
Abstract
Antimicrobial stewardship isn’t strictly observed in most Egyptian hospitals, raising antibiotic resistance. Epidemiology of Egyptian MRSA isolates, or associations with resistance to other antibiotics remain largely unknown. We identified MRSA genotypes in Alexandria Main University Hospital (AMUH) and investigated rates of moxifloxacin resistance, an alternative MRSA treatment, among different genotypes. Antibiotic susceptibility of 72 MRSA clinical isolates collected in 2015 from AMUH was determined by disc diffusion and broth microdilution. spa- and Staphylococcal Cassette Chromosome mec (SCCmec) typing were performed; with multi-locus sequence typing conducted on isolates representing major genotypes. Resistance to moxifloxacin, levofloxacin and ciprofloxacin were 69%, 78% and 96%, respectively. spa type t037 (57%) was commonest, followed by t127 (12.5%), t267 (8%) and t688 (6%). SCCmec III predominated (57%), all of these were moxifloxacin resistant and 97.6% t037 (ST241). SCCmec IV, IV E and V represented 15%, 7% and 11% of the isolates, respectively, 79% of these were moxifloxacin susceptible and of different spa types. t127 (ST-1) was associated with SCCmec V in 56% of the isolates, mostly moxifloxacin susceptible. Moxifloxacin resistance was high, most resistant isolates belonged to t037 and SCCmec III, suggesting local dissemination and antibiotic pressure. We recommend caution in treating MRSA infections with moxifloxacin.
Similar content being viewed by others

Introduction
Staphylococcus aureus (S. aureus) is a notorious Gram positive pathogen, with clinical presentations ranging from minor skin infections to life threatening invasive infections, including pneumonia, endocarditis, osteomyelitis and bacteremia1. Methicillin resistant Staphylococcus aureus (MRSA) strains emerged in the early 1960s, and were mainly confined to hospitals (HA-MRSA)2. This changed in the 1990s when community acquired MRSA (CA-MRSA) emerged as a clear and present danger3. Since then, cases of CA-MRSA have been reported globally4 . In Egypt, MRSA prevalence is about 52%, a rate that is higher than most other African countries (< 50%)5,6.
Methicillin resistance in S. aureus occurs due to the expression of an additional penicillin binding protein 2a (PBP2a) with a low affinity to methicillin and most other β-lactams7. This ensures continuous cell wall synthesis even in the presence of the antibiotic8. PBP2a is encoded by a chromosomal gene mecA present in a mobile genetic element called the staphylococcal cassette chromosome (SCCmec)9. Over the years, methicillin resistance has driven the use of other antibiotic classes for treatment of MRSA infections, which resulted in the evolution of multi-drug resistant strains10. Consequently, only a few agents are still available nowadays to treat MRSA infections. While the glycopetides remain the mainstay of treatment for MRSA infections, there is limited availability of options for oral therapy. These include clindamycin, trimethoprim-sulfamethoxazole, tetracycline, and linezolid11. Another option, available orally, to treat MRSA infections is the fourth generation fluoroquinolone: moxifloxacin12. Unfortunately, resistance to these agents is being increasingly reported globally11,13.
Several studies have reported on moxifloxacin resistance rates12,14,15,16,17. However, very little is known about the level of moxifloxacin resistance among Egyptian MRSA isolates. In 2007, a study reported moxifloxacin resistance rate of 30.8% among S. aureus isolates from Egyptian cancer patients18. Another study conducted in three university hospitals in Upper Egypt found 40% of isolated MRSA from patients with hospital acquired (HA) pneumonia to be moxifloxacin resistant19. A more recent study, however, reported a lower level of resistance (6.3%) among S. aureus collected between 2015 and 2018 in Zagazig, Egypt20. None of these studies commented on the genotype of the studied isolates.
Bacterial typing is essential for understanding the epidemiologic and evolutionary relationships between bacterial strains, and hence for devising infection prevention strategies21. Moreover, MRSA genotype could affect the strain’s virulence profile with an impact on the clinical outcome making the identification of MRSA genotype important to optimize the effectiveness of MRSA therapy22,23.
Different molecular typing methods are currently used for S. aureus and MRSA typing, including SCCmec typing, staphylococcus protein A typing (spa typing), multilocus sequence typing (MLST) and pulsed-field gel electrophoresis (PFGE)24. Sequence based genotyping methods such as MLST and spa typing are better at describing the evolutionary relationships and help understand the molecular epidemiological dynamics of S. aureus transmission than other methods25,26.
