
Abstract
Natural disturbances are an important driver of population dynamics. Because it is difficult to observe wildlife during these events, our understanding of the strategies that species use to survive these disturbances is limited. On October 10, 2018, Hurricane Michael made landfall on Florida’s northwest coast. Using satellite and acoustic telemetry, we documented movements of 6 individual turtles: one loggerhead sea turtle, one Kemp’s ridley sea turtle, three green sea turtles and one diamondback terrapin, in a coastal bay located less than 30 km from hurricane landfall. Post-storm survival was confirmed for all but the Kemp’s ridley; the final condition of that individual remains unknown. No obvious movements were observed for the remaining turtles however the loggerhead used a larger home range in the week after the storm. This study highlights the resiliency of turtles in response to extreme weather conditions. However, long-term impacts to these species from habitat changes post-hurricane are unknown.
Similar content being viewed by others
Introduction
Natural disturbances can impact animal populations in many ways including altering community structure1, displacing individuals2, reducing reproductive output3, lowering individual fitness4, and changing movement patterns5. In addition, these events can alter habitat and environmental conditions, which can create suboptimal conditions for organisms. Because it is difficult to observe wildlife during these events, our understanding of the strategies that species use to survive or mitigate these disturbances is limited6,7,8,9,10. This knowledge gap hampers our ability to identify optimal conditions required by imperiled species for survival during extreme events.
On October 10, 2018, Hurricane Michael made landfall near Tyndall Air Force Base on Florida’s northwest coast. As a Category 5 storm with maximum winds greater than 140 kts, Hurricane Michael was the third most intense storm to make landfall in the contiguous United States11. Storm surge was estimated at 9–14 ft in Bay and Gulf Counties, Florida11 and the combination of water and wind resulted in the destruction of man-made structures and natural habitats. An estimated 39% of longleaf pine habitat in Florida showed immediate impacts from the storm12 and salinity of wetlands in the region was altered13. Marine habitats may also have been impacted, which could have negative consequences for the species that rely on them for survival14,15.
Northwest Florida’s estuarine and marine systems are rich in natural resources. The dominant feature of the region is the Apalachicola River, which has the greatest volume of all Florida’s rivers16. The nutrients deposited by the river sustain a high biological diversity throughout the region including the highest species density of amphibians and reptiles on the continent north of Mexico17. These reptiles include a variety of turtle species that inhabit estuarine and marine habitats such as diamondback terrapins (Malaclemys terrapin) and loggerhead (Caretta caretta), Kemp’s ridley (Lepidochelys kempii) and green sea turtles (Chelonia mydas).
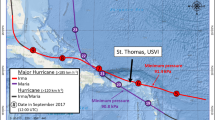
This area is also susceptible to hurricane impacts. Since hurricane records have been kept, the coasts of Mississippi, Alabama and Northwest Florida have had direct hits by more than one-third of the 273 storms that have affected the US mainland18. Hurricanes can displace freshwater and terrestrial reptiles19,20 and cause direct mortality of many species20,21,22. Understanding the conditions needed for species survival during hurricanes in this region is critical to population recovery. Using satellite and acoustic telemetry, we documented movements of four turtle species as Hurricane Michael moved through Northwest Florida (Fig. 1).

Hurricane Michael made landfall on October 10, 2018 approximately 15 km west of St. Joseph Bay, Florida where marine and estuarine turtles were being tracked using satellite and acoustic telemetry (hurricane data from Bevan et al. 2019). This map (with self-created symbols/shapes/text) was created using ESRI (Environmental Systems Resource Institute; http://www.esri.com/software/arcgis) ArcMap software, version 10.7.1. Basemap sources include: ESRI, Maxar, GeoEye, Earthstar Geographics, CNES/Airbus DS, USDA, USGS, AeroGRID, IGN, and the GIS User Community. Hurricane data contributed by Beven et al. (2019).
