
Abstract
The bulk of body fat in mammals is in the form of triacylglycerol. Diacylglycerol O-acyltransferase 2 (DGAT2) catalyses the terminal step in triacylglycerol synthesis. The proximity of DGAT2 with stearoyl-CoA desaturase (SCD) in the endoplasmic reticulum may facilitate provision of de novo SCD-mediated fatty acids as substrate for DGAT2. Here, we first searched for sequence variants in the DGAT2 gene to then validate their effect on fat content and fatty acid composition in muscle, subcutaneous fat and liver of 1129 Duroc pigs. A single nucleotide polymorphism in exon 9 (ss7315407085 G > A) was selected as a tag variant for the 33 sequence variants identified in the DGAT2 region. The DGAT2-G allele increased DGAT2 expression in muscle and had a positive impact on muscular C14 and C16 fatty acids at the expense of C18 fatty acids. Although there was no evidence for an interaction of DGAT2 with functional SCD genotypes, pigs carrying the DGAT2-G allele had proportionally more palmitoleic acid relative to palmitic acid. Our findings indicate that DGAT2 preferentially uptakes shorter rather than longer-chain fatty acids as substrate, especially if they are monounsaturated, and confirm that fatty acid metabolism in pigs is subjected to subtle tissue-specific genetic regulatory mechanisms.
Similar content being viewed by others

Introduction
Meat fat content and composition are important attributes that contribute to the nutritional quality and the consumer perception of meat products. Total fat dietary intake, particularly intake of saturated fatty acid (SFA), has been associated with obesity, circulating LDL-cholesterol and increased risk of coronary heart disease1. In line with this, public health recommendations advocate for low-fat diets and dietary substitution of SFA with monounsaturated fatty acids (MUFA) and polyunsaturated fatty acids (PUFA)2,3,4,5. On the other hand, although most consumers already prefer leaner meat products6, intramuscular fat (IMF) content and fatty acid composition exert a great influence on meat organoleptic and technological properties7,8. Increasing the knowledge of the genetic basis of the regulation of fat metabolism is key to improve the quality of meat products.
Triacylglycerols (TG) are the main stores of metabolic energy, serving as a reservoir of essential and non-essential fatty acids, and the precursors of phospholipids. In mammals, TG are primarily synthesized in the liver, intestine and adipose tissue9. Diacylglycerol O-acyltransferase (DGAT1 and DGAT2) genes are involved in intestinal fat absorption and play a critical role in the synthesis of TG in the adipocytes10,11. In particular, DGAT1 and DGAT2 encode membrane proteins that catalyse the formation of the ester bond between the hydroxyl group of 1,2-diacylglycerol and a long-chain fatty acyl-CoA (Fig. 1)9,12,13, the final step in the production of TG in mammals12,13,14,15. Although DGAT1 and DGAT2 have similar functions in the adipocytes, DGAT2 has been found to be more critical in mice, where it is essential for post-natal survival and can as well compensate for disruptions of DGAT116,17,18. Recent findings indicate that DGAT2 is specifically engaged in de novo lipogenesis by favouring both the synthesis of TG from glycerol 3-phosphate and the incorporation of endogenous fatty acids into TG, particularly the MUFA that result from the desaturation of palmitate (C16:0) and stearate (C18:0) through the action of stearoyl-CoA desaturase (SCD)19. DGAT2 and SCD can localize in the endoplasmic reticulum membrane and this proximity may facilitate the provision of de novo SCD-mediated MUFA as substrate for DGAT2 to form TG.

The role of DGAT2 in the synthesis pathway of triacylglycerol. Triacylglycerol is (1) synthesized de novo by sequential addition of fatty acyl moieties to a glycerol 3-phosphate (the G3P pathway) or (2) formed through the re-esterification of partial glycerides that results from either the hidrolysis of pre-existing triacylglycerol or via the monoacylglycerol (MAG) pathway. De novo transformation of diacylglycerol to triacylglycerol is catalyzed by DGAT2 (1) but re-esterification of partial glycerides is done by either DGAT2 (2a) or DGAT1 (2b). However, while DGAT1 specifically incorporates pre-formed fatty acids, DGAT2, which is able to form protein complexes with MGAT2, FATP1 and SCD, can use both endogenous and pre-formed fatty acids as substrates. DGAT1 diacylglycerol O-acyltransferase 1, DGAT2 diacylglycerol O-acyltransferase 2, ELOVL fatty acid elongase, FATP1 fatty acid transport protein 1, MGAT2 monoacylglycerol acyltransferase 2, SCD stearoyl-CoA desaturase.
