
Abstract
Coleoid cephalopods camouflage on timescales of seconds to match their visual surroundings. To date, studies of cephalopod camouflage-to-substrate have been focused primarily on benthic cuttlefish and octopus, because they are readily found sitting on the substrate. In contrast to benthic cephalopods, oval squid (Sepioteuthis lessoniana species complex) are semi-pelagic animals that spend most of their time in the water column. In this study, we demonstrate that in captivity, S. lessoniana Sp.2 (Shiro-ika, white-squid) from the Okinawa archipelago, Japan, adapts the coloration of their skin using their chromatophores according to the background substrate. We show that if the animal moves between substrates of different reflectivity, the body patterning is changed to match. Chromatophore matching to substrate has not been reported in any loliginid cephalopod under laboratory conditions. Adaptation of the chromatophore system to the bottom substrate in the laboratory is a novel experimental finding that establishes oval squid as laboratory model animals for further research on camouflage.
Similar content being viewed by others
Introduction
Camouflage is an important defense mechanism for many animals, both terrestrial and aquatic1,2,3,4. While for most animals, body patterns are fixed, and are some partially variable5, coleoid cephalopods such as octopuses, cuttlefish, and squid, display rapid dynamic pattern and texture changes6,7,8. This alteration in color, texture and associated posture and movement, in response to their visual surroundings can occur in less than a second9,10,11,12. To date, the study of cephalopod camouflage has focused chiefly on benthic species of cuttlefish and octopus13. Research on cuttlefish, both in the lab and in the wild, has described a wide variety of pattern and texture displays, among which are background matching, disruptive patterning, masquerading, countershading, and mimicry9,14,15,16,17,18,19. In contrast, because of the technical challenges of working with and filming octopus, most camouflage and background matching research in octopus has been conducted in the wild20,21.
The rapidity of cephalopod camouflage, when compared to other camouflaging animals, is due to the unique neural control of the color and texture changes. Color and pattern changes are achieved by neurally-controlled chromatophores, iridophores, and leucophores which are differentially distributed throughout their bodies12,22. Certain regions of the squid’s body have higher densities of leucophores, which may be obscured or exposed by the chromatophores23. The combined chromatic expression of these chromatophores, iridophores, and leucophores allows cephalopods to display a wide repertoire of body patterns, often specifically matched to camouflage them in visually diverse environments such as coral reefs12,24,25.
By altering their color, texture, body shape, motion patterns, cephalopods are capable of several types of camouflage strategies. Two major types of camouflage behavior that have been frequently observed in cephalopods are: (1) “camouflage in motion,” where the camouflage changes dynamically as the animal moves through the environment26,27; and (2) “situational motionless camouflage,” where the animal actively selects a site and camouflages at that site without moving from it. In general, motionless camouflage is more common in nature5,28,29 because visually enabled organisms are skilled at movement detection, and rapid changes in color and shape are impossible for most of animals30,31,32.
Camouflage in cuttlefish and octopuses has often been described in the literature, but squid camouflage is less studied13. Scientific studies of camouflage behavior in squid species are limited, as most species of squid are primarily pelagic, making both direct observation and laboratory studies challenging. To date, only the Caribbean reef squid, Sepioteuthis sepioidea, a semi-pelagic species, has been reported to exhibit background matching by mottled pattern, countershading, and translucency; disruptive coloration against soft coral; masquerading to soft coral; and mimicry to coral, in the wild23,33 and the longfin inshore squid, Doryteuthis pealeii, using disruptive pattern34. In addition to basic countershading reflex35, mesopelagic deep-sea squid, Onychoteuthis banksii, was reported to control pigmentation and transparency based on ambient light conditions to yield optimal countershading36 while bioluminescent squids are controlling countershading using photophores37.
Oval squid, Sepioteuthis lessoniana Férussac in Lesson, 1830, form a not fully defined species complex38. Three members of the species complex are traditionally and routinely recognized by fishermen in Okinawa, Japan, which are morphologically, molecularly, and behaviorally distinct39,40,41,42,43,44,45 awaiting a taxonomical revision46. Here, we have focused on S. lessoniana sp.2, also called “Shiro-ika” or white-squid. We use the name white-squid within this article in order to distinguish it clearly from other members of the S. lessoniana species complex that might be behaviorally distinct. Unlike any other squid of the Loliginidae family, white-squid are bred in captivity over multiple generations47,48,49 and thus represent an attractive model species for studying the distinctive biology of loliginid squids.
