
Abstract
Ungulates in alpine ecosystems are constrained by winter harshness through resource limitation and direct mortality from weather extremes. However, little empirical evidence has definitively established how current climate change and other anthropogenic modifications of resource availability affect ungulate winter distribution, especially at their range limits. Here, we used a combination of historical (1997–2002) and contemporary (2012–2015) Eurasian roe deer (Capreolus capreolus) relocation datasets that span changes in snowpack characteristics and two levels of supplemental feeding to compare and forecast probability of space use at the species’ altitudinal range limit. Scarcer snow cover in the contemporary period interacted with the augmented feeding site distribution to increase the elevation of winter range limits, and we predict this trend will continue under climate change. Moreover, roe deer have shifted from historically using feeding sites primarily under deep snow conditions to contemporarily using them under a wider range of snow conditions as their availability has increased. Combined with scarcer snow cover during December, January, and April, this trend has reduced inter-annual variability in space use patterns in these months. These spatial responses to climate- and artificial resource-provisioning shifts evidence the importance of these changing factors in shaping large herbivore spatial distribution and, consequently, ecosystem dynamics.
Similar content being viewed by others

Introduction
In temperate and boreal ecosystems, both the latitudinal and altitudinal distribution of species are often determined by winter conditions1,2. Ungulates and other large herbivores are particularly challenged by the presence of snow, which decreases food accessibility3, affects vegetation phenology4, increases the costs of thermoregulation5, and hampers movement6. While these constraints have driven some morphological adaptation in ungulates living in snow-covered environments7, most temperate ungulates rely on space-use plasticity to meet the challenges posed by snow. Winter space-use tactics, whether migration8 or refuges within winter ranges9, minimize the costs associated with winter conditions and are therefore crucial to ungulate survival in temperate regions.
Ongoing changes in winter conditions stemming from anthropogenic climate change thus carry extensive implications for space use in ungulates10, and are most evident at the latitudinal and altitudinal range limits of species11. These changes, and particularly a reduction in the snowpack12, have been particularly noticeable in the mid-elevations (~ 1000–1700 m, here and hereafter given as above sea level) of mountainous regions in the Northern Hemisphere such as the European Alps12, which form the altitudinal range limit of many species. Moreover, in these alpine regions, climatic changes often co-occur with the alteration of ungulate resource availability through supplemental feeding13,14. This practice consists of the provisioning of abundant plant-based food (grain or maize seeds, grain and fiber pellets, hay, fruit), usually placed in ad hoc feeding sites during winter months. Thus, the natural distribution of food resources, normally scarce in winter, is altered by point-source supplies of highly concentrated food15. In spite of its high ecological relevance, studying the interplay between long-term anthropogenic changes in the physical environment (the snowpack) and food availability (supplemental feeding) in shaping ungulate space use patterns is challenging, due to the gradual nature of these changes and the resulting need to have comparable measurements of space use that have been collected many years apart. Such datasets are rare, as the advent of large herbivore tracking technology is relatively recent16.
In this study, we had the unique opportunity to contrast and compare “historical” (1999–2002) and “contemporary” (2012–2015) space use patterns of Eurasian roe deer (Capreolus capreolus) in a valley at the species’ altitudinal range limit in the Italian Alps (Fig. 1) under ongoing and forecasted changes in winter conditions and food availability (Fig. 2). Roe deer exhibit high ecological plasticity17,18, making them uniquely suited as a bellwether of the ongoing effects of such changes on ungulate winter spatial distribution. Winter harshness (particularly deep snow) severely restricts roe deer movement and thus its spatial distribution, because of their small body mass (18–49 kg) and short legs (50–60 cm)6. Under these constraints, roe deer living in snowy areas adopt movement tactics at multiple spatiotemporal scales. First, roe deer exhibit a seasonal partial migration strategy, with all individuals overwintering in ranges characterized by less extreme snow conditions, and only some migrating to summer ranges (typically at higher elevations)8. Second, within winter ranges, we have previously documented third-order selection19 for shallow snow depth and dense forest canopy20,21. Where supplemental feeding is practiced, the presence of these anthropogenic resources also interacts with winter severity to affect roe deer space use14.

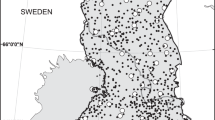
Study area in the Italian Alps, with insets of Italy (top right) and Trentino Province (bottom right). Polygons are shown for the overlapping historical (blue) and contemporary (red) roe deer population (kernel densities 99% based on radio-telemetry relocations). Supplemental feeding sites are indicated for each period as well as the 23 weather stations used to run GEOtop simulations (Supplementary S2).

Snow cover duration for each winter season (Nov–Apr) as a function of altitude in the study area, calculated using GEOtop 2.0 (Supplementary S2). Central continuous lines denote the 50th percentile, while range between 25 and 75th percentiles is shown by the colored areas. RCP scenarios refer to the last decade of the projection period (2046–2065).
In our study area, winter temperatures (Fig. S1.2a) and the variability of snow cover at low-to-intermediate elevations have increased over the last two decades (Fig. 2a), as has the prevalence and distribution of supplemental feeding sites (Fig. 1). We performed an analysis of roe deer space-use data covering this timespan. Specifically, our analysis used (i) a GPS dataset of contemporary roe deer relocations (2012–2015, hereafter “contemporary period”) in a valley in the Italian Alps (Fig. 1); (ii) a VHF dataset of historical roe deer relocations (1999–2002, hereafter “historical period”) in a largely overlapping area22; (iii) a detailed mapping of supplemental feeding site distribution and canopy cover in both periods; and (iv) an advanced surface-process predictive hydrological model of snow depth23, to fit resource selection functions (RSFs) modelling relative probability-of-use across the study area. We used these RSFs to predict and compare (by means of kappa coefficients24 and subtraction maps) the winter (November to April) distribution of roe deer at a high temporal resolution scale (monthly) over the course of the two study periods. Utilizing two climate forecasts of “intermediate” or “severe” thermal loading (the IPCC’s “Representative Concentration Pathways” for 2046–2065, modelling either intermediate or no restraint in emissions; hereafter RCP4.525 and RCP8.526, respectively), we then used the model built with contemporary data to evaluate roe deer habitat selection under predicted future snow depth scenarios. We thus evaluated the following set of hypotheses.
