
Abstract
The razor clam Sinonovacula constricta is a commercially important bivalve in Japan. The current distribution of this species in Japan is limited to Ariake Bay, where the fishery stock is declining. It is necessary to understand the genetic population structure of this species in order to restore the fishery stock while preserving the genetic diversity of the clam. Here, we report for the first time the genetic population structure of S. constricta in Ariake Bay, Japan. Paired-end restriction site-associated DNA sequencing (RAD-Seq) analyzed samples of S. constricta collected from seven mudflats located along Ariake Bay. Two different genetic populations exist in Ariake Bay, one inhabiting wild habitats and the other inhabiting the transplanted area of artificial seedlings. Our results suggest that genetic differentiation occurred between these two populations (Fst value = 0.052), and a high level of genetic differentiation is maintained between the two groups. In the future, monitoring the interbreeding status of the two genetically distinct populations and the genetic differentiation within each population is important for conserving the genetic diversity of S. constricta in Japan.
Similar content being viewed by others
Introduction
The technique of artificially producing seedlings and releasing them into fishing grounds is an effective method of aquaculture enhancement. This technique has been implemented with a variety of fishery species (e.g., finfish1,2, flounder3, shellfish4,5,6,7). In artificial seed production, large numbers of seedlings are often produced by a few broodstocks and released into natural seawater, which reduces the genetic diversity of wild populations in the field. Therefore, when seedlings are produced and released into natural seawater, there is a need to understand the genetic structure within the wild population in order to preserve genetic diversity.
The razor clam Sinonovacula constricta (Lamarck, 1818) is an important economic species in East Asian coastal waters8,9,10 and is widely farmed in China10. This species was once distributed from central to western Japan11,12 but is now restricted to the mudflats around Ariake Bay on the west coast of Kyushu in western Japan. In Japan, razor clams are not farmed, and the catch depends on the amount of resources in natural seawaters. According to a report by Ministry of the Environment, Japan13, the annual catch of S. constricta in Ariake Bay often recorded more than 1000 tons per year from the 1900s to the early the 1920s, but it decreased to less than 500 tons in most years from the latter half of the 1920s to the latter half of the 1980s, and no catches have been made since 1994 because of the marked decline in the abundance of this species. In order to restore the S. constricta resources, the Saga Prefectural Ariake Fisheries Research and Development Center has developed seed production techniques14,15, and since 1988, artificial seedlings of S. constricta have been released into the transplanting areas established in natural seawaters16 and have been conserved in transplanting areas as populations to produce subsequent generations. However, the genetic population structure of S. constricta within Ariake Bay has not been examined.
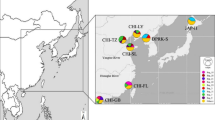
In recent years, technological innovations have made the genome-wide analysis of non-model organisms more widely available, and these techniques are useful for examining the genetic population structure of non-model aquatic organisms17,18,19,20. In the present study, to clarify the genetic population structure of S. constricta in Ariake Bay, we conducted field sampling surveys at 11 mudflats located along the bay in 2019 (Fig. 1) and used restriction-site-associated DNA sequencing (RAD-Seq) to analyze population genetics and phylogenetics. We propose guidelines concerning the optimal seed production and seed release methods for the conservation of the genetic diversity of S. constricta in Japan.

Sampling sites. The dotted line shows the area explored.
Results
Sampling data
We collected 28 razor clams from seven of the 11 mudflats and found no razor clams on the remaining four mudflats (Table 1, Fig. 1). The mean CPUE of the seven mudflats where individuals could be collected was 3.2 (± 3.0 SD) ind. person–1 h–1, and the range of CPUE at each site was 0.3 to 8.5 ind. person–1 h–1. The shell length of the collected individuals was 39.7 to 94.6 mm, implying that they contained specimens from 1 to 5 years old. The mean shell length of individuals collected from the transplanted area was 86.0 (± 4.5 SD) mm for the 3-year-old seedlings produced in 2016 (i.e., sample ID: 2016.1–2016.7) and 92.9 (± 1.1 SD) mm for the 5-year-old seedlings produced in 2014 (i.e., sample ID: 2014.1–2014.5).