Polymorphisms within DGAT2 have the potential of affecting lipogenesis. Thus, DGAT2 has been investigated as a candidate gene for fat-related traits such as obesity in humans11,20 and for lean content and muscle and backfat fatty acid composition in livestock species, particularly in pigs14,21,22. As a result of a genome-wide association study (GWAS) for fatty acid composition in Duroc pigs, we found that the region on SSC9 where DGAT2 lies was associated with MUFA content. Here, using this Duroc line as a model, we investigate the role of DGAT2 variants on fat metabolism in pigs. We first searched for sequence variants in the promoter, coding, and 3′-untranslated regions of porcine DGAT2 and then validated their effect on fat content and composition using data and samples from a large biorepository23. Given that this line segregates for a functional variant in the SCD gene24, potential interactions of DGAT2 with SCD were also assessed.
Materials and methods
Ethics statement
Pigs used in the study were raised and slaughtered in commercial units following applicable regulations and good practice guidelines on the protection of animals kept for farming purposes, during transport and slaughter (Royal Decree 37/2014, Spain) and tissue samples used were collected from these pigs at the slaughterhouse. In agreement with the European directive 2010/63/UE, tissue sampling from non-experimental animals slaughtered following legal procedures is not under the scope of European regulation on animal experimentation and does not require specific project authorization. The experimental protocol was approved by the Ethical Committee on Animal Experimentation of the University of Lleida. The study was carried out in compliance with the ARRIVE guidelines.
Animals and phenotypes
A total of 1129 pigs from 158 sires and 559 dams of the same Duroc line were used in the experiment. Pigs were raised in 15 fattening batches between 2006 and 2016 following a similar protocol for data recording and tissue sampling25. In each batch, pigs were raised from 75 days of age until slaughter in the same farm under identical conditions. During this period, pigs had ad libitum access to commercial feed (Esporc, Riudarenes, Girona, Spain). At 206 (SD 8) days of age, all pigs were weighted and backfat (BT) and loin (LT) thickness were ultrasonically measured at 5 cm off the midline between the third and fourth last ribs using the portable equipment Piglog 105 (Frontmatec, Kolding, Denmark). Pigs were slaughtered at 211 (SD 9) days in a commercial slaughterhouse equipped with a carbon dioxide stunning system. Carcass composition traits were recorded, including carcass weight, BT and LT. Both BT and LT were measured at 6 cm off the midline between the third and fourth last ribs using an ultrasound automatic scanner (AutoFOM, SFK-Technology, Denmark). Immediately after slaughter, a sample of semimembranosus muscle (SM, n = 40) and subcutaneous fat (SF, n = 226) were collected, snap-frozen and stored at -80ºC. After chilling for about 24 h at 2 °C, a large sample of the muscles gluteus medius (GM, n = 1093) and longissimus thoracis (LM, n = 526) were collected, vacuum packaged, and stored at − 20 °C until required. The IMF content in GM and LM, as well as the fatty acid composition in GM, LM and SF were determined in duplicate by quantitative gas chromatography26. The amount of each fatty acid was expressed as the percentage of each individual fatty acid relative to total fatty acid. The proportion of SFA (C14:0, C16:0, C18:0 and C20:0), MUFA (C16:1n-7, C18:1n-9, C18:1n-7 and C20:1n-9) and PUFA (C18:2n-6, C18:3n-3, C20:2n-6 and C20:4n-6) were calculated.
Genome-wide association study
In order to identify candidate genomic regions associated with fat-related traits, GWAS for each individual fatty acid were performed using a subset of 254 pigs. The genomic DNA of these pigs was isolated as described in Ref.24 and used for single nucleotide polymorphism (SNP) genotyping with either the PorcineSNP60 v2 Genotyping BeadChip (n = 138) or the GGP Porcine HD Array (n = 116) porcine arrays (Illumina, San Diego, CA, USA). Those SNPs that displayed a minor allele frequency below 0.10, a call rate below 0.95, or that could not be mapped to Sus scrofa reference genome (Sscrofa11.1)27 were filtered out. A total of 36,000 SNPs remained after data quality control. For each trait, a GWAS was performed by fitting a linear mixed model using GEMMA28, where phenotypes were adjusted for batch (11 levels) and IMF content as a covariate. The association of each SNP was tested using the Wald statistic considering the Bonferroni correction for multiple testing. Significance was set to a level of P ≤ 1.4 × 10–6. Two regions were found to be associated with C16:1n-7, one on SSC9 and another on SSC14, the latter corresponding to a reported polymorphism in the SCD gene that is known to affect MUFA and particularly C16:1n-724. Candidate genes mapping within the SSC9 region were explored with Ensembl Genes Database using BioMart (https://www.ensembl.org/biomart/martview). Functional analyses like Gene Ontology and Reactome Pathway Enrichment Analysis were performed using Enrichr29. DGAT2 was retrieved as the most promising candidate gene for C16:1n-7 on SSC9.