In the current study, we report the prevalence and level of fluoroquinolone resistance among 72 MRSA isolates collected in 2015 from Alexandria, Egypt, as well as the associations between methicillin resistance genotype and fluoroquinolone resistance.
Results
Antibiotic susceptibility among the bacterial isolates
The majority of the isolates were from male patients (n = 43, 60%) and were isolated from non-ICU locations (n = 56, 78%), the median age was 35 years. We were able to categorize 56 of the isolates as being either CA or HA, of these 80% (n = 45) were HA and 20% (n = 11) were CA (Table 1).
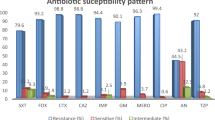
More isolates were susceptible to moxifloxacin (31%) than to the other tested fluoroquinolones. Moxifloxacin MICs ranged from < 0.125 to 32 μg/ml, with MIC50 and MIC90 values of 4 and 8 μg/ml, respectively. Ninety six percent of the isolates were ciprofloxacin resistant with MICs between 8 and 128 μg/ml and both MIC50 and MIC90 of 128 μg/ml. Seventy eight percent of the isolates were levofloxacin resistant with MICs ranging from 0.25 to 64 μg/ml and MIC50 and MIC90 of 16 μg/ml (Table 2). Seventy eight percent of the HA isolates were moxifloxacin resistant, compared to 64% of CA isolates (Fig. 1).

Moxifloxacin resistance among the isolates in hospital acquired (HA) and community acquired (CA) infections. Moxifloxacin resistance was seen among 78% of HA isolates, 64% of CA isolates and 50% of isolates of unknown infection type.
spa typing of the isolates
Eleven spa types were identified; spa type t037 (57%), t127 (12.5%), t267 (8%), t688 (6%), t223 (4%), t044 (4%) , t304 (3%), t786 (1%), t416 (1%), t6978 (1%), and a newly assigned spa type (t16221) with repeat succession (07–56-12–17-16–33-31–57-21–12) (1%). BURP analysis identified a single cluster, representing spa types t223 (n = 3) and t6978 (n = 1), and 8 singletons representing 93% of isolates. One spa type, t416 (n = 1) was excluded from the BURP analysis. Based on spa phylogeny, the isolates belonged to three clades: the first included spa types t044, t267, t786 and t127, the second clade comprised types t304, t037 and the newly identified t1622 while the rest of the identified spa types (t688, t223 and t6978) belonged to the third clade (Fig. 2).

Phylogenetic tree of the spa types detected using the neighbour-joining method. The isolates were grouped in three clades, clade 1 containing isolates belonging to spa types t044, t267, t786 and t127, clade 2 containing isolates of spa t304, t037 and t1622 and clade 3 containing isolates from spa t688, t223 and t6978.
SCCmec typing of the isolates
Forty-one isolates (57%) harbored SCCmec type III, the vast majority (98%) of which were also spa type t037 (Supplementary Table S1). All of the isolates with SCCmec III were moxifloxacin resistant and 72.5% were from HA infections. The remaining isolates were identified as SCCmec types IV (15%), IVE (7%), V (11%) and an unidentified group (10%) and were mainly moxifloxacin susceptible (Fig. 3).

Distribution of moxifloxacin resistance among the different SCCmec types detected in the isolate collection. All SCCmec III isolates were moxifloxacin resistant whereas SCCmec IV, IVE and V isolates were mostly moxifloxacin susceptible.
Isolates belonging to spa type t127 were predominantly moxifloxacin susceptible (78%). Sixty seven percent of spa type t267 isolates had SCCmec type IV, and were moxifloxacin susceptible. Fifty six percent (n = 5) of isolates belonging to spa type t127 also belonged to SCCmec type V and all but one isolate were moxifloxacin susceptible. Moxifloxacin resistance was variable among isolates belonging to spa types t688, t223, t044 and t304. One isolate of spa type t688 was identified as SCCmec IVE and the remaining isolates were of unknown SCCmec types. All isolates of spa type t223 and spa type t044 carried SCCmec IVE and SCCmec IV, respectively. Two isolates belonged to spa type t304; one isolate was moxifloxacin resistant and SCCmec III while the other isolate was moxifloxacin susceptible and SCCmec IV. spa types t416, t6978, t786 and t16221 were identified only once and in moxifloxacin susceptible isolates (Supplementary Table S1, Figs. 3–4). The newly identified spa type 16,221 was associated with SCCmec V.

