
Abstract
Clues used by migrant birds to select sites for stopover are much less known than their reasons for leaving. Habitat characteristics and geographical location may affect the decision to use an island as a stopover site in different ways for different species. Thus, abundance and composition of migrants may be expected to differ between islands. Using standardized ringing from 9 western Mediterranean islands we evaluate drivers of abundance of trans-Saharan migrant passerines, specifically the role of species continental abundance, island characteristics and geographical location. Although continental abundance is a main driver of migrant composition on all islands migrant composition differs between them. Redundancy analysis and species response models revealed that the main drivers were distance to the nearest land toward the south, which has a positive effect on the number of migrants of most species, and island area, which appears as an important cue used for selecting a stopover island. Species whose abundance is positively related to island area have more pointed wings while species affected by distance to land toward the south have relatively more rounded wings. This suggests a hypothesis on the mechanism that may generate differences in passerine migrant composition between islands based on better efficiency of more pointed wings for long-distance flight.
Similar content being viewed by others

Introduction
Most long-distance migrants breeding in the western Palearctic need to cross two large barriers in each migration, the Sahara Desert and the Mediterranean Sea. The Sahara may be crossed in a continuous flight1 or doing stopovers at appropriate sites2. When crossing the sea, the only possible stopover sites, apart from urgent stops on boats, are islands. In both cases, the characteristics of habitats at stopover sites may strongly influence the probability of success of the migratory trip3. Species composition and abundance of bird migrants at stopover sites may depend on factors operating at several scales. At a wide spatial scale, continental abundance of each migrant species directly determines the global number of migrants and might be a potential driver of the relative abundance of each species at stopover sites. However, different specific migration strategies or decision rules for stopover4 could cause the relative abundance of migrants to deviate from mirroring their continental abundance.
To optimize migratory success, migrant birds must take at least two types of important decisions: whether or not to stop at a potential stopover site appearing over the course of a trip and, once landed, when to leave the site5,6. There is a huge amount of research about the factors that influence the decision to leave7. These studies have identified several intrinsic factors, such as fuel load8,9 or refueling rate6,10,11, and extrinsic factors such as weather including wind12, among other factors. On the contrary, the clues used by migrants to select particular sites for stopover are much less known5. Several studies have identified some important variables for mainland stopover sites, such as habitat characteristics, in particular forest cover8,13,14,15, and meteorological factors, such as rain16, but such factors are far less understood for island stopover sites (see 17 for an exception including islands close to the coast). Furthermore, wing morphology is related to the efficiency and range of flight18,19 so that flight performance related to wing shape could influence travel time or distance when crossing wide barriers20 and therefore is expected to affect species responses to island characteristics.
If habitat characteristics affect the decision to use a stopover site in different ways for different species, it may be expected that abundance and composition of migrants will differ between islands. The number and species composition of migrants that stop on islands in a given season would depend on a large diversity of variables. Some of these variables fluctuate greatly over time within a migratory season, such as daily intensity of the migratory passage, the weather along the migratory path and the physical conditions of migrants. For instance, migrants in poor condition may be more prone to landing on an island than healthier birds5,21 and unfavorable winds may promote the landing of more migrants22. The average distribution of these fluctuating variables depends on other island features that are more stable over time. Some of these features are related to the geographical location of the islands17, which determines its location relative to the main migration flyways, their distance to the mainland source of migrants, prevailing winds and, to a large extent, their climate. On the other hand, islands located in the same geographical region may differ in some specific characteristics that may also have an effect on migrant decisions to land. These include their size and topography, which are likely related to the diversity of habitats that a migrant may find, and to their vegetation cover. The interaction between the particular characteristics of an island, the specific habitat selection rules of the different migrant species and the physical condition of individuals should determine the probability of the decision to land (Fig. 1). However, the number of migrants actually landing would depend on the intensity of the migratory passage over the island on a given day. Therefore, stable and changing island features interact to determine the abundance and composition of the migrants that stop on a given island each day along a migratory passage, and consequently the number of migrants landing on an island is highly variable on a daily scale23.

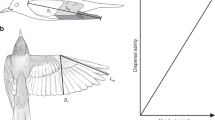
Conceptual model of the factors affecting the number of migrants of a species that make stopovers on an island. Continuous line circles and arrows identify factors that are constant over the long term. Dashed line circles and arrows identify factors that are highly variable, and change from day to day. Habitat characteristics are intermediate because some habitat components are stable (e.g. dominant vegetation types) while others may change through the migration season or between seasons (e.g. abundance of fruits or insects). Arrows ending over other arrows represent the modulating effect of some variables on the relationship of habitat characteristics and the landing decision. The interaction of the daily intensity of migratory passage and the probability of the decision to land of individual birds should determine the number of migrants actually landing each day. The sum of these numbers yields the number of migrants in a given season.
The stable features of islands would determine in part the abundance and composition of migrants and, in the long term, we expect that a general pattern would emerge for each island if migration seasons of several years are analyzed. However, the effect of species continental abundance might still be detectable and contribute to making migrant composition similar between islands. Therefore, the first aim of this paper is to assess if the abundance distribution of trans-Saharan migrant passerines that make stopovers at western Mediterranean islands is determined by the continental abundance of the species. The second objective is to evaluate beta diversity of these migrants, to estimate how it is partitioned among spatial and temporal components and determine if there are differences in the composition of migrants between this set of islands. We focus on trans-Saharan passerines in spring migration because ringing campaigns on these islands are timed to detect the passage of this group of species and do not sufficiently cover the phenology of short-distance migrants.
