
Abstract
Regular hepatocellular carcinoma (HCC) surveillance by ultrasonography in combination with the α-fetoprotein (AFP) examination is unsatisfactory in diagnostic sensitivity for early-stage HCC especially in cirrhotic patients. We conducted a prospective study in a tertiary medical center in Taiwan and consecutively collected serum samples from patients with chronic hepatitis, liver cirrhosis (LC), or HCC for new biomarker discovery. Overall, 166 patients were enrolled, including 40 hepatitis, 30 LC, and 96 HCC. Four acute-phase serum amyloid A (A-SAA) derived biomarkers including total A-SAA, A-SAA monomer and oligomer, and protein misfolding cyclic amplification (PMCA) signal were measured and compared between patients with and without HCC. A-SAA biomarkers significantly increased in the HCC group when compared to the hepatitis and LC groups, and generally increased in more advanced tumor stages. Among A-SAA biomarkers, the area under the receiver operator characteristic curves (AUROCs) for PMCA signal in discrimination of all-stage and early-stage HCC were 0.86 and 0.9 in cirrhotic patients, which is comparable to AFP. For cirrhotic patients with low AFP (< 7 ng/mL), PMCA signal maintained good capacity in prediction of early-stage HCC (AUROC: 0.94). Serum A-SAA and its prion-like property showed a potential to complement AFP in detection of early-stage HCC.
Similar content being viewed by others

Introduction
Hepatocellular carcinoma (HCC) is the most common primary liver cancer and the third leading cause of cancer-related death worldwide1. The incidence of HCC depends on variable prevalence of chronic hepatitis B virus (HBV) and hepatitis C virus (HCV) infection2. Liver cirrhosis developed after prolonged hepatic injuries caused by HBV or HCV infections, alcoholic hepatitis, nonalcoholic steatohepatitis, or autoimmune hepatitis3. Cirrhosis is the major risk for HCC, and approximately 80% to 90% of HCC patients already have established cirrhosis4. The Barcelona Clinic Liver Cancer (BCLC) system classifies HCC into five stages based on the number, size, and extension of tumors, liver reserve, and the overall performance status of the patients5. Currently, curative therapies are only feasible for very early and early-stage (BCLC stage 0-A) HCC6, suggesting the importance of early HCC detection. At present, the clinical guidelines of European Association for the Study of the Liver (EASL)6 and American Association for the Study of Liver Diseases (AASLD)7 recommend the high-risk patients to receive abdominal ultrasound imaging every six months with or without the measurement of the α-fetoprotein (AFP) level. However, ultrasonography is limited by its image quality and operator-dependent8. Serum AFP level remains normal in over 30% HCC patients9. The sensitivity of ultrasonography for HCC detection with or without consideration of the AFP level was 63% or 45%, respectively, which was unsatisfactory10. There is an urgent need to develop new biomarkers for early HCC detection.
Serum amyloid A (SAA) is a family of positive acute-phase reactants produced by hepatocytes11. Among four family members, SAA1 and SAA2, also referred to as “acute-phase SAA” (A-SAA), share high sequence identity12 and the expression increases significantly during an acute-phase response13. As an amyloid protein, A-SAA is prone to aggregate and form insoluble fibrils with cross-β structures and these fibrils can be probed by an amyloid dye thioflavin T (ThT)14. Furthermore, formation of amyloid fibrils is inducible and can be amplified by a method called protein misfolding cyclic amplification (PMCA)15. The serum SAA level increases in several malignancies including lung16, pancreas17, colon18, kidney19,20, and stomach21. SAA has also been detected in HCC cell lines and tissues22,23,24. The serum SAA level positively correlates with tumor stages25,26. Serum SAA can be used to monitor the recurrence of nasopharyngeal cancer and breast cancer after treatments27,28 and to predict the overall survival29. Serum SAA intensifies in HBV-related HCC30, and may serve as a prognostic marker for HCC31. These results suggest SAA is a pan-cancer marker; however, the contribution of A-SAA among the four SAAs was not specified. In addition, the role of A-SAA in detection of early-stage HCC remains unclear.
Here, we aimed to evaluate the potential of A-SAA as a biomarker for HCC. We first measured and analyzed the concentration of total serum A-SAA in patients with hepatitis, liver cirrhosis, and HCC. Then we specifically examined A-SAA monomer and oligomer of the three patient groups. Since SAA is an amyloidogenic protein, by using the amyloidogenic property we further investigated the capacity of a method namely protein misfolding cyclic amplification (PMCA), with prion-like amplification of A-SAA to discriminate HCC from non-HCC groups. PMCA is a method to allow amplification of the signal in body fluids15. Finally, we evaluated the predictability of A-SAA in patients with low AFP level and found PMCA signal maintained good capacity in prediction of early-stage HCC.
Results
Total A-SAA concentration is significantly higher in HCC patients than in hepatitis and LC patients
To assess the potential of A-SAA as an HCC biomarker, a total of 166 patients were prospectively enrolled, and they were categorized into three groups: hepatitis (n = 40), LC (n = 30), and HCC (n = 96). Their demographic and clinical characteristics are listed in Table 1. The mean age was 47 in the hepatitis group, compared to 60 in patients of the LC or the HCC groups. There were 43%, 63%, and 72% of male in the hepatitis, LC, and HCC groups, respectively. Regarding the etiology, more than 63% of patients in individual groups had HBV infection. Among the HCC group, 86 patients (90%) had cirrhosis, and 42 patients (44%) had early-stage HCC (BCLC 0 and A). We first employed ELISA for measurement of total A-SAA concentration in the serum collected from the three patient groups. Among the three groups, total A-SAA concentration in the HCC patients (mean ± standard deviation: 223.8 ± 76.0 ng/mL) was significantly higher than in the hepatitis (176.3 ± 73.2 ng/mL, P = 0.0013) and LC (167.8 ± 40.6 ng/mL, P = 0.0006) patients (Fig. 1a). No significant difference was found in total A-SAA concentration between the hepatitis and LC cohorts.