Distribution of moxifloxacin resistance among the spa types detected in the isolate collection. All isolates belonging to spa type t037 were moxifloxacin resistant. Isolates belonging to all other spa types were mostly (71%) moxifloxacin susceptible.
Multi-locus sequence typing of representative isolates
Nine MLST profiles (ST-22, ST-1, ST-5, ST-6, ST-80, ST-97, ST-239, ST-241, ST-1502) were identified in addition to a new profile (3-1-1-1-1-99-3) which was assigned as MLST ST-4808. The MLST phylogenetic tree showed that the identified MLST types were related to each other within four groups; ST-239 and ST-241 were single locus variants (SLV) and moxifloxacin resistant, and were grouped under the same clonal complex (CC-8). Both ST-80 and ST-1502 had no clonal complex, yet only ST-1502 was moxifloxacin resistant. The novel type, ST-4808 was a SLV of ST-97, both of which belonging to CC-97 and a triple locus variant (TLV) of ST-1, and all were moxifloxacin susceptible. ST-5 and ST-6 were in one group (CC-5) with double locus variant (DLV) relation, both represented by isolates that were moxifloxacin susceptible. On the other hand, ST-22 was identified as a singleton with no relation to other types and was represented by a moxifloxacin susceptible isolate (Fig. 5).

Evolutionary relationships and relatedness of MLST types using eBURST algorithm in phyloviz software. The identified MLST types belonged to four groups of ST and one singleton ST (ST-22 in CC-22). One group consisted of ST-80 and ST-1502, a second group of ST-5 and ST-6 within CC-5, a third group contained ST-1 (CC-1) and ST-97 and ST-4808 (CC-97) and a fourth group of ST-239 and ST-241 (CC-8).
Discussion
MRSA remains a major causative agent of infections on a global scale, with worse outcomes relative to methicillin susceptible infections27,28. The prevalence of MRSA in Egypt is high (52%) compared to other North African and Mediterranean countries such as Morocco (19%), Libya (31%), Algeria and Tunisia (45%)6. In this scenario, the treatment of infections caused by MRSA requires the use of an alternative agent such as moxifloxacin12, that is readily available on the Egyptian market. Few reports have described the levels of moxifloxacin resistance among Egyptian S. aureus18,19,20. Tackling moxifloxacin resistance in Egypt requires a better understanding of the molecular lineages of Egyptian MRSA clones21. In this study, moxifloxacin resistance among 72 Egyptian MRSA clinical isolates was determined relative to their genotype.
Rates of resistance to fluoroquinolones were higher than previously reported29,30, and although moxifloxacin had the highest level of in-vitro susceptibility, sixty nine percent of the isolates were moxifloxacin resistant. SCCmec type III was the most common SCCmec type and almost always associated with spa type t037 (ST-241) and moxifloxacin resistance. This is not surprising as the spa t037/SCCmec III clone is a predominant clone in Africa25. The association between spa t037/SCCmec III/ST-241 and moxifloxacin resistance seen among the studied isolates is in agreement with previous findings that SCCmec III isolates showed 89% resistance to ciprofloxacin31. Moreover, a previous study described a spa t037/SCCmec III/ST-241 clone that was moxifloxacin resistant among isolates from Nigeria32. Almost 70% of the SCCmec type III isolates in the current study were HA-MRSA, in accordance with previous reports that HA-MRSA strains mainly belong to SCCmec types I, II and III, while CA-MRSA strains are mainly of SCCmec types IV and V33,34,35. In the current study, isolates belonging to SCCmec types IV, IVE and V were found among HA-MRSA as well as CA MRSA infections. Upon isolate collection, data on prior hospitalization was not obtained which could have changed the classification as HA or CA and might explain the discordance. However the association between SCCmec type and community- or hospital onset of infection is also less clear cut, possibly due to spread of hospital strains into the community and vice versa. In addition, the limited number of CA MRSA isolates in the current study makes it harder to draw solid conclusions.
The moxifloxacin resistant ST-241-III/t037 clone might represent the majority of the tested isolates as spa type t037 and SCCmec III were the most predominant types. This agrees with a study that showed that the major clonal complex causing HA-MRSA in Africa was ST-239/ST-241- III36. However, to the best of the authors’ knowledge this is the first report of the moxifloxacin resistant ST-239-III/t304 clone.