The third objective of this paper is to identify stable island characteristics that determine the gradients in migrant species composition by using community ordination techniques. These gradients would be shaped by species-specific responses to island characteristics that would determine the abundance of each species at stopover sites. Therefore, the fourth objective of this paper is to identify drivers of migrant species abundance at these islands. To attain these two objectives, we evaluate the relative importance of variables related to the geographical location of the islands and variables related to the habitats existing in them. Within the former, we include the distance to large land masses (continents or large islands) that could act as a source of migrants or could affect the decision to land on an island. In spring migration, larger distances to sources of migrants situated south of the islands are expected to increase the number of birds likely to make stopovers, since they would have been flying for a longer period of time1. On the other hand, on islands that are close to land masses located in other, non-southerly directions it is possible that some of the migrants prefer to continue flying a short distance to arrive at larger pieces of land that may offer more resources. In these islands migrant abundance is expected to decrease with distance to these land masses. We also analyze the effect of the location of the island in the west–east gradient since migrant species differ in their main routes of migration, with some crossing the Mediterranean in a wide front while others are more concentrated on western or eastern routes24. Concerning the specific characteristics of the islands, we analyze the size and topography of the island, which are likely related to the diversity of habitats existing there, and the Normalized Difference Vegetation Index (NDVI), which is an indicator of productivity and is expected to be related to food resource abundance for landed birds25. Finally, since species responses to island characteristics may be affected by wing morphology, we explored if species whose abundance is related to the different island variables also differ in wing pointedness and aspect ratio.
Materials and methods
Study islands and bird data
Systematic ringing in spring on Mediterranean islands has been promoted by the Piccole Isole project since 198826. Standard methods of the project involve ringing between 16th April and 15th May attempting to include the peak of the spring passage of long-distance migrants. Ringing is performed from dawn to nightfall using a constant number of nets within ringing stations placed at stable sites located at representative habitats in each island (Supplementary Table S1). The use of tape-lures is not allowed. We have compiled ringing data for all the Spanish Mediterranean islands that have been applying this methodology, with the exception of Mallorca and Menorca where the ringing stations were located in wetlands and captured a large percentage of local birds (Fig. 2, Table 1). The nine study islands are spread along a south-west to north-east gradient and, with the exception of Columbrets, they are distributed in pairs of similar longitude but different latitudes (Fig. 2). Ringing stations have been operating over a variable number of years (5–27 years), with the maximum number of ringing stations operating at the same time occurring between 2003 and 2010. To include between-year variation on islands that started ringing campaigns more recently we used data from the years 2003–2018.

Image source: Google Earth. Data SIO, NOAA, US Navy, NGA, GEBCO. Image Landsat/Copernicus.
Geographical location of studied islands in the western Mediterranean.
The ringing period within each spring also varied in most islands, owing to funding or logistic limitations; thus, to reduce the possible effects on migrant composition we only used data from the standard period of the Piccole Isole project and from years that included at least one week of ringing in the fortnight of each month within this interval. This procedure excluded the use of some years for several islands, and the final number of data years for islands ranged between 5 and 16 (Table 1).
We used only data for trans-Saharan nocturnal migrant passerines, which form the bulk of species ringed on Mediterranean islands during the standard period. The standard ringing period only covers the tail end of the short-distance migrants’ passage; thus, these species were excluded as their contribution to composition of migrants could vary mainly due to between-year variation in migration phenology. Diurnal migrants, like hirundinids and fringillids, also represent a small fraction of birds ringed and may use different cues to select stopover islands. In addition, some of these species nest in some of the islands studied and birds ringed could include breeding birds. To avoid the distorting effect of species that are captured accidentally in very small numbers, we considered only the species that were ringed in at least five separate years, or on five different islands, which limited the species considered to 35 (Supplementary Table S2). This led to the exclusion of just two species (Ficedula semitorquata with three individuals ringed in two islands and Locustella luscinioides with one individual ringed in Aire island). In addition, we only considered the number of ringed birds, since the proportion of recaptures varies among islands, likely reflecting variation in the duration of stopovers21, which could bias the comparison of the patterns of migrant species composition.
Island descriptors
We obtained two groups of variables describing the characteristics of the study islands (Tables 2, 3): (1) Variables related to geographical location: latitude, longitude, straight distance and minimum distance to the North African coast, minimum distance to the closest large body of land (continent or large island) in any direction and to the closest large body of land situated in a southerly angle between SW and SE. (2) Variables related to the habitat characteristics of the islands: area, maximum altitude and Normalized Difference Vegetation Index (NDVI). We estimated NDVI from Landsat 8 Images taken during the standard ringing period in the years 2015 and 2016. Pixels containing shoreline were excluded and the average NDVI was calculated for the rest of the pixels.
Continental abundance data
Abundance estimates for western Europe were obtained from the European Red List of Birds27. We used the mean of the minimum and maximum number of pairs estimated for the 27 EU Member States as a measure of continental abundance (Supplementary Table S2).
Data analysis
All analyses were done using R 3.6.128. We built a matrix of island-year x species containing the number of individuals of each selected species ringed in the study period in each island and year. Average number of individuals of each species ringed at each island was calculated and was used (log-transformed) as a dependent variable in a linear model with continental abundance (log-transformed), island and their interaction as predictors. This model was simplified using AICc as criteria to identify the best model.
To analyze variation of species composition, the matrix of island-year x species was transformed using the chord transformation29 with the function decostand in the vegan package30.
Using the function beta.div of the adespatial package31 we calculated beta diversity, including temporal and between-island variability (BDI,T), as the total variance of the aforementioned transformed matrix (BDTotal in29), which varies between 0 and 1 when chord distance is used. Considering that yijk is the chord transformed abundance of the species j in the island i and year k and \(\overline{{y }_{j}}\) is the mean for species j in all islands and years altogether, then:
where N is the total number of samples. The function beta.div also provides an estimation of contribution of localities (LCBD) and species (SCBD) to beta diversity (Table 3). Yearly LCBD (log transformed because of skewed distribution) of each island were averaged and compared between islands using ANOVA and a post-hoc Tukey test.
We partitioned the above sum of squares in several ways. First, we calculated a beta diversity that considered only between-island variability, excluding temporal variability (BDI), by averaging the chord transformed abundances of each species j in each island along study years (\({\overline{y} }_{ij}\)) and applying the same procedure, but using the number of studied islands (n):
Second, we calculated a beta diversity due to inter-annual variation of migrant composition within islands (BDT) as:
where Y is the total number of study years and n is the number of studied islands (9). We also calculated a temporal beta diversity for each island i (BDTi) as the sum of squares due to variation within the island divided by the number of the island study years (Yi) minus 1:
Differences in temporal variability between islands could be due to different predominance of species that are more or less variable between years. To check this, we calculated Spearman’s rank correlation between the percentage of captures of each species in the total ringed on each island and BDTi and LCDB indices, for species present on all islands.
