an intellectual disability syndrome. Am. J. Hum. Genet. 100, 650–658 (2017).
Manthiram, K., Zhou, Q., Aksentijevich, I. & Kastner, D. L. The monogenic autoinflammatory diseases define new pathways in human innate immunity and inflammation. Nat. Immunol. 18, 832–842 (2017).
Taniguchi, K. & Karin, M. NF-κB, inflammation, immunity and cancer: coming of age. Nat. Rev. Immunol. 18, 309–324 (2018).
Liu, T., Zhang, L., Joo, D. & Sun, S. C. NF-κB signaling in inflammation. Signal. Transduct Target Ther. 2, 17023 (2017).
Li, D. et al. WIP1 phosphatase plays a critical neuroprotective role in brain injury induced by high-altitude hypoxic inflammation. Neurosci. Bull. 33, 292–298 (2017).
Andreone, B. J. et al. Alzheimer’s-associated PLCγ2 is a signaling node required for both TREM2 function and the inflammatory response in human microglia. Nat. Neurosci. 23, 927–938 (2020).
Poh, L. et al. Evidence that NLRC4 inflammasome mediates apoptotic and pyroptotic microglial death following ischemic stroke. Brain Behav. Immun. 75, 34–47 (2019).
Greenhalgh, A. D. et al. Peripherally derived macrophages modulate microglial function to reduce inflammation after CNS injury. PLoS Biol. 16, e2005264 (2018).
DePaula-Silva, A. B. et al. Differential transcriptional profiles identify microglial- and macrophage-specific gene markers expressed during virus-induced neuroinflammation. J. Neuroinflammation 16, 152–019–1545-x (2019).
Zhao, D. et al. Transcriptome analysis of microglia in a mouse model of Rett syndrome: differential expression of genes associated with microglia/macrophage activation and cellular stress. Mol. Autism 8, 17–017–0134-z. eCollection 2017 (2017).
Wilk, A. J. et al. A single-cell atlas of the peripheral immune response to severe COVID-19. Nat Med. 26(7), 1070–1076. https://doi.org/10.1038/s41591-020-0944-y (Epub 2020 Jun 8).
GTEx Consortium. The GTEx Consortium atlas of genetic regulatory effects across human tissues. Science 369, 1318–1330 (2020).
Zeisel, A. et al. Molecular Architecture of the Mouse Nervous System. Cell 174, 999-1014.e22 (2018).
La Manno, G. et al. Molecular architecture of the developing mouse brain. Nature 596, 92–96 (2021).
Javed, K., Reddy, V. & Lui, F. in StatPearls (StatPearls Publishing LLC, 2021).
Huang, X., Hussain, B. & Chang, J. Peripheral inflammation and blood-brain barrier disruption: Effects and mechanisms. CNS Neurosci. Ther. 27, 36–47 (2021).
Dixon, G. A. & Pérez, C. A. Multiple sclerosis and the choroid plexus: Emerging concepts of disease immunopathophysiology. Pediatr. Neurol. 103, 65–75 (2020).
Hubert, V. et al. MRI coupled with clinically-applicable iron oxide nanoparticles reveals choroid plexus involvement in a murine model of neuroinflammation. Sci. Rep. 9, 10046–019–46566–1 (2019).
Steeland, S. & Vandenbroucke, R. E. Choroid plexus tumor necrosis factor receptor 1: A new neuroinflammatory piece of the complex Alzheimer’s disease puzzle. Neural Regen. Res. 14, 1144–1147 (2019).
Sundaram, B. & Kanneganti, T. D. Advances in understanding activation and function of the NLRC4 inflammasome. Int. J. Mol. Sci. 22, 1048. https://doi.org/10.3390/ijms22031048 (2021).
Andrade, W. A. & Zamboni, D. S. NLRC4 biology in immunity and inflammation. J. Leukoc. Biol. 108, 1117–1127 (2020).
Sandall, C. F., Ziehr, B. K. & MacDonald, J. A. ATP-binding and hydrolysis in inflammasome activation. Molecules 25, 4572. https://doi.org/10.3390/molecules25194572 (2020).
Liang, J. et al. Novel NLRC4 mutation causes a syndrome of perinatal autoinflammation with hemophagocytic lymphohistiocytosis, hepatosplenomegaly, fetal thrombotic vasculopathy, and congenital anemia and ascites. Pediatr. Dev. Pathol. 20, 498–505 (2017).