Results
Hurricane Michael made landfall approximately 30 km west of SJB on October 10, 2018 at approximately 13:30 local time [Eastern Daylight Time (ET); 17:30 UTC; Fig. 1]. Barometric pressure at landfall was estimated at 919 mb and maximum sustained winds were from the southwest (from approximately 210°) at 140 kts11. The National Weather Service suggested the storm was still intensifying as it made landfall11. Barometric pressure dropped from 1133 mb on October 1 to 1020 mb on October 2 and then again to 1014 mb on October 8 as the storm entered the southern GoM11 (see Fig. 1). One loggerhead (July 27, 2018), one Kemp’s ridley (September 26, 2018) and one diamondback terrapin (July 10, 2018) were captured and satellite tagged in SJB prior to the hurricane (Table 1). All three individuals were carrying active satellite tags during the storm. In addition, three green turtles were captured and tagged on October 4, 2018 and were carrying active acoustic tags during the storm (Table 1). Using size as an estimator of life-stage23,24,25, the loggerhead was a large juvenile, the Kemp’s ridley was either a large juvenile or an adult female, and the green turtles were juveniles. The terrapin was an adult female. Because terrapins are highly exploited in the commercial pet trade26,27, and providing location data through publication of research results can direct poachers to capture sites28, we chose not to present specific location data for the terrapin tracked during this study. We believe that information is not necessary to understand the movement patterns of this individual during the storm.
The last location received for the Kemp’s ridley was on October 10, 2018 at 22:26 ET which is approximately 9.0 h after hurricane landfall (Table 1). The last location received from the terrapin was on December 3, 2018 which is 56 days after landfall. The loggerhead’s tag continued transmissions until July 2019 and the green turtles were still being detected > 6 months after hurricane landfall.
Locations during landfall
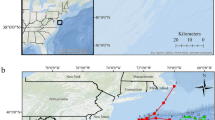
Locations were received from the loggerhead’s tag approximately 5 h before, 2 h before and 6 h after landfall (Fig. 2). The Kemp’s ridley tag transmitted approximately 21 h before and 6 h after landfall. Three additional locations were then received 7–9 h after landfall; those three locations represent the final transmissions from the tag. The terrapin tag transmitted 4 h before and 27 h after landfall.

Raw locations/detections for a Kemp’s ridley (a), loggerhead (b) and diamondback terrapin (c) that were carrying satellite tags and three green turtles (d–f) that were carrying acoustic tags in St. Joseph Bay, Florida when Hurricane Michael made landfall at approximately 14:00 h (EDT) on October 10, 2018 in Northwest Florida. The star represents the capture location (with capture date) for each turtle (except for the Kemp’s ridley which is marked by a pink circle). Triangles (regardless of color) show locations of acoustic receivers (R# is the receiver identifier). The location of receiver 3 (R3) is included in the Kemp’s ridley and loggerhead map for reference. Green circles represent locations/detections in the days prior to the storm; all previous locations (except filtered locations; see Methods) for the Kemp’s ridley are included. Red icons represent the last available location/detection for each turtle immediately prior to hurricane landfall. Blue icons represent the first available location/detection available for each turtle immediately after hurricane landfall. Yellow icons represent locations/detections in the day(s) after landfall occurred. Dates and times (in ET) for these locations/detections are also included. The arrow indicates wind direction during hurricane landfall. Because terrapins are an exploited species, and specific location data would reveal capture locations, we chose not to display these details on a satellite image. These maps (with self-created symbols/shapes/text) were created using ESRI (Environmental Systems Resource Institute; http://www.esri.com/software/arcgis) ArcMap software, version 10.7.1. Basemap sources for all Figures, except c (terrapin), include: Esri, Maxar, GeoEye, Earthstar Geographics, CNES/Airbus DS, USDA, USGS, AeroGRID, IGN, and the GIS User Community. The basemap source for Figure c is the Florida Fish and Wildlife Research Institute (https://myfwc.com/research/gis/). Hurricane data contributed by Beven et al. (2019).
