
Abstract
The Toca das Onças cave is one of the most important Quaternary mammal deposits of Brazil. Two different hypotheses have been proposed to explain the preservation mode of its skeletal remains: either the animals climbed down into the cave, or it could have functioned as a natural trap. Evaluation of pathological modifications on three articulated vertebrae of a single adult giant ground sloth Eremotherium laurillardi reveals a particular type of bone fracture caused by compressive force on the vertebral column, which split the vertebral bodies in the sagittal plane. This diagnosis suggests that the animal accidentally fell into the cave, in accordance with the second hypothesis proposed to the incorporation mode of skeletal remains into the cave.
Similar content being viewed by others

Introduction
The natural world is a tough place, where wild animals need to constantly adapt to different biotic and abiotic conditions to survive. In past animals, evidences of this “fight for survival” can be revealed through different types of analyses of bones preserved in the fossil record (e.g., ichnologic1; and isotopic2). An interesting category of evidence about the daily life of ancient animals are bone trauma – any break in the bone continuity caused by a force or mechanism extrinsic to the body3. This type of bone injury is able to reveal several paleoecological information such as intra and interspecific conflicts (e.g.,4,5), occupational activities (e.g.,6,7) and accidents8.
Despite its paleobiological significance, bone fractures in past animals remain underexplored when compared with other types of paleopathological conditions (e.g., arthritis9, and infections10), especially concerning the Pleistocene megafauna from South America. Recently, few cases of fractures have been described to this fauna, including a case in the ground sloth Nothrotherium maquinense11 and two independent cases in the giant ground sloth Eremotherium laurillardi12,13. Regarding the giant ground sloths, the record of complete and articulated Eremotherium skeletons in the Toca das Onças site has enabled inferences about the death and mode of incorporation of these remains into this cave deposit14,15.
This paper provides a new case of bone trauma in Eremotherium laurillardi, the only Pan-American megatheriine giant ground sloth and one of the taxa most often found in the Brazilian Quaternary fossil deposits16,17. In addition to detailing the fractures’ origin mechanism, we discuss how this particular case of bone injury can be used to shed light on the possible cause of death of the individual studied, on the incorporation of its bone remains into the taphocoenosis of the cave, and on the depositional aspects of caves.
Results
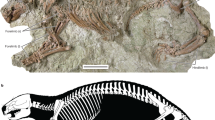
The three vertebrae investigated (DGEO UFPE 5769, DGEO UFPE 7167 and DGEO UFPE 9048) belong to the same individual and show lesions with similar appearance, location, and size. All lesions are in the caudal endplate of each vertebra near the left edges of the vertebral bodies running in the dorsoventral direction with a gently oblique position (Fig. 1). Each lesion appears as a deep and narrow bone discontinuity which has its gap widening at the midpoint before narrowing again, something that is most noticeable in the vertebra DGEO UFPE 9048 (Fig. 1a). Each lesion has approximately the following measurements of length: (1) 13.3 cm (12th thoracic vertebra, DGEO UFPE 9048); (2) 13.8 cm (13th thoracic vertebra, DGEO UFPE 7167); and (3) 8.4 cm (1st lumbar vertebra, DGEO UFPE 5769). In addition, it is possible to observe that all lesions have smooth and rounded edges (see images in detail in Fig. 1). There is no macroscopic evidence of new bone formation on the surface of the vertebrae.

Thoracic T12 and T13, and lumbar L1 vertebrae of Eremotherium laurillardi. (a–c) Photography of T12, T13 and L1 in caudal view, (d–f) schematic drawing of vertebrae in caudal view and close-up view of the traumatic lesions. Scale bars: 10 cm.