The majority of the isolates belonging to SCCmec types IV, IVE and V were susceptible to moxifloxacin with resistance rates of 18%, 20% and 25%, respectively. This concurs with Kilic et al. who showed levofloxacin resistance of 16.8% among SCCmec type IV isolates37. The moxifloxacin susceptible ST-1-V/t127 MRSA clone has been reported among human isolates in a European study, yet the porcine MRSA isolates from the same study were 95% resistant to ciprofloxacin38. The ST-80-IV/t416, ST-97-V/t267, ST-5-IVE/t688, ST-22-IVE/t223 and ST-22-IVE/t6978 clones reported here were all moxifloxacin susceptible. ST-97-V/t267 was previously detected in a ciprofloxacin susceptible bovine isolate from Italy39, the current study represents the first detection of this clone in Egypt from humans. ST-4808 is a SLV from ST-97 and the moxifloxacin susceptible clone ST-4808-IV/t267 is a novel clone firstly identified in Egypt from a human specimen in the present work. The ST-5-IVE/t688 is another new clone reported in the current study.
In conclusion, there were five new MRSA clones identified in this study; ST-239-III/t304, ST-1502-IV/t044, ST-4808-IV/t267, ST-22-IVE/t223 and ST-22-IVE/t6978 in addition to ST-97-V/t267 clone that was reported for the first time among human MRSA isolates. Resistance to fluoroquinolones was common (69% resistance to moxifloxacin), and appears to be driven by the predominant spa t037/SCCmec III clone which is moxifloxacin resistant. This may point to a potential clonal dissemination of this strain within hospitals. spa type t16221 and MLST ST-4808 type were newly identified in the present study, and were both moxifloxacin susceptible. The high rates of moxifloxacin resistance detected among the isolates calls for stricter implementation of antimicrobial stewardship guidelines and infection control practices among Egyptian hospitals.
Materials and methods
Sample collection and identification
Seventy two MRSA isolates collected from the Medical Microbiology laboratory at Alexandria Main University Hospital (AMUH) between September and December 2015 were included in the study. AMUH is the largest teaching hospital in northern Egypt with four satellite hospitals and a total capacity of 3500 beds. The isolates represented all non-duplicate MRSA isolates obtained from different clinical specimens, including pus, blood, sputum, urine, tissue, aspirate and broncho-alveolar lavage (BAL). The identity of the isolates was confirmed using Gram staining, bacterial growth and fermentation of mannitol salt agar, growth on DNase agar and slide coagulase testing using Dryspot Staphytect Plus (Oxoid Ltd, England)40. The isolates were classified as obtained from HA versus CA infections based on a 48 h window between admission and specimen collection.
Antibiotic susceptibility testing
Methicillin resistance was confirmed by cefoxitin disc diffusion testing (Oxoid Ltd, England), and susceptibility of the isolates to moxifloxacin, ciprofloxacin and levofloxacin was determined using disc diffusion and confirmed by Minimum Inhibitory Concentration (MIC) determination using a broth microdilution method41. All susceptibility tests were performed and interpreted according to the 2015 Clinical and Laboratory Standards Institute (CLSI) guidelines41.
SCCmec typing
DNA was extracted by boiling a suspension of colonies in 300 µl sterile distilled water for 30 min at 95 °C, followed by immediate cooling at -20 °C for 30 min. The suspension was centrifuged at 16,000 × g for 10 min and the supernatant was used in subsequent polymerase chain reactions (PCR)42. SCCmec typing was performed by multiplex PCR according to the protocol published by Milheirico et al.43. DNA extracts from six reference strains (Strains BAA-3843, BAA-168144, BAA-3943, BAA-168045, WIS(WBG8318)46 and BAA-4243) representing six SCCmec types (I, II, III, IV, V and VI) were included in each PCR reaction.
spa typing
The polymorphic X region of spa gene was amplified according to Harmsen et al.47 and the PCR products were Sanger sequenced using the ABI 3130XL Genetic analyzer (Inqaba Biotechnologies, South Africa;). Chromatograph sequence files were processed using the BioEdit Sequence Alignment software for creation of the consensus sequence. Sequence analysis was performed using spatyper online (http://spatyper.fortinbras.us) and/or the Ridom StaphType software (Munster, Germany) and spa clonal complexes (spa-CC) were allocated using the ‘based upon repeat pattern’ (BURP) algorithm that is implemented within the software47,48. spa types that were too short to presume ancestry (< 5 repeats within the hypervariable Xr region of the spa gene) were excluded from the BURP analysis. A phylogenetic tree of the identified spa types was constructed using the Molecular Evolutionary Genetic Analysis X software (MEGA X)49 using the neighbour-joining method50. The new spa type was submitted to the Ridom spaServer under accession number 17427851.