In initial early observations of our laboratory housed white-squid, we recorded substrate color matching and camouflage in motion of adult white-squid, reared in captivity from wild collected eggs (Supplementary Video S1, Fig. 1; recorded in 2017). In the same year, a video of a squid from the S. lessoniana species complex was uploaded in the Seaunseen YouTube channel (https://www.youtube.com/watch?v=DzqhiRv-zOk; filmed by Renee Blundon in Kankadya Reef in Dar es Salaam, Tanzania). As the squid fed on passing fish it, at times, appeared to match the nearby substrate. These separate observations led us to design an experiment examining squid camouflage to substrate in motion in controlled laboratory conditions.

Squid substrate matching. The captive white-squid are swimming above a partially cleaned bottom of their tank, which is brighter than the uncleaned area, with dark and light body patterns over the matching dark and light substrate. This is a single frame from the Supplementary Video S1 of our initial observation in 2017.
Materials and methods
Animal maintenance
This project was conducted with the approval of the Okinawa Institute of Science and Technology Graduate University Animal Welfare Committee (OIST; AAAE 2016-137). Second-generation laboratory-bred adult white-squid (S. lessoniana Sp.2, Shiro-ika) were used in this study.
Egg strings of white-squid were collected in August 2017 from intertidal seagrass bed in Tancha village on the west coast of Okinawa Island, Ryukyu Archipelago, Japan. The eggs were transported to the Okinawa Institute of Science and Technology Graduate University Marine Science Station (OMSS), where the white-squid life cycle was closed in a flow-through system49, and the white-squid were reared through multiple generations. Outdoor tanks were illuminated solely by natural light and fed by ocean water pumped through a plumbing system with a sand filter. Because of this minimal processing, the tank water was representative of the seawater conditions in the surrounding ocean that are recorded by the Japan Meteorological Agency (https://www.jma.go.jp/).
White-squid were fed up to four times a day, with food type depending on their age. Subadult and adult white-squid were offered defrosted subadult and adult silver-stripe round herring Spratelloides gracilis and occasionally live tiger prawn Marsupenaeus japonicus, mysid, larval anchovy, or ghost shrimp to enhance the animals’ feeding and hunting motivation, rather than as a daily source of nutrition. Dead white-squid, waste, and any remaining food particles or live food organisms were removed from the tank after feeding session or upon discovery. During maintenance, we observed that the white-squid color regularly matched substrate, starting from the initial generation that hatched in captivity from wild collected eggs, as shown on the video from November 2017, containing 8 white-squids in the tank (Supplementary Video S1).
Data acquisition
We housed 3 sub-adult white-squids from the second laboratory-bred generation (two females and one male, hatched on August 5–8, 2018) starting from October 30, 2018 in a square-bottomed blue fiberglass tank with rounded corners and 156 cm lateral dimension (Aqua Culture system, Earth Corporation, Japan) sited outdoors. The depth of water was maintained at 40 cm. A SONY HD video camera (PXW-X70, SONY Corporation, Japan) was positioned above the water surface to record white-squid behavior and set at HD (1920 × 1080 at 30fps) resolution. We also placed GoPro (HERO6, GoPro inc. USA) cameras underwater for close-up HD videos (1980 × 1080 at 30fps) either perpendicular to the tank bottom on an outflow water pipe located at the center of the tank, or oriented parallel to the bottom surface at the bottom of the tank. The filming area was shaded by fabric to minimize light reflection from the water surface.