Discussion
Since the bone discontinuities noted in the three vertebrae analyzed show no clear sign of bone overgrowth, it is pivotal to rule out the possibility that we are dealing with preservation damages before proposing an accurate diagnosis for the lesions. The close-up view examination of the abnormalities shows that their edges have clear signs of smoothing and rounding (Fig. 1), which represent important evidence of osteoblastic activity18,19. Additionally, the similar color of the cortical damage and normal bone can be used as secondary evidence to rule out post-mortem processes as a possible origin of the alterations, since recent destructive processes are lighter than the rest of the bone19. Therefore, as taphonomic processes can be ruled out, the pointed evidence strongly suggests that the discontinuities observed are of pathological origin. More specifically, these breaks found in all three vertebrae are indicative of bone fracture.
Based on fracture analysis criteria applied here20, which consider the location and morphological pattern of the fractures, we classified the fractures noted in all vertebrae as traumas belonging to Type A (vertebral body compression), Group A2 (split fractures), and subgroup A2.1 (sagittal split fracture). This diagnosis implies that the traumatic episode was likely caused by a compressive force on the vertebral column, which split the vertebral bodies in the sagittal plane. This type of injury is considered stable—i.e., the fracture does not have a tendency to displace after reduction—and neurological deficit is uncommon20,22,23. Although stable traumas cause only moderate pain, without generating significant movement limitations20, the Eremotherium individual here analyzed died with unhealed bones, as there is no evidence of callus formation.
The absence of other skeletal signs that point to the presence of another type of disease concomitantly to the fractures allows us to reject the possibility that they have been generated as a result of a pre-existing disease (e.g., infection, neoplasm). We also consider that the vertebral injuries were not caused by repetitive force (stress fractures) because this type of injury is commonly characterized as a nondisplaced line or crack in the bone, called hairline fracture3. Those refer to situations where the broken bone fragments are not visibly out of alignment and exhibit very little relative displacement21. Although the Eremotherium vertebrae fractures’ can be described as nondisplaced, they also have a noticeable gap between their edges that is mostly narrow with wider parts in the middle, something found in split fractures20 but that is not characteristic of hairline fractures. Lastly, the subgroup C1.2.1 (rotational sagittal split fracture) might be a source of confusion due to similar morphological pattern with subgroup A2.1 (sagittal split fracture). However, in subgroup C1.2.1 there are compressive and rotational forces acting simultaneously, producing total separation into two parts20, which clearly did not occur in the vertebrae analyzed here.
In humans, compression fractures are most commonly caused by osteoporosis, although infection, neoplasm and trauma can also be etiological factors23,24,25. However, as aforementioned, the absence of other pathological skeletal marks is an important characteristic to take note as it serves to disregard the possibility of the fractures’ genesis to be secondary to another pathology. As such, in this case, osteoporosis, infection and neoplasm are unlikely etiologies. On the other hand, a compression fracture in a healthy individual is commonly generated after a severe traumatic event such as a fall from great height23,26. This scenario seems to better explain the origin of the vertebral fractures in the case of the Eremotherium ground sloth herein studied.
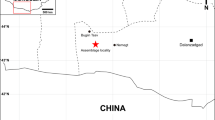
The three fractured vertebrae were recovered in the Toca das Onças site (Fig. 2), a small cave considered as one of the richest paleontological sites of the Brazilian Quaternary15. Two complete skeletons of Eremotherium laurillardi and fragments belonging to at least thirteen other individuals, together with several other bones assigned to different smaller species are known to this cave14. It comprises of a single dry chamber that can only be entered through vertical entrances approximately 4.5 m high (Figs. 2b–d and 3). Two different hypotheses concerning the depositional process of Toca da Onças were previously proposed: (1) the animals climbed down into the cave in search of water14; or (2) due to the vertical character of the cave entrance, it could have functioned as a natural trap where animals accidentally fell into the cave15.

Location map of the Toca das Onças site and images of the cave. (a) Detail of the location, (b) cave entrance area view, (c) view from inside the cave, (d) Cave entrance detail. Scale bars 10 m in (b) and 5 m in (c). This figure was generated by Adobe Photoshop CS6 software (https://www.adobe.com/br/products/photoshop.html).