MLST
Eleven isolates representing the most common spa and SCCmec types were selected for MLST typing according to Enright et al.52. Briefly, seven housekeeping genes were targeted using their specific primers. The PCR products were Sanger sequenced and the consensus sequence of each gene as generated by the BioEdit Sequence Alignment software was identified as a specific allele type. The loci were combined and identified as ST types using PubMLST online (https://pubmlst.org/saureus/)53. A phylogenetic tree was constructed using PHYLOViZ software (http://www.phyloviz.net/) by UPGMA and eBURST54 to investigate the evolutionary history of the identified MLST types and the degree of relatedness. The eBURST algorithm method was applied using eBURST v3 online tool (http://eburst.mlst.net/default.asp).
Data availability
The datasets used and/or analyzed during the current study are available from the corresponding author on reasonable request.
Abbreviations
- AMUH:
-
Alexandria Main University Hospital
- BAL:
-
Broncho Alveolar Lavage
- CA:
-
Community Acquired
- CC:
-
Clonal Complex
- CLSI:
-
Clinical and Laboratory Standards Institute
- DLV:
-
Double Locus Variant
- DNA:
-
Deoxyribo Nucleic Acid
- HA:
-
Hospital Acquired
- MIC:
-
Minimum Inhibitory Concentration
- Min:
-
Minute
- MLST:
-
Multilocus sequence typing
- MOX:
-
Moxifloxacin
- MRSA:
-
Methicillin Resistant Staphylococcus aureus
- PBP2a:
-
Penicillin Binding Protein 2a
- PCR:
-
Polymerase Chain Reaction
- PFGE:
-
Pulsed Field Gel Electrophoresis
- S:
-
Second
- SCCmec :
-
Staphylococcal cassette chromosome mec
- SLV:
-
Single Locus Variant
- spa :
-
Staphylococcus protein A
- TLV:
-
Triple Locus Variant
- UPGMA:
-
Unweighted Pair Group Method with Arithmetic Mean method
References
Lowy, F. D. Staphylococcus aureus infections. N. Engl. J. Med. 339, 520–532. https://doi.org/10.1056/NEJM199808203390806 (1998).
Eriksen, K. R. “Celbenin”-resistant staphylococci. Ugeskr. Laeger 123, 384–386 (1961).
Herold, B. C. et al. Community-acquired methicillin-resistant Staphylococcus aureus in children with no identified predisposing risk. JAMA 279, 593–598. https://doi.org/10.1001/jama.279.8.593 (1998).
Planet, P. J., Diaz, L., Rios, R. & Arias, C. A. Global spread of the community-associated methicillin-resistant Staphylococcus aureus USA300 Latin American variant. J. Infect. Dis. 214, 1609–1610. https://doi.org/10.1093/infdis/jiw418 (2016).
Borg, M. A. et al. Prevalence of methicillin-resistant Staphylococcus aureus (MRSA) in invasive isolates from southern and eastern Mediterranean countries. J. Antimicrob. Chemother. 60, 1310–1315 (2007).
Falagas, M. E., Karageorgopoulos, D. E., Leptidis, J. & Korbila, I. P. MRSA in Africa: filling the global map of antimicrobial resistance. PLoS ONE 8, e68024. https://doi.org/10.1371/journal.pone.0068024 (2013).
Chambers, H. F. Methicillin resistance in staphylococci: molecular and biochemical basis and clinical implications. Clin. Microbiol. Rev. 10, 781–791 (1997).
Stapleton, P. D. & Taylor, P. W. Methicillin resistance in Staphylococcus aureus: mechanisms and modulation. Sci. Prog. 85, 57–72 (2002).
Katayama, Y., Ito, T. & Hiramatsu, K. A new class of genetic element, staphylococcus cassette chromosome mec, encodes methicillin resistance in Staphylococcus aureus. Antimicrob. Agents Chemother. 44, 1549–1555 (2000).
Livermore, D. M. Antibiotic resistance in staphylococci. Int. J. Antimicrob. Agents 16(Suppl 1), S3-10 (2000).
Liu, C. et al. Clinical practice guidelines by the Infectious Diseases Society of America for the treatment of methicillin-resistant Staphylococcus aureus infections in adults and children. Clin. Infect. Dis. 52, e18-55. https://doi.org/10.1093/cid/ciq146 (2011).