Hu, Z. et al. Crystal structure of NLRC4 reveals its autoinhibition mechanism. Science 341, 172–175 (2013).
Popplewell, L. F. et al. Genetic analysis of nucleotide-binding leucine-rich repeat (NLR) receptors in multiple sclerosis. Immunogenetics 72, 381–385 (2020).
Moghaddas, F. et al. Autoinflammatory mutation in NLRC4 reveals a leucine-rich repeat (LRR)-LRR oligomerization interface. J. Allergy Clin. Immunol. 142, 1956-1967.e6 (2018).
Cardoso, M., Paulo, P., Maia, S. & Teixeira, M. R. Truncating and missense PPM1D mutations in early-onset and/or familial/hereditary prostate cancer patients. Genes Chromosomes Cancer 55, 954–961 (2016).
Earl, R. K. et al. Clinical phenotype of ASD-associated DYRK1A haploinsufficiency. Mol. Autism 8, 54–017–0173–5. eCollection 2017 (2017).
O’Roak, B. J. et al. Multiplex targeted sequencing identifies recurrently mutated genes in autism spectrum disorders. Science 338, 1619–1622 (2012).
Cerboni, S., Gehrmann, U., Preite, S. & Mitra, S. Cytokine-regulated Th17 plasticity in human health and diseases. Immunology (2020).
Khor, B. et al. The kinase DYRK1A reciprocally regulates the differentiation of Th17 and regulatory T cells. Elife 4. https://doi.org/10.7554/eLife.05920 (2015).
Cutforth, T., DeMille, M. M., Agalliu, I. & Agalliu, D. CNS autoimmune disease after Streptococcus pyogenes infections: Animal models, cellular mechanisms and genetic factors. Future Neurol. 11, 63–76 (2016).
Platt, M. P. et al. Th17 lymphocytes drive vascular and neuronal deficits in a mouse model of postinfectious autoimmune encephalitis. Proc. Natl. Acad. Sci. USA 117, 6708–6716 (2020).
Choi, D. W. et al. WIP1, a homeostatic regulator of the DNA damage response, is targeted by HIPK2 for phosphorylation and degradation. Mol. Cell 51, 374–385 (2013).
Xu, L., Fang, H., Xu, D. & Wang, G. HIPK2 sustains inflammatory cytokine production by promoting endoplasmic reticulum stress in macrophages. Exp. Ther. Med. 20, 171 (2020).
Tang, Y. et al. Wip1-dependent modulation of macrophage migration and phagocytosis. Redox Biol. 13, 665–673 (2017).
Zhang, Q. et al. Wip 1 inhibits intestinal inflammation in inflammatory bowel disease. Cell. Immunol. 310, 63–70 (2016).
Yi, W. et al. Phosphatase Wip1 controls antigen-independent B-cell development in a p53-dependent manner. Blood 126, 620–628 (2015).
Brichkina, A. & Bulavin, D. V. WIP-ing out atherosclerosis with autophagy. Autophagy 8, 1545–1547 (2012).
Stolarova, L. et al. CHEK2 germline variants in cancer predisposition: Stalemate rather than checkmate. Cells 9, 2675. https://doi.org/10.3390/cells9122675 (2020).
Helgeland, H. et al. Transcriptome profiling of human thymic CD4+ and CD8+ T cells compared to primary peripheral T cells. BMC Genomics 21, 350–020–6755–1 (2020).
Davari, K., Frankenberger, S., Schmidt, A., Tomi, N. S. & Jungnickel, B. Checkpoint kinase 2 is required for efficient immunoglobulin diversification. Cell Cycle 13, 3659–3669 (2014).
Erman, B. & Çipe, F. Genetic screening of the patients with primary immunodeficiency by whole-exome sequencing. Pediatr. Allergy Immunol. Pulmonol. 33, 19–24 (2020).
Delmonte, O. M., Villa, A. & Notarangelo, L. D. Immune dysregulation in patients with RAG deficiency and other forms of combined immune deficiency. Blood 135, 610–619 (2020).
Abraham, R. S. et al. Adult-onset manifestation of idiopathic T-cell lymphopenia due to a heterozygous RAG1 mutation. J. Allergy Clin. Immunol. 131, 1421–1423 (2013).
Martín-Nalda, A. et al. Severe autoinflammatory manifestations and antibody deficiency due to novel hypermorphic PLCG2 mutations. J. Clin. Immunol. 40, 987–1000 (2020).