Comparison of A-SAA derived biomarker levels among hepatitis, LC, and HCC patients. A total of 166 serum samples from 40 hepatitis (indicated as H), 30 LC, and 96 HCC patients were measured and analyzed. (a) Total A-SAA. (b) A-SAA monomer. (c) A-SAA oligomer. The statistical analysis was performed by one-way ANOVA and Tukey’s Post Hoc Test, where P < 0.05 (*), < 0.01 (**), and < 0.001 (***). NS not significant.
A-SAA monomer, but not the oligomer, differentiates HCC from LC
Due to the amyloidogenic property of A-SAA, we specifically examined the serum samples in Western blot to observe the possible association between different SAA assemblies and the three patient groups. The serum samples for SDS-PAGE were prepared without the addition of reducing agent β-mercaptoethanol and boiling to preserve oligomeric state of A-SAA. The Western blot results showed two main assemblies of A-SAA proteins in patients’ sera, A-SAA monomer migrated at ~ 9 kDa and its high-molecular-weight oligomer at ~ 130 kDa (Supplementary Fig. S1a). Since monomeric A-SAAs in patients’ sera displayed a size smaller than the theoretic molecular mass of SAA1 (11.7 kDa) and SAA2 (11.6 kDa), we compared the sera along with our purified recombinant full-length human SAA1 in Western blot (Supplementary Fig. S2). The result showed that the serum A-SAA migrated in similar distance to the recombinant SAA1 confirming that serum A-SAA detected is a full-length monomer. The faster migration in SDS-PAGE may be due to protein conformation and/or hydrophobicity.
After Western blot, we quantified the amount of A-SAA monomer and oligomer for further analysis. Since aggregation is a concentration-dependent event, we wondered if the presence of oligomers was due to the increase of A-SAA concentration in serum. Therefore, we conducted a correlation analysis on A-SAA monomer and oligomer. A Spearman’s coefficient of 0.296 (P = 0.0001) indicated only a modest positive correlation between A-SAA monomer and oligomer (Supplementary Fig. S1b). Next, we compared the level of both monomer and oligomer of A-SAA among the three groups. The level of A-SAA monomer was significantly higher in the HCC group (3.8 ± 4.6 fold) when compared to that in the hepatitis (1 ± 1.2 fold, P = 0.0002) and the LC (1 ± 1.5 fold, P = 0.0008) groups (Fig. 1b). No significant difference was found in A-SAA monomer level between the hepatitis and LC groups. The result is consistent with the ELISA result. For A-SAA oligomer, the HCC group (1.6 ± 1.2 fold) had a significantly higher level compared to the hepatitis group (1 ± 0.6 fold, P = 0.0125) but not to the LC group (1.3 ± 0.8 fold, P = 0.4486) (Fig. 1c).
PMCA of SAA1 showed potential to differentiate HCC from LC
PMCA is a method that utilizes nucleation-dependent amyloid aggregation process to accelerate the conversion of monomeric amyloid protein into fibrils15. The principle of PMCA relies on the application of ultrasound waves to fragment the preformed amyloid fibrils, increasing the amount of misfolded seeds present in the samples containing normal monomeric protein and further induce the amplification of aggregation or accelerate the aggregation reaction (Fig. 2a). PMCA has been applied to detect variant Creutzfeldt-Jakob disease and demonstrated a diagnostic capacity with high sensitivity and specificity15,32. Recently, PMCA has also been applied to differentiate α-synuclein in Parkinson’s disease and multiple system atrophy33. Since SAA also possesses amyloidogenic property, we performed PMCA on SAA and examined the potential of PMCA as detection method for HCC. Our purified recombinant full-length human SAA1 was used as a substrate and serum from each patient as seeds for PMCA. The patient’s serum was mixed with purified SAA1 in a volume ratio of 1:20 in a solution containing amyloid dye ThT that recognized amyloid fibrils. The samples underwent 24 PMCA cycles in which each cycle comprised 29 min 20 s of incubation followed by a 40-s sonication pulse. The final ThT fluorescence was detected and normalized to the signal in hepatitis group. Our results showed that patients with HCC had an increased PMCA signal (1.1 ± 0.2 fold) in comparison with the hepatitis (1 ± 0.1 fold, P = 0.0129) and LC (0.9 ± 0.1 fold, P < 0.0001) patients (Fig. 2b). Meanwhile, we investigated the relationship of these A-SAA biomarkers and clinical characteristics. The A-SAA biomarkers were not associated with age, sex, HBV or HCV infection, except a significantly higher PMCA signal in HBV carriers (P < 0.05) (Supplementary Fig. S3). There was low to moderate correlation between AST/ALT and the total A-SAA or A-SAA monomer levels, but the A-SAA oligomer, or the PMCA signal did not correlate with ALT level, indicating A-SAA might be associated with liver inflammation but the correlation was low. The A-SAA biomarkers did not correlate with liver fibrosis measured by the Fib-4 index. (Supplementary Fig. S4).

