
Abstract
Ischyromyids are a group of large rodents with the earliest fossil record known from the late Paleocene (Clarkforkian) of North America; they are considered the earliest fossil representatives of Rodentia of modern aspect. Ischyromyids dominated early Paleogene small-mammal assemblages of North America and in the latest Paleocene migrated to western Europe and to Asia; in the latter they survived only to the beginning of the late Eocene, but were never abundant. Here we describe for the first time the calcanei of ischyromyids from the early middle Eocene of the Erlian Basin in Nei Mongol, northern China. These calcanei document the existence of three species. The morphology of the studied tarsal bones overall suggests ambulatory locomotion for these animals (‘slow cursors’), similar to that of the coypu and porcupines, but one form shows more marked cursorial capabilities. These differences show that Chinese ischyromyids, although rare, had attained greater taxonomic diversity by the middle Eocene in Nei Mongol than estimated from dental remains. We also address the question of the morphological and ecological divergence of these ischyromyids in relation to their North American counterparts, as well as the issue of a direct dispersal route from North America to Asia in the early Eocene.
Similar content being viewed by others


Introduction
Ischyromyidae are the group that includes the earliest and most basal rodents, sometimes regarded as a stem rodent group1,2 or grouped with ctenodactyloids (Cocomys) as representing the first true rodents and a sister clade to Alagomyidae3; the latter were removed from rodents to Rodentiaformes by Meng and Wyss2. Whether ischyromyids are more derived than basal ctenodactyloids has been a matter of dispute4,5,6,7,8. Nevertheless, the appearance of ischyromyids predates that of the earliest ctenodactyloids by ca. one million years, which still constitutes the earliest fossil record of rodents3,6,9,10.
As currently presumed, ischyromyids originated in North America. Their earliest fossil record consists of Acritoparamys atavus (Bear Creek, Montana, USA) and Paramys adamus (Big Multi Quarry, Wyoming, USA), both dated at the latest Paleocene, early Clarkforkian North American Land Mammal Age (NALMA; see6,11). The group rapidly became diversified and thrived in North America during the Eocene6,12. Ischyromyids quickly migrated to Europe, where they are known at least from the beginning of the Ypresian (earliest Eocene; Dormaal [MP 7] faunal level; see13,14) and possibly even from the latest Paleocene (MP 6b faunal level15). However, they were much less diverse and abundant in Europe than in North America. Ischyromyids appeared in Asia in the earliest Eocene16; thus, the Asian fossil record of this group17,18,19 is only marginally younger than the European one. Also, they were never species-rich or common faunal elements on this continent17,19,20,21,22. Recently, Mein and Pickford23 reported the first record of an ischyromyid from the middle Eocene of Namibia. If the family attribution of Namaparamys is correct, Africa would be the final continent colonized by this group.
Here we present the postcranial material of ischyromyids from Asia for the first time. Our findings come from the middle Eocene localities in the Erlian Basin (Fig. 1), which is one of the classic Paleogene fossiliferous areas in Nei Mongol, northern China10,19,24,25,26,27,28. Despite the overall diversity and abundance of rodents during the Eocene in Nei Mongol29, ischyromyids were always very rare there (except for Asiomys;19,29). Our study of the calcanei of ischyromyids from the Erlian Basin shows two or three different taxa (either genera or species). This diversity points to a greater species richness of this group in northern China during the middle Eocene than was previously suggested by the dental material19,21, comparable to the record from southern China (Shanghuang;30). Furthermore, differences in the calcaneal structure between Asian and North American taxa imply a different paleoecology for Asian taxa with respect to their North American predecessors and a different composition of Eocene rodent paleocommunities on both continents.

Stratigraphic and geographic distribution of Ischyromyidae in China. (A) map of ischyromyid findings in China; yellow denotes Irdinmanhan findings in a province, green denotes older (?Bumbanian) findings; (B) panoramic view of Huheboerhe area; (C) detailed map of studied sites in the Erlian Basin; (D) stratigraphy of studied localities in the Erlian Basin, Nei Mongol, China; Irdinmanhan strata marked in yellow. (Maps and stratigraphic section created in Corel Draw X4 (v.14.0.0.567) by Łucja Fostowicz-Frelik; photograph taken by Łucja Fostowicz-Frelik).
Institutional abbreviations AMNH, American Museum of Natural History, New York, NY, USA; ISEZ, Institute of Systematics and Evolution of Animals, Polish Academy of Sciences, Cracow, Poland; IVPP, Institute of Vertebrate Paleontology and Paleoanthropology, Chinese Academy of Sciences, Beijing, China; ZMCAS, Zoological Museum of the Chinese Academy of Sciences, Beijing, China.
Results
Systematic paleontology
Order Rodentia Bowdich, 182131
Family Ischyromyidae Alston, 187632
Genus Asiomys Qi, 198733
Asiomys dawsoni Qi, 198733
Figure 3A–E
Material. Fragment of right calcaneus (IVPP V24417), early Middle Eocene, Huheboerhe, Irdin Manha Formation, Erlian Basin, China.