Butt, T., Ahmad, R. N. & Usman, M. Potential role of moxifloxacin in methicillin-resistant Staphylococcus aureus (MRSA) infections. J. Pak. Med. Assoc. 54, 42 (2004).
Tenover, F. C., Biddle, J. W. & Lancaster, M. V. Increasing resistance to vancomycin and other glycopeptides in Staphylococcus aureus. Emerg. Infect. Dis. 7, 327–332. https://doi.org/10.3201/eid0702.700327 (2001).
Chang, V. S., Dhaliwal, D. K., Raju, L. & Kowalski, R. P. Antibiotic resistance in the treatment of Staphylococcus aureus keratitis: a 20-year review. Cornea 34, 698–703. https://doi.org/10.1097/ICO.0000000000000431 (2015).
Entenza, J. M., Que, Y. A., Vouillamoz, J., Glauser, M. P. & Moreillon, P. Efficacies of moxifloxacin, ciprofloxacin, and vancomycin against experimental endocarditis due to methicillin-resistant Staphylococcus aureus expressing various degrees of ciprofloxacin resistance. Antimicrob. Agents Chemother. 45, 3076–3083. https://doi.org/10.1128/AAC.45.11.3076-3083.2001 (2001).
Lemaire, S. et al. Activity of moxifloxacin against intracellular community-acquired methicillin-resistant Staphylococcus aureus: comparison with clindamycin, linezolid and co-trimoxazole and attempt at defining an intracellular susceptibility breakpoint. J. Antimicrob. Chemother. 66, 596–607. https://doi.org/10.1093/jac/dkq478 (2011).
Vola, M. E. et al. Prevalence and antibiotic susceptibility of methicillin-resistant Staphylococcus aureus in ocular infections. Arq. Bras. Oftalmol. 76, 350–353 (2013).
Ashour, H. M. & el-Sharif, A. Microbial spectrum and antibiotic susceptibility profile of gram-positive aerobic bacteria isolated from cancer patients. J. Clin. Oncol. 25, 5763–5769 (2007).
Agmy, G. et al. Bacterial profile, antibiotic sensitivity and resistance of lower respiratory tract infections in Upper Egypt. Mediterr. J. Hematol. Infect. Dis. 5, e2013056 (2013).
El-Sokkary, R. H. et al. Community acquired pneumonia among adult patients at an Egyptian university hospital: bacterial etiology, susceptibility profile and evaluation of the response to initial empiric antibiotic therapy. Infect. Drug Resist. 11, 2141–2150 (2018).
Foxman, B., Zhang, L., Koopman, J. S., Manning, S. D. & Marrs, C. F. Choosing an appropriate bacterial typing technique for epidemiologic studies. Epidemiol. Perspect. Innov. 2, 10. https://doi.org/10.1186/1742-5573-2-10 (2005).
Li, L., Yeaman, M. R., Bayer, A. S. & Xiong, Y. Q. Phenotypic and genotypic characteristics of methicillin-resistant Staphylococcus aureus (MRSA) related to persistent endovascular infection. Antibiotics (Basel) https://doi.org/10.3390/antibiotics8020071 (2019).
Xiong, Y. Q. et al. Phenotypic and genotypic characteristics of persistent methicillin-resistant Staphylococcus aureus bacteremia in vitro and in an experimental endocarditis model. J. Infect. Dis. 199, 201–208. https://doi.org/10.1086/595738 (2009).
Rodriguez, M. et al. Discriminatory indices of typing methods for epidemiologic analysis of contemporary Staphylococcus aureus strains. Medicine (Baltimore) 94, e1534. https://doi.org/10.1097/MD.0000000000001534 (2015).
Asadollahi, P. et al. Distribution of the most prevalent spa types among clinical isolates of methicillin-resistant and -susceptible Staphylococcus aureus around the world: a review. Front. Microbiol. 9, 163 (2018).
Strommenger, B. et al. spa typing of Staphylococcus aureus as a frontline tool in epidemiological typing. J. Clin. Microbiol. 46, 574–581. https://doi.org/10.1128/JCM.01599-07 (2008).
Hassoun, A., Linden, P. K. & Friedman, B. Incidence, prevalence, and management of MRSA bacteremia across patient populations-a review of recent developments in MRSA management and treatment. Crit. Care 21, 211. https://doi.org/10.1186/s13054-017-1801-3 (2017).
van Hal, S. J. et al. Predictors of mortality in Staphylococcus aureus Bacteremia. Clin. Microbiol. Rev. 25, 362–386 (2012).