Although tagged on October 4, the first detection of green turtle 4320’s tag was approximately 19 h after hurricane landfall (Fig. 2). Green turtle 4321’s tag was detected before hurricane landfall on October 6 and then approximately 23 h after landfall. Green turtle 4322’s tag was also detected before landfall on October 6 but was not detected again until 47 h (October 12) after landfall.
Home ranges
Home range centroids for the loggerhead, terrapin and green turtle 4321 for the week immediately after hurricane landfall (Oct 10) were located north or northeast of the turtle’s original capture location (Fig. 3). The centroid for green turtle 4322 was located southwest of the turtle’s capture location and green turtle 4320 had no data available for that week.

Weekly home range centroids for turtles carrying a satellite tag [loggerhead (a, b), terrapin (c); Minimum Convex Polygon] or an acoustic tag [green turtles (d–f); Kernel Density Estimation] during Hurricane Michael in St. Joseph Bay, Florida. Due to a lack of data, home range maps are not provided for the Kemp’s ridley. The storm made landfall on October 10, 2018; the centroid for the week of October 10 (October 10–16) is shown by a red dot. All weekly centroids for the loggerhead are shown on the left (a) and a close-up of loggerhead centroids on the right (red square; shown in b) immediately before, during and after the storm. Capture locations for each individual are denoted with a star. If weekly centroids are not included on the map, it is due to gaps in data. Because terrapins are an exploited species and providing exact locations for individuals may contribute to poaching, we chose not to display locations on satellite imagery. These maps (with self-created symbols/shapes/text) were created using ESRI (Environmental Systems Resource Institute; http://www.esri.com/software/arcgis) ArcMap software, version 10.7.1. Basemap sources for all Figures, except c (terrapin), include: Esri, Maxar, GeoEye, Earthstar Geographics, CNES/Airbus DS, USDA, USGS, AeroGRID, IGN, and the GIS User Community. The basemap source for Figure c is Florida Fish and Wildlife Conservation Commission-Fish and Wildlife Research Institute (https://myfwc.com/research/gis/).
Home range sizes varied among individuals (Table 2) with the largest home range exhibited by green turtle 4320 (mean 4.45; SD 4.330; range 0.004–11.180) and the smallest by the terrapin (mean 0.525; SD 0.867; range 0.003–3.191). Home range sizes also varied among weeks for each individual (Supplemental Figure 1). The loggerhead used a larger home range the week after the storm than in all of the previous weeks (but not subsequent weeks; see Discussion). For the remaining turtles, there were no obvious changes in home range sizes in the weeks immediately before or after Hurricane Michael (Table 2).
Discussion
The long-term effects of Hurricane Michael on Northwest Florida habitats are still unknown. Despite the record-breaking winds and extreme storm surge, tags on five of the six turtles tracked during landfall continued transmissions post-storm thereby confirming turtle survival. There are multiple strategies that turtles utilize to survive during and after extreme weather events including altering dive behavior, increasing home range size and shifting habitat use29,30,31. These studies have typically involved larger samples sizes30 or have documented long-distance movements31. The turtles tracked in SJB during Hurricane Michael showed no large-scale movements or changes in behavior in response to the storm. Individual variation in hurricane response for hawksbill sea turtles30, and other reptiles such as alligators32 has been documented. This includes some individuals that show no obvious response32. Our results suggest turtles in SJB responded little, at least with obvious and/or large-scale movements, to the storm both before and after landfall. Our small sample sizes, paired with limitations of non-GPS capable satellite tags in identifying fine-scale movements33 and the potential impacts of shallow and rough water on acoustic receiver detections34, make it difficult for us to confirm fine-scale changes in behavior. In addition, it is difficult to say if behaviors observed in this study (e.g. larger home range used by the loggerhead after the storm) were in response to the storm or some other factor. We recognize these limitations however, we agree with Sergio et al.35 that “it is ever more urgent and fundamental to increase our knowledge of species responses to disturbance, especially to predict potential future responses by endangered organisms”.