At the beginning of December, we removed algal growth from 50% of the tank bottom surface, yielding a distinctly reflective substrate on each 50% of the tank bottom: one with the light blue color of the fiberglass tank, and the other with the dark green color of algal growth (see Fig. 2). The experiment took place under overcast skies over on four following days at the end of December (Supplementary Table 1). Individuals are easy to recognize by size differences (Supplementary Tables 1, 2). For every trial of filming, a small amount of food (Engraulis japonicus) was fed to the white-squid from time-to-time to evaluate their health condition and confirm that it remained good throughout the experiment. During the experiment, white-squid were encouraged to cross the border between substrates by food or other attention-stimulating activities of the experimenter, initially on the first day also by a gentle submersion and a slow movement of PVC pipe (daily used for the tank cleaning) with an attached black ball on the opposite side of the tank than was the intended white-squid destination (Supplementary Video S2). This was no longer necessary by the second day of experiment when the white-squid reacted to mere presence and gesticulation of the experimenter. Over the 4 days, we were able to record 5 videos from above the tank and simultaneously side-view videos underwater (Supplementary Video S3, Supplementary Table 1).

Diagram of the experimental arena. Video camera (A) is placed above the tank. Outflow tube (B) appears at the center of the tank. Algal cover creates the dark substrate in the tank (C). The area where the bottom was cleaned, exposing the bright light blue color of the tank, which creates the light substrate (D).
Data analysis
We examined three parameters: (1) white-squid coloration changing according to substrate, (2) duration of white-squid color-changing event, and (3) effect of swimming orientation on the position of white-squid in the environment during color-changing event.
The recordings were assessed at times after the white-squid had sufficiently recovered from initial disturbances generated by camera installation and other activities and started regularly crossing the divider between substrates under SONY camera.
-
(1)
RGB scores of a ~ 1 cm2 (average 17.3 × 17.3, s.d. 2.11, pixel area) region of skin located between eyes and an equivalent region above the ink sac of each white-squid were measured by Digital Color Meter on iMac. The average RGB scores were calculated for each selected area independently (Fig. 3). These locations were selected because they are well defined, not transparent, and fully visible on the top view SONY camera recording. Each score was obtained within one second after a white-squid fully crossed the border between the two distinct substrates, as recorded by the cameras and displayed in SONY videos (Supplementary Table 1). The total lengths of the videos varied; to standardize we scored 10 min from each SONY video.
Figure 3 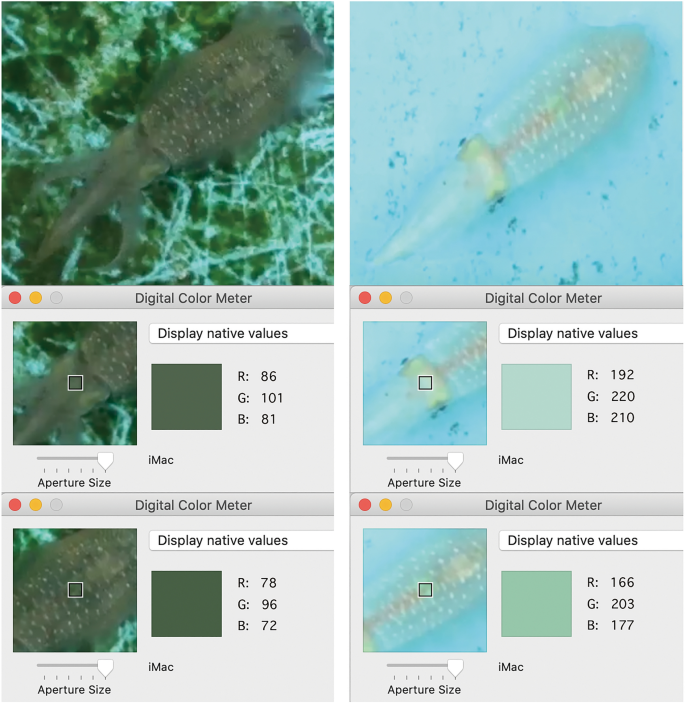
Example pictures of the subjects over dark or light substrates showing the RGB score as provided by the Digital Color Meter on iMac. When white-squid moved over different substrates, we measured the area between eyes of the animal, and the area right above the ink sac on the top view SONY camera recording (Supplementary Video S3), because these areas are never transparent and thus their color-change corresponds with activity of animal chromatophores.
