
Abstract
This study was aimed to evaluate the differences in the composition of gut microbiota, tryptophan metabolites and short-chain fatty acids in feces between volunteers who frequently ate chicken and who frequently ate pork. Twenty male chicken-eaters and 20 male pork-eaters of 18 and 30 years old were recruited to collect feces samples for analyses of gut microbiota composition, short-chain fatty acids and tryptophan metabolites. Chicken-eaters had more diverse gut microbiota and higher abundance of Prevotella 9, Dialister, Faecalibacterium, Megamonas, and Prevotella 2. However, pork-eaters had higher relative abundance of Bacteroides, Faecalibacterium, Roseburia, Dialister, and Ruminococcus 2. In addition, chicken-eaters had high contents of skatole and indole in feces than pork-eaters, as well as higher contents of total short chain fatty acids, in particular for acetic acid, propionic acid, and branched chain fatty acids. The Spearman’s correlation analysis revealed that the abundance of Prevotella 2 and Prevotella 9 was positively correlated with levels of fecal skatole, indole and short-chain fatty acids. Thus, intake of chicken diet may increase the risk of skatole- and indole-induced diseases by altering gut microbiota.
Similar content being viewed by others
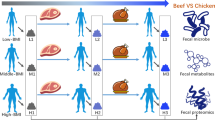


Introduction
In recent decades, eating habits have changed dramatically1. These changes have various effects on human health2. Several chronic diseases, such as cardiovascular diseases and diabetes are closely related to long-term unhealthy dietary habits3. In many countries, meat is an indispensable part of routine meals because meat is rich in high-quality protein, vitamins and minerals4,5. Preferences for different types of meat vary between and within countries depending on demographic and cultural influences. Protein composition differs between different types of meat, which may modulate the composition of gut microbiota and formation of their metabolites6.
Meat and meat products are the most important source for L-tryptophan (L-trp), which can be transformed into skatole and indole by the gut microbiota in the large intestine. Skatole was shown to be pneumotoxic to humans and associated with hepatic encephalopathy and saccharo-butyric putrefaction, whereas increased indole and oxindole were observed in cirrhotic patients’ blood7,8,9,10. Dietary fiber and peptides can be degraded by the gut microbiota into short-chain fatty acids (SCFAs). SCFAs may affect lipid metabolism, bacterial fermentation and the capacity of intestinal absorption including the absorption rate of skatole in gut11,12.
Many studies have been focused on the effects of meat consumption on the composition of gut microbiota, however, only few studies investigated an impact of meat consumption on tryptophan metabolites. In a previous study, we found that a high fat high chicken or pork protein diet increased the abundance of gut microbiota associated with skatole and indole production in Wistar rats13. In the present study, we investigated the differences in composition of gut microbiota, tryptophan metabolites and SCFAs in feces in volunteers who frequently consume chicken or pork products.
Results
Skatole, indole and SCFAs in fecal samples
The concentrations of fecal skatole and indole were higher in chicken-eaters (P < 0.05, Fig. 1). Chicken-eaters also had significantly higher concentrations of total SCFAs as well as acetic acid, propionic acid, isovaleric acid, and branched chain fatty acids (BCFAs) than pork eaters (P < 0.05, Table 1).

Chicken-eaters had higher levels of skatole and indole in feces than pork-eaters. The means were compared by Student’s t test. The asterisk (*) indicates significant difference between two groups (P < 0.05).
Gut microbiota composition and its correlations with skatole, indole and SCFAs
Richness and diversity
A total of 3787 and 1630 OTUs were identified in the fecal samples from chicken and pork eaters, respectively. The number of shared OTUs between the two groups was 1472. The numbers of OTUs specific for chicken- and pork-eaters were 2315 and 158, respectively (Fig. 2A). The diversity of gut microbiota in pork-eaters was relatively low, indicating that pork consumption reduced the diversity of gut microbiota. PCoA reveals responses of volunteers’ gut microbiota to the two dietary modes (Fig. 2B). The first and second principal components (PC1 and PC2) explained 56.1% of the total variance in gut microbiota composition, in which PC1 accounted for 43.9% and PC2 accounted for 12.2%. PC1 mainly explained the inter-group variation due to dietary effects, while PC2 explained the intra-group variation from individual volunteers. The two groups can be well separated in the first principal component, indicating that the gut microbiota responded differently to the two dietary modes. Analysis of the α diversity of samples in the two groups revealed that both Chao and Shannon indexes of pork-eaters were significantly lower than those in chicken-eaters (P < 0.01, Table 2), suggesting that pork protein reduced the diversity of gut microbiota.