Kemp’s ridley
Because the satellite tag on the Kemp’s ridley stopped transmitting approximately 5 h after hurricane landfall, the fate of this individual is unknown. There are many reasons why satellite tags stop transmitting including tag malfunction (e.g. salt-water switch), battery life, antenna damage or animal mortality36. The satellite tag was deployed on the Kemp’s ridley in SJB only two weeks before the hurricane and outputs from the tag showed strong battery, no signs of biofouling, and no salt-water switch failures. When sea turtles die, they quickly sink37. The abrupt cessation of tag transmissions soon after landfall could indicate mortality or tag damage. Unfortunately, it is difficult to discriminate between those two possible events36.
Typically, juvenile Kemp’s ridleys inhabit nearshore, shallow waters38,39,40. From 2013 to 2017, seven juvenile Kemp’s ridleys were tracked in SJB; those individuals established home ranges that were within 1.5 km of shore and in relatively shallow water (mean 3.6 m). In the weeks prior to hurricane landfall, a Harmful Algal Bloom (HAB) of the toxic dinoflagellate Karenia brevis impacted the northwest Florida coast (https://oceanservice.noaa.gov/hazards/hab/florida-2018.html). Termed “red tide” events, these HAB’s cause disproportionate mortality to Kemp’s ridleys as compared to green turtles and loggerheads and exposure to brevetoxins can result in neurological impairment in sea turtles37,41. This HAB may have impacted the Kemp’s ridley’s behavior during our tracking period.
Loggerhead
The loggerhead we tracked during Hurricane Michael appeared to remain in relatively shallow water throughout the storm and was near the water’s surface within two hours of hurricane landfall. Loggerheads are strong swimmers and can maintain their position despite high winds and storm currents42,43,44,45 however our results suggest that even in Category 5 winds, loggerheads are able to not only survive but to remain in place in relatively shallow water.
In the week after the storm, the loggerhead used a larger home range that included deep water north of her capture location (see Supplemental Figure S1). Short-term (i.e. weekly) expansions in space-use following environmental perturbations have been documented for many species35 including hawksbill sea turtles30 and may reflect increased foraging effort required after disturbances or use of new resting locations30. The expanded area used the week after the storm was also used by the loggerhead during the weeks of November 14 and 21. At this time, the loggerhead began moving north and by the end of November, the turtle was out of SJB. These November movements were in response to falling temperatures, which have been previously documented for loggerheads in SJB46. Mean air temperatures [recorded at the National Oceanographic and Atmospheric Administration’s weather buoy (APCF1) located approximately 30 km east of SJB], dropped from 21.5 °C (Nov 8–14), to 17.6 °C (Nov 15–21) and then even further to 13.8 °C (Nov 22–28) during this time period. It is well reported that loggerheads use deep water as a thermal refuge46,47,48 and this expanded area may have also served as a post-storm refuge for the loggerhead immediately after Hurricane Michael.
Terrapin
The diamondback terrapin is the smallest of all species tracked during the hurricane however this individual did not appear to be impacted during the storm’s passage. While displacement of aquatic turtles, such as wood turtles (Glyptemys insculpta) and Johnstone River snapping turtles (Elseya irwini), has been documented during extreme flooding, the majority of individuals in these studies were not impacted by the floods20,49,50. It has been suggested that individuals bury themselves or remain on the bottom in deep water to avoid extreme surface currents20 and this may have been the strategy utilized by the terrapin in SJB. We received a high-quality location (LC 3) approximately 2.5 h before and then again 18 h after hurricane landfall, which suggests the terrapin was at the water’s surface or on land at those times. Many turtles, including terrapins, are able to remain submerged for long periods of time through aquatic respiration or regulation of metabolism51,52. It is likely the terrapin in SJB remained submerged or buried through hurricane landfall to avoid displacement from winds and currents.