First, we hypothesized that winter distribution would be influenced by both (i) snow depth and (ii) the proximity of supplemental feeding sites20. We further hypothesized (iii) that there would be an interaction between these two factors, with the availability of supplemental feeding affecting the response of roe deer to snow depth. Specifically, we expected that the relative probability of space use would decrease with deep snow (> ~50 cm; Prediction 1) and increase in the proximity of supplemental feeding sites (Prediction 2). Following hypothesis (iii), we expected that as the availability of supplemental feeding sites increased between study periods, the relative probability of space use would concentrate around these sites irrespective of the presence of snow (Prediction 3)—i.e., the attraction of supplemental feeding sites at higher frequency would prevail over the negative effects of snow. Consequently, we also expected that the predicted spatial distribution of roe deer would display higher variability between the historical and the contemporary period than within either period, in response to the inter-decadal changes in snow depth and the substantial increase in supplemental feeding site availability across the study area (Prediction 4). Further, because the year-to-year differences within a given period should depend on snow depth variability (given supplemental feeding sites are constant), we predicted that the increases in winter variability in alpine regions (Fig. 2a) would produce more variability within the contemporary period than in the historical one (Prediction 5a), and even further variability under forecasted warming scenarios (Prediction 5b).
Results
Snow trends
We observed inter-decadal trends in snow cover duration (days with snow depth > 5 cm) and absolute snow depth, both as a function of elevation and of the winter months (November–April). The decade-aggregated average of “snow cover days” from November–April (Fig. 2) showed higher variability in snow cover duration within our study area from 2006 to 2015 (encompassing the contemporary study period) than in the previous decade (from 1996 to 2005, encompassing the historical study period). This pattern was particularly evident at low-to-intermediate elevations (500–1500 m; Fig. 2a). The same aggregated average also predicts that snow cover duration will decrease as a function of continued climate change, again especially at lower elevations (Fig. 2b). However, the comparison of snow depth between historical and contemporary study periods as a function of elevation (Fig. S2.2) showed that in early (November–December) and late (April) winter, snow depth variability decreased in the contemporary period, as a consequence of scarcer snowfall in these months in recent years. In contrast, the variability of snow depth over the central winter months (January–March) was relatively high during both study periods.
Roe deer space use patterns
Contemporary and historical resource selection functions (hereafter “RSFc” and “RSFh”, respectively) confirmed the hypothesized effects of snow and supplemental feeding (hypotheses i and ii) on relative probability of space use by roe deer. In particular, roe deer avoided areas with high snow depth (Prediction 1: βc = − 0.018 ± 0.002/cm; βh = − 0.174 ± 0.010/cm, both p < 0.001) and selected for areas close to supplemental feeding sites (Prediction 2: βc = 351.750 ± 18.540; βh = 529.691 ± 26.650, both p < 0.001, for the reciprocal of distance from the closest feeding site, “1/DistFS”). Further, in accordance with hypothesis (iii), the selection for supplemental feeding sites was stronger at higher snow depths, but less so as the availability of supplemental feeding sites increased between periods, as shown by the different magnitude of the interaction terms (Prediction 3: βc = 4.282 ± 0.752; βh = 140.411 ± 8.432, both p < 0.001) (Fig. 3 and Fig. S3.1). The models also indicated strong positive selection for canopy cover (βc = 0.8033 ± 0.067; βh = 1.217 ± 0.084, where 0 = open, 1 = forested, reference category: open; both p < 0.001), which changed relatively little between study periods.

3D prediction plots of relative probability-of-use of forested habitats (for unforested habitats, see Fig. S3.1) in the two periods (1999–2002: (a); 2012–2015: (b)) covered by the analysis.
The robustness of both RSFs for predicting relative probability-of-use under a variety of conditions was established via spatial validation (each RSF in its own study period) and temporal validation (RSFc hindcasted to the historical period, see “Methods” section). Each validation was performed through a Kendall’s tau (τ) rank correlation analysis (spatial RSFh: τ = 0.80, SD = 0.01, p < 0.05 for all months; spatial RSFc: τ = 0.70, SD = 0.23, all p < 0.05 except November 2014, p = 1; temporal RSFc: τ = 0.76, SD = 0.13; all p < 0.05 except April 1999, p = 0.13; see “Methods” section for explanation of τ and Supplement S5 for further information). The goodness-of-fit of the fitted RSFs was confirmed by Receiver Operating Characteristic (ROC) area-under-the-curve (AUC) values (AUCh = 0.86, AUCc = 0.82, ROC plots Fig. S3.2) and k-fold cross-validation, also using Kendall’s tau (fivefold, mean historical τh = 0.75, two historical k-fold subsets p < 0.001, three subsets p < 0.05; mean contemporary τc = 0.96, all contemporary k-fold subsets p < 0.001).