We tested for the existence of differences between islands in migrant species composition using Permutational Multivariate Analysis of Variance (PERMANOVA) using the function adonis2 in the vegan package. We performed a multivariate test of homogeneity of variances using the betadisper function (vegan package) with the adjustment for small sample bias, to test if temporal variability in species composition differed between islands. We made post-hoc comparisons between islands with False Discovery Rate (FDR) correction using the function pairwise.perm.manova of the package RVAideMemoire32.
To identify gradients in migrant species composition and the island characteristics that were associated with them, we employed Redundancy Analysis using the rda function (vegan package). We used the chord transformed matrix of species x island-year as a response matrix. We used two explanatory matrices, one including variables of geographical location and the other the variables related to habitat characteristics of the islands. We evaluated the relative importance of each group of variables to explain migrant species composition by performing a variation partitioning analysis, using the varpart function (vegan package). For that analysis, we followed the steps and R scripts recommended in33.
Variables describing island characteristics were transformed using natural logarithms and collinearity within each group was evaluated with variance inflation factor (VIF)34. All the habitat variables presented VIF < 3 and were retained in subsequent analyses, but some of the geographical location variables presented VIF values much larger than 10, so we removed the variable with the highest VIF and recalculated VIF values. This procedure was repeated until all variables presented VIF < 10 and led to the exclusion of minimum distance to Africa and latitude. Maximum VIF in the remaining variables was 2.1. We then performed a separate forward selection of variables within each group using the vegan function ordistep and all variables were selected.
A second redundancy analysis was performed with all the explanatory variables together to identify the particular variables most related to gradients of composition of migrants. Because we have repeated measures for each island, we fitted linear mixed models to identify the explanatory variables significantly related to each RDA axis, using the function glmmTMB (glmmTMB R package35). In these models, scores of each RDA axis were dependent variables, each island descriptor was tested as an explanatory variable and the island identity was included as a random effect. To evaluate the species whose abundance was significantly related to RDA gradients, we calculated correlation coefficients of each species abundance (chord transformed) and RDA axis scores.
Models of species response
To evaluate the response of each study species to the geographical and habitat characteristics of the islands, we calculated the average number of individuals captured daily per 100 m of mist net in each study year in each island. However, the capture index from Columbrets Island is not comparable to the other islands because most birds were trapped in just three nets placed around a clump of isolated Opuntia cactus, which strongly attracted birds landing on this island. On the other islands, nets were set in several straight lines across the available habitat, and thus sampling was more uniformly distributed. The relationship between the percentage of total captures and the number of birds captured per 100 m of net is similar in all islands except Columbrets (Supplementary Fig. S1 online) and thus it was not possible to include Columbrets in the models for the captures/100 m net.
To check if excluding Columbrets from these analyses changed the results, the species response to island variables was modeled twice: using as a dependent variable the proportion of captures, including Columbrets, and using an abundance index calculated as the average number of birds captured daily per 100 m of mist net (excluding Columbrets). In both cases, we used generalized linear mixed models (GLMM, R function glmmTMB). In the proportion of captures models, we included the natural logarithm of the total number of birds ringed of the selected species in each island and year as an offset. For the captures/100 m net models, the offset was calculated as the natural logarithm of the result of multiplying the total length of nets (in hundreds of meters) by the number of days that the nets were operating in each spring. Island identity and year, considered as a factor, were included in the models as random effects. Each of the island variables included in the RDA, after excluding colinear variables, was included in turn as a fixed effect in these GLMMs. We considered three potential family distributions: Poisson and two parameterizations of the Negative Binomial distribution (nbinom1: variance increases linearly with the mean; nbinom2: variance increases quadratically with the mean). A null model including only the random effects was run with each family distribution and the one with the lowest AICc was selected. Then, linear and quadratic models including by turn one of the island descriptors were fitted and retained if it had an AICc lower than the null model. For each species we considered as equally plausible the models with ΔAICc ≤ 2. We used the performance_aicc function from the performance package36 to calculate AICc. Significance of regression coefficients in quadratic models was checked by comparing quadratic and linear models with the anova function.
Effect of wing shape on island selection
We used the Kipp index as an indicator of wing pointedness37. This index is calculated as the percentage of the distance from the tip of the first secondary feather to the tip of the longest primary feather (primary projection) of the wing length (Kipp = 100*primary projection/wing length)38. Primary projection was measured with a transparent rule placed over the folded wing and wing length was measured as the maximum length with a butted ruler38, both with a precision of 0.5 mm. Primary projection data are only available for Tabarca Island from 2018 onwards, so these measures were lacking for some of the species less frequently ringed in the study area. The primary projection of great reed warblers (Acrocephalus arundinaceus) and wood warblers (Phylloscopus sibilatrix) were obtained from a nearby ringing station in the mainland (Hondo Natural Park). The Kipp index of individuals were averaged to obtain mean and SD for each species. We also used the wing aspect ratio (WAR) available in39 for most of the species studied.
We tested if mean Kipp index and wing aspect ratio differed between groups of species whose abundance was related to different RDA axes or selected island variables using phylogenetic generalized least-squares (PGLS) models40 constructed using the R package ape41 and nlme42. These models allow to account for potential non-independence among species due to shared evolutionary history by estimating a phylogenetic signal index, Pagel’s λ43, that measures phylogenetic dependence of observed trait data. For that aim, following44 2500 trees were downloaded from Birdtree web site45 for the set species with Kipp index available and the same quantity of trees for the species with WAR estimates available. A consensus phylogenetic tree was built for each set of species using the R package phytools46 (Supplementary Figs. S4 and S5 online). Iduna opaca was not available in Birdtree data and was substituted by the closely related Hippolais pallida (currently named Iduna pallida), from which it was split. Bivariate PGLS models including interaction were fitted and interaction was removed if not significant. Models were fitted with estimated Pagel’s λ and with λ fixed to zero (i.e. no phylogenetic signal). Likelihood ratio test was used to compare these models and if not significant the simplest model was selected47.
Results
The total number of birds ringed of each selected species in the study years is shown in Supplementary Table S2. Half of the species (18 out of 35) were ringed on all islands and represent 98.6% of all ringed birds analyzed. The species most often ringed on all the islands was the willow warbler (Phylloscopus trochilus), representing between 30 and 61% of all birds ringed.