RAD-Seq data
We used 30 individuals for RAD-Seq analysis, including 16 individuals collected from wild habitats, 12 individuals collected from the transplanted area of artificial seedlings, and two individuals stored at the Saga Prefectural Ariake Fisheries Research and Development Center (Table 1). In all, 401,109,332 raw reads were generated from the HiSeq X run, and 249,040,001 reads were retained after discarding reads with low quality and adapters. The final dataset included 137,246 loci, with an average of 679.38 bp (SE ± 0.18) of genotyped sites per locus.
Population genetics structure
The genetic relationships among sampling sites were clearly divided into individuals collected from wild habitat and those from the transplanted area of artificial seedlings along the axis of PC1 (Fig. 2a). The PC2 axis represents the differences within each habitat type. In the PCA plot, the coordinates of the transplanted samples were more widely scattered than those of the wild samples were, showing more diverse genetic relationships among transplanted samples. When evaluated by shell length (Fig. 2b), genetic diversity was higher in the 75–90 mm size group than that in the younger age group with smaller shells. The results of admixture analysis also showed a significant genetic difference between the individuals collected from the wild versus those collected from the transplanted areas of artificial seedlings (Fig. 3). The Fst value between populations consisting of individuals collected from the wild and the transplanted areas was 0.052. Owing to the calculation of the pairwise Fst values for each individual within the two populations, there was no significant difference in the mean values between these populations (Fig. 4). The pairwise Fst values within the population were more varied in the transplanted area (from 0.44 to 0.64) than those in the wild habitat (from 0.53 to 0.56). There were no significant differences in shell morphology between these two genetically different populations (Supplementary Fig. S1).

Biplots of principal component analysis. (a) Showing by sampling areas. (b) Showing by shell sizes.

Admixture analysis of S. constricta in the Ariake Bay, Japan.

Pairwise Fst values between individuals in each population.
Phylogenetic tree analysis
ML tree and SVDquartets analysis of RAD-Seq data showed that the phylogenies were highly consistent with each other (Fig. 5a,b). The obtained phylogenetic trees were clearly divided into individuals collected from the wild habitat and those collected from the transplanted area of artificial seedlings, with the exception of one sample (i.e., sample ID: 2016.7).

Phylogenetic trees of S. constricta inferred from RAD-Seq dataset (a) Maximum likelihood phylogeny estimated using RAxML. (b) Species tree inferred using SVDquartets. Numbers at the nodes indicate bootstrap values (1000 replicates).
Discussion
Genetic population structure of S. constricta in the Ariake Bay, Japan
All our analyses of the RAD-Seq data showed that individuals collected from wild habitats were genetically different from those collected from sites where artificial seedlings were transplanted. Only one sample collected from the area transplanted in 2016 (sample ID: 2016.7) showed different results from other seedlings collected in the same area, and belonged to a population consisting of individuals collected from the wild habitat (Figs. 3, 5a,b). Since this individual had a distinctly larger shell length than other 3-year-old seedlings in the same area, and its size was the same as that of 5-year-old seedlings (Table 1), it was likely a wild individual inhabiting the transplanted area prior to the 2016 transplantation. Therefore, we consider that there were two genetically distinct populations in Ariake Bay in 2019.
The reason for the differences between these two populations is the different parental origins of the seedlings. The parental origins can be divided into the Nanaura mudflat and the Kashima mudflat for a number of reasons. Artificial seedlings of S. constricta have been released by the Saga Prefectural Ariake Fisheries Research and Development Center into transplanting areas established in several mudflats along the coast of Ariake Bay. The seedlings collected from the transplanted area in this study (Takori_b and Takori_c) originated from parents at the Nanaura mudflat (hereinafter referred to as Nanaura lineage population), since the parents were collected from the Nanaura mudflat before 2010 and released into the transplanting areas of the Takori mudflat. In contrast, individuals collected from the wild habitat of Kashima, Nanaura, and Takori mudflats are likely to have originated from parents of Kashima mudflat (hereinafter referred to as the Kashima lineage population). In 2017 and 2018, the seedlings produced by parents collected from Kashima mudflat were released into new transplanting areas at Takori and Nanaura mudflats due to declining populations in wild and transplanted areas in these mudflats. Therefore, the 1- to 2-year old individuals collected in wild habitats in 2019 were likely the offspring of the transplanted Kashima seedlings in 2017 and 2018.