Chicken-eaters had higher diversity of the gut microbiota than pork-eaters. (A) Venn diagram of fecal microbiota at the OTU level. (B) PCoA plot of fecal microbiota at the OTU level. Each point represents one sample.
Gut microbiota composition
At the phylum level, Firmicutes, Bacteroidetes, Proteobacteria and Actinobacteria were the four most abundant phyla, with the range of 35.3–94.7%, 2.6–62.1%, 0.2–14.3% and 0.1–6.9%, respectively (Fig. 3A). In pork-eaters, Firmicutes had the highest relative abundance (58.2%), followed by Bacteroidetes (34.4%), Actinobacteria (2.8%) and Proteobacteria (2.6%). The dominant phyla in chicken-eaters were Firmicutes (71.2%), Bacteroidetes (21.3%), Proteobacteria (4.2%) and Actinobacteria (1.8%). Relative abundance of Firmicutes in pork-eaters was significantly higher than that in chicken-eaters (P < 0.01), but relative abundance of Bacteroidetes was lower in pork-eaters (P < 0.01).

Composition of fecal microbiota at the phylum and genus levels. (A) Hierarchical clustering of fecal microbiota at the phylum level. Each line and bar represents one sample. (B) Composition of fecal microbiota at the genus level. Each column represents one sample and each color represents one genus.
At the genus level, the dominant bacteria are quite different (Fig. 3B). In chicken-eaters, relative abundance of Prevotella 9 was the highest (22.45%), followed by Dialister (9.5%), Faecalibacterium (9.0%), Megamonas (4.6%), Prevotella 2 (4.2%), Roseburia (3.6%), Alloprevotella (3.2%), Ruminococcaceae UCG-002 (3.2%), [Eubacterium] coprostanoligenes group (2.5%), and Succinivibrio (1.7%). In pork-eaters, relative abundance of Bacteroides was the highest (17.3%), followed by Faecalibacterium (14.6%), Roseburia (7.4%), Dialister (4.6%), Ruminococcus 2 (3.7%), Blautia (2.6%), Megamonas (2.6%), Agathobacter (2.5%), Subdoligranulum (2.1%), and [Eubacterium] coprostanoligenes group (2.1%). Interestingly, relative abundance of Olsenella was higher in chicken-eaters (0.5%) than in pork-eaters (0.03%). Olsenella scatoligenes has been reported a skatole-producing bacterium in the intestine of pigs14.
Linear discriminant analysis of fecal microbiota
LEfSe analysis showed a significant difference in 142 species between the two groups (P < 0.05, Fig. 4). Among them, 55 species had higher abundance in chicken-eaters while 87 species had higher abundance in pork-eaters. Chicken-eaters had higher abundance of Prevotellaceae, Prevotella 9, Bacteroidales, Dialister, Prevotella 2, Ruminococcaceae UCG 002, Lactobacillus, and Olsenella. Pork-eaters had higher abundance of Clostridiales, Bacteroides, Firmicutes, Lachnospiraceae, Faecalibacterium, Roseburia, Ruminococcus 2, and Blautia.

Chicken-eaters had more different genera than pork-eaters. (A) Biomarker taxons generated from LEfSe analysis (LDA > 2). Red bars represent samples from pork-eaters and green bars represent samples from chicken-eaters. (B) Cladogram obtained from LEfSe analysis with presenting various levels (phylum, class, order, family and genus) from inner to outer rings. Different color represents different groups. Different letters indicate specific gut microbial taxa from phylum to genus associated with different diets.
Key species involved in regulating skatole, indole and SCFAs
In order to find relationships among gut microbiota, skatole, indole and SCFAs, we performed Spearson’s correlation analysis. As shown in Fig. 5A, at the phylum level, relative abundance of Elusimicrobia, Actinobacteria, Bacteroidetes, Cyanobacteria and Tenericutes were positively correlated with skatole level. Both abundance of these bacteria and skatole level were higher in chicken-eaters than those in pork-eaters (P < 0.05). Relative abundance of Firmicutes and Fusobacteria were negatively correlated with skatole level. These two phyla were more abundant in pork-eaters which also had lower skatole level. Relative abundance of Actinobacteria, Bacteroidetes and Tenericutes were higher in chicken-eaters and were positively correlated with acetic acid levels. Relative abundance of Verrucomicrobia, Proteobacteria and Firmicutes was negatively correlated with acetic acid levels. Relative abundance of Bacteroidetes and Firmicutes was positively and negatively correlated with propionic acid levels, respectively. Relative abundance of Elusimicrobia was positively correlated with valeric acid level. Relative abundance of Verrucomicrobia, Fusobacteria and Patescibacteria was negatively correlated with valeric acid levels. Relative abundance of Fusobacteria and Patescibacteria was negatively correlated with isobutyric acid levels. Fusobacteria was abundant in pork-eaters and its relative abundance was negatively correlated with isovaleric acid level. Relative abundance of Synergistetes was negatively correlated with isovaleric acid level.