Temporal variation in patterns of space use
We used kappa coefficients (κ) to compare similarity between predicted relative probability-of-use (Rp) maps within and between periods (κ = 1 for identical maps, κ = − 1 for reverse-image maps). These coefficients indicated substantial inter-decadal shifts in Rp by roe deer across our study area. In agreement with Prediction 4, inter-decadal variability in Rp was significantly higher (lower κ) than year-to-year variability in all months of the contemporary period and in April of the historical period (Fig. 4; November not shown because of limited sample size). However, the majority of historical year-to-year monthly comparisons showed relatively low κ values, indicating high variability (Fig. 4a–d). Therefore, contrary to our expectations (Prediction 5a), we found significantly lower year-to-year variability in the contemporary period than in the historical one during the months of December, January, and April (both pDec and pJan < 0.001, pApr = 0.036, Fig. 4a,b,e), while the year-to-year variability in February and March was not significantly different between the two periods (Fig. 4c,d). Similarly, we found this reduction in variability continued in the forecasted warming scenarios (counter to Prediction 5b), with the inter-annual variability within forecasted scenarios being significantly lower than in the historical period (Fig. 4).

Kappa coefficients comparing the relative probability-of-use by month. YH: year-to-year (y–y in the legend) comparisons for historical study period using RSFh; ID: inter-decadal comparisons; YC: year-to-year comparisons for contemporary study period using RSFc; YI: year-to-year comparisons for intermediate forecast scenario; YS: year-to-year comparisons for severe forecast scenario. Sample size varies based on the number of years for which data was collected in that month (November not shown due to low sample size). Asterisks denote pairwise significance of difference between categories of comparison (***, p < 0.001; **, p < 0.01; *, p < 0.05).
We have included subtraction maps comparing year-to-year relative Rp, in order to visualize the sole effect of snow variation on roe deer winter distribution under constant supplemental feeding intensity (Fig. 5). For both the historical and contemporary periods, when comparing a low-snow and high-snow winter (1999/2000 and 2000/2001, respectively, for the historical period; 2014/2015 and 2013/2014 for the contemporary period; see also Fig. S2.2), low-snow years exhibited an upslope shift of areas of high Rp out of the central valley. This was particularly true in the snowier months of mid-winter (February in Fig. 5, panels b and e respectively for the historical and contemporary periods). The comparison between the same two contemporary winters and the severe warming forecasted scenario confirmed this pattern is likely to continue in the future (Fig. 5g–l). Roe deer winter habitat suitability increased under predicted warming conditions, especially when compared to the particularly snowy winter of 2013/2014 (Fig. 5g–i), while smaller shifts in high-Rp areas were seen when comparing the forecast to the low-snow winter of 2014/2015 (Fig. 5j–l).

Maps of change in relative probability-of-use (Rp). Each map represents the subtraction of one binned Rp map from another (later map–earlier map). Pixel colors range from − 9, indicating extreme preference for the pixel in the earlier year, to + 9, extreme preference in a later year. The spatial resolution is 10 m, and the contour lines indicate 250 m changes in elevation, with lowest values in the central valley (see also Fig. 1). Panels (a–f) show subtractions within the historical period (a–c, low-snow year subtracted from high-snow year) and within the contemporary period (d–f, high-snow year subtracted from low-snow year); panels (g–l) show subtractions of contemporary years (high-snow: 2013/2014 winter; low-snow: 2014/2015 winter) from the same year predicted using the severe forecast scenario.
Discussion
By making use of two tracking datasets separated by a decade and a half of anthropogenic change, we were able to empirically link an ungulate’s changing winter distribution to long-term ongoing and forecasted changes in abiotic constraints and anthropogenic resource availability. Specifically, our findings forecast a distributional upslope shift in relative probability of space use for an opportunistic, snow-constrained ungulate under current climate change, as a consequence of snow depth and snowpack changes. In our empirical setting, this response was made more complex by the attraction of animals to supplemental feeding sites, evidencing how snow depth and supplemental feeding can combine to modify the seasonal spatial distribution of ungulates and other mammals.
Resource selection models confirmed Predictions 1 and 2 in both study periods, showing a steep drop-off in relative probability-of-use at snow depths exceeding ~ 50 cm and at increasing distances from supplemental feeding sites. However, in the historical period, the attraction to supplemental feeding sites increased with snow depth (i.e., the relative probability-of-use was higher close to feeding sites in the case of deep snow), suggesting a mitigating behavioral response to the constraints associated with widespread heavy snowfall3,6. Conversely, in the contemporary period, these sites now attract roe deer almost irrespective of snow depth (i.e., the relative probability-of-use remained high close to feeding sites under almost any snow condition) (Prediction 3). As expected, we observed a substantial shift in roe deer’s relative probability of space use (i.e. distribution) from one decade to the next in response to an increased number and spread of supplemental feeding sites, together with the upwards shift of the snowline (Prediction 4). However, increased snow variability in contemporary years (given that the distribution of feeding sites is relatively static within this period) did not produce the expected increase in inter-annual Rp variability. Conversely, we found a decrease in inter-annual Rp variability in the early and late winter months (December, January, and April), and projections that this variability will continue decreasing in the forecasted scenarios (Prediction 5).
In the historical period, supplemental feeding sites mitigated the negative effects of deep snow, with higher Rp values extending in a radius of up to ~ 600–700 m from supplemental feeding sites (Fig. 3a). In the contemporary period, instead, higher Rp values extend only up to 200–300 m from feeding sites under the same snow conditions (Fig. 3b). Notably, Rp in close proximity to feeding sites remains high in the contemporary period under almost any snow conditions, showing a decreased mitigation effect, but stronger dependence on these artificial sources of food. These findings raise the question of the extent to which anthropogenic modifications of resource availability affect ungulate populations—particularly those facing particularly limiting winter conditions27. Potential drawbacks of supplementing feeding include, but are not limited to, transforming feeding sites into “ecological traps”28, where disease transmission29,30 or competition-induced stress increases15,31 can severely affect population dynamics. Behavioral responses may enable animals to adapt to anthropogenic changes32,33,34, but in many cases, these responses may be insufficient and even maladaptive28,35. For instance, supplemental feeding sites may make animals more susceptible to harsh snowfall in the studied population, as they congregate in areas they would normally avoid in the winter and lose the capacity to rely on natural food resources15,36, potentially with negative effects on their health37,38. Anecdotally, five of our collared roe deer died in one week in March 2014 following an intense snowfall, despite sustained use of feeding sites. Increased predation risk due to artificially elevated local density of roe deer around feeding sites may prove another unanticipated drawback of the practice39. While this effect is currently not a major issue in our study system (only brown bears, Ursus arctos, may predate on adult roe deer, but they hibernate in winter), it could be increasingly be the case as large carnivores expand through Europe40,41. Thus, even as the positive effects of supplemental feeding practice remain controversial42, there is rising concern around the pitfalls of the practice and its far-reaching effects on behavior43,44,45.