Artificial transplantation of aquatic organisms has been reported to alter the geographic genetic structure of wild populations21,22,23,24, including that of S. constricta (in this case, along the Chinese coast)25. Anthropogenic transplantation likely strongly influenced the distribution of the genetic population structure of S. constricta in Japan. Comparison of the genetic distribution of species with a life history similar to that of S. constricta but those that are not exploited for fisheries can enhance our understanding of anthropogenic impacts on the genetic distribution of S. constricta in the Ariake Bay.
Genetic differentiation among and within populations
For artificial seed production and release of seedlings, it is necessary to understand the distribution of populations with different genetic structures in natural seawater. Therefore, it is necessary to select broodstocks and release areas according to the degree of genetic differentiation among and within populations. Fst values are widely used as a measure of the degree of genetic differentiation among and within populations26,27,28. According to Niu et al.25, the Fst value among populations of S. constricta at ten sites along the coast of mainland China is 0.044. The Fst value between the two populations in the present study was 0.052, suggesting that a similar level of genetic differentiation occurred between the two populations in Ariake Bay, Japan.
For these two populations, the genetic differentiation within each population was maintained at a high level, and its variation was greater within the population consisting of individuals collected from the transplanted area (Fig. 4). There are three possible reasons why the variation in the degree of genetic differentiation within populations differed between the two populations. The first is that the degree of variation in genetic differentiation in these populations originally may have been different. Second, the methods of seedling production may lead to greater variation of genetic differentiation within populations in the transplanted area. In seedling production, broodstocks were randomly selected with closely related individuals collected from the same transplanted area or distantly related individuals collected from different transplanted areas. After stimulating the mature individuals to spawn, 20 to 30 individuals were placed in the same tank to produce fertilized eggs. Thus, the degree of genetic differentiation may have been more varied in the transplanted populations than in the wild habitat population. Third, the sample size may have been too small to accurately estimate the population. However, in the current situation in which the number of wild populations is small, it is difficult to verify the validity of a larger sample size.
Guidelines concerning the best seed production and seed release methods
The current abundance of S. constricta is estimated to be very low based on the CPUE results of this study (Table 1). These results suggest that artificial seed production and release of seedlings could be an effective strategy for the recovery of S. constricta abundance in Ariake Bay, Japan. The present study suggested genetic differentiation between the two populations of S. constricta in Ariake Bay, and a high level of genetic differentiation within each population was maintained. Therefore, maintaining the two populations without interbreeding seems to be an appropriate conservation strategy. As of 2019, the Kashima lineage population is widely distributed in wild habitats, whereas the Nanaura lineage population is limited to the Takori mudflats. In order to avoid hybridization with the Kashima lineage population, the release of seedlings of the Nanaura lineage population should continue to be restricted to the mudflats within the current cove, with a transplanting area.
To produce seedlings to slow down the decline in genetic diversity within a population, mathematical models have evaluated five strategies: (I) avoidance of line breeding, (II) selective use of wild-born individuals, (III) line breeding, (IV) avoidance of artificial breeding of males, and (V) selective use of wild-born males29. Among them, strategy II has been considered the most effective29,30. The best way to produce seedlings of the Kashima lineage population would be to follow strategy II; all parental individuals would be randomly collected from wild habitats and previously produced seedlings would not be used as parents. In contrast, the Nanaura lineage population has already been maintained by a similar approach to strategy I; all parental individuals will be randomly collected from transplanted areas without excluding released seedlings. This approach gradually reduces the genetic diversity within the population. Indeed, lower pairwise Fst values were detected in the transplanted population (Fig. 4). Therefore, in the Nanaura lineage population, countermeasures should be taken to increase the number of parents used for seed production or to avoid using the produced seedlings as parents of the next generation.