Sequence variation in DGAT2
Variant discovery in porcine DGAT2 was examined by retrieving all sequence variants from the coding region, the 3′-untranslated region and 500 bp upstream on the proximal promoter of the gene (SSC9 10,031,627 to 10,068,464 bp) in a subset of 199 pigs of the same line with whole-genome sequencing data available. Sequenced pigs covered all representative sire families used in the experiment. DNA samples were submitted to Centre Nacional d’Anàlisi Genòmica (CNAG-CRG, Barcelona, Spain) for sequencing. Libraries were prepared and sequenced with paired-end reads with a NovaSeq 6000 instrument (Illumina) according to the manufacturer’s protocol. Libraries were aligned to the Sscrofa11.127 using the BWA-MEM algorithm30. The average realized sequencing coverage was 6.8x (SD = 1.2×; min = 4.4×; max = 12.2×). Variants were identified following GATK HaplotypeCaller 3.8.0 software31,32. The SNP ss7315407085 G > A in exon 9 (SNP5, 9:10,065,826; Table 1) was selected for further validation with the whole set of pigs.
Genotyping DGAT2
All pigs (n = 1129) used in the experiment were genotyped for SNP ss7315407085 in exon 9 of DGAT2 using the primers described in Supplementary Table S1. Amplifications were performed by real-time PCR (QuantStudio3, Applied Biosystems, Thermo Scientific, Waltham, MA, USA) with High-Resolution Melt analysis (Luminaris Colour HRM Master Mix, Thermo Scientific) using 20 ng of genomic DNA and 0.4 µM of each primer in 5 µL final volume reaction. Thermocycling conditions were 50 °C 2 min, 95 °C 10 min, and 40 cycles of 95 °C 15 s, 60 °C 1 min, followed by a high-resolution melting curve starting with a denaturation at 95 °C for 15 s, annealing at 60 °C for 1 min and a slow ramp at 0.015 °C/s up to 95 °C. High Resolution Melt software v3.1 (Applied Biosystems, Thermo Scientific) was used for melting data analysis and sample genotyping. All pigs were also genotyped for the SCD (rs80912566 T > C; on SSC14) and leptin receptor (rs709596309 C > T; on SSC6) SNPs following the protocols described in Refs.24,33, respectively.
DGAT2 expression
DGAT2 expression was measured by quantitative real-time PCR (qPCR) in the SM of 40 pigs from a single batch. RNA was isolated with TRI-Reagent according to the manufacturer’s protocol and purity was assessed by spectrophotometry with a Nanodrop-1000. Total RNA (1.5 µg) was reverse-transcribed using SuperScript IV Reverse Transcriptase (Invitrogen, Thermo Scientific) with 100 µM random hexamers at 23 °C for 10 min, 50 °C for 20 min and 80 °C for 10 min. The cDNA was diluted 1:30 in water. Real-time PCR assays were carried out in triplicate using SYBR Green Supermix (Bio-Rad, Hercules, CA, USA), 0.2 µM of each primer and 3 µl of diluted cDNA. The B2M and RPL32 genes were used as reference genes to quantify and normalize the DGAT2 expression data. Primers used for DGAT2 expression (qDGAT2) are given in Supplementary Table S1.
Estimation of DGAT2 tag-SNP effects
The effect of the DGAT2 ss7315407085 G > A SNP genotypes on production traits (body weight, carcass weight, BT and LT) and IMF content and fatty acid composition was estimated using a mixed model that included the batch (15 levels) and the genotype for DGAT2 (GG, AG and AA), SCD (TT, CT and CC) and leptin receptor (TT, CT and CC) as fixed effects and the sire and the dam as random effects. The leptin receptor polymorphism was included in the model because of its impact on fat content and fatty acid composition33. The slaughter age, for production traits, and IMF content, for fatty acids, were added as covariates. The additive and dominant effects of the DGAT2 SNP were estimated by replacing the genotype effect with the covariates (1, 0, − 1) and (0, 1, 0) for the GG, AG and AA genotypes, respectively. Gene expression was analysed only as a function of the DGAT2 genotype. The effects of the genotype and covariates were tested using the F-statistic. Multiple pairwise comparisons among DGAT2 genotypes were tested with the Tukey HSD test. Results are presented as least-square means ± standard error. All the analyses were performed using the statistical package JMP Pro 14 (SAS Institute Inc., Cary, NC).
Consent for publication
All the authors read and agree to the content of this paper and its publication.