We examined correlations between RGB scores and the two measured areas by Pearson correlation coefficient (Supplementary Table 1). To confirm our general observation of color-change correlation with substrate, we applied a linear mixed-effect model (lmer) in the lme4 v. 1.1-21 package50 of R version 3.6.251 in RStudio v. 1.2.503352 with restricted maximum likelihood estimation of variance components, and type III Wald F-tests with Kenward-Roger degrees of freedom appropriate for finite sample size53. We modelled the substrate as a fixed effect, day and individual as random effects. White-squid bodies are semi-transparent; top-view SONY videos do not reach single chromatophore resolution, but side-view GoPro videos allowed us to distinguish the chromatophores and observe directly their expansions and contractions during experiment (Supplementary Video S2).
The top-view SONY videos were further used to estimate the (2) duration of white-squid color-changing events, and the (3) effect of swimming orientation on the position of white-squid in the environment during color-changing event, because we noticed that the white-squid color does not always change at the same distance from the border between substrates. For purpose of these analyses, SONY recordings of 180 boundary crossing events were uploaded to Adobe Premier Pro 2020 for frame-by-frame analysis.
In order to assess squid color-change, RGB scores of the ~ 1 cm2 region of skin located between the eyes of each white-squid were recorded (Fig. 3). Single frames were exported as jpeg image files. The images were examined using image analysis software, Image J54. All video frames remain in their original HD video format (1080 × 1920 pixel), which allowed constant and accurate plotting. The central axis of the white-squid crossing the line connecting the center of the eyes was plotted for all targeted white-squid on the images in pixel. The plotted location in pixels was converted to millimeters (mm) with Image J54. The 60 mm GoPro camera width was used to calculate pixel per mm in each video-record.

-
(2)
For estimates of duration of squid color-changing events, we studied 10 events per animal per day of experiment (120 events in total). These events correspond with the 10 initial events per animal per day from the analysis of white-squid coloration changing according to the substrate (Supplementary Table 2). The beginning of the white-squid color-changing event was determined by the first video frame wherein color change is detected after a color remains stable for at least 15 video frames. The last frame was determined when the color change is completed, so that subsequently the white-squid color remains stable for at least 15 video frames. The number of frames from beginning to completion was counted and divided by 30 frames per second to determine each event's duration in seconds. We compared the duration of these color-changing events between days and animals by two-way ANOVA in R version 3.6.251 in RStudio v. 1.2.503352.
-
(3)
Because of differences between days in swimming orientation, the effect of swimming orientation (Arms-leading or Mantle-leading) on the position of the white-squid in the environment during color-changing event was examined by focusing on 30 initial events per animal in the fourth day of experiment which provides a series of well-balanced events across both types of swimming orientation and both types of substrates. We estimated the effect of swimming orientation on the distance of the color-measured point on the white-squid, once its color-change is complete, from the nearest point of the border between substrates (see Supplementary Table 2) by a linear mixed-effect model (lmer) in the lme4 v. 1.1-21 package50 of R version 3.6.251 in RStudio v. 1.2.503352 with restricted maximum likelihood estimation of variance components, and type III Wald F-tests with Kenward-Roger degrees of freedom appropriate for finite sample size53. We modelled the swimming orientation and type of substrate as fixed effects, individual as a random effect.
Ethics approval
All applicable international, national, and/or institutional guidelines for animal testing, animal care and use of animals were followed by the authors.
Results
White-squid coloration changing according to substrate
Our selected mathematical model revealed a highly significant interaction between the fixed effect of substrate (Fig. 3) and all the measured RGB color channel values (p < 0.0001) on the head that appear all highly positively correlated with each other and with the mantle (r > 0.94, p < 0.0001; Supplementary Table 1) and matching the destination substrate (Fig. 4).

Average head RGB values per day (1–4) per white-squid (A–C) and the substrate on dark and light substrate areas. Error-bars represent a 95% confidence interval for the mean.
Duration of white-squid color-changing event
White-squid changed their color during all examined boundary-crossing events, and all examined color-changing events are corresponding to these boundary-crossing events. The shortest measured duration of a white-squid color-changing event was approximately a third of a second, but two seconds was the average duration of white-squid color-changing event (Supplementary Table 2). We found significant differences in the duration of body pattern transformation between days covered by our experiments (d.f. = 3, F = 4.831, p = 0.0034), but we did not detect significant differences between individuals in our experiment over all days. The average velocity of white-squid during the color-changing event was approximately 0.16 m/s.