Abundance of the gut microbiota was significantly associated with levels of metabolites. (A) at the phyla level; (B) at the genus level. Green color represents positive correlation and red color represents negative correlation. Correlation was considered significant when the absolute value of Spearman’s rank correlation coefficient (Spearman’s r) was > 0.6 and statistically significant (P < 0.05). The asterisks indicate significant correlation, *P < 0.05; **P < 0.01; ***P < 0.001.
As shown in Fig. 5B, relative abundance of genera Prevotella 9, Prevotella 7, Prevotella 2, Alloprevotella, Succinivibrio, Lactobacillu, Ruminococcaceae UCG-014, Ruminococcaceae UCG-005 and Catenibacterium were positively correlated with skatole level, and their abundance was higher in chicken-eaters (P < 0.05). Relative abundance of Bacteroides, Ruminococcus 2, Lachnoclostridium, Alistipes, Parasutterella, Fusobacterium and [Eubacterium] hallii group were negatively correlated with skatole level, and their relative abundance was higher in pork-eaters but skatole level was lower. Relative abundance of Dialister was positively correlated with indole and isobutyric acid levels. As mentioned above, Chicken-eaters had higher levels of indole and isobutyric acid, and higher relative abundance of Dialister. Relative abundance of [Eubacterium] eligens group and Methanobrevibacter were negatively correlated with indole level. Relative abundance of Prevotella 9, Prevotella 2, Alloprevotella, Succinivibrio, Lactobacillus and Collinsella were positively correlated with levels of acetic acid and propionic acid, while relative abundance of Bacteroides, Blautia, Phascolarctobacterium, Escherichia–Shigella, Parabacteroides, Veillonella, Fusicatenibacter and [Eubacterium] hallii group were negatively correlated with levels of acetic acid and propionic acid. Relative abundance of Roseburia, Ruminococcus 2, Subdoligranulum, Veillonella, and Haemophilus were negatively correlated with butyric acid level. Finally, relative abundance of Dialister, Ruminococcaceae UCG 002 and Lactobacillus were positively correlated with levels of valeric acid and isovaleric acid.
Discussion
Meat is an important source of high quality protein and essential amino acids such as tryptophan in human body15,16. Consumers may have different dietary habits, which has a close association with the composition of gut microbiota. Preferences for meat type differ between consumers. In a rat study with strict design, we observed that high meat protein diets caused the dysbiosis of gut microbiota and tryptophan metabolism13. In the present study, we for the first time investigated the gut microbiota and tryptophan metabolism in volunteers who frequently eat chicken or pork products.
Eating habits are associated with the gut microbiota composition and the profiles of gut metabolites. Dietary components, e.g., protein, fat and sugars may mediate the diet effect on host physiology and metabolism by altering the gut microbiota composition17. SCFAs are important metabolites of dietary fiber and protein fermented by gut microbiota18 and can be used as an important energy source of intestinal epithelial cells, but also affect the function of intestinal barrier19,20.
Meat contains high amount of tryptophan, which plays an important role in the balance between intestinal immune tolerance and the maintenance of gut microbiota. Tryptophan is absorbed in the small intestine, and the unabsorbed part reaches the cecum and colon, and is metabolized by intestinal bacteria to indole and indolic compounds, including skatole21. Indolic compounds can play a protective role in liver diseases including alcoholic liver disease, non-alcoholic liver disease and non-alcoholic fatty liver disease through a variety of mechanisms. Indole can promote L cells to release GLP-1 and enter the circulatory system to play an antioxidant role22. Small molecules produced by tryptophan metabolism, such as skatole, are sources for activating aromatic hydrocarbon receptors in different immune cells. These metabolites can regulate immunity and immune response through aromatic hydrocarbon receptors and maintain intestinal immune balance23,24,25,26. Skatole is an aryl hydrocarbon receptor agonist, which induces the expression of aryl hydrocarbon receptor regulatory genes, such as CYP1A2. Indole affect intestinal permeability and host immunity27. Skatole were shown to be pneumotoxic to humans and associated with hepatic encephalopathy and saccharo-butyric putrefaction7,8,9,10. Therefore, the balance of tryptophan metabolites in the gut is beneficial to maintain intestinal homeostasis and plays an important role in body health.