The model for the relationship of continental abundance and the number of birds ringed on each island that included an additive effect of island was better than the model with the interaction (ΔAICc = 12.04), but the model without an effect of island was even better (ΔAICc = 6.05). The model with only the island predictor and the null model were clearly worse (ΔAICc > 128 in both cases), thus this result supports the hypothesis that continental abundance has a positive significant effect on species abundance at stopover sites and that this effect is similar in all islands studied (Supplementary Fig. S4 online). To check if this could be an effect of the large continental abundance of the willow warbler, which is almost an order of magnitude greater than any other species and could be an influential point, models were fitted excluding this species but the result was the same. Therefore, a model relating logarithm of continental abundance of each species to logarithm of number ringed on all islands was finally fitted (Log(ringed) = 0.8875 × Log(Continental abundance) −3.1019; F1,33 = 26.03; p < 0.001; r = 0.664; Fig. 3a).

(a) Relationship between continental abundance (horizontal axis) and total number ringed (both in logarithmic scale). (b) Relationship between the total number ringed (horizontal axis, logarithmic scale) and their SCBD (Species Contribution to Beta Diversity). Hollow circles: species not captured on all islands. Filled circles represent species captured on all islands differentiating between species correlated (red circles) with RDA 1 (see Fig. 4) or not correlated with that axis (blue circles). Regression line in b fitted for species ringed on all islands only. Acronym identification in Supplementary Table S2. For clarity, only species acronyms for red circles are shown in a.
Beta diversity
Total beta diversity was low (BDI,T = 0.098). Beta diversity excluding temporal variability (BDI = 0.056) was very similar to the average beta diversity within islands caused by temporal variability (BDT = 0.056). The average contribution of islands to beta diversity (LCBD, Table 3) was different among islands (F8,65 = 3.49, p < 0.01), due mainly to significant differences between Cabrera and Columbrets and Aire (Tukey-post-hoc tests p < 0.01) and marginally significant differences between Tabarca and the same two islands (Tukey-post-hoc tests p < 0.1). BDTi and LCDB were highly correlated (r = 0.912, p < 0.001) showing that islands that were more variable interannually contributed more to beta diversity. The only island variable related to BDTi and LCDB was the maximum altitude of the island (BDTi: r = 0.787; LCDB: r = 0.729, both p < 0.05). However, this result is due to the high variability of Dragonera, the highest island, and if Dragonera is dropped from the analysis, this correlation disappears. Neither BDTi nor mean LCBD were correlated with the number of years of study on each island (BDTi: r = −0.361; LCDB; r = −0.339, n.s.).
Species Contribution to Beta Diversity (SCBD) was positively correlated to their ringed total (r = 0.613 p < 0.001). The main outlier of this trend was the willow warbler, by far the most ringed species, which had a SCBD lower than expected for its abundance (Table 4, Fig. 3b). Species ringed on all islands accumulated almost all SCBD (0.9966) showing that beta diversity was due to changes in abundance of these migrants between islands. The only species with a significant correlation with indices of temporal variability between islands was the willow warbler (LCDB; rs = −0.700, p = 0.043; BDTi: rs = −0.817, p = 0.008; df = 7). After excluding the most variable island (Dragonera), the outcome is similar (LCDB; rs = -0.690, p = 0.058; BDTi: rs = −0.857, p = 0.007; df = 6). Therefore, the beta diversity of islands where willow warbler represents a larger proportion of migrants tends to be less variable between years.
The PERMANOVA test yielded significant differences in migrant composition between islands (F8,65 = 7.89, p < 0.001) and the permutational post-hoc test detected significant differences between all islands except between Dragonera and Colom and only marginally significant differences between Aire and Columbrets. However, as the betadisper test for multivariate homogeneity of variance was significant (F8,65 = 2.53, p < 0.05, Tukey post-hoc test: only differences between Dragonera and Grosa and Aire p < 0.05), this result could be influenced by the heterogeneity of variances. This analysis was repeated excluding Dragonera and then the betadisper test was no longer significant (F7,61 = 1.59, p > 0.1) while PERMANOVA produced a very similar result (F7,61 = 8.88, p < 0.001). Post-hoc comparisons yielded the same result of migrant species composition being different between all islands with the exception of Aire and Columbrets, whose difference was marginally significant (p < 0.1).
Gradients of migrant species composition
The partitioning variance analysis performed with all variables indicated that the island descriptors explained close to 40% of variance (R2adj = 0.391) that was partitioned in R2adj = 0.140 explained by geographical variables (ANOVA permutational test for rda: F4,66 = 5.02, p < 0.001), R2adj = 0.127 explained by habitat variables (ANOVA permutational test for rda: F3,66 = 5.82, p < 0.001) and R2adj = 0.124 (not testable) explained jointly by the two data sets. Geographical and habitat characteristics have weak correlations except for NDV and Longitude; therefore, the position of the study islands along the west–east direction appears to be the main link between habitat and geographical variables.
The RDA performed with all selected island variables together identified as significant the first three axes (p < 0.01 in all cases), which accounted for 86.5% of the variance explained by the model and 38.9% of total variance (Table 4). The first RDA axis (RDA1) was related to the island habitat variables and Longitude (Table 5). Axis 2 (RDA2) and 3 (RDA3) were related to distance to the closest large body of land situated in a southerly direction (MinDSouthLand), but this relationship was negative in the former and positive in the latter (Table 5). RDA2 was also negatively related to the distance to the large body of land in any direction (MinDistLand). None of the axes were related to the distance to the African coast.
Yearly variation in migrant composition in each island tends to be distributed in a particular space in the resulting ordination (Fig. 4). Points of smaller islands are mostly located towards the right side of the first axis and those of the larger islands (Cabrera, Dragonera and Formentera) towards the left. Ordination in axis 2 and 3 separates islands that are very close to large bodies of land located south (Conillera, Dragonera and Colom) from islands that are far from the nearest coast in a southerly direction. The only species related positively to RDA1 was the willow warbler, while the rest of the species significantly related to this axis show a negative correlation (Table 4), with whinchat (Saxicola rubetra), garden warbler (Sylvia borin) and pied flycatcher (Ficedula hypoleuca) being the most important. The most abundant migrant species on those islands were correlated with RDA1 (Fig. 3b), thus its abundance seems related to habitat characteristics of islands. The most significant correlations of species with RDA2 were negative, supporting the idea that abundance of these species is greater on islands that are far from a southern coast.