The week after the storm however the terrapin’s home range centroid was approximately 1.5 km east (i.e. landward) of any other weekly centroid we calculated, but this did not correspond to an increase in home range size. Although reptiles are occasionally displaced by storms those dislocations are typically farther (i.e. tens of kilometers) than the small distance (~ 1.5 km) observed here2,19,49. Flooding from SJB eastward was severe during and immediately after Hurricane Michael11 and this may have pushed the terrapin landward. As flood waters receded, she could have returned to her usual location near her capture site where she was documented the following week (October 17). However, this small shift in centroid location may also simply reflect variability in Argos locations. Regardless of whether the terrapin moved after landfall, terrapins in SJB did not appear to suffer significant consequences from the storm. After several hurricanes impacted the Florida Keys, few mangrove terrapins (Malaclemys terrapin rhizophorarium) were captured and it took several years for terrapins to return53; whereas, 6 days after Hurricane Michael, we captured two adult male terrapins in SJB, and within three weeks we captured an additional 17 individuals, including 6 females.
Green turtles
None of the tagged green turtles were detected on any receivers for several days immediately before and during the storm. We had acoustic receivers placed throughout most of the southern end of the bay, although receivers were spaced far enough apart (ex: 1200 m in some cases) where gaps were available. Therefore, the tagged green turtles could have either remained in the southern end of the bay but in undetectable areas or moved out of the array during this time period. Unfortunately, we do not have the data to answer this question.
Both of the green turtles that were detected the day after the storm (October 11; 4320 and 4321) were located at the most northerly receiver (R3). Perhaps the green turtles moved north into deeper waters where they were undetected. Similar to loggerheads, green turtles in SJB use deep water as a thermal refuge during cold temperatures46,54 and these same areas may have also been used as a refuge during Hurricane Michael30. Again, without acoustic detections during the storm, it is impossible to know for sure how these individuals responded. After the storm passed, the turtles traveled back to their original capture location, with turtles 4321 and 4322 arriving by October 12 and turtle 4320 arriving by October 21.
Interestingly, Matley et al.30 reported that larger hawksbills did not shift habitat use after Hurricanes Irma and Maria. In our study, the green turtles (range 34.0–40.3 cm SCL) were much smaller than the loggerheads (76.5 cm SCL) however none of these individuals displayed obvious shifts in habitat use. Larger individuals may be more resilient to environmental perturbations30,55 due to increased energy reserves, competition or simple physical strength. For example, a 150 kg loggerhead may be less susceptible to dislocation from storm surge than a 6 kg green turtle. In SJB, not even the terrapin, which was the smallest turtle we tracked during the storm (0.98 kg), was clearly impacted by the storm. However, terrapins, unlike sea turtles, are able to bury themselves completely in response to environmental perturbations. It has been suggested that sea turtles bury themselves in sediment during periods of extreme cold56 but it is more likely that turtles simply rest on the seafloor for extended periods rather than actually bury themselves47,57. The ability of terrapins to utilize hibernacula as a refuge during extreme weather conditions most likely explains the lack of obvious impacts we observed for the small terrapin during Hurricane Michael.
Although we have confirmation that most of the turtles we tracked during the storm survived, long-term impacts from the hurricane may still affect these individuals20. Population declines in various species, including aquatic turtles, have been linked to habitat alterations caused by hurricanes58,59. Seagrass and salt marsh habitat used by loggerheads, Kemp’s ridleys, green turtles and terrapins may be altered by storms, although these impacts appear to be site- or storm-specific60,61. Seagrass habitat in Alabama was unaffected by Hurricane Katrina whereas habitat in the Florida Keys experienced significant damage from Hurricane Georges60. Challener et al.61 found no post-storm impacts to seagrass-associated, sea urchin populations in SJB immediately after Hurricane Michael and reported no evidence that the seagrass was impacted. The results of our opportunistic study highlight the resiliency of marine and estuarine turtles62,63. More research on hurricane response by marine and estuarine turtles is needed, particularly for Kemp’s ridleys as the individual in our study was the only turtle that may not have survived.
Methods
Turtle tagging
St. Joseph Bay (SJB), located in Northwest Florida in the northern Gulf of Mexico (GoM; Fig. 1), covers approximately 26,000 ha and is surrounded by mostly undeveloped land and the small town of Port St. Joe (population 350064). It has a mean depth of 7 m64. Seagrass beds cover approximately one-sixth of the bay (4000 ha) and are most abundant in the southern end64. Barometric pressure, wind speed and direction data were downloaded from NOAA C-MAN station (APCF1; https://www.ndbc.noaa.gov/station_page.php?station=APCF1) located approximately 32 km east of St. Joseph Bay.