Comparison of PMCA signals among hepatitis, LC, and HCC patients. (a) Diagram illustrating the principle of PMCA technique. A standard PMCA procedure starts with incubation of oligomeric seeds with monomeric amyloid protein to allow growth of the polymers. The aggregates are then fragmented by sonication to increase the number of seeds for further aggregate growth. After repeating PMCA cycles, the concentration of initial preformed oligomeric protein can be amplified and thus easy to be detected. (b) Comparison of PMCA signals among hepatitis, LC, and HCC patients. A total of 146 serum samples from 30 hepatitis (indicated as H), 30 LC, and 86 HCC patients were measured and analyzed. The statistical analysis was performed by one-way ANOVA and Tukey’s Post Hoc Test, where P < 0.05 (*) and < 0.0001 (****). NS not significant.
Correlation between A-SAA derived biomarkers and HCC stages
The level of A-SAA biomarkers including total A-SAA, A-SAA monomer, A-SAA oligomer, and A-SAA PMCA signal was further compared within HCC patients. The mean concentration of total A-SAA for HCC patients at BCLC stage 0, A, B, C, and D were 195.7 ± 70.9, 192.7 ± 57.9, 242.7 ± 63.9, 244.8 ± 99.8, and 284.5 ± 53.5 ng/mL, respectively. No significant difference between any two BCLC stages was detected (Fig. 3a). The mean level of A-SAA monomer increased significantly in BCLC stage B (5.2 ± 4.6 fold), stage C (5.8 ± 5.3 fold), and stage D (8.8 ± 6.0 fold) when compared to BCLC stage 0 (0.5 ± 0.4 fold, P = 0.0163, 0.0075, and 0.0025), or BCLC stage A (1.7 ± 2.9 fold, P = 0.0139, 0.0061, and 0.0042) (Fig. 3b). The mean level of A-SAA oligomer also significantly increased in BCLC stage C (2.2 ± 1.1 fold) when compared to BCLC stage 0 (1.0 ± 1.0 fold, P = 0.0366) and stage A (1.1 ± 0.5 fold, P = 0.0052) (Fig. 3c). The PMCA signals were comparable at different BCLC stages without significance detected between any two groups (Fig. 3d). Overall, A-SAA monomer and oligomer showed an increased level in more advanced stage HCC in comparison with early-stage HCC.

The level of A-SAA derived biomarkers at different HCC BCLC stages. A total of 96 serum samples from stage 0 (n = 11), stage A (n = 31), stage B (n = 28), stage C (n = 21), and stage D (n = 5) were measured and analyzed. (a) Total A-SAA. (b) A-SAA monomer. (c) A-SAA oligomer. (d) PMCA signal. The statistical analysis was performed by one-way ANOVA and Tukey’s Post Hoc Test, where P < 0.05 (*) and < 0.01 (**).
The prediction of all-stage and early-stage HCC in all patients using A-SAA derived biomarkers
Receiver Operating Characteristic (ROC) analysis was performed to assess the prediction value of A-SAA derived biomarkers in discriminating all-stage and early-stage HCC in all patients (Fig. 4). As a comparison, among all the biomarkers, AFP showed the highest area under the ROC curve (AUROC) of 0.9 (95% CI 0.84–0.95, P < 0.0001) to distinguish all-stage HCC (Supplementary Table S1). The best A-SAA biomarker was PMCA signal which demonstrated an AUROC of 0.77 (95% CI 0.7–0.85, P < 0.0001) (Fig. 4a and Supplementary Table S1). With the optimal cut-off values, AFP showed a higher sensitivity of 78% compared to 58% for PMCA signal with similar specificities of 93% and 92%, respectively.

ROC curves of A-SAA derived biomarkers in differentiating all-stage and early-stage HCC in all patients. (a) Detection of all-stage HCC. (b) Detection of early-stage HCC. AUROC value for each curve is shown in parentheses.
To further investigate the performance of A-SAA biomarkers in detecting early-stage HCC, we only included 42 early-stage HCC (BCLC 0-A) patients along with hepatitis and LC patients in this subgroup analysis. Among all the biomarkers, the highest AUROC was 0.89 (95% CI 0.81–0.96, P < 0.0001) for AFP followed by PMCA signal (0.82; 95% CI 0.74–0.91, P < 0.0001) (Fig. 4b and Supplementary Table S1). The sensitivity of AFP was 74% compared to 68% of PMCA signal with specificity of 93% and 92%, respectively. To summarize, for all patients, AFP demonstrated the best performance in detection of all-stage HCC whereas PMCA signal showed a comparable capacity in early-stage HCC prediction.
The prediction of HCC using A-SAA derived biomarkers among cirrhotic patients
Since patients with cirrhosis had the highest risk of HCC, we investigated the HCC predictability of A-SAA biomarkers among cirrhotic patients. From 166 patients, we excluded 50 non-cirrhotic patients and only included 116 cirrhotic patients in this subgroup analysis. We found a significantly lower concentration of total A-SAA in the LC group when compared to the intermediate (BCLC B, P = 0.0005) and the advanced HCC group (BCLC C-D, P < 0.0001), but not to the early HCC group (BCLC 0-A, P = 0.2569) (Fig. 5a). A-SAA monomer showed a pattern similar to total A-SAA with a significantly lower level in the LC group when compared to the intermediate (P = 0.0012) and the advanced HCC groups (P < 0.0001), but not to the early HCC group (P = 0.9021) (Fig. 5b). For A-SAA oligomer, a significantly lower level was found in the LC group only when compared to the advanced HCC group (P = 0.0065) (Fig. 5c). PMCA signal of the LC group showed a significantly lower level when compared to the early (P < 0.0001), the intermediate (P = 0.0001), and the advanced HCC groups (P = 0.0399) (Fig. 5d). Together, among the A-SAA derived biomarkers, PMCA signal is the only one able to differentiate LC and the early-stage HCC.