Triplot of the RDA relating migrant species composition to island variables. Each point corresponds to a spring migration year on one island. Pairs of islands located at similar longitude are depicted with the same color. Within each pair, points of the island with a more southern position are represented with a filled circle and hollow polygons and points of the island in the northern position with a hollow circle and shaded polygons. Dashed dark green arrows represent island variables significantly related to at least one of the axes represented (Table 5). Continuous black arrows represent the species most correlated to the axis (Table 4). The small unlabeled arrow in panel a corresponds to PHYSIB and in b corresponds to LUSMEG. Species acronyms identification in Supplementary Table S2. Figures were produced using R (R 3.6.1, https://www.R-project.org).
Response of species to island characteristics
The variable most often included in significant models (Supplementary Tables S3 and S4; Fig. S5 online) is the distance to the main area of land located south (Table 6), both for explaining the percentage of captures and the standardized number of birds ringed (abundance index). The position of the island in the east–west gradient is also important in both groups of models. Among the island characteristics, island area and NDVI are the variables most often included in both groups of models. Variables had a more consistent effect when used to model abundance index, thus in 20 of the studied species abundances increased with distance to southern land, in 10 species it increased with island area and in 9 with longitude. Distance to the northern coast of Africa was the least important variable.
There was a strong correlation (r = 0.885, p < 0.001) between the Kipp index for the study species ringed in Tabarca and wing aspect ratio compiled by39 from wild birds in Central Europe (Supplementary Table S5), which supports the idea that the Kipp index is a good surrogate for wing aspect ratio. PGLS models comparing mean Kipp index and wing aspect ratio of groups of species that were related to the main RDA axis or to the most important variables in species response models yielded very similar results. Interaction was not significant in any case and was removed from all the models. According to likelihood ratio test Pagel’s λ was retained in the model relating Kipp index to RDA axes (λ = 0.802) and the model relating WAR to Area and distance to the main area of land located south (λ = −0.645). Species whose abundance is negatively correlated with RDA1 had on average more pointed wings (Fig. 5) than species not correlated with this RDA axis (Kipp index: t22 = 2.78, p < 0.05; WAR: t21 = 2.22, p < 0.05), while there is no effect of correlation with RDA2 (Kipp index: t22 = 1.20, n.s.; WAR: t21 = 1.38, n.s.). When species were classified as related or not related to island area and to the distance to the main area of land located south there was a significant effect of island area (Kipp index: t22 = 3.34, p < 0.01; WAR: t21 = 3.70, p < 0.01) and MinDSouthLand (Kipp index: t22 = 2.17, p < 0.05: WAR: t21 = 4.11, p < 0.01). Species whose abundance responded to island area had more pointed wings while species related to distance to the land located south had more rounded wings (Fig. 5).

Mean (± SE) Kipp index (a) and wing span ratio (c) for species classified as presenting a significant negative correlation with RDA1 or RDA2 or not (No effect) (Table 4). Mean (± SE) Kipp index (b) and wing span ratio (d) for species classified as having a significant positive effect of the variables area or MinDSouthLand on its abundance index or not (No effect) (Table 3S). Minimum values in Y axis of both variables correspond approximately to the values for two short distance migrant species (Erithacus rubecula and Phylloscopus collybita).
Discussion
Most of the migrant species were present on all islands and summed to more than 98% of total birds ringed during the study years. The abundance index of passerine species at stopover sites was correlated with their continental abundance on all islands. Therefore, beta diversity is low and well below its possible maximum value of one. Despite this, when relative abundance of migrants is considered, virtually every island has its own pattern of migrant passerine composition, even islands that are located near each other. On the contrary, the most similar islands in terms of migrant composition (Aire and Columbrets) are situated almost at the extremes of the longitudinal gradient studied. Therefore, distance between islands at the scale of this study is not a determinant for their differences in migrant composition and other causes should be involved.
About half of the beta diversity is explained by inter-annual variation within islands. The difference between islands in temporal variability is not an artifact due to their different lengths of monitoring periods. Cabrera and Dragonera were the most variable islands, even though they were studied for 8 and 5 years respectively, while Aire is one of the least variable islands despite having been monitored for the longest period of the study set (16 years). Differences in temporal variability of migrant composition between islands could be explained by geographical or habitat characteristics, but we did not detect consistent effects of any of these variables. An alternative is that differences in temporal variability originate from differences in species composition. Given that continental abundance determines to a large extent migrant abundance, if populations of some species are more variable interannually48,49,50 and these species are more abundant on some islands, those islands would present more variation in migrant composition between years. In addition, some species could be more sensitive than others to variation in meteorological conditions51,52 which could cause more interannual variability in migrant composition. However, these potential effects are not detected because we did not find positive correlations between any species and island temporal variability indices, but only a consistent negative correlation with willow warbler relative abundance. Therefore, islands where willow warbler represents a larger proportion of migrants tend to be less variable between years. Given that among the species analyzed in this study the willow warbler is, by far, the most abundant in Europe and at the stopover islands, its high abundance might buffer between-year variability.
The variance partition analysis showed that geographical location and habitat characteristic variables explained one third each of the variation in migrant composition on the study islands, while the remaining third was explained conjointly by both sets of variables. Thus, the two types of variables are not completely independent, since some habitat characteristics change consistently with geographic position. This is due to the correlation between longitude and NDVI, which in this set of islands increases eastwards according to an increasing rainfall gradient from southwest to northeast.
The main gradient in species composition is strongly related to all the habitat variables and to a lesser extent to the longitudinal position of the island. The other two important RDA axes are only related to geographical variables, the distance to large pieces of land in any direction (MinDistLand) and in a southerly direction (MinDSouthLand). Willow warbler is the only species correlated to the gradient of habitat characteristics (RDA1) whose abundance appears to increase on small and dry islands. This is particularly striking since the remaining 16 species that present significant correlations with this axis are positively associated with island area. From Supplementary Table S2, it can be calculated that these 16 species together represent a similar number of birds (on average 43% of birds ringed on the study islands; range 24–64%) to the number of willow warblers ringed. Given that the chord transformed abundance is, like the percentage, a relative abundance index, when the abundance of the group of species positively related to larger islands increases, the relative abundance of willow warblers should decrease and in fact their chord transformed abundances are negatively correlated (r = −0.897).