Turtles were captured in SJB from July through October 2018 (Table 1) using a set net (green turtle) or by hand (terrapin, loggerhead, Kemp’s ridley). All sea turtles were individually marked with a metal Inconel tag placed along the trailing edge of each front flipper and a Passive Integrated Transponder (PIT) tag placed subcutaneously in the left shoulder. The terrapin received a PIT tag inserted behind the left bridge, and its carapace was notched. All turtles were measured using two methodologies: (1) straight carapace length (SCL) and width (SCW) using calipers and (2) curved carapace length (CCL; measured from the nuchal notch to the tip of the left pygial scute) and width (CCW) using a cloth tape measure. Turtles were also weighed using a hand-held Pesola (Kapuskasing, Ontario, Canada) spring scale.
All turtle capture, handling and sampling methods were carried out in accordance with relevant guidelines and regulations described by the National Marine Fisheries Service (permit #17183) and Florida Fish and Wildlife Conservation Commission (permits #094, #33447). All experimental protocols were approved by USGS Institutional Animal Care protocols (Protocol USGS/WARC/GNV 2019-15; USGS/WARC/GNV 2018-04).
Satellite tags were fitted to the anterior portion of the carapace using a cool-setting epoxy (Superbond; FCGE Fiberglass Coatings, Inc, St. Petersburg, Florida) and following established protocols65. The terrapin received a SPOT-387 tag, the loggerhead received SPOT-287 tag, and the Kemp’s ridley was carrying a SPLASH10 tag (Wildlife Computers, Redmond, WA). We ensured that the total weight of the satellite tag and epoxy on all turtles did not exceed 5% of the individual’s body weight. Prior to satellite tag attachment, we removed epibionts, sanded the attachment site and cleaned it with isopropyl alcohol. We streamlined the attachment materials to reduce drag on the turtle66. All tagged turtles were released at or near their capture location. Each tag was set to be active for 24 h d−1.
In July 2018, we placed nine VR2W acoustic receivers (Innovasea; Bedford, Nova Scotia, Canada) throughout the southern portion of SJB (see Fig. 2). Receivers were attached to permanent channel markers using plastic electrician zip-ties. Green turtles were captured on October 4, 2018 and fitted with V13 coded acoustic transmitters (Innovasea; 6 g in water; 13 mm diameter × 36 mm length) on the right rear marginal scute of the carapace. Acoustic transmitters were attached with stainless steel wire fitted through two 3-mm diameter holes drilled in the marginal scutes. Location of transmitters did not interfere with flipper movements. A small amount of epoxy was placed under and then over the transmitter for additional adhesion. Receivers were downloaded after the storm on December 5, 2018.
Data analysis
We calculated home ranges, along with home range centroids, for each turtle on a weekly basis. Weekly analyses were chosen based on data availability and following Matley et al.30. Satellite data were filtered by location class prior to analyses. We included location classes (LC) 3, 2, 1, 0, A, and B and rejected LC Z for which no error estimation was available. Argos assigns accuracy estimates of 250 m for LC 3, 250 to 500 m for LC 2, 500 to 1500 m for LC 1, and 1500 m for LC 067. The estimated accuracy is unknown for LCs A and B (but these LCs can be useful; see68,69), and locations failing the Argos plausibility tests are tagged as class LC Z. Kalman-filtering of location data was performed by Argos. Kalman-filtering algorithm provides more estimated positions and significantly improves position accuracy, most significantly for locations obtained in LCs A and B70.