Description. The bone is damaged and most probably that of a juvenile as it shows loss of the tissue in the extremities of the bone such as the calcaneal tuber and calcaneal eminence, which are usually less calcified in juveniles. The bone is relatively large (Table 1), with an elongated calcaneal tuber and a relatively short body (Fig. 3A–D). The sustentaculum tali is partly damaged; it has a subcircular articulation facet, which was probably more extended craniocaudally than mediolaterally. The caudal margin of the sustentaculum tali is inclined cranially, similar to the condition seen in species A and more than in species B (Fig. 3A). The sustentacular facet overlaps about one-half of the craniocaudal reach of the ectal facet. The groove for the ‘spring ligament’ (sensu Szalay and Decker34), which runs along the medial edge of the sustentaculum tali, is poorly pronounced. Likewise, the calcaneal groove for the tendon of the flexor fibularis muscle is shallow and poorly marked, most probably due to poor preservation. The ectal facet is relatively wide and similarly shaped as in species B (below). The peroneal process is completely damaged.

Linear measurements of the calcaneus. Abbreviations: AEW, ectal facet anterior width; BL, calcaneal body length; BW, calcaneal body width; CCL, calcaneocuboid facet length; CCW, calcaneocuboid facet width; CL, calcaneus length; CMT, calcaneus maximum thickness; CW, calcaneal width; EL, ectal facet length; TEW, ectal facet total width; TL, tuber calcanei length; TT, tuber calcanei thickness; TW, tuber calcanei width; TWM, tuber calcanei width in mid-length. (Figure created in Corel Draw X4 (v.14.0.0.567) by Łucja Fostowicz-Frelik).

Ischyromyid calcanei from the early middle Eocene of the Erlian Basin, Nei Mongol, China. (A–E), Asiomys dawsoni (IVPP V24417), right calcaneus, juvenile?; (F–K), species A (IVPP V24416), right calcaneus, adult; (L–Q), species B (IVPP V24418), right calcaneus, adult. In: A, F and L, dorsal; B, G and M, medial; C, H and N, lateral; D, I and O plantar; J and P caudal; E, K and Q, cranial views. Explanatory line drawings (right side) show important morphological features. Note sustentacular facet marked pale yellow. Scale bar equals 10 mm. (Photographs taken by Łucja Fostowicz-Frelik; drawings created in Corel Draw X4 (v.14.0.0.567) by Łucja Fostowicz-Frelik).
The calcaneal tuber is strongly compressed, but it resembles in shape those of species A and B. A long groove for the calcaneofibular ligament is impressed on its lateral side.
The anterior plantar tubercle is large and swollen, similar to that in species A, and touches the brim of the calcaneocuboid surface. The latter, only slightly damaged laterally, is round in outline, without a distinct pit, and inclined about 20–30°.
Systematic remark: The fossil was associated with Asiomys dentition found in the same spot. We attribute specimen IVPP V24417 to Asiomys dawsoni, based on this fact and its distinctive size (Asiomys being the largest rodent in the assemblage). Asiomys is the only ischyromyid rodent known from the basal strata of the Irdin Manha Formation of Huheboerhe.
Genus indet.
Species A
Figure 3F–K
Material. Right calcaneus (IVPP V24416), early Middle Eocene, Irdin Manha Escarpment, Irdin Manha Formation, Erlian Basin, China.
Description. The right almost complete calcaneus of an adult specimen is relatively large (Table 1), comparable in length to the calcaneus of a coypu (Myocastor coypus) or Asiatic brush-tailed porcupine (Atherurus macrourus). The bone has a characteristically elongated calcaneal tuber and rather short body (Fig. 3F–I). The calcaneal tuber is quite slender in comparison with the structure found in the coypu and porcupines. The shape of the bone resembles most closely the calcaneus of Paramys wortmani (see35: Fig. 12B), although in Paramys the calcaneal tuber is more compressed mediolaterally.
The sustentaculum tali is large and eminent, reaching far medially and tapering, although its medial end forms a blunt edge parallel to the long axis of the bone. This medial edge also bears a well-marked but not deep groove of the calcaneonavicular (or ‘spring’) ligament (Fig. 3G). The sustentacular facet (facies articularis talaris media in Fostowicz-Frelik36: Fig. 12B2) is round, with only slight anteroposterior compression. It occupies almost the whole dorsal surface of the sustentaculum, encroaching slightly onto the calcaneal body. In that it differs from Notoparamys and Paramys wortmani, which both have a much more medially placed sustentacular facet, which does not encroach on the calcaneal body. The range of the sustentacular facet overlaps less than one-third of the ectal facet (posterior facies articularis talaris in Fostowicz-Frelik36: Fig. 12B2) on its anterior and medial sides. The calcaneal eminence is slightly longer than that in Marmota and Sciurus, in proportions closer to that of porcupines and of similar size as in Paramys wortmani. The ectal facet is wide, long, and has a distinctly helical course, even more strongly marked than in North American ischyromyids (see Rose and Chinnery35: Fig. 12A). It is, however, inclined more strongly mediolaterally than in Notoparamys and Paramys, and faces strongly medially. On the dorsal side of the calcaneal eminence, posterolateral to the ectal facet, there is a flattened rough area (finely pitted), marking the place of attachment of the lateral collateral ligaments binding the distal fibula and the astragalus with the calcaneus and stabilizing the astragalocalcaneal joint.
A calcaneal body is short and stocky with poorly marked tendon ridges at the dorsal surface. A large peroneal process is partly damaged at its lateral margin. The process is placed closer to the cuboid surface than the sustentaculum tali. The position of the sustentaculum tali and the proportions of the calcaneal body of specimen IVPP V24416 resemble rather closely the calcaneus of Paramys wortmani (see35).
The calcaneal tuber is not ‘pinched’ at its dorsal side but moderately compressed, thus there is no coracoid ridge posterior to the ectal facet. At the lateral side of the tuber, there is a long groove for the calcaneofibular ligament running askew, towards the dorsal surface of the calcaneal tuber. The groove for the calcaneofibular ligament is more weakly expressed than in the North American paramyines and arboreal sciurids, but similar to that of Marmota.