Results
The preliminary GWAS revealed two regions associated with C16:1n-7, one on SSC9 and another on SSC14 (Fig. 2). The region on SSC9 (9.8 Mb to 12.8 Mb) contained 18 significant SNPs (P < 1.4 × 10–6). Candidate gene DGAT2 mapped within this region, specifically at 10.0–10.1 Mb. This association was not identified in previous analyses in this Duroc line32 and evidence for potential implications of sequence variation in DGAT2 on fat content and composition in pigs is still very scarce14,21. On the other hand, the region on SSC14 matched the variant in SCD described above24. No evidence of association was found between the genomic region of DGAT1 (on SSC4) and fat content and fatty acid composition.

Genome-wide associations for palmitoleic acid content in the gluteus medius muscle. Different chromosomes are distinguished with different colours. The red line indicates the Bonferroni‐corrected genome‐wide significance threshold.
Sequence variation in DGAT2
Using whole-genome sequencing data from pigs of the same line, we identified a total of 33 sequence variants in DGAT2. Three of them were exonic, although synonymous (Table 1) and, from these, only SNP5 in exon 9 (ss7315407085) had a minor allele frequency greater than 0.25 (0.33). With 1000 samples, which are around the current sample size of our biorepository, 0.25 is the minimum minor allele frequency required to detect (P < 0.05) a difference of around 5% (0.5 SD) between genotypes for C16:1n-7 with a power of at least 80%. This SNP has been previously reported in other pig breeds and crossbreds14,21 and, in line with our results, the A allele was the minor allele in all genetic types, with a frequency that ranged from 0 to 0.32. The other 30 variants were located in the promoter region (2 SNPs) and in the 3′-UTR (21 SNPs and 7 indels). Using only the sequenced pigs, we confirmed that DGAT2 exerted an additive influence on C16:1n-7 (Supplementary Table S2), with SNP5 and the sequence variants in the interval between INDEL13 and SNP22 presenting the most relevant associations. Linkage disequilibrium of all variants was analysed with Haploview 4.2 software (Supplementary Fig. S1), showing that SNP5 was in linkage disequilibrium with both INDEL13 (r2 = 0.51) and SNP22 (r2 = 0.47). In view of these results, we selected SNP5 (ss7315407085) as a tag variant of this haplotype for further validation.
Validation of DGAT2 tag SNP
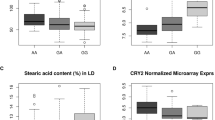
The expression of DGAT2 by ss7315407085 genotype is shown in Fig. 3. Despite the limited sample size, the results obtained indicate that the DGAT2-G show a favourable additive effect as compared to DGAT2-A on DGAT2 mRNA expression in muscle (+ 0.65 ± 0.27, P < 0.05). The DGAT2-GG pigs displayed around 1.5-fold increase in DGAT2 expression compared to the DGAT2-AA pigs (+ 1.48 ± 0.58, P < 0.05). Gene expression was measured in SM because it was the only muscle that could be sampled immediately after slaughter.

Relative expression of DGAT2 across ss7315407085 genotypes. Numbers in boxes indicate the number of pigs used per genotype (n). Error bars represent standard errors. Columns lacking a common letter differ (P < 0.05).
The effect of the DGAT2 SNP on fatty acid composition was validated in GM (Table 2) and LM (Table 3) using the whole set of pigs in the experiment. In line with preliminary association results in Supplementary Table S2, the DGAT2-G allele exerted a positive additive effect on C16:1n-7 in both muscles (+ 0.12% ± 0.03, in GM, and + 0.19% ± 0.03, in LM, P < 0.0001). The effect of the DGAT2-G allele had also a positive impact on C14:0 (+ 0.04% ± 0.01, in GM, and + 0.05% ± 0.01, in LM, P < 0.01), C16:0 (+ 0.12% ± 0.06, in GM, P = 0.05) and C18:1n-7 (+ 0.06% ± 0.02, in GM, P = 0.01). However, the effect of the DGAT2-G allele was proportionally greater in C16:1n-7 than in C16:0, as evidenced by subsequent changes in the C16:1n-7 to C16:0 ratio (+ 0.04 ± 0.01, in GM, and + 0.07 ± 0.01, in LM, P < 0.01, values × 10). In contrast, the DGAT2-G allele affected negatively C18:0 and C18:1n-9, particularly in LM (− 0.13% ± 0.06, P < 0.05, and − 0.40% ± 0.15, P < 0.01, respectively).
The DGAT2 SNP did not show dominance effects nor evidence for interaction with the SCD genotype for any of the fatty acids. The additive behaviour of the DGAT2-G allele was mantained across all SCD genotypes (Fig. 4). Thus, the maximum difference in C16:1n-7, which accounted for around 25% of the mean, occurred between the two extreme DGAT2/SCD haplotypes (from 4.05% ± 0.05, for GG/TT, to 3.23% ± 0.08, for AA/CC, P < 0.0001). Despite this, the difference between DGAT2-GG and AA genotypes was greater for the SCD-CC genotype (+ 0.32% ± 0.09, P < 0.001) than for the SCD-TT genotype (+ 0.15% ± 0.10, P = 0.13).