The present study indicated great differences in the composition of gut microbiota between chicken-eaters and pork-eaters. Pork-eaters had lower diversity of gut microbiota than chicken-eaters. Bacteroides, Faecalibacterium, Roseburia, Dialister and Ruminococcus 2 were found more abundant in the feces of pork-eaters. On the other hand, Prevotella 9, Dialister, Faecalibacterium, Megamonas, and Prevotella 2 were more abundant in the feces of chicken-eaters. Such a difference could be attributed to dietary structure that is more complex than a strictly controlled diet in our previous rat study13. In that study, chicken and pork protein diets increased the abundance of Lactobacillus, the Family XIII AD3011 group, and Desulfovibrio in rat large intestine relating to the production of skatole and indole13. A previous comparative study revealed that differences in dietary habits led to significant differences in the composition of gut microbiota, in which African children who eat foods rich in plant fiber had higher levels of Prevotella and Xylanibacte, as well as SCFAs and Enterobacter28. A study comparing the gut microbiota of Japanese and Indian with different dietary preferences showed that the abundance of Bacteroides was higher in Japanese, while the abundance of Prevotella was higher in Indian29. The altered gut microbiota may affect tryptophan metabolism30,31. Chicken-eaters had higher fecal skatole and indole, and higher abundance of Prevotella 2 and Prevotella 9. Chicken-eaters exhibited significantly higher contents of T-SCFAs. This could be because more diet-derived substances went into the large intestine for the fermentation of gut microbiota.
In summary, the effect of meat diet mode was explored on human gut microbiota composition and tryptophan metabolites, i.e., skatole and indole. The dietary mode had a significant effect on the gut microbiota composition and the levels of skatole and indole. Chicken-eaters had higher relative abundance of Prevotella 9, Prevotella 2, Dialister, Faecalibacterium, and Megamonas, and higher levels of SCFAs and tryptophan metabolites. In particular, relative abundance of Prevotella 2 and Prevotella 9 was positively correlated with levels of fecal skatole, indole and SCFAs. However, pork-eaters had higher relative abundance of Bacteroides, Dialister, Faecalibacterium, Roseburia, and Ruminococcus 2. The results give an indication that long-term intake of chicken diet may increase the risk of skatole- and indole-induced diseases by altering gut microbiota.
Methods
Volunteers and sample collection
Male volunteers of 18 and 30 years old were recruited to carry out a dietary questionnaire (supplementary file 1) to investigate the dietary habits and medical history, and to collect their living habitual and long-term dietary information. In the study, 40 eligible healthy volunteers who did not consume any antibiotics for 3 months participated. Informed consent was issued to volunteers who met the sampling standards, and the experiment was conducted after obtaining written consent. According to the survey results of dietary structure, volunteers were divided into two groups, namely the chicken-eaters (n = 20) and pork-eaters (n = 20). Chicken-eaters prefer chicken as the main meat dishes, whereas pork-eaters prefer pork as the main meat dishes.
The sterilized feces collectors were distributed to volunteers. Fecal samples were collected at morning and frozen in liquid nitrogen, and stored at − 80 °C for further analyses. This study followed the Declaration of Helsinki. The research protocol was approved by the Ethics Committee of Nanjing Agricultural University (Nanjing) and registered in the World Health Organization clinical trial registration platform (WHO ICTRP) first-level registration institutions (registration number: ChiCTR1800015339). All the procedures were conducted in strict accordance with the approved guidelines.
Skatole and indole quantification
Ultraperformance liquid chromatography (UPLC, Agilent) was used to quantify skatole and indole in feces as we previously described13. Briefly, skatole and indole were extracted by mixing fecal samples (50 mg) with 500 μL of methanol and then centrifuging for 10 min at 3000g. The supernatants (4 µL) were separated in a C18 column (1.8 μm, 50 × 2.1 mm, Agilent, California) and analytes were detected by a UPLC with a fluorescence detector (G1321B, λex = 280 nm, λem = 360 nm). The mobile phase A was ultra-pure water, and mobile phase B was acetonitrile. The elution conditions were as follows: 0–6 min, 90% A; 6–10 min, 60% A; and 10–12 min, 40% A. By comparing with retention time of standards, contents of skatole and indole in samples were qualitatively determined, and the concentration was calculated by the standard curve using an external standard method.