While the increase in supplemental feeding sites played the largest role in driving inter-decadal shifts in space use, snow variability exerted a fundamental role in shaping year-to-year variation in relative probability of space use. Remarkably, we observed the high variability expected in the contemporary period (Prediction 5a) only during the central months of the winter (February–March), while in the historical period deer exhibited relatively high space use variability all winter long. The low variability in contemporary space use patterns detected for the months of December, January, and April (Fig. 4) was likely due to scarce-to-absent snowfall in these months in contemporary years. Therefore, milder conditions at the beginning and end of winter reduced the constraint of snow cover on roe deer winter distribution and allowed relative probability-of-use to remain highly consistent from year to year during these months. Conversely, the unexpected variability in historical space use is arguably the standard variability to be expected from year-to-year changes in snow patterns, and should be contrasted with the climate change-driven lowered year-to-year variability of the contemporary period and forecasted scenarios. These patterns presage the speed with which climate-driven changes in snow cover and related animal behavioral responses are happening. The continued upward shift of the snowline along the elevation gradient in our climate change forecasts (Prediction 5b) confirms this trend and in turn forecasts substantial upslope expansion of suitable winter habitat for roe deer in the Alps.
Our work empirically supports the theoretically predicted effects of a decrease in snow coverage in the Northern Hemisphere46 on distribution range shifts and movement patterns of varied mammal species47. Such changes have been found to affect the probability of seasonal migration from summer to winter ranges in several ungulate species8,48,49, with deep snowpacks driving moose (Alces alces) to modify their fine-scale movements to search for conifer patches within mixed stands50, and Sitka deer (Odocoileus hemionus sitkensis) selecting for different successional stages of forest within their winter range according to snow depth variability51.
This study expands these findings by providing rare empirical evidence of the interplay between anthropogenic changes in climate and food supply32 in driving behavioral responses in a large herbivore at the altitudinal limit of its distribution. Moreover, it examines behavioral responses at the inter-decadal scale at which changes in the snowpack become measurable and sustained. Although our analysis is confined to the alpine watershed in which our study was executed, it is applicable to a broad guild of temperate and arctic ungulates. More generally, it provides an example of the general effects of interacting pressures on space use patterns at the range limits of all species11,27. Under predicted changes, species may change their distribution47, undergo local extirpations52, or adapt within their current range, as observed in this study.
Therefore, the topic is of great relevance—particularly to ungulates, as their proximate behavioral responses will likely have cascading effects on local ecosystems27, e.g. through changing browsing pressure, nitrogen cycling, and seed dispersal53, or through inter-specific competition54. We therefore encourage other animal ecologists to make use of localized climatological and movement data to shed light on these and other important questions linking environmental change with space use trends. Further, future research should consider the demographic consequences of these ongoing changes, particularly by linking expanding habitat suitability to population dynamics at the range limits of snow-constrained species under climate change.
Methods
Study area
All research was carried out in a 400 km2 mountainous area ranging in altitude from 400 to 3500 m in the north-eastern Italian Alps (Val Rendena and Valli Giudicarie, Autonomous Province of Trento, Italy; see Supplement S1 for details on climate, flora, and fauna). Over the study period, the area saw an increase in supplemental feeding for roe deer, from 20 to 95 actively managed sites (Fig. 1), due to modifications in hunting management regulations. The overall population density of roe deer in the area has remained relatively low and constant throughout the last two decades (i.e., between three and five deer/100 ha)55,56.
Movement data
The dataset of contemporary roe deer relocations was collected from December 18th 2012 to April 20th 2015, for a total of 33,399 winter GPS relocations (1 Nov–30 Apr; one fix every 3 h) from 23 roe deer (9 males, 14 females). Animals were captured using box traps and drop nets during two field campaigns in the winters of 2012–2013 and 2013–2014 and outfitted with GPS-GSM radio-collars (VECTRONIC Aerospace GmbH).
The dataset of historical roe deer relocations was collected from March 7th 1999 to April 30th 2002, for a total of 4529 winter VHF radio-collar relocations (one fix per animal per day22) from 32 roe deer (10 males, 22 females), captured using box traps and drop nets, then outfitted with VHF transmitters (Televilt International AB).
All animal handling practices for capturing and marking roe deer complied with Italian laws on animal welfare and were approved by the Wildlife Committee of the Autonomous Province of Trento.
Environmental data
We used the GEOtop 2.0 Hydrological Model23 (hereafter “GEOtop”) to generate a 25-year long time series (1989–2015) of daily variation in snow depth at a spatial resolution of 100 m. GEOtop is a water- and energy-balance model that produces snow depth area maps from meteorological data, taking into account snow melting processes across the landscape (see Supplement S2 for details). We also used GEOtop in conjunction with a regional climate model (COSMO-CLM)57 to generate two further 25-year long time series (hereafter “intermediate” and “severe” scenarios) using forecasts of radiative forcing from greenhouse gas emission scenarios of differing severity (IPCC RCP4.525 and RCP8.526). We assessed the accuracy of GEOtop output for our study area by comparing predicted snow depth data to empirically measured snow depth measurements collected from 2012 to 201520, and we applied a correction coefficient to remedy slight overprediction of GEOtop in forested areas (see Supplement S2). For each generated timeseries separately, we obtained monthly snow depth maps for the entire study area by averaging the daily GEOtop snow depth values over each winter month (November–April).