In conclusion, we detected two genetically distinct populations of S. constricta in the Ariake Bay, Japan. Additionally, our results suggests that genetic differentiation within each population is maintained at a high level. In the future, monitoring the interbreeding status of the two genetically distinct populations and the genetic differentiation within each population is important to enhance the stock of S. constricta in Japan by producing and releasing seedlings, while preserving genetic diversity. As there was no significant difference in the morphology of the shells of these genetically distinct populations (Supplementary Fig. S1), DNA markers may be effective for discriminating between the two populations.
Methods
Sampling and genomic DNA extraction
Sampling of the razor clam S. constricta was conducted at 11 mudflats located along Ariake Bay (Fig. 1). Information on the sampling day and habitat types at each site is shown in Table 1. Habitats are divided into two types: transplanted area of artificial seedlings, and wild habitat. As mentioned above, artificial seedlings of S. constricta have been released into the transplanting areas established in natural seawaters since 1988. Therefore, artificially produced individuals live within the transplanted area, and the wild habitat would be populated by natural individuals and offspring of seedlings from the transplanted areas. The ages of individuals in the transplanted areas were estimated by calculating backwards from the year of release. At each site, razor clams were explored by hand and their abundance was assessed using the catch per unit effort index (CPUE), calculated as the number of clam individuals collected divided by the number of explorers and sampling time (i.e., ind. person–1 h–1). The samples were stored at − 80 °C until genomic DNA extraction. In addition to the collected individuals, the oldest seedlings (produced in 2005, n = 2) stored at the Saga Prefectural Ariake Fisheries Research and Development Center, were included in the RAD-Seq analysis for comparison with seedlings produced in the past. Genomic DNA was extracted from finely chopped foot tissue of the specimens using DNAs-ici!-F (RIZO, Tsukuba, Japan), and the concentration of the DNA was assessed using a Qubit 3.0 Fluorometer (Thermo Fisher Scientific).
RAD-Seq analysis
The preparation for the EcoRI RAD-Seq library has been described previously28, based on the established method31. The library was sequenced with 150 bp paired-end reads in one lane of an Illumina HiSeq X (Illumina, San Diego, CA, USA). The RAD-Seq data were analyzed using the Stacks program (version 2.52)32. The data were quality filtered using the process_radtags program in the Stacks package (-r -c -q,—adapter_1, adapter_2, and adapter_mm2). Filtered sequences were registered in the DRA (DRA accession number: DRA010922). The data were aligned with the reference genome of S. constricta33 (NCBI/DDBJ BioProject number: PRJNA508451) using BWA-MEM (version 0.7.17)34. Aligned data were analyzed using the ref_map.pl script of the Stacks package.
Population genetics analysis
Principal component analysis (PCA) was conducted based on a variant call format (VCF) file using the SNPrelate program (v.1.18.1)35. The VCF file was created by the populations program of the Stacks package (–rm-pcr-duplicates, −R 0.7, –write-single-snp, –min-maf 0.05)36. The admixture analysis was conducted based on the PLINK file using ADMIXTURE (v. 1.3.0)37. The PLINK file was created with the same Stacks populations program that created the VCF file38. The genetic diversity of razor clams among and within populations was assessed using AMOVA Fst values obtained from ‘populations.fst_files’, which were calculated by the Stacks populations program. To visualize the distribution of all data points, pairwise Fst values between individuals in each population are shown in boxplot with beeswarm using R (package: ‘beeswarm’)39.
Phylogenetic tree analysis
Phylogenetic tree analysis was performed using two different approaches: the maximum likelihood (ML) method and SVDquartets analysis. Multiple alignments within the cluster were obtained using the Stacks package populations program mentioned earlier. The phylogenetic tree based on the ML method was constructed using the RAxML program (version 8.2.12)40 (−f = a, −x = 12,345, −p = 12,345, −N (bootstrap value) = 1000, and −m = GTRGAMMA). The SVDquartets analysis based on the coalescent model was computed as implemented in PAUP ver. 4.0a 167 using multiple alignments41,42. The number of bootstrap analyses replicates was 1000. In each analysis, the oldest samples (i.e., sample ID, 2005.1; 2005.2), were used as the root.