The caudal surface of the calcaneal tuber is subcircular (only slightly more extended dorsoplantarly than mediolaterally, see Fig. 3 and Table 1). The groove for the calcaneal tendon (= Achilles tendon) is deep and placed asymmetrically at the surface (Fig. 3J). Also, the medial process of the calcaneal tuber is much better developed and extending medially.
The plantar surface of the bone is almost straight with a delicate flexure cranially to a well-developed plantar heel process (Fig. 3G). The anterior plantar tubercle is relatively large, swollen, but shifted medially, towards the sustentaculum tali. It is placed very close to the cuboid surface, almost touching its margin; such location and the medial shifting resembles the condition in some ground squirrels, e.g., Cynomys (see Fostowicz-Frelik et al.8: Fig. 3D–F). The anterior plantar tubercle is also somewhat flattened and inclined medially and forms a well-marked calcaneal groove for the tendon of the flexor fibularis muscle.
The calcaneocuboid articular surface is semicircular, slightly wider mediolaterally than long dorsoplantarly, which distinguishes species A from Marmota and paramyines (see35). It is almost transversally positioned, not inclined, as in most of the rodent taxa (coypu and porcupines included), and gently concave; it is also confluent and level with the cuboid pit, forming one round surface at the cranial end of the bone.
Genus indet.
Species B
Figure 3L–Q
Material Right calcaneus (IVPP V24418), early Middle Eocene, Daoteyin Obo, Irdin Manha Formation, Erlian Basin, China.
Description The bone is complete, slightly larger than in species A (Table 1), matching in length the calcaneus of the coypu. Its overall structure is very similar to the calcaneus of Paramys (either P. wortmani or P. taurus, see Rose and Chinnery35: Fig. 12B, C). It has a long and strong calcaneal tuber and a relatively strong but short calcaneal body (Fig. 3L). The tuber is more compressed mediolaterally than in species A; thus, the caudal surface of the tuber is extended more dorsoplantarly than mediolaterally (Fig. 3P). The attachment for the calcaneal tendon forms a rounded concavity at the caudal side of the tuber, and is more horizontally and symmetrically located at the surface than in species A. The lateral surface of the calcaneal tuber bears a marked scar from the calcaneofibular ligament, although the scar is convex, not concave as in species A and in other compared taxa (e.g., Cynomys).
The sustentaculum tali is large and round; it is located relatively close to the calcaneal body, not extending as far medially as in the North American paramyines (see35). It is slightly longer anteroposteriorly and located more caudally (closer to the ectal facet) than in species A. Thus, the sustentacular surface overlaps ca. one-half of the cranial part of the ectal facet. The medial edge of the sustentacular shelf bears a deep groove for the ‘spring ligament’.
The ectal facet is large, equally wide throughout its length, long and helical, although its course is straighter along the proximodistal direction than in species A. The ectal surface faces mediodorsally, with a slightly weaker medial component than in species A. The dorsal surface of the tuber, just caudal to the ectal facet, is not typically ‘pinched’ into a sagittally oriented crest, but it is, nevertheless, more mediolaterally compressed than in the species A, similar to Marmota.
The calcaneal body forms about one-third of the bone length. Its dorsal surface is carved by deep longitudinal marks indicating the position of the extensor digitorum brevis muscle (Fig. 3). A middle-size peroneal process is located cranially at the calcaneal body. It is strong and long anteroposteriorly, reaching almost the edge of the calcaneocuboid surface. Its lateral edge shows a deep groove for the tendon of the peroneus longus muscle, while its dorsal surface forms a groove for the peroneus brevis muscle tendon (Fig. 3). Species B differs from the ground squirrels in the shape and location of the peroneal process, which is less extended laterally in species B than e.g., in marmots, although it is relatively much larger than in the coypu and porcupines.
The anterior plantar tubercle looks less swollen than in species A; it is located at the very margin of the calcaneocuboid surface and as in species A is shifted medially (Fig. 3O, Q). The calcaneocuboid surface is slightly inclined (ca. 25°) anteromedially, which distinguishes the bone from species A, Marmota, and Notoparamys, which all have the calcaneocuboid facet almost transversal and perpendicular to the long axis of the calcaneus. In this respect, the calcaneocuboid surface resembles more closely the calcaneus of Paramys taurus (Rose and Chinnery35: Fig. 12C). The calcaneocuboid surface is almost round, slightly wider mediolaterally, resembling that of species A. A relatively small calcaneal pit (extending only to a half of the anterior plantar tubercle base, see Fig. 3Q), smaller but deeper than in species A, forms a shallow sink at the medial side of the surface, cranially to the sustentaculum tali.
PCA analysis
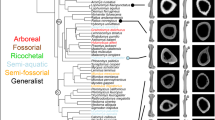
A Principal Component Analysis (PCA) was performed based on 14 measurements of the calcaneus. The analysis included the calcaneal measurements of five ischyromyid species (two described here as species A and B, and three comparative species from North America) and 16 extant large rodent species (Supplementary Table S1). The extant taxa represent six basic types of locomotor adaptations found in rodents: ambulatorial (terrestrial generalists), amphibious (swimming), arboreal (tree climbing), cursorial (four-pedal runners), ricochetal (bipedal jumpers), and semi-fossorial (burrowing).