Comparison of A-SAA derived biomarkers in patients with cirrhosis. A total of 116 serum samples from 30 LC and 86 HCC patients were measured and analyzed. Early HCC indicates BCLC stage 0-A (n = 38). Intermediate HCC indicates BCLC stage B (n = 24). Advanced HCC indicates BCLC stage C-D (n = 24). (a) Total A-SAA. (b) A-SAA monomer. (c) A-SAA oligomer. (d) PMCA signal. The statistical analysis was performed by one-way ANOVA and Tukey’s Post Hoc Test, where P < 0.05 (*), < 0.01 (**), < 0.001 (***), and < 0.0001 (****). NS not significant.
For detection of all-stage HCC in cirrhotic patients, the highest AUROCs were comparable for PMCA signal (0.86; 95% CI 0.79–0.93, P < 0.0001) and AFP (0.84; 95% CI 0.76–0.92, P < 0.0001) (Fig. 6a and Supplementary Table S2). The HCC detection sensitivity at the optimal cut-off values was also similar for PMCA signal (81%) and AFP (80%) as well as their specificity (80% vs. 83%, respectively). For early-stage HCC detection, AFP and PMCA signal again had a statistically comparable AUROC of 0.81 (95% CI 0.7–0.93, P = 0.0002) and 0.91 (95% CI 0.84–0.98, P < 0.0001), respectively (Fig. 6a and Supplementary Table S2). The sensitivities were close for AFP (74%) and PMCA signal (73%) with a higher specificity for PMCA signal (97%) compared to AFP (83%). In patients with cirrhosis, PMCA signal and AFP showed a similar detection capacity for all-stage HCC. For early HCC detection, with their optimal cut-off values, PMCA signal demonstrated a better performance than AFP with a similar sensitivity but a higher specificity.

ROC curves of A-SAA derived biomarkers in discrimination of all-stage and early-stage HCC in cirrhotic patients with or without low AFP level. (a) Detection of HCC in all cirrhotic patients. (b) Detection of early-stage HCC in cirrhotic patients with low AFP. AUROC value for each curve is shown in parentheses.
Performance of A-SAA derived biomarkers in cirrhotic patients with low AFP level
Because it is difficult to detect HCC in patients with low AFP level, and A-SAA biomarkers, especially PMCA signal, had better HCC predictability in cirrhotic patients, we only included the patients with both cirrhosis and low AFP (< 7 ng/mL, optimal cut-off decided by AUROC) in this subgroup analysis. For all-stage HCC detection, PMCA signal again showed the highest AUROC of 0.93 (95% CI 0.85–1, P < 0.0001) followed by total A-SAA (0.79; 95% CI 0.63–0.94, P = 0.0017) (Fig. 6b and Supplementary Table S2). The sensitivity and specificity for PMCA signal was 81% and 88%. For early-stage HCC detection, the AUROC of PMCA signal reached 0.94 (95% CI 0.86–1, P < 0.0001) with a sensitivity and specificity of 90% and 81% (Fig. 6b and Supplementary Table S2).
Furthermore, we performed multivariable logistic regression analysis to evaluate the association between PMCA signal and HCC after adjusted to age, gender, and AST/ALT concentration (Supplementary Table S3). We separated the PMCA signals for all cases in quartiles (PMCA signal < 0.91 fold [Quartile 1, Q1], 0.91–1.02 fold [Q2], 1.02–1.14 fold [Q3], and ≥ 1.14 fold [Q4]). The adjusted odds ratios for Q2 to Q4 are 8.43 (95% CI 2.01–44.77), 15 (95% CI 3.86–74.36), and 101 (95% CI 19.61–753.6) for all-stage HCC, respectively. After excluding the intermediate and advanced-stage HCC, the PMCA signals were separated in quartiles where the signal < 0.89 [Q1], 0.89–1.00 fold [Q2], 1.00–1.13 fold [Q3], and ≥ 1.13 fold [Q4]. The adjusted odds ratios to predict early-stage HCC for Q2 to Q4 are 17.43 (95% CI 2.32–339.5), 25.96 (95% CI 3.49–512.8), and 344.8 (95% CI 38.47–8286), respectively. After adjustment to confounders, PMCA signal still maintained strong association with all-stage and early-stage HCC.
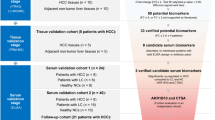
To conclude, among A-SAA biomarkers, PMCA signal demonstrated the highest HCC detection capacity, especially in the subgroup of cirrhotic patients with low AFP level. Based on our results, we designed a surveillance flow chart to demonstrate HCC detection focused on cirrhotic patients by AFP in combination of PMCA signal (Supplementary Fig. S5). We began with cirrhotic patients including 30 LC and 79 HCC cases. With AFP lower than 7 ng/mL, there are still 16 out of 79 HCC patients (approximately 20% of all HCC patients) remained undetected. In the subgroup of cirrhotic patients, the optimal cut-off value for PMCA signal decided by AUROC was 0.9 fold. In the following detection by PMCA signal with the cut-off, only 1 HCC cases (approximately 1% of all HCC patients) remained undetected.
Discussion
SAA has been shown to increase in the patients with different cancers and correlate with the progression of tumorigenesis. The majority of studies shows the value of SAA as a biomarker in prognosis of cancers31,34,35,36. In the present study, we attempted to investigate the association between SAA and different stages of liver diseases, especially among three closely related pathogenic conditions: hepatitis, liver cirrhosis, and HCC, and the value of SAA for early HCC detection. From our results, A-SAA derived biomarkers generally showed a higher level in the HCC group compared to the hepatitis and the LC groups. Furthermore, the level of A-SAA monomer increased in more advanced HCC (stage B, C, and D), compared to early HCC (stage 0 and A). The increase in SAA level in more advanced HCC suggests that SAA may not only be an indicator but also be involved in disease progression.