We derived canopy cover maps for the historical and contemporary periods, respectively, from CORINE 2000 and 2012 land cover maps (European Environment Agency 2000 and 2012, available at https://land.copernicus.eu/pan-european/corine-land-cover; hereafter "CLC 2000" and "CLC 2012"). For both maps, we ranked broad-leaf forest, coniferous forest, and mixed forest land classes as “closed” canopy, and all other classes as “open”.
Finally, we gathered supplemental feeding site locations from local hunters and wildlife managers. We used ArcGIS software (ESRI, Redlands, CA, USA) to compute two maps (one for each study period) of 3D path-distance to the nearest supplemental feeding site.
Predicting habitat selection
We estimated two resource selection functions (RSFs) to predict roe deer space use patterns across the study area in the historical and in the contemporary periods. RSFs are functions proportional to the probability a given animal will select a resource unit. They are commonly used to understand species’ distributions and space use by characterizing the relative probability-of-use of a given site as a log-linear product of its characteristics and appropriate selection or avoidance coefficients (see Eq. 1 below)58. We calculated these RSF coefficients by fitting a logistic regression to telemetry-derived locations used by roe deer (case = 1) between 1999 and 2002 (historical RSF, “RSFh”) or 2012–2015 (contemporary RSF, “RSFc”) and to available locations (case = 0)59. After confirming the consistency between daily and nocturnal versions of the GPS-based RSFc (see Supplement S4), we retained only GPS relocations between sunrise and sunset for the RSFc used in the analysis (daylight deduced using the suncalc R package60). Limitation to daylight hours was done to permit strict comparison of the VHF- (historical) and GPS-based (contemporary) models, since VHF data had been only gathered during daylight hours. For both RSFs, we randomly selected one used point per animal per day (final used sample: GPS 4625 points, VHF 4013 points) in order to reduce autocorrelation between points and to further strengthen the comparison between VHF- and GPS-based datasets, given their different temporal resolutions. For both RSFs, available points were randomly sampled at a 1:1 ratio to used points61 within 99% kernel density polygons (bivariate normal with reference bandwidth) of all VHF and GPS roe deer locations, respectively.
We estimated the historical (RSFh) and contemporary (RSFc) relative probability of space use (used/available) at a monthly scale, as a function of (i) monthly averages of GEOtop-derived snow depth (SD) (Prediction 1), (ii) the reciprocal of the distance between the location and the nearest supplemental feeding site (1/DistFS, “proximity”) (Prediction 2), and (iii) the interaction between these two covariates (SD/DistFS) (Prediction 3), while controlling for canopy cover as a fixed effect (CC). We chose 1/DistFS rather than the distance to supplemental feeding sites because, when predicting over large areas, an ever-decreasing linear effect of supplemental feeding sites on habitat selection becomes biologically unrealistic beyond perception limits. All predictions of relative probability-of-use (Rp) were based on a linear scaling (from 0 to 1, Eq. 2) of the log-linear form in Eq. 1 (typical for RSFs59):
where
We evaluated the reliability of both RSFh and RSFc using Receiver Operating Characteristics (ROC) curves and k-fold cross-validation59 with Kendall’s tau (τ) coefficients62 (which are typically more robust than Spearman’s rho). In k-fold cross-validation, data are partitioned into k equal subsets (5 in this study), and each subset is used to define a model. τ coefficients measure the correlation between rankings of model predictions in the remaining data and area-adjusted frequencies of observed habitat use59. We also used RSFh to predict Rp for 1999–2002 and RSFc to predict Rp for 1999–2002 and 2012–2015, further using τ coefficients to spatially and temporally validate the models’ ability to predict actual space use (Supplement S5). We thus considered significant τ coefficients produced within a model’s respective study period to be a spatial validation of that model (spatial RSFh τ in 1999–2002, spatial RSFc τ in 2012–2015) and significant τ coefficients produced in 1999–2002 for RSFc to be a temporal validation of the contemporary model’s predictive ability (temporal RSFc).
Finally, we also forecasted Rp using RSFc and snow depths estimates from both forecast scenarios of climate change (intermediate scenario = GEOtop monthly average outputs 2012–2015 with RCP4.5 correction + CLC 2012 + distribution of supplemental feeding sites in 2012–2015; severe scenario = GEOtop monthly average outputs 2012–2015 with RCP8.5 correction + CLC 2012 + distribution of supplemental feeding sites in 2012–2015).
Evaluating temporal changes in patterns of space use
We used weighted Kappa coefficients24 to examine how changes in snow patterns and supplemental feeding site distribution have affected variability in spatial patterns of Rp of identical months between periods (“inter-decadal”, Prediction 4) and within a study period (“year-to-year”, Prediction 5). Kappa coefficients (κ) provide a proportion of agreement (“similarity”) between two maps’ binned Rp values (pixel-by-pixel), accounting for the probability of overlap between randomly-distributed Rp values (e.g., 0.1 * 0.1 = 1% of the lowest 10% of Rp values should overlap by chance in any two maps). κ coefficients take on a value of 1 for perfect agreement and values near 0 for agreement no larger than expected by chance24, thus providing a robust means of comparison between entire spatial surfaces.