References
Fushimi, H. Production of juvenile marine finfish for stock enhancement in Japan. Aquaculture 200, 33–53. https://doi.org/10.1016/S0044-8486(01)00693-7 (2001).
Masuda, R. & Tsukamoto, K. Stock enhancement in Japan: review and perspective. Bull. Mar. Sci. 62, 337–358. https://www.ingentaconnect.com/content/umrsmas/bullmar/1998/00000062/00000002/art00005 (1998).
Sekino, M., Saitoh, K., Yamada, T., Hara, M. & Yamashita, Y. Genetic tagging of released Japanese flounder (Paralichthys olivaceus) based on polymorphic DNA markers. Aquaculture 244, 49–61. https://doi.org/10.1016/j.aquaculture.2004.11.006 (2005).
Arnold, W. S. Bivalve enhancement and restoration strategies in Florida, USA. Hydrobiologia 465, 7–19. https://doi.org/10.1023/A:1014596909319 (2001).
Castell, L. L., Naviti, W. & Nguyen, F. Detectability of cryptic juvenile Trochus niloticus Linnaeus in stock enhancement experiments. Aquaculture 144, 91–101. https://doi.org/10.1016/S0044-8486(96)01320-8 (1996).
McCormick, T. B., Herbinson, K., Mill, T. S. & Altick, J. A review of abalone seeding, possible significance and a new seeding device. Bull. Mar. Sci. 55, 680–693. https://www.ingentaconnect.com/content/umrsmas/bullmar/1994/00000055/f0020002/art00035 (1994).
Zohar, Y. et al. The Chesapeake Bay blue crab (Callinectes sapidus): A multidisciplinary approach to responsible stock replenishment. Rev. Fish. Sci. 16, 24–34. https://doi.org/10.1080/10641260701681623 (2008).
Funge-Smith, S., Briggs, M. & Miao, W. Regional Overview of Fisheries and Aquaculture in Asia and the Pacific 2012 (RAP Publication (FAO), 2012).
Mao, Y. et al. Chapter 4. Bivalve production in China. In Goods and Services of Marine Bivalves (eds Smaal, A. C. et al.) 51–72 (Springer, 2019).
Ran, Z. et al. Fatty acid and sterol changes in razor clam Sinonovacula constricta (Lamarck 1818) reared at different salinities. Aquaculture 473, 493–500. https://doi.org/10.1016/j.aquaculture.2017.03.017 (2017).
Suzuki, T., Inoue, K. & Ozawa, T. Environmental degradation in Ise and Mikawa Bays after 1960’s as viewed from intertidal molluscan community. Boll. Nagoya Univ. Museum 22, 31–64. https://doi.org/10.18999/bulnum.022.04 (2006).
Nakamura, T. et al. Marine reservoir effect deduced from 14C dates on marine shells and terrestrial remains at archeological sites in Japan. Nucl. Instrum. Methods Phys. Res. B 259, 453–459. https://doi.org/10.1016/j.nimb.2007.01.186 (2007).
Ministry of the Environment, Japan. Ariake Sea and Yatsushiro Sea Comprehensive Survey Evaluation Committee Report. https://www.env.go.jp/council/20ari-yatsu/rep061221/all.pdf (2006).
Ito, S., Eguchi, T. & Kawahara, I. Rearing experiment on planktonic larvae of the JackKnife clam, Sinonovacula constricta. Boll. Saga Prefect. Ariake Fish. Res. Dev. Cent. 20, 49–53. https://agriknowledge.affrc.go.jp/RN/2010813185.pdf (2001).