Expression of SAA, especially A-SAA, is induced and regulated by proinflammatory cytokines produced by a variety of immune cells, which are abundant in the tumor microenvironment37,38. Moreover, cancer cells also have the capability to produce inflammatory cytokines39. SAA has been shown to stimulate the production of matrix metalloproteinases (MMP)40 that is positively associated with tumor progression41,42. The alteration and degradation of extracellular matrix by MMPs is also closely related to tumor initiation and development43. Besides, SAA has been shown to induce tumor cell migration44 and angiogenesis45, which are involved in cancer exacerbation. Therefore, the significant increase in serum A-SAA of the HCC group is likely to be a feedback loop between SAA and hepatocarcinogenesis. However, the mechanistic insights for the current study are still unclear and required further intensive investigation.
The unique aspect of our study is that we utilized PMCA in combination with the prion-like property of A-SAA as a predictive method for HCC. PMCA method was originally designed to detect amyloidosis-related diseases46 and has never been applied and assessed for cancer detection. The basic principle of PMCA relies on an amyloidogenic protein can serve as a seed and induces structural transformation of the native substrate proteins to form ThT dye-recognizable aggregates. From our results, PMCA method showed a better capacity for HCC discrimination compared to total A-SAA measured by ELISA or A-SAA monomer detected by Western blot, especially for early-stage HCC. Since the cancer patients contain more serum A-SAA as evidenced by the level of A-SAA monomer and seeding-liable oligomer, the PMCA procedure may further amplify the signals to enhance differences between the HCC and non-cancer groups. Besides, a more amyloidogenic conformation of A-SAA may be adopted due to the aberrant environment for protein folding or maturation in cancer cells. However, it is possible that the amplification can be induced by other amyloidogenic proteins, but not SAA, in the serum although the hetero-seeding is in general less efficient.
Cirrhotic patients had high risk to develop HCC; however, the low sensitivity of ultrasonography to detect early HCC in cirrhotic background remains a challenge to clinicians. Previous studies have shown that the sensitivity and specificity of AFP for HCC surveillance largely depended on the selected serum level threshold47. In our study, even with low threshold of AFP (< 7 ng/mL), approximately 20% of HCC patients still cannot be detected. From our analysis, the PMCA method demonstrated the best HCC predictability in cirrhotic patients with low AFP. The PMCA method may be applied for HCC surveillance in this clinical scenario. However, there are limitations to the present study. In this study, we only included hepatitis, LC, and HCC patients with the background of HBV or HCV infection. Therefore, we do not know the A-SAA level in healthy control, or hepatitis patients with other etiologies, e.g., non-alcoholic fatty liver disease (NAFLD), autoimmune hepatitis (AIH), or alcoholic liver disease. Moreover, since the case numbers are small in this study, further large scale, prospective validation is needed.
In conclusion, by using several methods to investigate HCC predictive capacity of A-SAA, we found that A-SAA derived biomarkers, especially PMCA method, have potential to facilitate HCC detection. Even though currently AFP may still be the main serum biomarker for HCC surveillance, A-SAA biomarkers can be used as a complementary test in combination with imaging examination, especially for the cirrhotic patients with low AFP level.
Methods
Serum samples
This was a prospective study conducted at National Taiwan University Hospital (NTUH) since 2012 to investigate the biomarkers for HCC. Patients with chronic liver disease, liver cirrhosis, and HCC (with viable tumor) who were followed regularly in the liver clinic of NTUH were enrolled. Their sera were collected, processed, and stored at -80 °C until further use. Cirrhosis was diagnosed clinically by the nodular liver surface, coarse liver parenchymal texture, narrowed vessels with irregular intrahepatic vessel contour, and enlarged spleen size by abdominal ultrasonography48. HCC was diagnosed by either pathology or typical dynamic imaging studies according to the AASLD guideline. The liver function tests and AFP level were measured in the Department of Laboratory Medicine in NTUH. The study conformed to the ethical guidelines of the 1975 Declaration of Helsinki and was approved by the Institutional Review Boards of NTUH (201108073RC and 201207048RIB). All patients provided their written informed consents before enrolment.
Enzyme-linked immunosorbent assay (ELISA)
The assessment of total A-SAA concentration in the sera from hepatitis, LC, and HCC patients was conducted using Human SAA ELISA kit (ab100635, Abcam, Cambridge, MA, USA). One hundred µL of 20 × PBS-diluted serum sample was added into the antibody-pre-coated ELISA plate and incubated at 4 °C overnight. Next, 100 µL of biotinylated SAA detection antibody was applied to the plate and recognized by addition of 100 µL of horseradish peroxidase-conjugated streptavidin. Chromogenic substrate was added and the absorbance at 450 nm was then measured using SpectraMax M5 microplate reader (Molecular Devices, San Jose, CA, USA).
Western blot
Proteins in 20 × PBS-diluted serum samples were separated by 8–16% gradient SDS-PAGE precast gels (Bio-Rad, Hercules, CA, USA). To preserve oligomeric states of A-SAA during SDS-PAGE, the sample buffer did not contain β-mercaptoethanol and the samples were not boiled. A-SAA proteins were then recognized by an anti A-SAA antibody (ab687, Abcam) and a secondary antibody (goat anti-mouse IgG peroxidase conjugated antibody, Jackson ImmunoResearch Labs, West Grove, PA, USA) and visualized in a luminescence/fluorescence imaging system ImageQuant LAS 4000 biomolecular imager (GE Healthcare, Marlborough, MA, USA). The amount of A-SAA monomer and oligomer was quantified by ImageQuant TL 8.2 image analysis (GE Healthcare). The final results were calculated by normalization of the amount of A-SAA monomer and oligomer from each patient to the mean amount of the hepatitis group.