Novice, T. et al. A germline mutation in the C2 domain of PLCγ2 associated with gain-of-function expands the phenotype for PLCG2-related diseases. J. Clin. Immunol. 40, 267–276 (2020).
Lee, K., Vyas, Y., Garner, C. C. & Montgomery, J. M. Autism-associated Shank3 mutations alter mGluR expression and mGluR-dependent but not NMDA receptor-dependent long-term depression. Synapse 73, e22097 (2019).
Leblond, C. S. et al. Meta-analysis of SHANK Mutations in Autism Spectrum Disorders: A gradient of severity in cognitive impairments. PLoS Genet. 10, e1004580 (2014).
Guilmatre, A., Huguet, G., Delorme, R. & Bourgeron, T. The emerging role of SHANK genes in neuropsychiatric disorders. Dev. Neurobiol. 74, 113–122 (2014).
Bey, A. L. et al. Subacute Neuropsychiatric Syndrome in Girls With SHANK3 Mutations Responds to Immunomodulation. Pediatrics 145, e20191490. https://doi.org/10.1542/peds.2019-1490 (2020).
Kohlenberg, T. M. et al. Psychiatric illness and regression in individuals with Phelan-McDermid syndrome. J. Neurodev Disord. 12, 7–020–9309–6 (2020).
Redin, C. et al. Efficient strategy for the molecular diagnosis of intellectual disability using targeted high-throughput sequencing. J. Med. Genet. (2014).
Berryer, M. H. et al. Mutations in SYNGAP1 cause intellectual disability, autism, and a specific form of epilepsy by inducing haploinsufficiency. Hum. Mutat. 34, 385–394 (2013).
Santos-Gómez, A. et al. Disease-associated GRIN protein truncating variants trigger NMDA receptor loss-of-function. Hum. Mol. Genet. (2020).
Liao, X. & Li, Y. Genetic associations between voltage-gated calcium channels and autism spectrum disorder: A systematic review. Mol. Brain 13, 96–020–00634–0 (2020).
Ryan, T. J. et al. Evolution of GluN2A/B cytoplasmic domains diversified vertebrate synaptic plasticity and behavior. Nat. Neurosci. 16, 25–32 (2013).
Seo, B. A., Lee, J. H., Kim, H. M. & Kang, M. G. Neuronal calcium channel α(1) subunit interacts with AMPA receptor, increasing its cell surface localisation. Biochem. Biophys. Res. Commun. 498, 402–408 (2018).
Kristinsson, S. Y. et al. Autoimmunity and risk for Hodgkin’s lymphoma by subtype. Haematologica 94, 1468–1469 (2009).
Urayama, K. Y. et al. Genome-wide association study of classical Hodgkin lymphoma and Epstein-Barr virus status-defined subgroups. J. Natl. Cancer Inst. 104, 240–253 (2012).
Nanda, H., Ponnusamy, N., Odumpatta, R., Jeyakanthan, J. & Mohanapriya, A. Exploring genetic targets of psoriasis using genome wide association studies (GWAS) for drug repurposing. 3 Biotech 10, 43–019–2038–4. Epub 2020 Jan 10 (2020).
Li, B. et al. The role of Th17 cells in psoriasis. Immunol. Res. 68, 296–309 (2020).
Snowden, J. A., Badoglio, M. & Alexander, T. The rise of autologous HCT for autoimmune diseases: What is behind it and what does it mean for the future of treatment? An update on behalf of the EBMT Autoimmune Diseases Working Party. Expert Rev. Clin. Immunol. 15, 981–985 (2019).
Li, X. et al. GABRG2 deletion linked to genetic epilepsy with febrile seizures plus affects the expression of GABA(A) receptor subunits and other genes at different temperatures. Neuroscience 438, 116–136 (2020).
Jiang, Y. L. et al. Clinical utility of exome sequencing and reinterpreting genetic test results in children and adults with epilepsy. Front. Genet. 11, 591434 (2020).
Li, J. et al. Mutation in ε-Sarcoglycan induces a myoclonus-dystonia syndrome-like movement disorder in mice. Neurosci. Bull. (2020).
Nedergaard, M. & Goldman, S. A. Glymphatic failure as a final common pathway to dementia. Science 370, 50–56 (2020).
Manouchehrian, O., Ramos, M., Bachiller, S., Lundgaard, I. & Deierborg, T. Acute systemic LPS-exposure impairs perivascular CSF distribution in mice. J. Neuroinflam. 18, 34-021-02082-6 (2021).