Principal Components 1 and 2 (PC1 and PC2) represent 87.48% and 5.75% of the variance, respectively, whereas Principal Components 3–4 represent further 4% of the variance (Supplementary Table S2). All the variables are positively correlated with PC1 and their loadings are very balanced (Fig. 4). Thus, it implies that the PC1 represents a proxy for the size of the bone. PC2 is most strongly correlated with the length of the calcaneal body, BL (-0.86) and more weakly correlated with the width of the cuboid facet (CCW) and anterior width of the ectal facet (AEW), 0.31 and 0.21, respectively (Fig. 4). The correlation with the length of the calcaneal body is an especially important factor for estimating an animal’s vertical jumping ability; the species with elongated calcaneal bodies are generally better jumpers (see8,36). The strong negative correlation of the length of the calcaneal body in the second component is illustrated by grouping the species with a strong jumping locomotor repertoire (e.g., squirrels and chinchillas) towards the left side of the plot (Fig. 4). Incidentally, this phenomenon does not concern the calcanei of ricochetal species (see the position of Pedetes versus that of Sciurus and Chinchilla: Fig. 4), where the mechanics of a jump are differently realized, and the stabilisation and relative stiffness of the ankle joint plays the most important role (thus, the calcaneal body and calcaneal tuber are more similar in size).

Principal component analysis of 14 metric parameters of rodent calcanei. The morphospace including paramyid calcanei from Nei Mongol in yellow circle. Lines connecting all data points represent a minimum spanning tree (MST) based on a Euclidean distance matrix. The loadings of the Components 1 and 2 shown at the corresponding axes. Strictly fossil taxa marked in red and pink, extant in black. (Figure created in Corel Draw X4 (v.14.0.0.567) by Łucja Fostowicz-Frelik).
In the plot of PC1 against PC2, ischyromyids do not cluster together. Instead, the PCA morphospace is divided into two (or even three) broad groups of ischyromyid locomotor adaptations: the ambulatorial species and those with more pronounced jumping or cursorial ability. Chinese taxa fall among typically large ambulatorial rodents, such as the coypu (Myocastor) and porcupines (Atherurus and Hystrix). Closest to them there is the North American ischyromyid Quadratomus, which is somewhat shifted towards the cursorial species and can be thus distinguished as differently specialized (more cursorial). Two other North American ischyromyids, Ischyromys and Reithroparamys, are grouped with Chinchilla and Ondatra, respectively, which may imply some jumping and slightly scansorial locomotor adaptations for Ischyromys and those of typical agile generalist species for Reithroparamys.
Although the sample is limited, the results of the PCA analysis point to general differences in the structure of the calcaneus, and thus, locomotor specialisation, between Asian and North American ischyromyid species. Moreover, Asian species seem to differ less from each other than the North American ones do, reflecting the overall greater species diversity and coverage of a wider niche spectrum of the North American ischyromyids.
Functional and paleoecological implications
The studied calcanei add to our knowledge on the functional aspects of locomotion of ischyromyid rodents. Proximal tarsal morphology has been recently used to interpret the locomotor behavior of some extinct rodents (see e.g.,8,37,38,39). In the scheme of locomotor categories of Samuels and Van Valkenburgh40, attributions proposed for early ischyromyids fit into generally terrestrial41, arboreal42 or a mixture of those two35.
A relatively short calcaneal body, widely spread sustentaculum tali, and a large peroneal process observed in most ischyromyid species (including these studied herein) indicate rather poor cursoriality. Instead, their ankle joint structure allows for a large freedom of foot movements in different planes. A medially extended sustentaculum tali together with a long and helically twisted ectal facet indicate a large degree of sliding between the calcaneus and astragalus along their articular facets, which makes possible a great degree of foot torsion resulting in foot eversion and inversion. This effect is further enhanced by an extended calcaneocuboid facet that is gently concave and oriented perpendicularly to the long axis of the calcaneus in species A.
Such adaptations are helpful for both clinging to branches and adjusting to uneven or inclined substrate during climbing. A great degree of freedom of movement may be helpful also during burrowing, when the hind legs are used to push forward loose soil out of a burrow or an animal is forced to maintain a crouched posture, when it digs with its forelegs and head. Nevertheless, as much as the calcaneal structure may suggest some burrowing ability in ischyromyids (see Rose and Chinnery35), the rest of the postcranial skeleton known from the more complete specimens of North American representatives41 does not support fossorial adaptations. In particular, a long tail in the pre-Oligocene North American (see e.g., Paramys or Reithroparamys in Wood41: figs. 8 and 44, respectively) suggests some arboreal adaptations or at least occasional climbing, as such a tail greatly enhances balancing on uneven terrain. In contrast, typically fossorial mammals have reduced tails43.
The overall morphology of dental and mandibular remains16,18 of Asian ischyromyids is similar to that of their North American counterparts16,19. As complete or even partial postcranial skeletons are unknown for the Asian ischyromyids, we can surmise their general locomotor adaptations based on calcaneal morphology which, although not in striking contrast with their North American counterparts, shows some differences.
Overall, the calcaneal morphology of Chinese ischyromyids is closest to that of ground squirrels and especially porcupines (both Atherurus and Hystrix) and the coypu; the similarity to the last one is supported also by the PCA analysis. The calcaneal morphology and proportions may therefore reflect their locomotion behavior as generalized terrestrials, with a somewhat limited ability to climb (a rare but observed behavior in Hystrix) and to dig burrows (as does Atherurus43). A transverse and gently concave calcaneocuboid facet of species A facilitates foot rotation along the long axis, useful on an uneven, rocky terrain or while traversing branches, when an animal needs a flexible foot for a better grip (see Chester et al.44). On the other hand, the lack of both a characteristically bent calcaneal tuber and posteriorly located peroneal process in all ischyromyids (except for Notoparamys, see Rose and Chinnery35) argues against the arboreal adaptations characteristic of tree squirrels.