To adequately interpret these results, it is necessary to analyze the models for species response to island variables. Response models for the species correlated with the habitat characteristics gradient are in general consistently positively related to area, NDVI or maximum altitude, regardless of whether the percentage of captures or the number of captures per 100 m of net is used as the response variable. On the contrary, in the case of the willow warbler the model for the percentage of captures is negatively related to area but the number of captures is independent of this variable. Therefore, the apparent preference of willow warbler for smaller islands is an artifactual consequence of about one-third of the species studied stopping in larger numbers on larger islands, thus decreasing the relative abundance of willow warblers on larger islands. Those species appear to be more selective of island habitat characteristics while willow warblers would stop in large numbers on any island available. It has been suggested that birds in poorer condition would be less selective of island characteristics and land at the first available site21. According to this idea, individuals of species whose abundance is related to island characteristics are expected to be on average in better condition. But contrary to expected, the average fat score of the species related to island characteristics is lower both in Columbrets (mean 1.2, SD 0.57, range: 0.6–2.5) and the Dry Balearic Islands (a group that includes all the Balearic islands analyzed here; mean: 2.0, SD: 0.37, range: 1.3–2.5) than the fat score of willow warbler (Columbrets mean: 1.8; Dry Balearic Islands mean: 3.0; data in21). The lower fat load of these species suggests that they would be on average in greater need of refueling and their relation to island habitat variables supports that they are more selective of stopover sites, in accordance with5. Refueling rates are often negative on dry small islands21 and thus it would pay, at least for individuals that are not in a desperate situation, to continue migration to a better stopover site. Given that small islands may provide other benefits apart from refueling53,54, the larger on average fat load of the willow warbler could explain their independence of island characteristics and weak relationship to island location.
The habitat variable that presents the most frequent and consistent positive effect in species response models for standardized numbers of captures was island area. Species models with NDVI and maximum altitude often included quadratic terms and predicted maximum or minimum numbers of captures at intermediate values of these variables, making their effect more unclear than with island area. This result supports the idea that island area may be an important clue used by migrants to assess island quality as a stopover site, as it may be the easiest characteristic to evaluate from afar. Alternatively, larger islands could be simply easier to detect from greater distances and would attract a larger number of birds. However, this explanation seems unlikely in our study area because all the smaller islands, apart from Columbrets, are very close to larger islands or the mainland coast, which could attract migrants.
Species whose abundance is correlated to the other two RDA axes appear to be related to island geographical location. Species response models for the number of captures reveal that the distance to the closest large body of land situated in a broad southerly direction is significant and has a positive effect for most species (20 of the 35 species analyzed). Therefore, in most species, the farther the island from a large piece of land located broadly south, the larger the abundance of landed migrants, which is likely related to a greater proportion of individuals in need of resting. Conversely, this suggests that islands that are closely located northwards to a larger body of land (Conillera, Dragonera and Colom) tend to be overflown by a larger proportion of migrants that made stopovers on the land that appeared just before. This is easily seen in the graphics for species response models since the above-mentioned islands have a counterpart located south of the same large Balearic island (Formentera, Cabrera and Aire) where the abundance of these migrants is considerably greater, despite being practically at the same longitude and in some cases very close by (see Aire and Colom). This result agrees with the prediction of optimal migration theory for the time minimizing strategy, which predicts that fuel gain at the best sites would allow skipping less optimal sites5.
On the other hand, some of the islands are very close to the Iberian Peninsula or a large Balearic island located to the north or northwest (Grosa, Tabarca, Formentera and Aire). Thus, birds could easily overfly these islands to arrive at the larger land. This does not seem to occur because in general birds arrive in large numbers to these islands even though trophic resources would be more abundant in the mainland. This also occurs in the Tyrrhenian Islands and has been explained by the idea that recovery after a long flight would first require rest54. Stopover at those islands tends to be in any case very short21,54.
The pattern that emerges from our results is that variation in migrant composition is determined mainly by two variables, island area and island distance to large pieces of land located in a broad southerly direction. Island area affects about a third of studied species that are more abundant at larger islands. Furthermore, abundance of most migrant species increases the farther an island is from other land that they could have encountered previously. Different species appear to be responding in different ways to these two variables and the analyses of primary projections and wing aspect ratios reported here support that wing morphology contributes to defining species' responses to island characteristics and geographical location. This supports the proposal of a hypothesis on the mechanism that may generate the differences in passerine migrant composition between islands.
Wing pointedness is related to the efficiency of long-distance flight and pointed wingtips have been shown to be one of the factors that reduce flight costs for small birds during migration55,56. Moreover, mass-adjusted wing aspect ratio has been found to be positively related to migration distance57. This suggests that individuals of species with more pointed wings crossing the Mediterranean would be more able to make longer flights, which would give them a better capacity to skip low quality islands and select better stopover islands, if they appear, or fly until the continent. Landing at more extensive islands may be advantageous as these islands have a more developed vegetation and a larger diversity of habitats, that would allow migrants to select the best habitat and reach a higher fuel deposition rate. On the contrary, small Mediterranean islands offer a very limited variety of dry scrublands, where fuel deposition rates have been found to be lower than in large Balearic Islands21. Migrating passerines arriving at unfamiliar stopover areas must base their choices on instant decisions58 and island area may be a quick and easy indicator to use. Low-quality stopover sites can act as an ecological trap, especially for migrating birds with insufficient fuel load to leave in search for an alternative place of a higher quality58. Moving to an alternative stopover site may be even more costly and unpredictable when crossing the sea, thus the use of cues for improving the chance of selecting a high-quality island or avoiding low quality ones would be highly rewarding.
On the other hand, it is revealing that species whose abundance is positively related to distance to land toward the south have on average a less pointed wing than species that do not present that relationship. Therefore, the higher flight costs expected for these species would determine that a relatively larger proportion of individuals need to land on the first available island, a proportion that would increase with the distance that birds had flown previously. The capacity of these species to select stopping islands according to their characteristics would be reduced, in comparison to species with more pointed wings, and their choice of stopover island may be more opportunistic. According to this hypothesis, larger islands would be enriched in individuals of migrant species with more pointed wings while on islands farther from southern land migrants with more rounded wings would reach higher abundance. Nevertheless, despite the importance of wing shape, it would be worth investigating which other morphological, physiological or ecological characteristics determine variation in stopover strategies and how they interact with weather variation. The importance of these drivers of migrant composition may be spatially dependent and thus could be different on sets of islands with different spatial distribution, such as the Tyrrhenian Islands, spreading along a north–south gradient along the Italian coast24. Understanding how drivers of migrant composition change on other sets of islands would contribute to a more general view of factors determining landing decisions.