Ohkuma, H., Ymagachi, T., Kawahara, I., Eguchi, T. & Ito, S. A study on the development of techniques for mass production of seeds of jackknife clam, Sinonovacula constricta. Boll. Saga Prefect. Ariake Fish. Res. Dev. Cent. 22, 47–54. https://agriknowledge.affrc.go.jp/RN/2030813210.pdf (2004).
Tsukuda, M. et al. Variation in the Distribution of the JackKnife Clam, Sinonovacula constricta, on the Muddy Tidal flat of Ariake Sound off Saga Prefecture. Boll. Saga Prefect. Ariake Fish. Res. Dev. Cent. 28, 47–49. https://agriknowledge.affrc.go.jp/RN/2010925685.pdf (2017).
Holman, L. E., de la Garcia, S. D., Onoufriou, A., Hillestad, B. & Johnston, I. A. A workflow used to design low density SNP panels for parentage assignment and traceability in aquaculture species and its validation in Atlantic salmon. Aquaculture 476, 59–64. https://doi.org/10.1016/j.aquaculture.2017.04.001 (2017).
Li, Y. H. & Wang, H. P. Advances of genotyping-by-sequencing in fisheries and aquaculture. Rev. Fish Biol. Fish. 27, 535–559. https://doi.org/10.1007/s11160-017-9473-2 (2017).
Robledo, D., Palaiokostas, C., Bargelloni, L., Martínez, P. & Houston, R. Applications of genotyping by sequencing in aquaculture breeding and genetics. Rev. Aquac 10, 670–682. https://doi.org/10.1111/raq.12193 (2018).
You, X., Shan, X. & Shi, Q. Research advances in the genomics and applications for molecular breeding of aquaculture animals. Aquaculture 526, 735357. https://doi.org/10.1016/j.aquaculture.2020.735357 (2020).
Arthington, A. H. Ecological and genetic impacts of introduced and translocated freshwater fishes in Australia. Can. J. Fish. Aquat. Sci. 48, 33–43. https://doi.org/10.1139/f91-302 (1991).
Habtemariam, B. T., Arias, A., García-Vázquez, E. & Borrell, Y. J. Impacts of supplementation aquaculture on the genetic diversity of wild Ruditapes decussatus from northern Spain. Aquacult. Environ. Interact. 6, 241–254. https://doi.org/10.3354/aei00128 (2015).
Williams, S. L. & Orth, R. J. Genetic diversity and structure of natural and transplanted eelgrass populations in the Chesapeake and Chincoteague Bays. Estuaries 21, 118–128. https://doi.org/10.2307/1352551 (1998).
Yamakawa, A. Y. & Imai, H. PCR-RFLP typing reveals a new invasion of Taiwanese Meretrix (Bivalvia: Veneridae) to Japan. Aquat. Invasions 8, 407–415. https://doi.org/10.3391/ai.2013.8.4.04 (2013).
Niu, D. H., Feng, B. B., Liu, D. B., Zhong, Y. M., Shen, H. D. & Li, J. L. Significant genetic differentiation among ten populations of the razor clam Sinonovacula constricta along the coast of china revealed by a microsatellite analysis. Zool. Stud. 51, 406–414. http://zoolstud.sinica.edu.tw/Journals/51.3/406.pdf. (2012).
DeFaveri, J., Shikano, T., Ghani, N. I. A. & Merilä, J. Contrasting population structures in two sympatric fishes in the Baltic Sea basin. Mar. Biol. 159, 1659–1672. https://doi.org/10.1007/s00227-012-1951-4 (2012).
Jeffery, N. W. et al. RAD sequencing reveals genomewide divergence between independent invasions of the European green crab (Carcinus maenas) in the Northwest Atlantic. Ecol. Evol. 7, 2513–2524. https://doi.org/10.1002/ece3.2872 (2017).
Kato, D. et al. Evaluation of the population structure and phylogeography of the Japanese Genji firefly, Luciola cruciata, at the nuclear DNA level using RAD-Seq analysis. Sci. Rep. 10, 1533. https://doi.org/10.1038/s41598-020-58324-9 (2020).