Discussion
In contrast to the preceding fauna (Arshantan ALMA) of Nei Mongol, where the Glires assemblages were less diverse and dominated by the early ctenodactyloid Tamquammys8,10,19, the early middle Eocene (Irdinmanhan ALMA) witnessed a surge of new Glires taxa including, among others, ischyromyids29. In particular, ctenodactyloids diversified markedly in that interval10,29 and the first cricetids appeared45, along with the increase in lagomorph diversity46,47 and a slight mimotonid revival48,49. Mimotonids were much less abundant during the Arshantan and Irdinmanhan than in the Bumbanian 25, although their taxonomical diversity was higher 48,49.
Since their origin, ischyromyids were relatively large, based on their first molar length, which reached frequently over 4 mm6. Thus, in the Irdinmanhan biocenoses their closest ecological competitors should be other large Glires, such as Gomphos and Mimolagus48, although by then Gomphos was already much less abundant than in the preceding Bumbanian ALMA25,26,50. Among the Irdinmanhan rodents most ctenodactyloids (apart from Yuomys cavioides and Y. weijingensis) and cricetids were small, based on the M1/m1 length much below 2 mm19,45. The lagomorphs were even smaller47; their body mass was estimated at less than 150 g 48. Thus, these herbivores probably were not direct competitors of ischyromyids.
Asian ischyromyids are relatively rare compared to the North American record; in total, ten genera were reported from Asia (Fig. 5). Six taxa come from China16,19,21,30,51, one from India22, two plausible from Kazakhstan20, and one from Pakistan (see Dawson17:101). Thus far, all of them have been known either from isolated teeth or mandible fragments16. The first Chinese ischyromyids Taishanomys changlensis and Acritoparamys? wutui are known from the early Early Eocene of the Wutu Formation, Shandong Province, China16,18. The Wutu Fm. is coeval with the upper Nomogen Formation in Nei Mongol52, and characterized by the presence of Alagomyidae, an important faunal element typical of the Bumbanian and preceeding Gashatan ALMAs2,25,53. Early ingress of ischyromyids into the Erlian Basin in Nei Mongol (Nuhetingboerhe site) took place in the Arshantan19. The group, however, did not become established in this area until the beginning of the Irdinmanhan19. The northward expansion of ischyromyids during the middle Eocene to early Oligocene interval in China mimics the trend observed in other mammalian groups, in particular anagalids, a Paleogene group of primitive Euarchontoglires endemic to Asia54, which also appeared in Nei Mongol not earlier than in the Irdinmanhan49,55.

Temporal ranges of Ischyromyidae. Color of range line denotes subfamily: black, Microparamyinae; pink, Paramyinae; orange, Ischyromyinae; white, Ailuravinae; yellow stripe marks the Irdinmanhan ALMA. Geological and stratigraphic chart based on Woodburne11, Ni et al.52, and Cohen et al.64; classification based on Anderson6 and McKenna and Bell65. For fossils age data, see the text. (Drawing made in Corel Draw X4 (v.14.0.0.567) by Łucja Fostowicz-Frelik).
The affiliations of Chinese ischyromyids are vague, as well as are the exact routes of their dispersal. Taishanomys and Acritoparamys from the Wutu Formation, Anatoparamys from Shanghuang, and Asiomys from Nei Mongol are assigned to Paramyinae16,19,30,33. Eoischyromys from the middle Eocene of the vicinity of Beijing21 is considered Ischyromyinae. Hulgana from the Ulan Gochu Formation, Jhama Obo, Nei Mongol56, once considered an ischyromyine, may be a cylindrodont57. Furthermore, the lack of Ailuravinae in the Chinese19 and Central Asian fossil rodent record20,58 indicates that ischyromyids came to China with a different wave of migration than that which led to colonization of India22. The presence of Ischyromyinae and a greater similarity of Asiomys to North American ischyromyids than to European taxa19 imply that the migration route to northern China may have been directly from North America via the Beringian region, as for tarkadectine primates59.
The overall scarce ischyromyid findings in Asia indicate that the group did not adapt well to Asian environments and may have suffered from competition with small herbivores, either mimotonids or even minute perissodactyls, abundant in China at that time48,60. Also, it seems that early Asian ischyromyids preferred more humid and warm habitats; they were most diverse in the middle Eocene Shanghuang fissure fillings, southern China (five taxa;30), which yielded a rich mammalian fauna suggestive of a treed environment (e.g., bats and primates;61). The demise of ischyromyids in Asia may have been connected with a growing ecological pressure from other rodent groups, especially ctenodactyloids, which continued to dominate the late Eocene of China17,62.
Material and methods
The anatomical terminology (Figs. 2, 3) follows Fostowicz-Frelik36, Ginot et al.37, and Fostowicz-Frelik et al.8. The measurements (Fig. 2, Table 1, Supplementary Information) were taken with a digital caliper (with 0.1 mm accuracy). The comparative material used in this paper includes postcranial material of ischyromyid rodents from the AMNH collection as well as extant rodent taxa (coll. AMNH, ISEZ, IVPP, and ZMCAS; see Supplementary Information) chosen to include the calcanei of large extant taxa representing ambulatorial, semi-fossorial, ricochetal, cursorial, and arboreal locomotor adaptations. PCA analyses on the variance–covariance matrix were performed with PAST v. 2.1763.