For each winter month (Nov–Apr) we extracted monthly Rp values (generated with the corresponding period’s RSF) for 1000 random locations within the intersection of both study periods’ 99% kernel polygons and binned these into 10 quantiles established from the pooled Rp values generated from RSFc across the entire study. We then used these binned values to compute a κ coefficient, weighted by the degree of rank disagreement63. For a given winter month, we thus calculated a κ coefficient for each possible annual pairwise combination (a) within the historical study period (“intrah”), (b) within the contemporary study period (“intrac”), (c) between the two study periods (“inter-decadal”), (d) within the intermediate scenario (“intrai”), and (e) within the severe scenario (“intras”). For each month, we used a one-way ANOVA and post-hoc Tukey tests to evaluate the significance of differences between the κ coefficients of these five categories.
Finally, we created subtraction maps to visually represent the comparisons captured by the κ coefficients within each period (i.e. year-to-year), thus keeping fixed the availability of supplemental feeding sites. We chose three months to represent different parts of the winter: December (early winter), February (mid-winter), and April (late winter). For each month, we firstly created maps of Rp, binned into the same 10 quantiles developed for calculating the κ coefficients. Then, we computed four subtraction maps in order to visually illustrate the areas of highest difference in relative probability-of-use. Specifically, both for the historical study period (intrah) and for the contemporary one (intrac), we compared a dry and a snowy winter (1999/2000 and 2000/2001, respectively, for the historical period; 2014/2015 and 2013/2014 for the contemporary period; information on snow characteristics in 2014 and 2015 were extracted from GEOtop layer, see also Fig. S2.2). We also created subtraction maps to compare both contemporary winters with the maps predicted under the most severe warming scenario. In all cases, the earlier map was subtracted from the later map.
Kernel polygons and path distances between pixels were generated in ArcGIS spatial software (ESRI, Redlands, CA, USA). All other statistical analyses and spatial analyses were conducted using R 3.4.3 (key packages used: irr64, ResourceSelection65, rgdal66, Kendall67).
Data availability
The data are available in the Eurodeer repository (http://www.eurodeer.org), accessible following user registration. The datasets are also available in Zenodo athttps://doi.org/10.5281/zenodo.4637674. (Bright Ross et al68, https://doi.org/10.5281/zenodo.4637674).
References
Weiner, J. Physiological limits to sustainable energy budgets in birds and mammals: ecological implications. Trends Ecol. Evol. 7, 384–388 (1992).
Mcnab, B. K. Food habits, energetics, and the population biology of mammals. Am. Nat. 116, 106–124 (1980).
Hovey, F. W. & Harestad, A. S. Estimating effects of snow on shrub availability for black-tailed deer in southwestern British Columbia. Wildl. Soc. Bull. 20, 308–313 (1992).
Post, E. & Stenseth, N. Climatic variability, plant phenology, and northern ungulates. Ecology 80, 1322–1339 (1999).
Moen, A. N. Seasonal changes in heart rates, activity, metabolism, and forage intake of white-tailed deer. J. Wildl. Manag. 42, 715–738 (1978).
Holand, Ø., Mysterud, A., Wannag, A. & Linnell, J. D. C. Roe deer in northern environments: physiology and behaviour. In The European Roe Deer: Biology of Success (eds Andersen, R. et al.) 117–137 (Scandinavian University Press, 1998).
Foromozov, A. N. Snow Cover as an Integral Factor of the Environment and Its Importance in the Ecology of Mammals and Birds (The University of Alberta, 1963).
Cagnacci, F. et al. Partial migration in roe deer: migratory and resident tactics are end points of a behavioural gradient determined by ecological factors. Oikos 120, 1790–1802 (2011).
Dussault, C., Courtois, R., Ouellet, J.-P. & Girard, I. Space use of moose in relation to food availability. Can. J. Zool. 83, 1431–1437 (2005).
Mysterud, A. & Sæther, B.-E. Climate change and implications for the future distribution and management of ungulates in Europe. In Ungulate Management in Europe: Problems and Practices (eds Putman, R. et al.) 349–375 (Cambridge University Press, 2011).
Parmesan, C. Ecological and evolutionary responses to recent climate change. Annu. Rev. Ecol. Evol. Syst. 37, 637–669 (2006).
Scherrer, S. C., Wüthrich, C., Croci-Maspoli, M., Weingartner, R. & Appenzeller, C. Snow variability in the Swiss Alps 1864–2009. Int. J. Climatol. 33, 3162–3173 (2013).
Milner, J. M., van Beest, F. M., Schmidt, K. T., Brook, R. K. & Storaas, T. To feed or not to feed? Evidence of the intended and unintended effects of feeding wild ungulates. J. Wildl. Manag. 78, 1322–1334 (2014).
Ossi, F. et al. Plastic response by a small cervid to supplemental feeding in winter across a wide environmental gradient. Ecosphere 8, e01629 (2017).
Putman, R. & Staines, B. W. Supplementary winter feeding of wild red deer Cervus elaphus in Europe and North America: justifications, feeding practice and effectiveness. Mamm. Rev. 34, 285–306 (2004).
Cagnacci, F., Boitani, L., Powell, R. A. & Boyce, M. S. Animal ecology meets GPS-based radiotelemetry: a perfect storm of opportunities and challenges. Philos. Trans. R. Soc. B Biol. Sci. 365, 2157–2162 (2010).
Peters, W. et al. Migration in geographic and ecological space by a large herbivore. Ecol. Monogr. 87, 297–320 (2017).
Morellet, N. et al. Seasonality, weather and climate affect home range size in roe deer across a wide latitudinal gradient within Europe. J. Anim. Ecol. 82, 1326–1339 (2013).
Johnson, D. H. The comparison of usage and availability measurements for evaluating resource preference. Ecology 61, 65–71 (1980).
Ossi, F., Gaillard, J. M., Hebblewhite, M. & Cagnacci, F. Snow sinking depth and forest canopy drive winter resource selection more than supplemental feeding in an alpine population of roe deer. Eur. J. Wildl. Res. 61, 111–124 (2015).