SCFAs analysis
Fecal samples (40 mg) were diluted in 200 μL ddH2O and centrifuged for 10 min at 12,000g. An internal standard (crotonic acid) was prepared by mixing 34 μL of crotonic acid with 170 μL of the supernatant. Centrifugation was repeated once. The supernatant (1 mL) was injected into a gas chromatograph (GC-2010 Plus Shimadzu, Japan) equipped with a HP-INNOWax capillary column (30 m × 0.25 mm × 0.25 μm, Agilent Technologies, CA). The chromatographic parameters were performed with a capillary pipette column and a hydrogen ion flame detector. The temperatures for column, vaporization and detection were set 130 °C, 180 °C, and 180 °C, respectively. The carrier gas was nitrogen with a pressure of 60 kPa. The oxygen pressure was set at 50 kPa. The sensitivity (gear) was 10–1, and the attenuation was 3.0.
16S rRNA gene sequencing and data analysis
Total genome DNA was extracted from fecal samples with an Omega Bio-tek DNA extraction kit (Norcross, GA). DNA samples were amplified with universal primers F341 (5ʹ-CCTAYGGGRBGCASCAG-3ʹ) and R806 (5ʹ -GGACTACNNGGGTATCTAAT-3ʹ). The pair-end library was constructed and the amplicons were sequenced on an Illumina MiSeq platform. Sequencing data were analyzed as previously described13, including bacterial diversity (Shannon and Simpson) and community richness (Chao) of operation taxonomy units (OTUs). The UniFrac for principal coordinate analysis (PCoA) was done to estimate the beta diversity of gut microbiota. Linear discriminant analysis effect size (LEfSe) was performed to identify the bacterial biomarkers between chicken-eaters and pork-eaters. The Spearman’s correlation coefficients between gut microbiota and skatole, indole or SCFAs (r > 0.6, P < 0.05) were visualized on the R program (version 3.2.4, https://www.r-project.org).
Statistical analysis
The differences between the two groups in measured variables were evaluated by Student’s t-test. The significance level was set as 0.05. The SAS program was applied for the analyses (SAS, version 8.0.1, Cary, NC).
References
Zhang, M. & Yang, X. J. Effects of a high fat diet on intestinal microbiota and gastrointestinal diseases. World J. Gastroenterol. 22(40), 8905–8909 (2016).
Zhang, Y. B. et al. Combined lifestyle factors, incident cancer, and cancer mortality: A systematic review and meta-analysis of prospective cohort studies. Br. J. Cancer 122(7), 1085–1093 (2020).
Clark, M. A., Springmann, M., Hill, J. & Tilman, D. Multiple health and environmental impacts of foods. Proc. Natl. Acad. Sci. 116(46), 23357–23362 (2019).
Bender, A. E. & Zia, M. Meat quality and protein quality. Int. J. Food Sci. Technol. 11(5), 495–498 (1976).
Dehnhard, M., Bernal-Barragan, H. & Claus, R. Rapid and accurate high-performance liquid chromatographic method for the determination of 3-methylindole (skatole) in faeces of various species. J. Chromatogr. B 566, 101–107 (1991).
Madsen, L., Myrmel, L. S., Fjære, E., Liaset, B. & Kristiansen, K. Links between dietary protein sources, the gut microbiota, and obesity. Front. Physiol. 8, 1047 (2017).
Ruangyuttikarn, W., Appleton, M. L. & Yost, G. S. Metabolism of 3-methylindole in human tissues. Drug Metab. Dispos. 19, 977–984 (1991).
Suyama, Y. & Hirayama, C. Serum indole and skatole in patients with various liver diseases. Clin. Chim. Acta 176, 203–206 (1988).
Herter, C. A. The occurrence of skatole in the human intestine. J. Biol. Chem. 4, 101–109 (1908).
Riggio, O. et al. Peripheral and splanchnic indole and oxindole levels in cirrhotic patients: A study on the pathophysiology of hepatic encephalopathy. Am. J. Gastroenterol. 105, 1374–1381 (2010).
Morrison, D. J. & Preston, T. Formation of short chain fatty acids by the gut microbiota and their impact on human metabolism. Gut Microbes 7, 189–200 (2016).