Effect of swimming orientation on the position of white-squid in the environment during color-changing event
We found that Mantle-leading swimming orientation resulted in the white-squid color-change occurring at a significantly (p < 0.0001) shorter distance from the substrate divider (mean = 13 ± 8 cm) when compared to Arm-leading swimming orientation (mean = 25 ± 8 cm), but there was no difference between traveling from light to dark or from dark to light substrate (Table 1, Supplementary Table 2).
Discussion
Although camouflage in squids has long been thought to be based on transparency, we demonstrated that they are also using chromatophores to adjust their color according to substrate. The animals in our experiment consistently (p < 0.0001) change their coloration while crossing between two distinct substrates. Although squid might have separate neural pathways to control their chromatic patterns on different body parts37,55,56, we observed a highly significant positive correlation between coloration on the head and mantle of camouflaging squid (r > 0.94, p < 0.0001; Supplementary Table 1), similarly to Josef et al. ’s26,27 observations in cuttlefish.
In a pelagic environment, the distinction between camouflage-to-substrate in motion behavior and countershading, which is widely associated with squids in the literature35,57, may be semantic. White-squid, however, reproduce in the optically heterogenous habitat of coral lagoons and seagrass meadows, returning to very shallow waters at times of maturity. In the latter habitat, countershading would be less effective than the camouflage-to-substrate capability of white-squid observed.
Squids face many different predators during their ontogeny and their defensive strategies change accordingly58. Our study focuses on mature or nearly mature animals. It has yet to be established when and how camouflage-to-substrate behavior emerges and/or changes during squid ontogenesis. The physiological factors that might be responsible for this behavior, and how environmental factors affect it have yet to be clearly defined.
The position of white-squid in the environment during color-changing events is significantly affected by squid swimming orientation (p < 0.0001; Table 1, Supplementary Table 2). The shortest white-squid color-changing event duration recorded by us in this study (0.3 s) corresponds with published single chromatophore expansion/contraction durations in loliginid squids59,60,61, but on average, the color-changing events in our study took four times longer. This variable and relatively long duration for the color-changing process which begins and is completed often centimeters from the divider (Supplementary Table 2) suggests that although squids anticipate the upcoming background, evidenced by becoming paler or darker before entering the light or dark substrate respectively, the final color is usually fully determined once the divider has been crossed, similarly to cuttlefish26,27. Alternatively, in our experiment, the duration of color-changing events might be affected by various external factors that have not been fully controlled, such as the mildness of stress induced by presence of experimenter, time since feeding, weather, natural lighting, water temperature, turbidity, chemistry, etc. Based on subjective visual examination of the video-recordings, we hypothesize that social interactions also influence squid camouflage behavior, including duration of body pattern transformation. Nonetheless, squids in our experimental setup are clearly changing color while in motion according to their proximity to the different substrates, regardless of coloration of their congeners farther away in the tank.
White-squid possess a rare combination of semitransparency and the ability to change body color via chromatophores (metachrosis) which enables them to successfully inhabit both pelagic and reef environments. Therefore, it represents an attractive model organism for studying the ecology, evolution, and neurobiology of versatile, dynamic camouflage.
Data availability
All data generated and analysed during this study are included in this published article [and its supplementary information files].
Code availability
No custom code was used in this study.
References
Endler, J. A. Interactions between predators and prey. In Behavioural Ecology: An Evolutionary Approach 3rd edn (eds Krebs, J. R. & Davies, N. B.) 169–196 (Blackwell, 1991).
Stevens, M. & Merilaita, S. Animal camouflage: Current issues and new perspectives. Philos. Trans. R Soc. Lond. B 364, 423–427 (2009).
Stevens, M. & Merilaita, S. Animal camouflage: Function and mechanisms. In Animal Camouflage: Mechanisms and Function (eds Stevens, M. & Merilaita, S.) 1–17 (Cambridge University Press, 2011).
Reiter, S. & Laurent, G. Visual perception and cuttlefish camouflage. Curr. Opin. Neurobiol. 260, 47–54 (2020).
Cott, H. B. Adaptive Coloration in Animals (Methuen, 1940).