Expression and purification of recombinant human SAA1
Full-length human SAA1 (residues 1–104) was cloned into a pET-14b vector which contains an N-terminal histidine tag (His-tag) and expressed in Escherichia coli BL21 cells. A TEV protease cleavage site (ENLYFQS) was added to the N-terminus of mature SAA1 sequence for His-tag removal. Since TEV protease recognizes and cleaves between Q and S, SAA1 after cleavage contains an additional serine residue at the N-terminus. The overexpression of His-tagged SAA1 protein was induced at 37 °C by the addition of 1 mM isopropyl β-D-1-thiogalactopyranoside at mid-exponential growth phase of E. coli. After 5-h induction, the bacteria were harvested by centrifugation and lyzed in lysis buffer (20 mM Tris, pH 8.0, 5 M urea, 50 mM imidazole) by a microfluidizer (model M110L, Microfluidics, Westwood, MA, USA). His-tagged SAA1 protein was purified by HisTrap HP column (GE Healthcare) in the presence of 5 M urea. The His-tag was removed from SAA1 by incubation of His-tagged TEV protease in a TEV:SAA molar ratio of 1:20 at 30 °C for 16 h. The cleaved proteins were further purified by HisTrap HP column as the flow-through. The pure SAA1 protein was then dialyzed into a fresh 50 mM phosphate buffer (pH 7.0) and stored at − 80 °C for subsequent experiments.
Protein misfolding cyclic amplification (PMCA)
For PMCA experiment, the sera of patients and purified recombinant human SAA1 were used as seeds and substrates, respectively. Samples were prepared by mixing patient’s serum with purified SAA1 in a volume ratio of 1:20. As substrate, SAA1 protein was prepared to a final concentration of 20 μM in a 50 mM phosphate buffer (pH 7.0). A final concentration 10 μM of ThT was added into samples to monitor amyloid fibril formation of SAA1. One hundred µL of SAA1-serum sample was loaded into a 96-well ELISA plate and PMCA reaction was automatically performed by a microsonicator (model Q700, Qsonica, Newtown, CT, USA). One PMCA cycle comprised 29 min 20 s of incubation at 25 °C, followed by a 40-s sonication pulse at a potency of 300 to 310 W. A complete PMCA round consisted of 24 cycles that equals to 12 h. The fluorescent emission of ThT was measured at 485 nm using SpectraMax M3 microplate reader (Molecular Devices) with an excitation wavelength of 442 nm. The final results were calculated by normalization of PMCA signal from each patient to the mean signal value of the hepatitis group.
Statistical analysis
GraphPad Prism version 7.04 and 8.4.2 (GraphPad Software, San Diego, CA, USA) were utilized for statistical analyses. One-way ANOVA was used to evaluate the difference for more than two groups. Post-hoc analyses were conducted by Tukey’s multiple comparison between two groups. Correlation between two variables was measured by the Spearman’s correlation coefficient. Receiver operating characteristic (ROC) curves and area under the ROC curve (AUROC) were generated to assess the predictive capacity of each biomarker, and the Youden index to indicate the optimal threshold for each ROC curve. Multivariable logistic regression was used to evaluate the association of biomarkers with the outcome under adjustment of age, gender, and AST/ALT concentration. A P-value (two-tailed) less than 0.05 was considered a statistical significance.
Data availability
The datasets used and/or analyzed during the current study available from the corresponding author on reasonable request.
References
Sung, H. et al. Global cancer statistics 2020: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA 71, 209–249. https://doi.org/10.3322/caac.21660 (2021).
El-Serag, H. B. Epidemiology of viral hepatitis and hepatocellular carcinoma. Gastroenterology 142, 1264–1273. https://doi.org/10.1053/j.gastro.2011.12.061 (2012).
Schuppan, D. & Afdhal, N. H. Liver cirrhosis. Lancet 371, 838–851. https://doi.org/10.1016/s0140-6736(08)60383-9 (2008).
Davis, G. L. et al. Hepatocellular carcinoma: Management of an increasingly common problem. Proceedings 21, 266–280. https://doi.org/10.1080/08998280.2008.11928410 (2008).
Forner, A., Reig, M. & Bruix, J. Hepatocellular carcinoma. The Lancet 391, 1301–1314. https://doi.org/10.1016/s0140-6736(18)30010-2 (2018).
European Association for the Study of the Liver. EASL Clinical Practice Guidelines: Management of hepatocellular carcinoma. J Hepatol 69, 182–236. https://doi.org/10.1016/j.jhep.2018.03.019 (2018).
Heimbach, J. K. et al. AASLD guidelines for the treatment of hepatocellular carcinoma. Hepatology 67, 358–380. https://doi.org/10.1002/hep.29086 (2018).
Li, J. et al. Serum thioredoxin is a diagnostic marker for hepatocellular carcinoma. Oncotarget 6, 9551–9563. https://doi.org/10.18632/oncotarget.3314 (2015).
Charrière, B., Maulat, C., Suc, B. & Muscari, F. Contribution of alpha-fetoprotein in liver transplantation for hepatocellular carcinoma. World J. Hepatol. 8, 881–890. https://doi.org/10.4254/wjh.v8.i21.881 (2016).
Tzartzeva, K. et al. Surveillance imaging and alpha fetoprotein for early detection of hepatocellular carcinoma in patients with cirrhosis: A meta-analysis. Gastroenterology 154, 1706-1718.e1701. https://doi.org/10.1053/j.gastro.2018.01.064 (2018).