Claus, R., Losel, D., Lacorn, M., Mentschel, J. & Schenkel, H. Effects of butyrate on apoptosis in the pig colon and its consequences for skatole formation and tissue accumulation. J. Anim. Sci. 81, 239–248 (2003).
Shi, J. et al. High meat protein high fat diet induced the dysbiosis of gut microbiota and tryptophan metabolism in Wistar rats. J. Agric. Food Chem. 68(23), 6333–6346 (2020).
Liu, D. et al. Indoleacetate decarboxylase is a glycyl radical enzyme catalysing the formation of malodorant skatole. Nat. Commun. 9(1), 4224 (2018).
Pereira, P. M. C. C. & Vicente, A. F. R. B. Meat nutritional composition and nutritive role in the human diet. Meat Sci. 93(3), 586–592 (2013).
Hou, Y. & Wu, G. Nutritionally essential amino acids. Adv. Nutr. 9(6), 849–851 (2018).
Tang, Z. Z. et al. Multi-omic analysis of the microbiome and metabolome in healthy subjects reveals microbiome-dependent relationships between diet and metabolites. Front. Genet. 10, 454 (2019).
Canfora, E. E., Meex, R., Venema, K. & Blaak, E. E. Gut microbial metabolites in obesity, NAFLD and T2DM. Nat. Rev. Endocrinol. 15(5), 261–273 (2019).
Ríos-Covián, D. et al. Intestinal short chain fatty acids and their link with diet and human health. Front. Microbiol. 7, 185 (2016).
Venegas, D. et al. Short chain fatty acids (SCFAs)-mediated gut epithelial and immune regulation and its relevance for inflammatory bowel diseases. Front. Immunol. 10, 277 (2019).
Roager, H. M. & Licht, T. R. Microbial tryptophan catabolites in health and disease. Nat. Commun. 9(1), 3294 (2018).
Hendrikx, T. & Schnabl, B. Indoles: Metabolites produced by intestinal bacteria capable of controlling liver disease manifestation. J. Intern. Med. 286(1), 32–40 (2019).
Gao, J. et al. Impact of the gut microbiota on intestinal immunity mediated by tryptophan metabolism. Front. Cell Infect. Microbiol. 8, 13 (2018).
Zelante, T. et al. Tryptophan catabolites from microbiota engage aryl hydrocarbon receptor and balance mucosal reactivity via interleukin-22. Immunity 39(2), 372–385 (2013).
Morris, G. et al. The role of the microbial metabolites including tryptophan catabolites and short chain fatty acids in the pathophysiology of immune-inflammatory and neuroimmune disease. Mol. Neurobiol. 54(6), 4432–4451 (2017).
Islam, J. et al. Dietary tryptophan alleviates dextran sodium sulfate-induced colitis through aryl hydrocarbon receptor in mice. J. Nutr. Biochem. 42, 43–50 (2017).
Kurata, K. et al. Skatole regulates intestinal epithelial cellular functions through activating aryl hydrocarbon receptors and p38. Biochem. Biophys. Res. Commun. 510(4), 649–655 (2019).
De Filippo, C. et al. Impact of diet in shaping gut microbiota revealed by a comparative study in children from Europe and rural Africa. Proc. Natl. Acad. Sci. 107(33), 14691–14696 (2010).
Pareek, S. et al. Comparison of Japanese and Indian intestinal microbiota shows diet-dependent interaction between bacteria and fungi. Npj Biofilms Microbiomes 5(1), 37 (2019).
Xie, Y. T., Zhou, G. H., Wang, C., Xu, X. L. & Li, C. B. Specific microbiota dynamically regulate the bidirectional gut−brain axis communications in mice fed meat protein diets. J. Agric. Food Chem. 67, 1003–1017 (2019).
Zhu, Y. et al. Beef, chicken, and soy proteins in diets induce different gut microbiota and metabolites in rats. Front. Microbiol. 8, 1395 (2017).
Acknowledgements
This work was supported by the Ministry of Science and Technology of China (10000 Talent).
Author information
Authors and Affiliations
Contributions
C.L. designed research; J.S., D.Z., F.Z., C.W., and G.Z. conducted research; J.S. analyzed data; J.S., C.L., and G.Z. wrote the paper; C.L. had primary responsibility for final content. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Shi, J., Zhao, D., Zhao, F. et al. Chicken-eaters and pork-eaters have different gut microbiota and tryptophan metabolites. Sci Rep 11, 11934 (2021). https://doi.org/10.1038/s41598-021-91429-3
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-021-91429-3
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