Data availability
The datasets generated during and/or analyzed during the current study are available from the corresponding author upon reasonable request.
References
Blackburn, E. et al. Spring migration strategies of Whinchat Saxicola rubetra when successfully crossing potential barriers of the Sahara and the Mediterranean Sea. Ibis 161, 131–146 (2019).
Salewski, V., Schmaljohann, H. & Liechti, F. Spring passerine migrants stopping over in the Sahara are not fall-outs. J. Ornithol. 151, 371–378 (2010).
Moore, F. R. Biology of landbird migrants: a stopover perspective. Wilson J. Ornithol. 130, 1–12 (2018).
Cohen, E. B. et al. A place to land: Spatiotemporal drivers of stopover habitat use by migrating birds. Ecol. Lett. 24, 38–49 (2021).
Chernetsov, N. Passerine Migration: Stopovers and Flight (Springer, 2012).
Alerstam, T. Optimal bird migration revisited. J. Ornithol. 152(Suppl 1), S5–S23 (2011).
Müller, F. et al. Towards a conceptual framework for explaining variation in nocturnal departure time of songbird migrants. Mov. Ecol. 4, 24. https://doi.org/10.1186/s40462-016-0089-2 (2016).
Ktitorov, P., Tsvey, A. & Mukhin, A. The good and the bad stopover: behaviours of migrant reed warblers at two contrasting sites. Behav. Ecol. Sociobiol 64, 1135–1143 (2010).
Klinner, T., Buddemeier, J., Bairlein, F. & Schmaljohann, H. Decision-making in migratory birds at stopover: an interplay of energy stores and feeding conditions. Behav. Ecol. Sociobiol. 74, 10. https://doi.org/10.1007/s00265-019-2784-7 (2020).
Delingat, J., Bairlein, F. & Hedenström, A. Obligatory barrier crossing and adaptive fuel management in migratory birds: the case of the Atlantic crossing in Northern Wheatears (Oenanthe oenanthe). Behav. Ecol. Sociobiol. 62, 1069–1078 (2008).
Schmaljohann, H. & Eikenaar, C. How do energy stores and changes in these affect departure decisions by migratory birds? A critical view on stopover ecology studies and some future perspectives. J. Comp. Physiol. A 203, 411–429 (2017).
Saino, N. et al. Spring migration decisions in relation to weather are predicted by wing morphology among trans-Mediterranean migratory birds. Funct. Ecol. 24, 658–669 (2010).
Bonter, D. N., Gauthreaux, S. A. & Donovan, T. M. Characteristics of important stopover locations for migrating birds: remote sensing with radar in the Great Lakes basin. Conserv. Biol. 23, 440–448 (2009).
Buler, J. J., Moore, F. R. & Woltmann, S. A multi-scale examination of stopover habitat use by birds. Ecology 88, 1789–1802 (2007).
Buler, J. J. & Moore, F. R. Migrant–habitat relationships during stopover along an ecological barrier: extrinsic constraints and conservation implications. J. Ornithol. 152, 101–112 (2011).
Andueza, M., Barba, E. & Arizaga, J. Using capture-mark-recapture models to assess the effect of age and weather on landing decisions of sedge warblers Acrocephalus schoenobaenus during migration. Ardeola 61, 269–283 (2014).
McCabe, J. D. & Olsen, B. J. Landscape-scale habitat availability, and not local geography, predicts migratory landbird stopover across the Gulf of Maine. J. Avian Biol. 46, 395–405 (2015).
Bowlin, M. S. Sex, wingtip shape, and wing-loading predict arrival date at a stopover site in the Swainson’s Thrush (Catharus ustulatus). Auk 124, 1388–1396 (2007).
Marchetti, K., Price, T. & Richman, A. Correlates of wing morphology with foraging behaviour and migration distance in the genus Phylloscopus. J. Avian Biol. 26, 177–181 (1995).
Buler, J. J., Lyon, R. J., Smolinsky, J. A., Zenzal, T. J. Jr. & Moore, F. R. Body mass and wing shape explain variability in broad-scale bird species distributions of migratory passerines along an ecological barrier during stopover. Oecologia 185, 205–212 (2017).
Gargallo, G. et al. (eds) Spring bird migration in the Western Mediterranean: results from the Piccole Isole Project. (Monografies del Museu de Ciències Naturals 6, (2011).
Clipp, H. L. et al. Broad-scale weather patterns encountered during flight influence landbird stopover distributions. Remote Sens. 12, 565. https://doi.org/10.3390/rs12030565 (2020).
Knudsen, E. et al. Characterizing bird migration phenology using data from standardized monitoring at bird observatories. Clim. Res. 35, 59–77 (2007).
Pilastro, A., Macchio, S., Massi, A., Montemaggiori, A. & Spina, F. Spring migratory routes of eight trans-Saharan passerines through the central and western Mediterranean; results from a network of insular and coastal ringing sites. Ibis 140, 591–598 (1998).
Briedis, M. et al. Broad-scale patterns of the Afro-Palaearctic landbird migration. Glob. Ecol. Biogeogr. 29, 722–735 (2020).
Spina, F., Massi, A., Montemaggiori, A. & Bacceti, N. Spring migration across central Mediterranean: general results from the “Proggeto Piccole Isole”. Vogelwarte 37, 1–94 (1993).
BirdLife International. European Red List of Birds. http://datazone.birdlife.org/info/euroredlist. (2015).
R Core Team. R: A language and environment for statistical computing. (R Foundation for Statistical Computing, Vienna, Austria. https://www.R-project.org (2019).
Legendre, P. & De Cáceres, M. Beta diversity as the variance of community data: dissimilarity coefficients and partitioning. Ecol. Lett. 16, 951–963 (2013).
Oksanen, J. et al. vegan: Community Ecology Package. R package version 2.5-5. (2019).