Hashem, R. A., Yassin, A. S., Zedan, H. H. & Amin, M. A. Fluoroquinolone resistant mechanisms in methicillin-resistant Staphylococcus aureus clinical isolates in Cairo, Egypt. J. Infect. Dev. Ctries. 7, 796–803. https://doi.org/10.3855/jidc.3105 (2013).
Mohamed, N. M., Zakaria, A. S., Edward, E. A. & Abdel-Bary, A. In vitro and in vivo activity of zabofloxacin and other fluoroquinolones against MRSA isolates from a university hospital in Egypt. Pol. J. Microbiol. 68, 59–69. https://doi.org/10.21307/pjm-2019-007 (2019).
Japoni, A. et al. Characterization of SCCmec types and antibacterial susceptibility patterns of methicillin-resistant Staphylococcus aureus in Southern Iran. Jpn. J. Infect. Dis. 64, 28–33 (2011).
Shittu, A. O. et al. Antibiotic resistance and molecular epidemiology of Staphylococcus aureus in Nigeria. BMC Microbiol. 11, 92. https://doi.org/10.1186/1471-2180-11-92 (2011).
Zhang, K., McClure, J.-A., Elsayed, S., Louie, T. & Conly, J. M. Novel multiplex PCR assay for characterization and concomitant subtyping of staphylococcal cassette chromosome types I to V in methicillin-resistant Staphylococcus aureus. J. Clin. Microbiol. 43, 5026–5033. https://doi.org/10.1128/jcm.43.10.5026-5033.2005 (2005).
Arakere, G. et al. Genotyping of methicillin-resistant Staphylococcus aureus strains from two hospitals in Bangalore, South India. J. Clin. Microbiol. 43, 3198–3202. https://doi.org/10.1128/JCM.43.7.3198-3202.2005 (2005).
Abou Shady, H. M., Bakr, A. E. A., Hashad, M. E. & Alzohairy, M. A. Staphylococcus aureus nasal carriage among outpatients attending primary health care centers: a comparative study of two cities in Saudi Arabia and Egypt. Braz. J. Infect. Dis. 19, 68–76. https://doi.org/10.1016/j.bjid.2014.09.005 (2015).
Abdulgader, S. M., Shittu, A. O., Nicol, M. P. & Kaba, M. Molecular epidemiology of Methicillin-resistant Staphylococcus aureus in Africa: a systematic review. Front. Microbiol. 6, 348. https://doi.org/10.3389/fmicb.2015.00348 (2015).
Kilic, A., Li, H., Stratton, C. W. & Tang, Y. W. Antimicrobial susceptibility patterns and staphylococcal cassette chromosome mec types of, as well as Panton-Valentine leukocidin occurrence among, methicillin-resistant Staphylococcus aureus isolates from children and adults in middle Tennessee. J. Clin. Microbiol. 44, 4436–4440. https://doi.org/10.1128/JCM.01546-06 (2006).
Franco, A. et al. Molecular characterization of spa type t127, sequence type 1 methicillin-resistant Staphylococcus aureus from pigs. J. Antimicrob. Chemother. 66, 1231–1235. https://doi.org/10.1093/jac/dkr115 (2011).
Feltrin, F. et al. A livestock-associated, multidrug-resistant, methicillin-resistant Staphylococcus aureus clonal complex 97 lineage spreading in dairy cattle and pigs in Italy. Appl. Environ. Microbiol. 82, 816–821. https://doi.org/10.1128/AEM.02854-15 (2016).
Kateete, D. P. et al. Identification of Staphylococcus aureus: DNase and Mannitol salt agar improve the efficiency of the tube coagulase test. Ann. Clin. Microbiol. Antimicrob. 9, 23. https://doi.org/10.1186/1476-0711-9-23 (2010).
CLSI. M100-S25 performance standards for antimicrobial susceptibility testing. (2015).
Pui, C. F. et al. Multiplex PCR for the concurrent detection and differentiation of Salmonella spp., Salmonella typhi and Salmonella typhimurium. Trop. Med. Health 39, 9–15 (2011).
Milheirico, C., Oliveira, D. C. & de Lencastre, H. Update to the multiplex PCR strategy for assignment of mec element types in Staphylococcus aureus. Antimicrob. Agents Chemother. 51, 3374–3377. https://doi.org/10.1128/AAC.00275-07 (2007).