Effect of DGAT2 by SCD genotype on palmitoleic acid (C16:1n-7) in muscle gluteus medius. Numbers in boxes indicate the number of pigs used per genotype (n). Error bars represent standard errors. Columns lacking a common letter differ (P < 0.05).
We did not find evidence of consistent effects of DGAT2 on body weight or carcass fat content (Table 4) or on the fatty acid composition of SF (Supplementary Table S3) or liver (Supplementary Table S4). Only a minor effect on live body weight was observed (Table 4). Although DGAT2-GG pigs weighted 2.0 kg ± 0.7 (P < 0.05) less than DGAT2-AG pigs, the G allele did not show a clear additive behaviour (− 1.0 kg ± 0.6, P = 0.08).
Discussion
Understanding the regulation of fat metabolism is a milestone in the prevention and treatment of human lipid diseases, such as obesity, and in animal science. In this context, DGAT2 has received attention as the enzyme that catalyzes the final step in the synthesis of TG, the most common type of body fat. Here, we validated the effects of the ss7315407085 G > A SNP on fat content and fatty acid composition as a tag SNP for the haplotype in DGAT2 that was identified following a preliminary GWAS and after mRNA expression and sequence variant association analyses. Using a larger sample, we confirmed that the minor allele A segregates in the studied Duroc line at a moderate frequency (0.33), and that it has a negative impact on C16:1n-7 content in muscle. The moderate minor allele frequency in the studied Duroc line is similar to estimates from other Duroc lines14 and White and Asian breeds and crossbreds14,21,22.
Our validation scheme evidenced a consistent effect of DGAT2 SNP on C16:1n-7 in muscle that is independent of IMF content. A genetic variant in SCD with relevant effects on MUFA also segregates in this line24. The DGAT2 SNP had a lower impact on C16:1n-7 than the SCD SNP variant, particularly in terms of genetic variance. While the additive value of DGAT2 for C16:1n-7 was around half of that of SCD, the additive variance only reached 22% of that attributed to SCD. It has been shown that SCD co-expresses with DGAT234 and that their respective enzymes locate very close in the endoplasmic reticulum11,17, two findings that support the hypothesis that SCD is involved in the TG synthesis by providing a more accessible pool of MUFA through substrate channelling12,17. This also would explain why cells overexpressing DGAT2 show a greater proportion of MUFA and very particularly of C16:1n-734. Our results are in line with this hypothesis, because C16:1n-7 was the fatty acid most affected by the DGAT2 SNP and the G allele, the one with higher DGAT2 expression, was the one that led to accumulate more C16:1n-7. However, we did not find evidence for a positive interaction between SCD and DGAT2 SNPs for C16:1n-7. Using RNAseq data from 40 other pigs of the same line, we were able to confirm that DGAT2 genotype (GG: n = 19, AG: n = 14, AA: n = 5) affected muscular DGAT2 expression, but we did not detect differential expression of SCD by DGAT2 genotype. In absence of interaction, the two genes mostly behaved in an allele-dose manner (Fig. 3).
The positive effect of the DGAT2 G allele on C16:1n-7 was not observed on C18:1n-9. Zhang et al.34 evidenced that the impact of DGAT2 on C16:1n-7 was proportionally greater as fatty acid composition turned more dependent on de novo fatty acids. For C18:1n-9, the same effect was only seen in mature adipocytes, especially when they were cultured in a fatty acid-rich medium. In line with these results, a plausible explanation to the differential trend between C16:1n-7 and C18:1n-9 could be that the most immediate (short-term) effect of DGAT2 is to alter fatty acid composition by selectively capturing as substrate the first available de novo fatty acids rather than to enhance the endogenous biosynthesis of longer-chain fatty acids. This may be the case here, since differences in DGAT2 expression across genotypes were relatively small and no correlated responses in IMF content and SCD expression were observed. The favourable effect of the G allele on C14 and C16 fatty acids would reinforce this hypothesis. On the other hand, the impact of de novo fatty acids on final fatty acid composition is more apparent in C16:1n-7 and C18:1n-7 than in C18:1n-9, which is proportionally more abundant in pig feed than C16:1n-7 and thus more likely to be influenced by the diet. Similarly, the effect of DGAT2 on fatty acid composition was only observed in IMF, which is precisely the adipose tissue that develops later35 and is less sensitive to dietary fat36.