Delgado-Alvarado, M. et al. A novel SGCE variant is associated with myoclonus-dystonia with phenotypic variability. Neurol. Sci. 41, 3779–3781 (2020).
Peall, K. J. et al. SGCE mutations cause psychiatric disorders: Clinical and genetic characterization. Brain 136, 294–303 (2013).
LeDoux, M. S. Population prevalence of deleterious SGCE variants. Tremor Other Hyperkinet Mov. (N. Y) 10, 50 (2020).
Fernández-Marmiesse, A. et al. A novel missense mutation in GRIN2A causes a nonepileptic neurodevelopmental disorder. Mov. Disord. 33, 992–999 (2018).
Cocoș, R. et al. CACNA1B gene variants in adult-onset isolated focal dystonia. Neurol. Sci. (2020).
Lewin, A. B., Storch, E. A. & Murphy, T. K. Pediatric autoimmune neuropsychiatric disorders associated with Streptococcus in identical siblings. J. Child Adolesc. Psychopharmacol. 21, 177–182 (2011).
Gan, L. et al. Twins discordant for myositis and systemic lupus erythematosus show markedly enriched autoantibodies in the affected twin supporting environmental influences in pathogenesis. BMC Musculoskelet. Disord. 15, 67–2474–15–67 (2014).
Moretti, R. et al. Blood-brain barrier dysfunction in disorders of the developing brain. Front. Neurosci. 9, 40 (2015).
Ribot JC, Lopes N, Silva-Santos B. γδ T cells in tissue physiology and surveillance. Nat. Rev. Immunol. 21(4):221–232 (2021)
Alves de Lima, K. et al. Meningeal γδ T cells regulate anxiety-like behavior via IL-17a signaling in neurons. Nat. Immunol. 21, 1421–1429 (2020).
Huett, A., Leong, J. M., Podolsky, D. K. & Xavier, R. J. The cytoskeletal scaffold Shank3 is recruited to pathogen-induced actin rearrangements. Exp. Cell Res. 315, 2001–2011 (2009).
Zhen, H. et al. Wip1 regulates blood-brain barrier function and neuro-inflammation induced by lipopolysaccharide via the sonic hedgehog signaling signaling pathway. Mol. Immunol. 93, 31–37 (2018).
Lombardo, M. V. et al. Maternal immune activation dysregulation of the fetal brain transcriptome and relevance to the pathophysiology of autism spectrum disorder. Mol. Psychiatry 23, 1001–1013 (2018).
Spann, M. N., Monk, C., Scheinost, D. & Peterson, B. S. Maternal immune activation during the third trimester is associated with neonatal functional connectivity of the salience network and fetal to toddler behavior. J. Neurosci. 38, 2877–2886 (2018).
Han, V. X., Patel, S., Jones, H. F. & Dale, R. C. Maternal immune activation and neuroinflammation in human neurodevelopmental disorders. Nat. Rev. Neurol. (2021).
Lünemann, J. D., Nimmerjahn, F. & Dalakas, M. C. Intravenous immunoglobulin in neurology–mode of action and clinical efficacy. Nat. Rev. Neurol. 11, 80–89 (2015).
Norris, P. A. A., Kaur, G. & Lazarus, A. H. New insights into IVIg mechanisms and alternatives in autoimmune and inflammatory diseases. Curr. Opin. Hematol. 27, 392–398 (2020).
Jones, H. F. et al. Maternal thyroid autoimmunity associated with acute-onset neuropsychiatric disorders and global regression in offspring. Dev. Med. Child Neurol. 61, 984–988 (2019).
Lin, J. R. et al. Integrated post-GWAS analysis sheds new light on the disease mechanisms of schizophrenia. Genetics 204, 1587–1600 (2016).
Sekar, A. et al. Schizophrenia risk from complex variation of complement component 4. Nature 530, 177–183 (2016).
Hafizi, S., Tabatabaei, D. & Lai, M. C. Review of clinical studies targeting inflammatory pathways for individuals with autism. Front. Psychiatry. 10, 849 (2019).
Sabourin, K. R. et al. Infections in children with autism spectrum disorder: Study to explore early development (SEED). Autism Res. 12, 136–146 (2019).
Mehra, C. et al. Childhood disintegrative disorder and autism spectrum disorder: A systematic review. Dev. Med. Child Neurol. 61, 523–534 (2019).