Geological settings
The ankle bone material studied here comes from three localities (Fig. 1) of the Erenhot area in the Erlian Basin28, the classical area of the Central Asiatic Expeditions since 192327,28,60. These are: Irdin Manha Escarpment, Daoteyin Obo (= Overnight Camp; see Wang et al.28: Fig. 1), and Huheboerhe (= ‘10 miles Southwest of Camp Margetts’). The stratigraphic range of these localities spans generally the upper Eocene (Bumbanian Asian Land Mammal Age [ALMA]) strata assigned to the Nomogen Formation through the beds of the lower middle Eocene belonging to the Irdinmanhan Formation26,27,28. The Huheboerhe outcrop covers the whole mentioned sequence, while the Irdin Manha Escarpment and Daoteyin Obo sections include only the Arshantan–Irdinmanhan beds, but the thickness of the Arshantan strata varies (Fig. 1). The material studied herein comes exclusively from the lower Irdinmanhan (IM-1) part of the section (Fig. 1). The specimens were collected, mostly through surface screening, by the IVPP field parties during 2004–2012 seasons. GPS data (geographic coordinates and the height above sea level) were recovered for each finding.
Data availability
All data generated or analysed during this study are included in this published article (and its Supplementary Information files).
References
Wyss, A. R. & Meng, J. Application of phylogenetic taxonomy to poorly resolved crown clades: a stem-modified node-based definition of Rodentia. Syst. Biol. 45, 559–568 (1996).
Meng, J. & Wyss, A. R. The morphology of Tribosphenomys (Rodentiaformes, Mammalia): phylogenetic implications for basal Glires. J. Mammal. Evol. 8, 1–71 (2001).
Dawson, M. R. & Beard, K. C. New late Paleocene rodents (Mammalia) from Big Multi Quarry, Washakie Basin, Wyoming. Palaeovertebrata 25, 301–321 (1996).
Marivaux, L., Vianey-Liaud, M. & Jaeger, J. J. High-level phylogeny of early tertiary rodents: dental evidence. Zool. J. Linn. Soc. 142, 105–134 (2004).
Wible, J. R., Wang, Y. Q., Li, C. K. & Dawson, M. R. Cranial anatomy and relationships of a new ctenodactyloid (Mammalia, Rodentia) from the early Eocene of Hubei Province, China. Ann. Carnegie Mus. 74, 91–150 (2005).
Anderson, D. Ischyromyidae in Evolution of Tertiary Mammals in North America. Volume 2: Small Mammals, Xenarthrans, and Marine Mammals (eds. Janis, C. M., Gunnell, G. F. & Uhen, M. D.) 311–25 (Cambridge University Press, 2008).
Antoine, P. O. et al. Middle Eocene rodents from Peruvian Amazonia reveal the pattern and timing of caviomorph origins and biogeography. Proc. R. Soc. Lond. B 279, 1319–1326 (2012).
Fostowicz-Frelik, Ł, Li, Q. & Ni, X. Oldest ctenodactyloid tarsals from the Eocene of China and evolution of locomotor adaptations in early rodents. BMC Evol. Biol. 18, e150 (2018).
Meng, J. & Wyss, A. R. Glires in The Rise of Placental Mammals; Origins, Timing, and Relationships of the Major Extant Clades (eds. Rose, K. D. & Archibald, J. D.) 145–158 (Johns Hopkins University Press, 2005).
Li, Q. & Meng, J. New ctenodactyloid rodents from the Erlian Basin, Nei Mongol, China and the phylogenic relationships of Eocene Asian ctenodactyloids. Am. Mus. Novit. 3828, 1–58 (2015).
Woodburne, M. O. (ed.) Late Cretaceous and Cenozoic Mammals of North America. Biostratigraphy and Geochronology (Columbia University Press, 2004).
Ivy, L. D. Systematics of the late Paleocene and early Eocene Rodentia (Mammalia) from the Clarks Fork Basin, Wyoming. Contrib. Mus. Paleontol. Univ. Mich. 28, 21–70 (1990).
Michaux, J. Les paramyidae (Rodentia) de l’Eocène inférieur du bassin de Paris. Palaeovertebrata 1, 135–193 (1968).
Escarguel, G. Les rongeurs de l’Eocène inférieur et moyen d’Europe occidentale. Systématique, phylogénie, biochronologie et paléobiogéographie des niveaux-repères MP 7 à MP 14. Palaeovertebrata 28, 89–351 (1999).
Smith, T. et al. First Clarkforkian equivalent land mammal age in the latest Paleocene basal Sparnacian facies of Europe: fauna, flora, paleoenvironment and (bio)stratigraphy. PLoS ONE 9, e86229 (2014).
Tong, Y. & Dawson, M. R. Early Eocene rodents (Mammalia) from Shandong Province, People’s Republic of China. Ann. Carnegie Mus. 64, 51–63 (1995).
Dawson, M. R. Paleogene rodents of Eurasia. Deinsea 10, 97–126 (2003).
Tong, Y. & Wang, J. Fossil mammals from the early Eocene Wutu formation of Shandong Province. Palaeontol. Sin. New Ser. C 192, 1–195 (2006).
Li, Q. & Meng, J. Eocene ischyromyids (Rodentia, Mammalia) from the Erlian Basin, Nei Mongol, China. Vert. PalAsiat. 51, 289–304 (2013).
Shevyreva, N. S. New early Eocene rodents from the Zaysan Basin in The Flora and Fauna of the Zaysan Basin (ed. Gabunia, L. K.) 77–114 (Metsniyereba Press, 1984).