Yokota, M., Harada, Y. & Iizuka, M. Genetic drift in a hatchery and the maintenance of genetic diversity in hatchery-wild systems. Fish Sci. 69, 101–109. https://doi.org/10.1046/j.1444-2906.2003.00593.x (2003).
Harada, Y., Yokota, M. & Iizuka, M. Genetic risk of domestication in artificial fish stocking and its possible reduction. Popul. Ecol. 40, 311–324. https://doi.org/10.1007/BF02763463 (1998).
Etter, P. D., Bassham, S., Hohenlohe, P. A., Johnson, E. A. & Cresko, W. A. SNP discovery and genotyping for evolutionary genetics using RAD sequencing, in: Molecular methods for evolutionary genetics. Methods Mol. Biol. Humana Press 772, 157–178. https://doi.org/10.1007/978-1-61779-228-1_9 (2011).
Rochette, N. C. & Catchen, J. M. Deriving genotypes from RAD-seq short-read data using stacks. Nat. Protoc. 12, 2640–2659. https://doi.org/10.1038/nprot.2017.123 (2017).
Ran, Z. et al. Chromosome-level genome assembly of the razor clam Sinonovacula constricta (Lamarck, 1818). Mol. Ecol. Resour. 19, 1647–1658. https://doi.org/10.1111/1755-0998.13086 (2019).
Li, H. Aligning sequence reads, clone sequences and assembly contigs with BWA-MEM. arXiv preprint arXiv: 1303.3997. https://arxiv.org/abs/1303.3997 (2013).
Zheng, X. et al. A high-performance computing toolset for relatedness and principal component analysis of SNP data. Bioinformatics 28, 3326–3328. https://doi.org/10.1093/bioinformatics/bts606 (2012).
Danecek, P. et al. The variant call format and VCFtools. Bioinformatics 27, 2156–2158. https://doi.org/10.1093/bioinformatics/btr330 (2011).
Alexander, D. H., Novembre, J. & Lange, K. Fast model-based estimation of ancestry in unrelated individuals. Genome Res. 19, 1655–1664. https://pubmed.ncbi.nlm.nih.gov/19648217/ (2009).
Purcell, S. et al. PLINK: A tool set for whole-genome association and population-based linkage analyses. Am. J. Hum. Genet. 81, 559–575. https://doi.org/10.1086/519795 (2007).
Eklund, A. R package beeswarm: the bee swarm plot, an alternative to stripchart. https://cran.r-project.org/web/packages/beeswarm/beeswarm.pdf (2016).
Stamatakis, A. RAxML version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 30, 1312–1313. https://doi.org/10.1093/bioinformatics/btu033 (2014).
Chifman, J. & Kubatko, L. Quartet inference from SNP data under the coalescent model. Bioinformatics 30, 3317–3324. https://doi.org/10.1093/bioinformatics/btu530 (2014).
Swofford, D. L. PAUP*. Phylogenetic analysis using parsimony (*and other methods). Version 4 (Sinauer Associates, 2002).
Acknowledgments
The authors are grateful to the Saga Prefectural Ariake Fisheries Research and Development Center for supporting the field survey and providing samples and information on seedling production. This work was supported by the “Projects for sophistication of production and utilization technology supporting local agriculture and marine industry” from Saga University.
Author information
Authors and Affiliations
Contributions
R.O. designed and wrote the original manuscript draft. R.O. and Y.N. performed the RAD-Seq analysis and bioinformatic analysis. All authors reviewed drafts of the manuscript and approved its final version.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Orita, R., Nagano, Y., Kawamura, Y. et al. Genetic diversity and population structure of razor clam Sinonovacula constricta in Ariake Bay, Japan, revealed using RAD-Seq SNP markers. Sci Rep 11, 7761 (2021). https://doi.org/10.1038/s41598-021-87395-5
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-021-87395-5
This article is cited by
-
Genetic diversity of loquat (Eriobotrya japonica) revealed using RAD-Seq SNP markers
Scientific Reports (2022)
-
Genome-wide SNP analysis reveals the selection signatures of two indigenous buffalo breeds in Sichuan
Conservation Genetics Resources (2022)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.

{kind=link}