Dray, S. et al. adespatial: Multivariate Multiscale Spatial Analysis. R package version 0.3-7. (2019).
Hervé, M. RVAideMemoire: Testing and Plotting Procedures for Biostatistics. R package version 0.9-73. (2019).
Borcard, D., Gillet, F. & Legendre, P. Numerical Ecology with R 2nd edn. (Springer, 2018).
Zuur, A. F., Ieno, E. N. & Elphick, C. S. A protocol for data exploration to avoid common statistical problems. Methods Ecol. Evol. 1, 3–14 (2010).
Brooks, M. E. et al. glmmTMB balances speed and flexibility among packages for zero-inflated generalized linear mixed modeling. R J. 9, 378–400 (2017).
Lüdecke, D., Makowski D., & Waggoner Ph. performance: assessment of regression models performance. R package version 0.2.0. (2019).
Lockwood, R., Swaddle, J. P. & Rayner, J. M. V. Avian wingtip shape reconsidered: Wingtip shape indices and morphological adaptations to migration. J. Avian Biol. 29, 273–292 (1998).
Salewski, V., Siebenrock, K.-H., Hochachka, W. M., Woog, F. & Fiedler, W. Morphological change to birds over 120 years is not explained by thermal adaptation to climate change. PLoS ONE 9, e101927 (2014).
Vincze, O., Vágási, C. I., Pap, P. L., Palmer, C., & Møller, A. P. Wing morphology, flight type and migration distance predict accumulated fuel load in birds. J. Exp. Biol. 222, jeb183517 (2019).
Symonds, M .R. E. & Blomberg, S. P. A Primer on Phylogenetic Generalised Least Squares in Modern Phylogenetic Comparative Methods and Their Application in Evolutionary Biology (ed. Garamszegi, L.Z.) 105–130 (Springer, New York, 2014).
Paradis, E., Claude, J., & Strimmer, K. ape 5.0: an environment for modern phylogenetics and evolutionary analyses in R. Bioinformatics 35, 526–528 (2019).
Pinheiro, J., Bates, D., DebRoy, S., Sarker, D. & R Development Core Team. nlme: linear and nonlinear mixed effects models. R package version 3.1–153 (2021).
Freckleton, R. P., Harvey, P. H. & Pagel, M. Phylogenetic analysis and comparative data: a test and review of evidence. Am. Nat. 160, 712–726 (2002).
Rubolini, D., Liker, A., Garamszegi, L. Z., Møller, A. P. & Saino, N. Using the BirdTree. org website to obtain robust phylogenies for avian comparative studies: a primer. Curr. Zool. 61, 959–965 (2015).
Jetz, W., Thomas, G. H., Joy, J. B., Hartmann, K. & Mooers, A. O. The global diversity of birds in space and time. Nature 491, 444–448 (2012).
Revell, L. J. phytools: An R package for phylogenetic comparative biology (and other things). Methods Ecol. Evol. 3, 217–223 (2012).
Münkemüller, T. et al. How to measure and test phylogenetic signal. Methods Ecol. Evol. 3, 743–756 (2012).
Gregory, R. D., Skorpilova, J., Vorisek, P. & Butler, S. An analysis of trends, uncertainty and species selection shows contrasting trends of widespread forest and farmland birds in Europe. Ecol. Indic. 103, 676–687 (2019).
Brlík, V. et al. Long-term and large-scale multispecies dataset tracking population changes of common European breeding birds. Sci. Data 8, 21 (2021).
Tirozzi, P., Orioli, V., Dondina, O., Kataoka, L. & Bani, L. Species traits drive long-term population trends of common breeding birds in northern Italy. Animals 11, 3426 (2021).
Nilsson, C., Bäckman, J. & Alerstam, T. Seasonal modulation of flight speed among nocturnal passerine migrants: differences between short- and long-distance migrants. Behav. Ecol. Sociobiol. 68, 1799–1807 (2014).
Packmor, F., Klinner, T., Woodworth, B.K., Eikenaar, C., & Schmaljohann, H. Stopover departure decisions in songbirds: do long-distance migrants depart earlier and more independently of weather conditions than medium-distance migrants? Mov. Ecol. 8, 6; doi:https://doi.org/10.1186/s40462-020-0193-1 (2020)
Linscott, J. A. & Senner, N. R. Beyond refueling: Investigating the diversity of functions of migratory stopover events. Ornithol. Appl. 123, 1–14 (2021).
Maggini, I., Trez, M., Cardinale, M. & Fusani, L. Stopover dynamics of 12 passerine migrant species in a small Mediterranean island during spring migration. J. Ornithol. 161, 793–802 (2020).
Bowlin, M. S. & Wikelski, M. Pointed wings, low wingloading and calm air reduce migratory flight costs in songbirds. PLoS ONE 3, e2154 (2008).
Weber, J. M. The physiology of long-distance migration: extending the limits of endurance metabolism. J. Exp. Biol. 212, 593–597 (2009).
Vágási, C. I. et al. Morphological adaptations to migration in birds. Evol. Biol. 43, 48–59 (2016).
Domer, A., Vinepinsky, E., Bouskila, A., Shochat, E. & Ovadia, O. Optimal stopover model: A state-dependent habitat selection model for staging passerines. J. Anim. Ecol. 90, 2793–2805 (2021).
Acknowledgements
The authors want to thank especially all the ringers and helpers that have participated in the ringing campaigns over those years. A quick calculation yields that a minimum of 33,750 field working hours were needed to obtain the data analyzed here. This work was done in its vast majority by volunteers and it would have been impossible to pay for this work. We also want to thank the institutions that provided funding for materials and maintenance costs of volunteers.
Author information
Authors and Affiliations
Contributions
G.M.L-I. conceived the study, analysed the data and wrote the paper. G.M.L-I., A.B., J.C., R.E., A.S. and M.S. organized and coordinated the ringing campaigns in the islands that contributed with data to the analyses. All the authors revised critically drafts of the paper and approved submission.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
López-Iborra, G.M., Bañuls, A., Castany, J. et al. Drivers of migrant passerine composition at stopover islands in the western Mediterranean. Sci Rep 12, 2943 (2022). https://doi.org/10.1038/s41598-022-06912-2
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-022-06912-2
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