Cloney, R. A. & Florey, E. Ultrastructure of cephalopod chromatophore organs. Z. Zellforsch. Mikrosk. Anat. 89, 250–280 (1968).
Borrelli, L., Gherardi, F. & Fiorito, G. A. Catalogue of Body Patterning in Cephalopoda (Firenze University Press, 2006).
Reiter, S. et al. Elucidating the control and development of skin patterning in cuttlefish. Nature 562, 361–366 (2018).
Barbosa, A., Allen, J. J., Mäthger, L. M. & Hanlon, R. T. Cuttlefish use visual cues to determine arm postures for camouflage. Proc. R Soc. B Biol. Sci. 279, 84–90 (2012).
Hanlon, R. T. Cephalopod dynamic camouflage. Curr. Biol. 17, R400-404 (2007).
Hill, A. V. & Solandt, D. Y. Myograms from the chromatophores of Sepia. J. Physiol. Lond. 83, 13–14 (1935).
Williams, T. L. et al. Dynamic pigmentary and structural coloration within cephalopod chromatophore organs. Nat. Commun. 10, 1–5 (2019).
Hanlon, R. T. et al. Rapid adaptive camouflage in cephalopods. In Animal Camouflage: Mechanisms and Functions (eds Stevens, M. & Merilaita, S.) 145–163 (Cambridge Univ Press, 2011).
Hanlon, R. T. & Messenger, J. B. Adaptive coloration in young cuttlefish (Sepia officinalis L.): The morphology and development of body patterns and their relation to behavior. Philos. Trans. R Soc. Lond. B 320, 437–487 (1988).
Ferguson, G., Messenger, J. B. & Budelmann, B. Gravity and light influence the countershading reflexes of the cuttlefish Sepia officinalis. J. Exp. Biol. 191, 247–256 (1994).
Shohet, A. J., Baddeley, R. J., Anderson, J. C., Kelman, E. J. & Osorio, D. Cuttlefish responses to visual orientation of substrates, water flow and a model of motion camouflage. J. Exp. Biol. 209, 4717–4723 (2006).
Barbosa, A. et al. Disruptive coloration in cuttlefish: A visual perception mechanism that regulates ontogenetic adjustment of skin patterning. J. Exp. Biol. 210, 1139–1147 (2007).
Chiao, C. C., Chubb, C. & Hanlon, R. T. Interactive effects of size, contrast, intensity and configuration of background objects in evoking disruptive camouflage in cuttlefish. Vis. Res. 47, 2223–2235 (2007).
Nakajima, R. & Ikeda, Y. A catalog of the chromatic, postural, and locomotor behaviors of the pharaoh cuttlefish (Sepia pharaonis) from Okinawa Island, Japan. Mar. Biodivers. 47, 735–753 (2017).
Packard, A. Chromatophore fields in the skin of the octopus. J. Physiol. 238, 38–40 (1974).
Caldwell, R. L., Ross, R., Rodaniche, A. F. & Huffard, C. L. Behavior and body patterns of the larger pacific striped octopus. PLoS ONE 10, e0134152 (2015).
Gutnick, T., Shomrat, T., Mather, J. A. & Kuba, M. J. The cephalopod brain: Motion control, learning, and cognition. In Physiology of Molluscs: A Collection of Selected Reviews Vol. 2 (eds Salleudin, S. & Mukai, S.) 139–177 (Apple Academic Press, 2016).
Hanlon, R. T. & Messenger, J. B. Cephalopod Behaviour 2nd edn. (Cambridge University Press, 2018).
Cloney, R. & Brocco, S. Chromatophore organs, reflector cells, iridocytes, and leucophores. Am. Zool. 23, 581–592 (1983).
Mäthger, L. M. & Hanlon, R. T. Malleable skin coloration in cephalopods: Selective reflectance, transmission and absorbance of light by chromatophores and iridophores. Cell Tissue Res. 329, 179 (2007).
Josef, N., Berenshtein, I., Fiorito, G., Sykes, A. V. & Shashar, N. Camouflage during movement in the European cuttlefish (Sepia officinalis). J. Exp. Biol. 218, 3391–3398 (2015).
Josef, N. et al. Size matters: Observed and modeled camouflage response of European Cuttlefish (Sepia officinalis) to different substrate patch sizes during movement. Front. Physiol. 7, 671 (2017).