Lina, G. et al. Bacterial competition for human nasal cavity colonization: role of staphylococcal agr alleles. Appl. Environ. Microbiol. 69, 18–23 (2003).
Oliveira, D. C. & de Lencastre, H. Multiplex PCR strategy for rapid identification of structural types and variants of the mec element in methicillin-resistant Staphylococcus aureus. Antimicrob. Agents Chemother. 46, 2155–2161. https://doi.org/10.1128/aac.46.7.2155-2161.2002 (2002).
Ito, T. et al. Novel type V staphylococcal cassette chromosome mec driven by a novel cassette chromosome recombinase, ccrC. Antimicrob. Agents Chemother. 48, 2637–2651. https://doi.org/10.1128/AAC.48.7.2637-2651.2004 (2004).
Harmsen, D. et al. Typing of methicillin-resistant Staphylococcus aureus in a university hospital setting by using novel software for spa repeat determination and database management. J. Clin. Microbiol. 41, 5442–5448 (2003).
Strommenger, B. et al. Assignment of Staphylococcus isolates to groups by spa typing, SmaI macrorestriction analysis, and multilocus sequence typing. J. Clin. Microbiol. 44, 2533–2540. https://doi.org/10.1128/JCM.00420-06 (2006).
Kumar, S., Stecher, G., Li, M., Knyaz, C. & Tamura, K. MEGA X: molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 35, 1547–1549. https://doi.org/10.1093/molbev/msy096 (2018).
Saitou, N. & Nei, M. The neighbor-joining method: a new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 4, 406–425. https://doi.org/10.1093/oxfordjournals.molbev.a040454 (1987).
Ridom, Ridom SpaServer https://spaserver.ridom.de/ (2016).
Enright, M. C., Day, N. P., Davies, C. E., Peacock, S. J. & Spratt, B. G. Multilocus sequence typing for characterization of methicillin-resistant and methicillin-susceptible clones of Staphylococcus aureus. J. Clin. Microbiol. 38, 1008–1015 (2000).
Jolley, K. A., Bray, J. E. & Maiden, M. C. J. Open-access bacterial population genomics: BIGSdb software, the PubMLST.org website and their applications. Wellcome Open Res. 3, 124. https://doi.org/10.12688/wellcomeopenres.14826.1 (2018).
Feil, E. J., Li, B. C., Aanensen, D. M., Hanage, W. P. & Spratt, B. G. eBURST: inferring patterns of evolutionary descent among clusters of related bacterial genotypes from multilocus sequence typing data. J. Bacteriol. 186, 1518–1530 (2004).
Acknowledgements
The authors would like to thank PD Dr. med. Wilma Ziebuhr for her helpful insights. This work was supported by the Egyptian Science and Technology Development Fund (STDF) [grant number 15135], The Ministry for Higher Education and Research Mobility grant 26-3-14 and the DFG- German Research Foundation grant ZI 665/3-1. The funding bodies had no role in the study design and collection, analysis and interpretation of data or in the writing of the manuscript.
Author information
Authors and Affiliations
Contributions
M.A. performed the experiments, participated in data analysis and contributed to manuscript writing. M.N. Participated in the genotyping experiments and data analysis. A.K. supervised the performance of the experimental work and reviewed the manuscript. M.E. participated in study design and reviewed the manuscript. A.W. designed the study, participated in data analysis and critically reviewed the manuscript. A.A. designed the study, analyzed the results and actively participated in manuscript writing. All authors revised the manuscript and approved the final form.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Alseqely, M., Newton-Foot, M., Khalil, A. et al. Association between fluoroquinolone resistance and MRSA genotype in Alexandria, Egypt. Sci Rep 11, 4253 (2021). https://doi.org/10.1038/s41598-021-83578-2
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-021-83578-2
This article is cited by
-
Unveilling genetic profiles and correlations of biofilm-associated genes, quorum sensing, and antibiotic resistance in Staphylococcus aureus isolated from a Malaysian Teaching Hospital
European Journal of Medical Research (2024)
-
Knowledge, attitudes and behaviour of Egyptians towards antibiotic use in the community: can we do better?
Antimicrobial Resistance & Infection Control (2023)
-
Recombination-mediated dissemination of Methicillin-resistant S. aureus clonal complex 1 in the Egyptian health care settings
Annals of Clinical Microbiology and Antimicrobials (2023)
-
Panton-Valentine Leukocidin (PVL) genes may not be a reliable marker for community-acquired MRSA in the Dakahlia Governorate, Egypt
BMC Microbiology (2023)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