Mysterud, A. & Østbye, E. Bed-site selection by European roe deer (Capreolus capreolus) in southern Norway during winter. Can. J. Zool. 73, 924–932 (1995).
Ramanzin, M., Sturaro, E. & Zanon, D. Seasonal migration and home range of roe deer (Capreolus capreolus) in the Italian eastern Alps. Can. J. Zool. 85, 280–289 (2007).
Endrizzi, S., Gruber, S., Dall’Amico, M. & Rigon, R. GEOtop 2.0: simulating the combined energy and water balance at and below the land surface accounting for soil freezing, snow cover and terrain effects. Geosci. Model. Dev. 7, 2831–2857 (2014).
Cohen, J. A coefficient of agreement for nominal scales. Educ. Psychol. Meas. 20, 37–46 (1960).
Thomson, A. M. et al. RCP 4.5: a pathway for stabilization of radiative forcing by 2100. Clim. Change 109, 77–94 (2011).
Riahi, K. et al. RCP 8.5—a scenario of comparatively high greenhouse gas emissions. Clim. Change 109, 33–57 (2011).
Thomas, C. D. Climate, climate change and range boundaries. Divers. Distrib. 16, 488–495 (2010).
Penteriani, V. et al. Evolutionary and ecological traps for brown bears Ursus arctos in human-modified landscapes. Mamm. Rev. 48, 180–193 (2018).
Sorensen, A., van Beest, F. M. & Brook, R. K. Impacts of wildlife baiting and supplemental feeding on infectious disease transmission risk: a synthesis of knowledge. Prev. Vet. Med. 113, 356–363 (2014).
Mysterud, A., Viljugrein, H., Solberg, E. J. & Rolandsen, C. M. Legal regulation of supplementary cervid feeding facing chronic wasting disease. J. Wildl. Manag. 83, 1667–1675 (2019).
Ceacero, F. et al. Benefits for dominant red deer hinds under a competitive feeding system: food access behavior, diet and nutrient selection. PLoS ONE 7, e32780 (2012).
Beever, E. A. et al. Behavioral flexibility as a mechanism for coping with climate change. Front. Ecol. Environ. 15, 299–308 (2017).
Loe, L. E. et al. Behavioral buffering of extreme weather events in a high-Arctic herbivore. Ecosphere 7, e01374 (2016).
Sih, A., Ferrari, M. C. O. & Harris, D. J. Evolution and behavioural responses to human-induced rapid environmental change. Evol. Appl. 4, 367–387 (2011).
Radchuk, V. et al. Adaptive responses of animals to climate change are most likely insufficient. Nat. Commun. 10, 3109 (2019).
Mysterud, A. Still walking on the wild side? Management actions as steps towards ‘semi-domestication’ of hunted ungulates. J. Appl. Ecol. 47, 920–925 (2010).
Felton, A. M. et al. Interactions between ungulates, forests, and supplementary feeding: the role of nutritional balancing in determining outcomes. Mamm. Res. 62, 1–7 (2017).
Ricci, S. et al. Impact of supplemental winter feeding on ruminal microbiota of roe deer Capreolus capreolus. Wildl. Biol. 2019, wlb.00572 (2019).
Lone, K. et al. Living and dying in a multi-predator landscape of fear: roe deer are squeezed by contrasting pattern of predation risk imposed by lynx and humans. Oikos 123, 641–651 (2014).
Chapron, G. et al. Recovery of large carnivores in Europe’s modern human-dominated landscapes. Science (80-) 346, 1517–1519 (2014).
Milanesi, P., Breiner, F. T., Puopolo, F. & Holderegger, R. European human-dominated landscapes provide ample space for the recolonization of large carnivore populations under future land change scenarios. Ecography (Cop.) 40, 1359–1368 (2017).
Pascual-Rico, R. et al. Is diversionary feeding a useful tool to avoid human-ungulate conflicts? A case study with the aoudad. Eur. J. Wildl. Res. 64, 1–7 (2018).
van Beest, F. M., Loe, L. E., Mysterud, A. & Milner, J. M. Comparative space use and habitat selection of moose around feeding stations. J. Wildl. Manag. 74, 219–227 (2010).
Jerina, K. Roads and supplemental feeding affect home-range size of Slovenian red deer more than natural factors. J. Mamm. 93, 1139–1148 (2012).
Ranc, N. et al. Preference and familiarity mediate spatial responses of a large herbivore to experimental manipulation of resource availability. Scientific Reports 10, 11946 (2020).
Brown, R. D. & Robinson, D. A. Northern Hemisphere spring snow cover variability and change over 1922–2010 including an assessment of uncertainty. Cryosphere 5, 219–229 (2011).
Schloss, C. A., Nuñez, T. A. & Lawler, J. J. Dispersal will limit ability of mammals to track climate change in the Western Hemisphere. Proc. Natl. Acad. Sci. U. S. A. 109, 8606–8611 (2012).
Gurarie, E. et al. A framework for modelling range shifts and migrations: asking when, whither, whether and will it return. J. Anim. Ecol. 86, 943–959 (2017).
Rivrud, I. M. et al. Leave before it’s too late: anthropogenic and environmental triggers of autumn migration in a hunted ungulate population. Ecology 97, 1058–1065 (2016).
Courtois, R., Dussault, C., Potvin, F. & Daigle, G. Habitat selection by moose (Alces alces) in clear-cut landscapes. Alces 38, 177–192 (2002).
Gilbert, S. L., Hundertmark, K. J., Person, D. K., Lindberg, M. S. & Boyce, M. S. Behavioral plasticity in a variable environment: snow depth and habitat interactions drive deer movement in winter. J. Mamm. 98, 246–259 (2017).