Poulton, E. B. The Colours of Animals: Their Meaning and Use, Especially Considered in the Case of Insects (D. Appleton, 1890).
Zhang, Y. & Richardson, J. S. Unidirectional prey–predator facilitation: Apparent prey enhance predators’ foraging success on cryptic prey. Biol. Lett. 3, 348–351 (2007).
Troscianko, T., Benton, C. P., Lovell, P. G., Tolhurst, D. J. & Pizlo, Z. Camouflage and visual perception. Philos. Trans. R Soc. B 364, 449–461 (2009).
Land, M. F. & Nilsson, D. E. Animal Eyes (Oxford University Press, 2012).
Cronin, T. W., Johnsen, S., Marshall, N. J. & Warrant, E. J. Visual Ecology (Princeton University Press, 2014).
Hanlon, R. T. & Messenger, J. B. Cephalopod Behaviour (Cambridge University Press, 1996).
Staudinger, M. D., Hanlon, R. T. & Juanes, F. Primary and secondary defences of squid to cruising and ambush fish predators: Variable tactics and their survival value. Anim. Behav. 81, 585–594 (2011).
Ferguson, G. P. & Messenger, J. B. A countershading reflex in cephalopods. Proc. R. Soc. B 243, 63–67 (1991).
Zylinski, S. & Johnsen, S. Mesopelagic cephalopods switch between transparency and pigmentation to optimize camouflage in the deep. Curr. Biol. 21, 1937–1941 (2011).
Young, R. E. & Roper, C. F. E. Bioluminescent countershading in mid water animals: Evidence from living squid. Science 191, 1046–1048 (1976).
Jereb, P. & Roper, C. F. E. Cephalopods of the World. An Annotated and Illustrated Catalogue of Cephalopod Species Known to Date. Myopsid and Oegopsid Squids Vol. 2 (FAO, 2010).
Okutani, T. Life history of the oval squid, Sepioteuthis lessoniana. Saibai Giken 13, 69–75 (1984) ((in Japanese)).
Segawa, S. Food consumption, food conversion and growth rates of the oval squid Sepioteuthis lessoniana by laboratory experiments. Nippon Suisan Gakkai Shi 56, 217–222 (1990).
Izuka, T., Segawa, S., Okutani, T. & Numachi, K. Evidence on the existence of three species in the oval squid Sepioteuthis lessoniana complex in Ishigaki Island, Okinawa, southwestern Japan, by isozyme analyses. Venus Jpn. J. Malacol/Kairuigaku Zasshi 53, 217–228 (1994).
Izuka, T. Biochemical study of the population heterogeneity and distribution of the oval squid Sepioteuthis lessoniana complex in southwestern Japan. Am. Malac. Bull. 12, 129–135 (1996).
Imai, H., & Aoki, M. Genetic diversity and genetic heterogeneity of bigfin reef squid “Sepioteuthis lessoniana” species complex in northwestern Pacific Ocean. in Analysis of Genetic Variation in Animals (Caliskan, M. ed). 151–166. (InTech, 2012).
Cheng, S. H. et al. Molecular evidence for co-occurring cryptic lineages within the Sepioteuthis cf. lessoniana species complex in the Indian and Indo-West Pacific Oceans. Hydrobiologia 725, 165–188 (2014).
Tomano, S. et al. Contribution of Sepioteuthis sp. 1 and Sepioteuthis sp. 2 to oval squid fishery stocks in western Japan. Fish Sci 82, 585–596 (2016).
Okutani, T. Past, present and future studies on cephalopod diversity in tropical west Pacific. Phuket Mar. Biol. Center Res. Bull. 66, 39–50 (2005).
Lee, P. G., Turk, P. E., Yang, W. T. & Hanlon, R. T. Biological characteristics and biomedical applications of the squid Sepioteuthis lessoniana cultured through multiple generations. Biol. Bull. 186, 328–341 (1994).
Nabhitabhata, J. & Ikeda, Y. Sepioteuthis lessoniana. In Cephalopod Culture (eds Iglesias, J. et al.) 315–347 (Springer, 2014).