Coetzee, G. A. et al. Serum amyloid A-containing human high density lipoprotein 3. Density, size, and apolipoprotein composition. J. Biol. Chem. 261, 9644–9651 (1986).
Parmelee, D. C. et al. Amino acid sequence of amyloid-related apoprotein (apoSAA1) from human high-density lipoprotein. Biochemistry 21, 3298–3303. https://doi.org/10.1021/bi00257a008 (1982).
Zhang, Y., Zhang, J., Sheng, H., Li, H. & Wang, R. Acute phase reactant serum amyloid A in inflammation and other diseases. Adv. Clin. Chem. 90, 25–80. https://doi.org/10.1016/bs.acc.2019.01.002 (2019).
Naiki, H., Higuchi, K., Hosokawa, M. & Takeda, T. Fluorometric determination of amyloid fibrils in vitro using the fluorescent dye, thioflavin T1. Anal. Biochem. 177, 244–249. https://doi.org/10.1016/0003-2697(89)90046-8 (1989).
Moda, F. et al. Prions in the urine of patients with variant Creutzfeldt-Jakob disease. N. Engl. J. Med. 371, 530–539. https://doi.org/10.1056/NEJMoa1404401 (2014).
Biaoxue, R., Hua, L., Wenlong, G. & Shuanying, Y. Increased serum amyloid A as potential diagnostic marker for lung cancer: A meta-analysis based on nine studies. BMC Cancer 16, 836. https://doi.org/10.1186/s12885-016-2882-0 (2016).
Yokoi, K. et al. Serum amyloid A as a tumor marker in sera of nude mice with orthotopic human pancreatic cancer and in plasma of patients with pancreatic cancer. Int. J. Oncol. 27, 1361–1369 (2005).
Toriola, A. T. et al. Biomarkers of inflammation are associated with colorectal cancer risk in women but are not suitable as early detection markers. Int. J. Cancer 132, 2648–2658. https://doi.org/10.1002/ijc.27942 (2013).
Wood, S. L. et al. Association of serum amyloid A protein and peptide fragments with prognosis in renal cancer. Br. J. Cancer 103, 101–111. https://doi.org/10.1038/sj.bjc.6605720 (2010).
Fischer, K., Theil, G., Hoda, R. & Fornara, P. Serum amyloid A: A biomarker for renal cancer. Anticancer Res. 32, 1801–1804 (2012).
Chan, D. C. et al. Evaluation of serum amyloid A as a biomarker for gastric cancer. Ann. Surg. Oncol. 14, 84–93. https://doi.org/10.1245/s10434-006-9091-z (2007).
Ganapathi, M. K. et al. Role of interleukin-6 in regulating synthesis of C-reactive protein and serum amyloid A in human hepatoma cell lines. Biochem. Biophys. Res. Commun. 157, 271–277. https://doi.org/10.1016/s0006-291x(88)80043-3 (1988).
Thorn, C. F., Lu, Z. Y. & Whitehead, A. S. Tissue-specific regulation of the human acute-phase serum amyloid A genes, SAA1 and SAA2, by glucocorticoids in hepatic and epithelial cells. Eur. J. Immunol. 33, 2630–2639. https://doi.org/10.1002/eji.200323985 (2003).
Thorn, C. F., Lu, Z. Y. & Whitehead, A. S. Regulation of the human acute phase serum amyloid A genes by tumour necrosis factor-alpha, interleukin-6 and glucocorticoids in hepatic and epithelial cell lines. Scand. J. Immunol. 59, 152–158. https://doi.org/10.1111/j.0300-9475.2004.01369.x (2004).
Biran, H., Friedman, N., Neumann, L., Pras, M. & Shainkin-Kestenbaum, R. Serum amyloid A (SAA) variations in patients with cancer: Correlation with disease activity, stage, primary site, and prognosis. J. Clin. Pathol. 39, 794–797 (1986).
Li, J. et al. Purification, identification and profiling of serum amyloid A proteins from sera of advanced-stage cancer patients. J. Chromatog. B 889–890, 3–9. https://doi.org/10.1016/j.jchromb.2012.01.002 (2012).
Cho, W. C. et al. Identification of serum amyloid a protein as a potentially useful biomarker to monitor relapse of nasopharyngeal cancer by serum proteomic profiling. Clin. Cancer Res. 10, 43–52. https://doi.org/10.1158/1078-0432.ccr-0413-3 (2004).
Bobin-Dubigeon, C., Lefrançois, A., Classe, J. M., Joalland, M. P. & Bard, J. M. Paired measurement of serum amyloid A (SAA) and paraoxonase 1 (PON1) as useful markers in breast cancer recurrence. Clin. Biochem. 48, 1181–1183. https://doi.org/10.1016/j.clinbiochem.2015.07.020 (2015).
Lin, H. Y., Tan, G. Q., Liu, Y. & Lin, S. Q. The prognostic value of serum amyloid A in solid tumors: A meta-analysis. Cancer Cell Int. 19, 62. https://doi.org/10.1186/s12935-019-0783-4 (2019).
He, Q. Y. et al. Toward the proteomic identification of biomarkers for the prediction of HBV related hepatocellular carcinoma. J. Cell. Biochem. 103, 740–752. https://doi.org/10.1002/jcb.21443 (2008).
Ni, X. C. et al. Serum amyloid A is a novel prognostic biomarker in hepatocellular carcinoma. Asian Pac. J. Cancer Prev. 15, 10713–10718. https://doi.org/10.7314/apjcp.2014.15.24.10713 (2014).