Changes in DGAT2 expression are related to lipid accumulation. Results in human and murine cells indicate that DGAT2 promotes TG synthesis and storage in cytosolic lipid droplets37 and accordingly DGAT2 is expressed more abundantly in cells with greater de novo fatty acid synthesis19. This is in line with findings in pigs, in which DGAT2 was most expressed in the liver of fatter breeds, where TG are expected to be more actively synthesized38. The expression of DGAT2 was twofold higher in fatter than in leaner breeds and around fivefold and tenfold higher in liver than in subcutaneous fat and muscle, respectively. However, the mRNA expression of DGAT2 was positively correlated with IMF content but not with BT. In view of these results, it would appear that the pigs carrying the DGAT2 A allele, which down-regulates the expression of DGAT2, should be less fatty and, specifically, have less IMF content. Even though this association was reported in other studies14,21, we did not find evidence that DGAT2 A entails a reduction in IMF content, BT or even fat content in liver. This may contradict biological expectations, but, in comparison with previous reports, validation here was performed using a much larger set of pigs from a single line so as to avoid breed or family biases. A 13-bp deletion in DGAT2 3′-UTR region has also been associated with increased DGAT2 mRNA expression and BT39. This deletion allele showed higher transcriptional activity, most likely owing to a less stable 3′-UTR secondary structure. This indel was located at 905 bp downstream from the stop codon and matches INDEL14 (Table 1). In line with the results observed for SNP5, none of these polymorphisms including INDEL14 (r2 = 0.57, Supplementary Fig. S1) affected BT in our Duroc line (Supplementary Table S2).
The results obtained are more appealing for lipid research than for pig breeding. In recent years, C16:1n-7 has received a lot of attention for its potential role as a lipokine, i.e. as a lipid hormone that acts in distant organs. Despite this, the effects of C16:1n-7 are still under debate. On the one hand, preclinical experiments with cell and rodent models show that C16:1n-7 supplementation has anti-inflammatory properties40,41 that protect against metabolic disorders. But, on the other hand, research in humans reported elevated blood levels of C16:1n-7 in patients with obesity and metabolic syndrome42,43. Findings in pigs may contribute to disentangle the biological implications of C16:1n-7 as well as to understand the genetic regulation of fat metabolism.
Conclusions
A sequence variant in the porcine DGAT2 influences muscular gene expression and fatty acid composition, with changes mostly limited to swapping C18 fatty acids for C14 and C16 fatty acids of the same degree of saturation. Differences across DGAT2 genotypes are quantitatively small, but the variant that enhances DGAT2 expression is associated with increased content of shorter-chain fatty acids, thereby indicating that DGAT2 preferentially uptakes the first available de novo fatty acids as substrate. Although DGAT2 and SCD act additively, the fact that DGAT2 overexpression impacts C16:1n-7 more than C16:0 indicates that DGAT2 and SCD are closely interlinked. Further enzyme characterization is needed, but our findings provide evidence that C16:1n-7 is the immediate substrate for DGAT2 and corroborate that fatty acid metabolism in pigs is subjected to subtle tissue-specific genetic regulatory mechanisms.
Data availability
Data that support the findings of this study are available within the article and Supplementary Information, or from the authors upon reasonable request.
Abbreviations
- BT:
-
Backfat thickness
- DGAT:
-
Diacylglycerol O-acyltransferase
- GM:
-
Gluteus medius muscle
- GWAS:
-
Genome-wide association study
- IMF:
-
Intramuscular fat
- LM:
-
Longissimus thoracis muscle
- LT:
-
Loin thickness
- MUFA:
-
Monounsaturated fatty acids
- PUFA:
-
Polyunsaturated fatty acids
- SCD:
-
Stearoyl-CoA desaturase
- SD:
-
Standard deviation
- SF:
-
Subcutaneous fat
- SFA:
-
Saturated fatty acids
- SM:
-
Semimembranosus muscle
- SNP:
-
Single nucleotide polymorphism
- TG:
-
Triacylglycerols
References
Zhu, Y., Bo, Y. & Liu, Y. Dietary total fat, fatty acids intake, and risk of cardiovascular disease: A dose-response meta-analysis of cohort studies. Lipids Health Dis. 18(1), 91 (2019).
Posner, B. M. et al. Dietary lipid predictors of coronary heart disease in men: The Framingham Study. Arch. Intern. Med. 151, 1181–1187 (1991).
Ockene, I. S. et al. Dietary fat intake and the risk of coronary heart disease in women. N. Engl. J. Med. 337, 1491–1499 (1997).
Griel, A. & Kris-Etherton, P. Beyond saturated fat: The importance of the dietary fatty acid profile. Nutr. Rev. 64, 257–262 (2006).
de Lorgeril, M. & Salen, P. New insights into the health effects of dietary saturated and omega-6 and omega-3 polyunsaturated fatty acids. BMC Med. 10(1), 50 (2012).