Wang, B. Y., Zhai, R. & Dawson, M. R. Discovery of Ischyromyinae (Rodentia: Mammalia) from the Middle Eocene of North China. Vert. PalAsiat. 36(1), 1–12 (1998).
Rana, R. S. et al. An ailuravine rodent from the lower Eocene Cambay formation at Vastan, western India, and its palaeobiogeographic implications. Acta Palaeontol. Pol. 53, 1–14 (2008).
Mein, P. & Pickford, M. Reithroparamyine rodent from the Eocene of Namibia. Commun. Geol. Sur. Namibia 18, 38–47 (2018).
Russell, D. E. & Zhai, R. The Paleogene of Asia: mammals and stratigraphy. Mém. Mus. Natl. Hist. Nat. Série C Sci. Terre 52, 1–490 (1987).
Meng, J. et al. Gomphos elkema (Glires, Mammalia) from the Erlian Basin: evidence for the Early Tertiary Bumbanian Land Mammal Age in Nei-Mongol, China. Am. Mus. Novit. 3425, 1–24 (2004).
Meng, J. et al. New stratigraphic data from the Erlian Basin: implications for the division, correlation, and definition of Paleogene lithological units in Nei Mongol (Inner Mongolia). Am. Mus. Novit. 3570, 1–31 (2007).
Wang, Y. Q. et al. Early Paleogene stratigraphic sequences, mammalian evolution and its response to environmental changes in Erlian Basin, Inner Mongolia, China. Sci. China Earth Sci. 53, 1918–1926 (2010).
Wang, Y. Q., Meng, J. & Jin, X. Comments on Paleogene localities and stratigraphy in the Erlian Basin, Nei Mongol, China. Vert. PalAsiat. 50, 181–203 (2012).
Li, Q. Eocene fossil rodent assemblages from the Erlian Basin (Inner Mongolia, China): biochronological implications. Palaeoworld 25, 95–103 (2016).
Dawson, M. R. & Wang, B. Y. Middle Eocene Ischyromyidae (Mammalia: Rodentia) from the Shanghuang fissures, Southeastern China. Ann. Carnegie Mus. 70(3), 221–230 (2001).
Bowdich, T. E. An Analysis of Natural Classifications of Mammalia for the Use of Students and Travelers (J. Smith, 1821).
Alston, E. R. On the classification of the order Glires. J. Zool. 44, 61–98 (1876).
Qi, T. The middle Eocene Arshanto fauna (Mammalia) of Inner Mongolia. Ann. Carnegie Mus. 56, 1–73 (1987).
Szalay, F. S. & Decker, R. L. Origins, evolution, and function of the tarsus in late Cretaceous Eutheria and Paleocene Primates in Primate Locomotion (ed. Jenkins F. A.) 223–259 (Academic Press, 1974).
Rose, K. D. & Chinnery, B. J. The postcranial skeleton of early rodents. Bull. Carnegie Mus. Nat. Hist. 36, 211–244 (2004).
Fostowicz-Frelik, Ł. The hind limb skeleton and cursorial adaptations of the Plio-Pleistocene rabbit Hypolagus beremendensis. Acta Palaeontol. Pol. 52, 447–476 (2007).
Ginot, S., Hautier, L., Marivaux, L. & Vianey-Liaud, M. Ecomorphological analysis of the astragalo-calcaneal complex in rodents and inferences of locomotor behaviours in extinct rodent species. PeerJ 4, e2393 (2016).
Candela, A. M., Muñoz, N. A. & García-Esponda, C. M. The tarsal-metatarsal complex of caviomorph rodents: anatomy and functional-adaptive analysis. J. Morphol. 278, 828–847 (2017).
Boivin, M. et al. Tarsal morphology and locomotor adaptation of some late middle Eocene caviomorph rodents from Peruvian Amazonia reveal early ecological diversity. J. Vert. Paleontol. 38, e1555164 (2018).
Samuels, J. X. & Van Valkenburgh, B. Skeletal indicators of locomotor adaptations in living and extinct rodents. J. Morphol. 269, 1387–1411 (2008).
Wood, A. E. The early Tertiary rodents of the family Paramyidae. Trans. Am. Phil. Soc. 52(1), 1–261 (1962).
Szalay, F. S. Rodent and lagomorph morphotype adaptations, origins, and relationships: some postcranial attributes analyzed in Evolutionary Relationships among Rodents. A Multidisciplinary Analysis (eds. Luckett, W. P. & Hartenberger, J. L.) 83–132 (Plenum Press, 1985).
Wilson, D. E. et al. (eds) Handbook of the Mammals of the World: Lagomorphs and Rodents I (Lynx Edicions, 2016).
Chester, S. G. B., Bloch, J. I., Boyer, D. M. & Clemens, W. A. Oldest known euarchontan tarsals and affinities of Paleocene Purgatorius to Primates. Proc. Natl. Acad. Sci. USA. 112, 1487–1492 (2015).
Li, Q. Middle Eocene cricetids (Rodentia, Mammalia) from the Erlian Basin, Nei Mongol, China. Vert. PalAsiat. 50, 237–244 (2012).
Fostowicz-Frelik, Ł & Li, Q. A new genus of stem lagomorphs (Mammalia: Glires) from the middle Eocene of the Erlian Basin Nei Mongol, China. Acta Zool. Cracov. 57, 29–42 (2014).