Schematic representation of the Toca das Onças site. (a) Ground plan of the cave illustrating its morphology and dimension, (b) Cross-section illustrating the abyss-shaped entrance.
The first hypothesis would indicate that the animal fell into the cave during an attempt to climb down. However, there is no report in the literature indicating that Eremotherium laurillardi could have been a climbing animal. In addition, the vertical morphology of the cave entrance would be a limiting factor for climbing behavior (see Fig. 3).
Therefore, based on the type of fracture (compression sagittal split fracture) observed in the three vertebrae of Eremotherium as well as the inferred origin mechanism (fall from a great height), the presence of the individual here analyzed in the fossil accumulation of Toca das Onças is more likely explained by the second hypothesis. This idea is not particularly new as ‘entrapment due to fall’ has been described as a fossil accumulation mode to several other caves worldwide (e.g.,27,28). However, the use of bones fractures as an indicator of fossil accumulation mode is an interesting novelty. Of course, a detailed taphonomic investigation in the Toca das Onças still needs to be conducted in order to accurately interpret the formation of this important Quaternary fossil accumulation from Brazil.
In sum, we suggest that the animal accidentally fell into the cave, fractured at least three sequential vertebrae (12th, 13th thoracic vertebrae and 1st lumbar vertebra) after the impact on the ground, survived for a while, but succumbed trapped inside the cave without food and water (Fig. 4). Other animals found in the cave, but without signs of bone fracture, may have fallen and not fractured their bones or not survived after the fall, especially the smaller ones. Finally, the proposal of falls to explain the unusual record of giant ground sloth fossils preserving much of its skeleton in caves, as reported for Toca das Onças site, contrasts with the better-documented pattern of skeletal accumulation via hydraulic action.

Artistic reconstruction of the suggested fall of the individual Eremotherium laurillardi into the cave. Artwork by Júlia d’Oliveira.
Materials and methods
The material herein analyzed consists of three articulated vertebrae of an adult—indicated by the presence of vertebral plates completely fused—individual of Eremotherium laurillardi, as follows: (1) the twelfth thoracic vertebra (DGEO UFPE 9048); (2) the thirteenth thoracic vertebra (DGEO UFPE 7167); and (3) the first lumbar vertebra (DGEO UFPE 5769). The specimens were recovered from the Toca das Onças site (WGS84-UTM 300,604 m E; 8,791,416 m N), a small cave deposit located at the village of Caatinga do Moura, municipality of Jacobina, State of Bahia, Brazil (Fig. 2) and are housed in the Paleontological Collection of Geology Department of the Universidade Federal de Pernambuco (DGEO UFPE), Recife city, State of Pernambuco, Brazil.
The Toca das Onças site was developed in the Neoproterozoic dolomitic rocks of the Salitre Formation, Una Group. The cave has a single vertical entrance with a drop of approximately 4.5 m and a total area of 16 m229. It consists of a single chamber with a linear development of 23 m and a volume of 63 m329.
Each vertebra was macroscopically examined to characterize the pathological changes. We adopt here the fracture classification system proposed by Magerl et al.20 to describe the lesions studied. This classification system was created specifically for thoracic and lumbar vertebral fractures and is based on morphological patterns that can be divided in a set of three main categories of lesions (Type A, Type B and Type C). Each category reflects the effect of a force and the mechanism that was involved in the origin of the type of lesion and its subtypes. The system presents a hierarchy of types, groups and subgroups that are ranked according to its progressive severity and are accompanied by detailed morphological features (see20 for more details). The length of each fracture was measured using the open-source platform Fiji30.
Data availability
All data analyzed during this study are included in this work.