Ngapo, T. M., Martin, J. F. & Dransfield, E. International preferences for pork appearance: I. Consumer choices. Food Qual. Prefer. 18, 26–36 (2007).
Wood, J. D. et al. Effects of fatty acids on meat quality: A review. Meat Sci. 66(1), 21–32 (2004).
Wood, J. D. et al. Fat deposition, fatty acid composition and meat quality: A review. Meat Sci. 78(4), 343–358 (2008).
Coleman, R. A. & Lee, D. P. Enzymes of triacylglycerol synthesis and their regulation. Prog. Lipid Res. 43(2), 134–176 (2004).
Harris, C. A. et al. DGAT enzymes are required for triacylglycerol synthesis and lipid droplets in adipocytes. J. Lipid Res. 52(4), 657–667 (2011).
Bhatt-Wessel, B., Jordan, T. W., Miller, J. H. & Peng, L. Role of DGAT enzymes in triacylglycerol metabolism. Arch. Biochem. Biophys. 655, 1–11 (2018).
Liu, Q., Siloto, R. M. P., Lehner, R., Stone, S. J. & Weselake, R. J. Acyl-CoA:diacylglycerol acyltransferase: Molecular biology, biochemistry and biotechnology. Prog. Lipid Res. 51(4), 350–377 (2012).
Karantonis, H., Nomikos, T. & Demopoulos, C. Triacylglycerol metabolism. Curr. Drug Targets. 10(4), 302–319 (2009).
Yin, Q., Yang, H. W., Han, X. L., Fan, B. & Liu, B. Isolation, mapping, SNP detection and association with backfat traits of the porcine CTNNBL1 and DGAT2 genes. Mol. Biol. Rep. 39, 4485–4490 (2012).
Wältermann, M., Stöveken, T. & Steinbüchel, A. Key enzymes for biosynthesis of neutral lipid storage compounds in prokaryotes: Properties, function and occurrence of wax ester synthases/acyl-CoA:diacylglycerol acyltransferases. Biochimie 89(2), 230–242 (2007).
Stone, S. J. et al. Lipopenia and skin barrier abnormalities in DGAT2-deficient mice. J. Biol. Chem. 279(12), 11767–11776 (2004).
Man, W. C., Miyazaki, M., Chu, K. & Ntambi, J. Colocalization of SCD1 and DGAT2: Implying preference for endogenous monounsaturated fatty acids in triglyceride synthesis. J. Lipid Res. 47(9), 1928–1939 (2006).
Chitraju, C., Walther, T. C. & Farese, R. V. The triglyceride synthesis enzymes DGAT1 and DGAT2 have distinct and overlapping functions in adipocytes. J. Lipid Res. 60(6), 1112–1120 (2019).
Wurie, H. R., Buckett, L. & Zammit, V. A. Diacylglycerol acyltransferase 2 acts upstream of diacylglycerol acyltransferase 1 and utilizes nascent diglycerides and de novo synthesized fatty acids in HepG2 cells. FEBS J. 279(17), 3033–3047 (2012).
Ning, T. et al. Genetic interaction of DGAT2 and FAAH in the development of human obesity. Endocrine 56(2), 366–378 (2017).
Renaville, B., Bacciu, N., Lanzoni, M., Corazzin, M. & Piasentier, E. Polymorphism of fat metabolism genes as candidate markers for meat quality and production traits in heavy pigs. Meat Sci. 110, 220–223 (2015).
Renaville, B., Bacciu, N., Lanzoni, M., Mossa, F. & Piasentier, E. Association of single nucleotide polymorphisms in fat metabolism candidate genes with fatty acid profiles of muscle and subcutaneous fat in heavy pigs. Meat Sci. 139, 220–227 (2018).
Gol, S., Pena, R. N., Rothschild, M. F., Tor, M. & Estany, J. A polymorphism in the fatty acid desaturase-2 gene is associated with the arachidonic acid metabolism in pigs. Sci. Rep. 8(1), 14336 (2018).
Estany, J., Ros-Freixedes, R., Tor, M. & Pena, R. N. A functional variant in the stearoyl-CoA desaturase gene promoter enhances fatty acid desaturation in pork. PLoS One 9(1), 1–11 (2014).
Ros-Freixedes, R., Reixach, J., Bosch, L., Tor, M. & Estany, J. Genetic correlations of intramuscular fat content and fatty acid composition among muscles and with subcutaneous fat in Duroc pigs. J. Anim. Sci. 92(12), 5417–5425 (2014).
Bosch, L., Tor, M., Reixach, J. & Estany, J. Estimating intramuscular fat content and fatty acid composition in live and post-mortem samples in pigs. Meat Sci. 82(4), 432–437 (2009).