Fostowicz-Frelik, Ł, Li, C. K., Li, Q., Meng, J. & Wang, Y. Q. Strenulagus (Mammalia: Lagomorpha) from the middle Eocene Irdin Manha formation of the Erlian Basin, Nei Mongol, China. Acta Geol. Sin. Engl. 89, 12–26 (2015).
Fostowicz-Frelik, Ł, Li, C. K., Mao, F. Y., Meng, J. & Wang, Y. Q. A large mimotonid from the middle Eocene of China sheds light on the evolution of lagomorphs and their kin. Sci. Rep. 5, e9394 (2015).
Li, Q., Wang, Y. Q. & Fostowicz-Frelik, Ł. Small mammal fauna from Wulanhuxiu (Nei Mongol, China) implies the Irdinmanhan-Sharamurunian (Eocene) faunal turnover. Acta Palaeontol. Pol. 61, 759–776 (2016).
Dashzeveg, D. & Russell, D. E. Palaeocene and Eocene Mixodontia (Glires) of Mongolia and China. Palaeontology 31, 129–164 (1988).
Tong, Y. Middle Eocene small mammals from Liguanqiao Basin of Henan Province and Yuanqu Basin of Shanxi Province, Central China. Palaeontol. Sin. New Ser. C 26, 1–256 (1997).
Ni, X. et al. Paleogene mammalian fauna exchanges and the paleogeographic pattern in Asia. Sci. China Earth Sci. 63, 202–211 (2020).
Dashzeveg, D. New trends in adaptive radiation of Early Tertiary rodents (Rodentia, Mammalia). Acta Zool. Cracov. 33, 11–35 (1990).
López-Torres, S. & Fostowicz-Frelik, Ł. A new Eocene anagalid (Mammalia: Euarchontoglires) from Mongolia and its implications for the group’s phylogeny and dispersal. Sci. Rep. 8, e13955 (2018).
Fostowicz-Frelik, Ł, Li, Q. & Saha, A. A gliriform tooth from the Eocene of the Erlian Basin (Nei Mongol, China) and the premolar morphology of anagalidan mammals at a crossroads. Diversity 12, 420 (2020).
Dawson, M. R. Oligocene rodents (Mammalia) from East Mesa, Inner Mongolia. Am. Mus. Novit. 2324, 1–12 (1968).
Flynn, L. J., Jacobs, L. L. & Cheema, I. U. Baluchimyinae, a new ctenodactyloid rodent subfamily from the Miocene of Baluchistan. Am. Mus. Novit. 2841, 1–58 (1986).
Averianov, A. O. Early Eocene Rodentia of Kyrgyzstan. Bull. Mus. Natl. Hist. Nat. Sec. C 18, 629–662 (1996).
Ni, X. et al. A new tarkadectine primate from the Eocene of Inner Mongolia, China: phylogenetic and biogeographic implications. Proc. R. Soc. Lond. B 277, 247–256 (2010).
Bai, B. et al. Biostratigraphy and diversity of Paleogene perissodactyls from the Erlian Basin of Inner Mongolia, China. Am. Mus. Novit. 3914, 1–60 (2018).
Qi, T. et al. The Shanghuang mammalian fauna, middle Eocene of Jiangsu: history of discovery and significance. Vert. PalAsiat. 34, 202–214 (1996).
Wang, B. Y. The mid-Tertiary Ctenodactylidae (Rodentia, Mammalia) of eastern and central Asia. Bull. Am. Mus. Nat. Hist. 234, 1–88 (1997).
Hammer, Ø., Harper, D. A. T. & Ryan, P. D. PAST: paleontological statistics software package for education and data analysis. Palaeontol Electron. 4(1), 1–9 (2001).
Cohen, K. M., Finney, S. C., Gibbard, P. L. & Fan, J. X. The ICS International Chronostratigraphic Chart. Episodes 36, 199–204 (2013).
McKenna, M. C. & Bell, S. Classification of Mammals above the Species Level (Columbia University Press, 1997).
Acknowledgements
We thank Jin Meng, Yuanqing Wang, Xijun Ni, Bin Bai, Xun Jin, Ping Li, Chuankui Li, K. C. Beard, D.L. Gebo, Wei Chen, Wei Zhou, Shejie Li, Qiang Cao, Wei Gao, and Chenkai Sun for their assistance during fieldwork. Furthermore, we thank Deyan Ge, Barbara Miękina, Neil Duncan, Eleanor Hoeger, Sara Ketelsen, and Marisa Surovy for the access to comparative osteological collections. We are grateful to Aleksandra Hołda-Michalska for help with Figs. 2, 3, 4. The study was supported by the Grant No. 2015/18/E/NZ8/00637 from the National Science Centre (Cracow, Poland) to ŁF-F, the Strategic Priority Research Program of Chinese Academy of Sciences (XDB26000000) and the National Natural Science Foundation of China Grant No. 41572013 to QL, and an AMNH Kalbfleisch Postdoctoral Research Fellowship to SL-T. The fieldwork was also supported by the Special Fund for Fossil Excavation and Preparation, Chinese Academy of Sciences.
Author information
Authors and Affiliations
Contributions
ŁF-F designed the study and wrote the initial draft of the paper. All authors examined specimens, analyzed and interpreted data, and approved the final version of the manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Fostowicz-Frelik, Ł., López-Torres, S. & Li, Q. Tarsal morphology of ischyromyid rodents from the middle Eocene of China gives an insight into the group’s diversity in Central Asia. Sci Rep 11, 11543 (2021). https://doi.org/10.1038/s41598-021-90796-1
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-021-90796-1
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