References
Araújo-Júnior, H. I., Barbosa, F. H. S. & Silva, L. H. M. Overlapping paleoichnology, paleoecology and taphonomy: Analysis of tooth traces in a Late Pleistocene-early Holocene megafaunal assemblage of Brazil and description of a new ichnotaxon in hard substrate. Palaeogeogr. Palaeoclimatol. Palaeoecol. 468, 122–128 (2017).
Tejada, J. V. et al. Isotope data from amino acids indicate Darwin’s ground sloth was not an herbivore. Sci. Rep. 11, 1–10 (2021).
Lovell, N. C. Trauma analysis in paleopathology. Yearb. Phys. Anthropol. 40, 139–170 (1997).
Farke, A. A., Wolff, E. D. S. & Tanke, D. H. Evidence of Combat in Triceratops. PLoS ONE 4, e4252. https://doi.org/10.1371/journal.pone.0004252 (2009).
Mackness, B. S., Cooper, J. E., Wilkinson, C. E. C. & Wilkinson, D. Palaeopathology of a crocodile femur from the Pliocene of eastern Australia. Alcheringa 34, 515–521 (2010).
Rothschild, B. M. & Laub, R. Pedal stress fractures in mastodons. J. Paleopathol. 20, 43–51 (2008).
Brown, C., Balisi, M., Shaw, C. A. & Van Valkenburgh, B. Skeletal trauma reflects hunting behaviour in extinct sabre-tooth cats and dire wolves. Nat. Ecol. Evol. 1, 0131. https://doi.org/10.1038/s41559-017-0131 (2017).
Rothschild, B. & Lambert, H. W. First documentation of a greenstick fracture in the fossil record. Possible gout also noted in Arkansaurus fridayii. Hist. Biol. 33, 1349–1351 (2019).
Tomassini, R. L. et al. Gregariousness in the giant sloth Lestodon (Xenarthra): multi-proxy approach of a bonebed from the Last Maximum Glacial of Argentine Pampas. Sci. Rep. 10, 1–16 (2020).
Barbosa, F. H. S., Porpino, K. O., Fragoso, A. B. L. & Santos, M. F. C. F. Osteomyelitis in Quaternary mammal from the Rio Grande do Norte State Brazil. Quat Int. 299, 90–93 (2013).
Barbosa, F. H. S., Porpino, K. O., Bergqvist, L. P. & Rothschild, B. M. Elucidating bone diseases in Brazilian Pleistocene sloths (Xenarthra, Pilosa, Folivora): First cases reported for the Nothrotheriidae and Megalonychidae families. Ameghiniana. 54, 331–340 (2017).
Dias, E. M. D., Dantas, M. A. T. & Barbosa, F. H. S. Diagnosis of bone diseases in two representatives of the Pleistocene megafauna of Bahia Brazil. Hist. Biol. 33, 3224–3227 (2020).
Andrade, L. C. et al. Revealing bone diseases in the Quaternary ground sloth Eremotherium laurillardi (Mammalia, Xenarthra). Hist Biol. 33, 1422–1430 (2021).
Cartelle, C. & Bohorquez, G. A. Eremotherium laurillardi Lund, 1842: Parte I. Determinação específica e dimorfismo sexual. Iheringia. 7, 45–63 (1982).
Auler, A. S. et al. U-series dating and taphonomy of Quaternary vertebrates from Brazilian caves. Palaeogeogr. Palaeoclimatol. Palaeoecol. 240, 508–522 (2006).
Cartelle, C. & De Iuliis, G. Eremotherium laurillardi: the Panamerican Late Pleistocene Megatheriid Sloth. J. Vert. Paleont. 15, 830–841 (1995).
Cartelle, C., De Iuliis, G. & Pujos, F. Eremotherium laurillardi (Lund, 1842) (Xenarthra, Megatheriinae) is the only valid megatheriine sloth species in the Pleistocene of intertropical Brazil: A response to Faure et al., 2014. C. R. Palevol. 14, 15–23 (2015).
Ortner, D. J. Identification of pathological conditions in human skeletal remains (Academic Press, 2003).