Concha-Marambio, L. et al. Detection of prions in blood from patients with variant Creutzfeldt-Jakob disease. Sci. Transl. Med. 8, 370ra183. https://doi.org/10.1126/scitranslmed.aaf6188 (2016).
Shahnawaz, M. et al. Discriminating alpha-synuclein strains in Parkinson’s disease and multiple system atrophy. Nature 578, 273–277. https://doi.org/10.1038/s41586-020-1984-7 (2020).
Meng, Y. Q. et al. Preoperative level of serum amyloid A is superior to C-reactive protein in the prognosis of esophageal squamous cell carcinoma. Dis. Esophagus 27, 670–677. https://doi.org/10.1111/dote.12128 (2014).
Cho, W. C., Yip, T. T., Cheng, W. W. & Au, J. S. Serum amyloid A is elevated in the serum of lung cancer patients with poor prognosis. Br. J. Cancer 102, 1731–1735. https://doi.org/10.1038/sj.bjc.6605700 (2010).
Yang, M. et al. Serum amyloid A expression in the breast cancer tissue is associated with poor prognosis. Oncotarget 7, 35843–35852. https://doi.org/10.18632/oncotarget.8561 (2016).
De Buck, M. et al. The cytokine-serum amyloid A-chemokine network. Cytokine Growth Factor Rev. 30, 55–69. https://doi.org/10.1016/j.cytogfr.2015.12.010 (2016).
Setrerrahmane, S. & Xu, H. Tumor-related interleukins: Old validated targets for new anti-cancer drug development. Mol. Cancer 16, 153. https://doi.org/10.1186/s12943-017-0721-9 (2017).
Dunlop, R. J. & Campbell, C. W. Cytokines and advanced cancer. J. Pain Symptoms Manage. 20, 214–232. https://doi.org/10.1016/s0885-3924(00)00199-8 (2000).
De Buck, M., Gouwy, M., Struyf, S., Opdenakker, G. & Van Damme, J. The ectoenzyme-side of matrix metalloproteinases (MMPs) makes inflammation by serum amyloid A (SAA) and chemokines go round. Immunol. Lett. 205, 1–8. https://doi.org/10.1016/j.imlet.2018.06.001 (2019).
McGowan, P. M. & Duffy, M. J. Matrix metalloproteinase expression and outcome in patients with breast cancer: Analysis of a published database. Ann. Oncol. 19, 1566–1572. https://doi.org/10.1093/annonc/mdn180 (2008).
González, L. O. et al. Overexpression of matrix metalloproteinases and their inhibitors in mononuclear inflammatory cells in breast cancer correlates with metastasis-relapse. Br. J. Cancer 97, 957–963. https://doi.org/10.1038/sj.bjc.6603963 (2007).
Malle, E., Sodin-Semrl, S. & Kovacevic, A. Serum amyloid A: An acute-phase protein involved in tumour pathogenesis. Cell Mol. Life Sci. 66, 9–26. https://doi.org/10.1007/s00018-008-8321-x (2009).
Hansen, M. T. et al. A link between inflammation and metastasis: Serum amyloid A1 and A3 induce metastasis, and are targets of metastasis-inducing S100A4. Oncogene 34, 424–435. https://doi.org/10.1038/onc.2013.568 (2015).
Lv, M. et al. Serum amyloid A stimulates vascular endothelial growth factor receptor 2 expression and angiogenesis. J. Physiol. Biochem. 72, 71–81. https://doi.org/10.1007/s13105-015-0462-4 (2016).
Soto, C., Saborio, G. P. & Anderes, L. Cyclic amplification of protein misfolding: Application to prion-related disorders and beyond. Trends Neurosci. 25, 390–394. https://doi.org/10.1016/s0166-2236(02)02195-1 (2002).
Zhang, J. et al. The threshold of alpha-fetoprotein (AFP) for the diagnosis of hepatocellular carcinoma: A systematic review and meta-analysis. PLoS ONE 15, e0228857. https://doi.org/10.1371/journal.pone.0228857 (2020).
Su, T. H. et al. Acoustic radiation force impulse US imaging: Liver stiffness in patients with chronic hepatitis B with and without antiviral therapy. Radiology 288, 293–299. https://doi.org/10.1148/radiol.2018171116 (2018).
Acknowledgements
We thank Ms. Mei-Hung Pan, Genomics Research Center, Academia Sinica, Taiwan to assist in multiple logistic regression analysis and Dr. Hwai-I Yang, Genomics Research Center, Academia Sinica, Taiwan to provide statistical suggestions. We thank Prof. Ding-Shinn Chen, National Taiwan University Hospital, to provide clinical suggestions.
Funding
The project and serum collection were supported by Bio-Signature Project from Academia Sinica (AS-BD-108-5); Genomics Research Center, Academia Sinica; Ministry of Science and Technology, Taiwan (MOST 109-2326-B-002 -012 -MY3); National Taiwan University Hospital (NTUH109-T20).
Author information
Authors and Affiliations
Contributions
J.L.W. performed all experiments and analyzed the data. T.H.S. collected all serum samples and clinical data. T.H.S. and P.J.C. provided critical research suggestions. J.L.W., T.H.S., P.J.C., and Y.R.C. wrote the manuscript. Y.R.C. conducted the research direction. All the authors interpreted the results and approved the final version of the article.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Wu, JL., Su, TH., Chen, PJ. et al. Acute-phase serum amyloid A for early detection of hepatocellular carcinoma in cirrhotic patients with low AFP level. Sci Rep 12, 5799 (2022). https://doi.org/10.1038/s41598-022-09713-9
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-022-09713-9
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