Warr, A. et al. An improved pig reference genome sequence to enable pig genetics and genomics research. Gigascience. 9(6), 1–14 (2020).
Zhou, X. & Matthew, S. Genome-wide efficient mixed-model analysis for association studies. Nat. Gene. 44, 821–824 (2012).
Chen, E. Y. et al. Enrichr: interactive and collaborative HTML5 gene list enrichment analysis tool. BMC Bioinformatics 15(14), 128 (2013).
Li, H. Aligning sequence reads, clone sequences and assembly contigs with BWA-MEM. arXiv:1303.3997v1 (2013).
DePristo, M. et al. A framework for variation discovery and genotyping using next-generation DNA sequencing data. Nat. Genet. 43(5), 491–498 (2011).
Poplin, R. et al. Scaling accurate genetic variant discovery to tens of thousands of samples. bioRxiv. 10, 1–22 (2017).
Ros-Freixedes, R. et al. Genome-wide association study singles out SCD and LEPR as the two main loci influencing intramuscular fat content and fatty acid composition in Duroc pigs. PLoS One 11(3), 1–18 (2016).
Zhang, Z. et al. Effect of diacylglycerol acyltransferase 2 overexpression in 3T3-L1 is associated to an increase in mono-unsaturated fatty acid accumulation. J. Anim. Sci. Biotechnol. 5(1), 1–8 (2014).
Bosch, L., Tor, M., Reixach, J. & Estany, J. Age-related changes in intramuscular and subcutaneous fat content and fatty acid composition in growing pigs using longitudinal data. Meat Sci. 91(3), 358–363 (2012).
Nürnberg, K., Wegner, J. & Ender, K. Factors influencing fat composition in muscle and adipose tissue of farm animals. Livest. Prod. Sci. 56(2), 145–156 (1998).
McFie, P. J., Banman, S. L., Kary, S. & Stone, S. J. Murine diacylglycerol acyltransferase-2 (DGAT2) can catalyze triacylglycerol synthesis and promote lipid droplet formation independent of its localization to the endoplasmic reticulum. J. Biol. Chem. 286(32), 28235–28246 (2011).
Cui, J. X. et al. The effects of DGAT1 and DGAT2 mRNA expression on fat deposition in fatty and lean breeds of pig. Livest. Sci. 140(1–3), 292–296 (2011).
Zhao, Z., Kang, L., Zhang, X., Yang, C. & Jiang, Y. Identification of a 13 bp indel polymorphism in the 3′-UTR of DGAT2 gene associated with backfat thickness and lean percentage in pigs. Gene 576(2), 729–733 (2016).
Çimen, I. et al. Prevention of atherosclerosis by bioactive palmitoleate through suppression of organelle stress and inflammasome activation. Sci. Transl. Med. 8(358), 1–15 (2016).
de Souza, C. O., Vannice, G. K., Rosa Neto, J. C. & Calder, P. C. Is palmitoleic acid a plausible non-pharmacological strategy to prevent or control chronic metabolic and inflammatory disorders?. Mol. Nutr. Food Res. 62(1), 1–31 (2017).
de Souza, C. O. et al. Palmitoleic acid has stronger anti-inflammatory potential in human endothelial cells compared to oleic and palmitic acids. Mol. Nutr. Food Res. 62(20), 1–20 (2018).
Frigolet, M. E. & Gutiérrez-Aguilar, R. The role of the novel lipokine palmitoleic acid in health and disease. Adv. Nutr. 8(1), 173S-181S (2017).
Acknowledgements
We acknowledge Dr Rayner González-Prendes for collaborating in preliminary GWAS and Pilar Sopeña for helping in the maintenance of the UdLGIM biobank. We also thank Josep Reixach and the personnel at Selección Batallé for their cooperation for the recording of on-farm data and sample collection.
Funding
This research was supported by the Spanish Ministry of Science, Innovation & Universities and the EU Regional Development Funds (Grant RTI2018-101346-B-I00). ES is recipient of a PhD scholarship from the University of Lleida.
Author information
Authors and Affiliations
Contributions
Conceptualization and funding acquisition: J.E. and R.R.F.; methodology and investigation: E.S., M.T. and R.N.P.; data editing and analyses: E.S., J.E. and R.R.F.; writing of original draft: E.S. and J.E. All authors reviewed, edited and approved the final manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Solé, E., Ros-Freixedes, R., Tor, M. et al. A sequence variant in the diacylglycerol O-acyltransferase 2 gene influences palmitoleic acid content in pig muscle. Sci Rep 11, 14797 (2021). https://doi.org/10.1038/s41598-021-94235-z
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-021-94235-z
This article is cited by
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