Chevin, L. M., Lande, R. & Mace, G. M. Adaptation, plasticity, and extinction in a changing environment: towards a predictive theory. PLoS Biol. 8, e1000357 (2010).
Bauer, S. & Hoye, B. J. Migratory animals couple biodiversity and ecosystem functioning worldwide. Science (80-) 344, 1242552 (2014).
Mason, T. H. E., Stephens, P. A., Apollonio, M. & Willis, S. G. Predicting potential responses to future climate in an alpine ungulate: Interspecific interactions exceed climate effects. Glob. Change Biol. 20, 3872–3882 (2014).
Carnevali, L., Pedrotti, L., Riga, F. & Toso, S. Banca dati ungulati: Status, distribuzione, consistenza, gestione e prelievo venatorio delle popolazioni di ungulati in Italia. Rapporto 2001–2005 Vol. 117 (Biologia e Conservazione della Fauna, 2009).
Provincia Autonoma di Trento. Analisi delle consistenze e dei prelievi di ungulati, tetraonidi e coturnice. Stagione Venatoria 2018 (Provincia Autonoma di Trento, 2018).
Rockel, B., Will, A. & Hense, A. The regional climate model COSMO-CLM (CCLM). Meteorol. Z. 17, 347–348 (2008).
Boyce, M. S. & McDonald, L. L. Relating populations to habitats using resource selection functions. Trends Ecol. Evol. 14, 268–272 (1999).
Boyce, M. S., Vernier, P. R., Nielsen, S. E. & Schmiegelow, F. K. A. Evaluating resource selection functions. Ecol. Modell. 157, 281–300 (2002).
Benoit, T. & Achraf, E. suncalc: compute sun position, sunlight phases, moon position and lunar phase. R package version 0.5.0. https://cran.r-project.org/package=suncalc (2019).
DeCesare, N. J. et al. Transcending scale dependece in identifying habitat with resource selection functions. Ecol. Appl. 22, 1068–1083 (2012).
Kendall, M. A new measure of rank correlation. Biometrika 30, 81–89 (1938).
Cohen, J. Weighted kappa: nominal scale agreement with provision for scaled disagreement or partial credit. Psychol. Bull. 70, 213–220 (1968).
Gamer, M., Lemon, J., Fellows, I. & Singh, P. irr: various coefficients of interrater reliability and agreement. R package version 0.84.1. https://cran.r-project.org/package=irr (2019).
Lele, S. R., Keim, J. L. & Solymos, P. ResourceSelection: resource selection (probability) functions for use-availability data. R package version 0.3-5. https://cran.r-project.org/package=ResourceSelection (2019).
Bivand, R., Keitt, T. & Rowlingson, B. rgdal: bindings for the ‘Geospatial’ Data Abstraction Library. R package version 1.4-8. https://cran.r-project.org/package=rgdal (2019).
McLeod, A. I. Kendall: Kendall rank correlation and Mann-Kendall trend test. R package version 2.2. https://cran.r-project.org/package=Kendall (2011).
Bright Ross, J. G., Peters, W., Ossi, F., Moorcroft P. R., Cordano, E., Eccel, E., Bianchini, F., Ramanzin, M., and Cagnacci, F. Datasets for "Climate change and anthropogenic food manipulation interact in shifting the distribution of a large herbivore at its altitudinal range limit." https://doi.org/10.5281/zenodo.4637674 (2021).
Acknowledgements
This paper represents a collaborative effort within the framework of the EURODEER project (paper no. 13 of the EURODEER paper series, www.eurodeer.org). The co-authors are grateful to all EURODEER members for their support throughout the project. The EURODEER spatial database is hosted by Fondazione Edmund Mach. F. Cagnacci, F. Ossi and W. Peters thank the Wildlife and Forestry Service and the Trentino Hunting Association for support and help during captures. Thanks to L. Zollo and P. Mercogliano (CMCC) for the use of climate scenarios. Travel for study was partially funded by a Herchel Smith Fellowship and a Harvard University Museum of Comparative Zoology Grant-In-Aid of Undergraduate Research, for which J. Bright Ross extends his most sincere thanks. F. Cagnacci was supported by the Sarah and Daniel Hrdy Fellowship (2015–2016 at Harvard University OEB) during part of the development of this manuscript. The majority of this analysis was undertaken as part of J. Bright Ross' undergraduate thesis with the Harvard University Department of Organismic and Evolutionary Biology. Lastly, the authors are grateful to the two anonymous reviewers whose suggestions improved the clarity and impact of this manuscript.
Author information
Authors and Affiliations
Contributions
F.C., W.P., and F.O. conceived the concept; F.O., M.R., and F.B. collected the data; J.B., W.P., and F.C. designed the methodology; J.B., W.P., and E.C. conducted the analysis; E.C. and E.E. generated climatological data; J.B., F.C., F.O., W.P., and P.M. led the writing of the manuscript. All authors contributed critically to drafts of the manuscript and approved the final version for publication.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Bright Ross, J.G., Peters, W., Ossi, F. et al. Climate change and anthropogenic food manipulation interact in shifting the distribution of a large herbivore at its altitudinal range limit. Sci Rep 11, 7600 (2021). https://doi.org/10.1038/s41598-021-86720-2
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-021-86720-2
This article is cited by
-
Links between energy budgets, somatic condition, and life history reveal heterogeneous energy management tactics in a group-living mesocarnivore
Movement Ecology (2024)
-
Land use drives differential resource selection by African elephants in the Greater Mara Ecosystem, Kenya
Movement Ecology (2024)
-
Evaluating density-weighted connectivity of black bears (Ursus americanus) in Glacier National Park with spatial capture–recapture models
Movement Ecology (2024)
-
Occupancy and activity rhythms of the Siberian roe deer
Biologia (2021)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