Waldron, T. Palaeopathology (Cambridge University Press, 2009).
Magerl, F., Aebi, M., Gertzbein, S. D., Harms, J. & Nazarian, S. A comprehensive classification of thoracic and lumbar injuries. Eur. Spine J. 3, 184–201 (1994).
Chowdhury, A. S., Bhandarkar, S. M., Robinson, R. W., Jack, C. Y. & Liu, T. Detection of hairline mandibular fracture using max-flow min-cut and Kolmogorov-Smirnov distance. In 2011 IEEE International Symposium on Biomedical Imaging: From Nano to Macro (eds IEEE) 1962–1965 (2011).
Simon, R. R. & Sherman, S. C. Emergency Orthopedics. (McGraw Hill, 2010).
Alexandru, D. & So, W. Evaluation and management of vertebral compression fractures. Perm J. 16, 46–51 (2012).
Nevitt, M. C. et al. The association of radiographically detected vertebral fractures with back pain and function: a prospective study. Ann. Intern. Med. 128, 793–800 (1998).
Gertzbein, S. D. et al. Thoracic and lumbar fractures associated with skiing and snowboarding injuries according to the AO comprehensive classification. Am. J. Sports Med. 40, 1750–1754 (2012).
Campagne, D. Fraturas de compressão da coluna. Manual MSD https://www.msdmanuals.com/pt-br/casa/lesões-e-envenenamentos/fraturas/fraturas-de-compressão-da-coluna (2021).
Andrews, P. Owls, caves and fossils: predation, preservation and accumulation of small mammal bones in caves, with an analysis of the Pleistocene cave faunas from Westbury-sub-Mendip (UK (University of Chicago Press, 1990).
Simms, M. J. Emplacement and preservation of vertebrates in caves and fissures. Zool. J. Linnean Soc. 112, 261–283 (1994).
Araújo, A. V., Nogueira, E. E. & Cajado, E. M. Caracterização do Geossítio Toca das Onças no município de Jacobina, Bahia, Brasil. Scientia Plena. 14, 125302. https://doi.org/10.14808/sci.plena.2018.125302 (2018).
Schindelin, J. et al. Fiji: A. open-source platform for biological-image analysis. Nat Methods. 9, 676–682 (2012).
Acknowledgements
We are grateful to Mr. Erickson Batista, Mr. Juliano de Jesus Silva and Sociedade Espeleológica Azimute who kindly give us the images and the map of the cave. This work was supported by the Fundação Carlos Chagas Filho de Amparo à Pesquisa do Estado do Rio de Janeiro under Grants [numbers E-26/201.881/2020; E-26/203.176/2017; and E-26/201.371/2021], Conselho Nacional de Desenvolvimento Científico e Tecnológico under Grant [number 301405/2018-2] and Universidade do Estado do Rio de Janeiro under Grant [number 38.944-5].
Funding
This article was funded by Fundação Carlos Chagas Filho de Amparo à Pesquisa do Estado do Rio de Janeiro (Grants no. E-26/201.881/2020; E-26/203.176/2017; and E-26/201.371/2021), Conselho Nacional de Desenvolvimento Científico e Tecnológico (Grant no. 301405/2018-2) and Universidade do Estado do Rio de Janeiro (Grant no. 38.944-5).
Author information
Authors and Affiliations
Contributions
F.H.S.B conceived the study, analyzed the data, prepared the figures and wrote the manuscript. I.C., H.I.A-J., A.V.A. and E.V.O. analyzed the data and wrote the manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
de S. Barbosa, F.H., de Araújo-Júnior, H.I., da Costa, I. et al. Spinal fracture reveals an accident episode in Eremotherium laurillardi shedding light on the formation of a fossil assemblage. Sci Rep 12, 4119 (2022). https://doi.org/10.1038/s41598-022-08107-1
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-022-08107-1
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