Lajbner, Z. et al. Captive breeding of the oval squid (Aori-ika; Sepioteuthis sp.). in Cephalopod International Advisory Council Conference 2018, Book of Abstracts, St. Petersburg. 152. (2018)
Bates, D., Mächler, M., Bolker, B. & Walker, S. Fitting linear mixed-effects models using lme4. J. Stat. Softw. 67, i01 (2015).
R Core Team. R: A Language and Environment for Statistical Computing. http://www.R-project.org (R Foundation for Statistical Computing, 2019).
RStudio Team. RStudio: Integrated Development for R. http://www.rstudio.com (RStudio, Inc., 2019)
Kenward, M. & Roger, J. Small sample inference for fixed effects from restricted maximum likelihood. Biometrics 53, 983–997 (1997).
Schneider, C. A., Rasband, W. S. & Eliceiri, K. W. NIH Image to ImageJ: 25 years of image analysis. Nat. Methods 9, 671–675 (2012).
Lin, C. Y., Tsai, Y. C. & Chiao, C. C. Quantitative analysis of dynamic body patterning reveals the grammar of visual signals during the reproductive behavior of the oval squid Sepioteuthis lessoniana. Front. Ecol. Evol. 5, 30 (2017).
Chung, W. S., Kurniawan, N. D. & Marshall, N. J. Toward an MRI-based mesoscale connectome of the squid brain. Iscience 23, 100816 (2020).
Messenger, J. B. Cephalopod chromatophores: Neurobiology and natural history. Biol. Rev. Camb. Philos. Soc. 76, 473–528 (2001).
York, C. A. & Bartol, I. K. Anti-predator behavior of squid throughout ontogeny. J. Exp. Mar. Biol. Ecol. 480, 26–35 (2016).
Suzuki, M., Kimura, T., Ogawa, H., Hotta, K. & Oka, K. Chromatophore activity during natural pattern expression by the squid Sepioteuthis lessoniana: Contributions of miniature oscillation. PLoS ONE 6, e18244 (2011).
Liu, Y.C., Wang, W.C., & Grasse, B. Electrical coupling between chromatophore muscle fibers allows for versatile control of chromatophore expansion in squid. bioRxiv 2020.02.17.951715 (2020).
Hadjisolomou, S. P., El-Haddad, R. W., Kloskowski, K., Chavarga, A. & Abramov, I. Quantifying the speed of chromatophore activity at the single-organ level in response to a visual startle stimulus in living, intact squid. Front. Physiol. 12, 675252. https://doi.org/10.3389/fphys.2021.675252 (2021).
Acknowledgements
We would like to thank OMSS staff for their dedicated support, namely Koichi Toda, Nobuo Ueda, and Kosuke Mori. We would like to thank Giovanni Masucci for helpful comments and Yuan Liu for help with graphics. This work was supported by the Physics and Biology Unit of the Okinawa Institute of Science and Technology Graduate University.
Funding
This work was supported by the Physics and Biology Unit of the Okinawa Institute of Science and Technology Graduate University.
Author information
Authors and Affiliations
Contributions
Z.L., R.N., M.K., J.M. contributed conception and design of the study; R.N. and Z.L. collected the data and organized the database; M.K., T.G., T.I., K.A., T.N. contributed to the data collection; Z.L. and R.N. performed statistical analyses; R.N. and Z.L. drew figures; R.N. and Z.L. wrote the first draft of the manuscript; M.K., T.I., J.M., T.G. wrote sections of the manuscript. All authors contributed to manuscript revision, read and approved the submitted version.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Supplementary Video S1.
Supplementary Video S2.
Supplementary Video S3.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Nakajima, R., Lajbner, Z., Kuba, M.J. et al. Squid adjust their body color according to substrate. Sci Rep 12, 5227 (2022). https://doi.org/10.1038/s41598-022-09209-6
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-022-09209-6
This article is cited by
-
Squid-inspired and wirelessly controllable display for active camouflage in aquatic-environment
npj Flexible Electronics (2024)
-
Peracetic acid treatment of squid eggs infected with parasitic copepod (Ikanecator primus gen. et sp. nov.)
Scientific Reports (2024)
-
Bioinspired designer surface nanostructures for structural color
Nanotechnology for Environmental Engineering (2024)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