Yin, J. et al. Next generation sequencing of 134 children with autism spectrum disorder and regression. Genes (Basel) 11, 853. https://doi.org/10.3390/genes11080853 (2020).
Dantzer, R. Neuroimmune interactions: From the brain to the immune system and vice versa. Physiol. Rev. 98, 477–504 (2018).
Niesler, B., Kuerten, S., Demir, I. E. & Schäfer, K. H. Disorders of the enteric nervous system—a holistic view. Nat. Rev. Gastroenterol. Hepatol. (2021).
Oshima, T. et al. Wip1 protects hydrogen peroxide-induced colonic epithelial barrier dysfunction. Cell Mol. Life Sci. 64, 3139–3147 (2007).
Quagliariello, A. et al. Gut microbiota profiling and gut-brain crosstalk in children affected by pediatric acute-onset neuropsychiatric syndrome and pediatric autoimmune neuropsychiatric disorders associated with streptococcal infections. Front. Microbiol. 9, 675 (2018).
Loffredo, L. et al. Oxidative stress and gut-derived lipopolysaccharides in children affected by paediatric autoimmune neuropsychiatric disorders associated with streptococcal infections. BMC Pediatr. 20, 127–020–02026–8 (2020).
Wei, S. C. et al. SHANK3 Regulates Intestinal Barrier Function Through Modulating ZO-1 Expression Through the PKCε-dependent Pathway. Inflamm. Bowel Dis. 23, 1730–1740 (2017).
Pellegrini, C. et al. Microbiota-gut-brain axis in health and disease: Is NLRP3 inflammasome at the crossroads of microbiota-gut-brain communications?. Prog. Neurobiol. 191, 101806 (2020).
Opipari, A. & Franchi, L. Role of inflammasomes in intestinal inflammation and Crohn’s disease. Inflamm. Bowel Dis. 21, 173–181 (2015).
Erikson, G. A. et al. Whole-genome sequencing of a healthy aging cohort. Cell 165, 1002–1011 (2016).
Drmanac, R. et al. Human genome sequencing using unchained base reads on self-assembling DNA nanoarrays. Science 327, 78–81 (2010).
Acknowledgements
The authors want to thank EXPAND for directing researchers to PANS cases throughout the European Union, and Erika Pedrosa and Stefanie Mittelstadt for copy editing the manuscript. HML is supported by the National Institute of Child Health and Human Development NIH/NICHD; P30 HD071593 to the Albert Einstein College of Medicine’s Rose F. Kennedy Intellectual and Developmental Disabilities Research Center. The Lachman lab also receives support from the Janice C. Blanchard Family Fund. P.J. van der Spek is supported by EU H2020 grants, an ImmunAID grant (ID: 7792950), and a MOODSTRATIFICATION grant (ID: 754740). The Bioinformatics infrastructure and team is supported by grants from KWF, NWO/ZonMW and the Dutch Heart foundation through the BDVA initiated H2020 Bigmedilytics program on Personalized Medicine. Janet Cunningham is a Gullstrand Fellow at Uppsala University Hospital. The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript. The authors dedicate this paper to Dr. Paul Janssen whose ideas about the role of the immune system in tics and psychosis help form the framework for this research project, and to participating families for their courage and support.
Funding
This article was funded by National Institutes of Health (P30 HD071593), Janice C. Blanchard Family Fund, Gullstrand Fellow (EU 2020), immunAID grant, 7792950, MOODSTRATIFICATION (7547540), KWF, NWO/ZonMW, BDVA H2020 Bigmedilytics program on Personalized Medicine.
Author information
Authors and Affiliations
Contributions
R.T. characterized U.S. cohort; exome sequencing; H.L. wrote manuscript, analyzed data, generated supplemental figures; O.M. collected patient data from European cohort J.C. major contributor to manuscript; D.Z. prepared figure S2, edited manuscript; A.Sp. ascertained patients in European cohort; P.C. ascertained patients in European cohort and carried out DNA analysis of family members; A.Sc. data analysis European cohort; R.W. edited manuscript, generated figure 1; S.S. analyzed genetic data in European cohort helped create figures 2-4; P.S. analyzed genetic data European cohort, helped create figures 2-4.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Trifiletti, R., Lachman, H.M., Manusama, O. et al. Identification of ultra-rare genetic variants in pediatric acute onset neuropsychiatric syndrome (PANS) by exome and whole genome sequencing. Sci Rep 12, 11106 (2022). https://doi.org/10.1038/s41598-022-15279-3
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-022-15279-3
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