We used State Space Modeling (SSM) approach to calculate home ranges (but not estimate behavioral state), following Hart et al.33. We applied a model used by Breed et al. 71, which is a modified version of a model described by Jonsen et al.72 that estimates model parameters by Markov Chain Monte Carlo (MCMC) using JAGS via the software program R. We fit the model to data for each individual turtle to estimate location every day from five independent and parallel chains of MCMC samples. Our samples from the posterior distribution were based on 3,000 samples after a burn-in of 7000 samples and thinning the remaining samples by twenty. If convergence wasn’t reached, we increased the sample size to 6,000. We used the psrf < 1.1 as a measure of convergence.
Home ranges were calculated for all individuals using the SSM location data during each week using Multiple Convex Polygon (MCP)70,73 or Kernel Density Estimation (KDE)72,74 in R (package adehabitatHR75) depending on data availability. For individuals that had between 5 and 19 locations MCP was used whereas KDE was used for individuals with ≥ 20 locations. Weeks were defined based on hurricane landfall (October 10, 2018 at approximately 13:30 EDT). Hence the first week before the hurricane ranged from October 3 to October 9 and all preceding and subsequent weeks followed (e.g. week after the hurricane October 10–October 16).
In ArcMap (v10.7.1) we plotted turtle locations to document movements in the days surrounding hurricane landfall. We plotted the following locations, depending on data availability: (1) the day(s) prior to landfall, (2) the most recent location immediately before landfall, (3) the most recent location immediately after landfall and (4) the day(s) after landfall. For the Kemp’s ridley, we plotted all available locations after filtering out obvious erroneous locations. Erroneous locations were defined as those that were > 2 km into the GoM or on land76.
We calculated weekly home range size (km2) and home range centroids for the temporal extent possible for each individual. We mapped centroid locations for the week immediately prior to landfall (October 3) through November 28. If data were not available for a turtle for that entire time period, we used the temporal extent of available data. We chose November 28 as the cut off for mapping centroid locations because sea turtles in SJB typically move out of the bay and into deeper waters in the GoM in mid to late November46. Terrapins also become less active and presumably enter hibernacula when temperatures drop77 although data on over-wintering behavior by terrapins in Northwest Florida are lacking.
References
Elliser, C. R. & Herzing, D. L. Replacement dolphins? Social restructuring of a resident pod of Atlantic bottlenose dolphins, Tursiops truncatus, after two major hurricanes. Mar. Mammal Sci. 27, 39–59 (2011).
Elsey, R. M. & Aldrich, C. Long-distance displacement of a juvenile alligator by Hurricane Ike. Southeast. Nat. 8, 746–749 (2009).
Jones, J., DeBruyn, R. D., Barg, J. J. & Robertson, R. J. Assessing the effects of natural disturbance on a Neotropical migrant songbird. Ecology 82, 2628–2635 (2001).
Swilling, W. R., Wooten, M. C., Holler, N. R. & Lynn, W. J. Population dynamics of Alabama beach mice (Peromyscus polionotus ammobates) following Hurricane Opal. Am. Midl. Nat. 140, 287–298 (1998).
Heupel, M. R., Simpfendorfer, C. A. & Hueter, R. E. Running before the storm: blacktip sharks respond to falling barometric pressure associated with Tropical Storm Gabrielle. J. Fish Biol. 63, 1357–1363 (2003).
Cannizzo, Z. J. & Griffen, B. D. Habitat-specific impacts of Hurricane Matthew on a range-expanding species. Hydrobiologia 809, 79–89 (2018).
Gunzburger, M. S., Hughes, W. B., Barichivich, W. J. & Staiger, J. S. Hurricane storm surge and amphibian communities in coastal wetlands of northwestern Florida. Wetl. Ecol. Manag. 18, 651–663 (2010).
Lawes, M. J., Adie, H., Russell-Smith, J., Murphy, B. & Midgley, J. J. How do small savanna trees avoid stem mortality by fire? the roles of stem diameter, height and bark thickness. Ecosphere 2, 1–13 (2011).
Lytle, D. A., Bogan, M. T. & Finn, D. S. Evolution of aquatic insect behaviours across a gradient of disturbance predictability. Proc. R. Soc. B Biol. Sci. 275, 453–462 (2008).
Schwilk, D. W. & Ackerly, D. D. Flammability and serotiny as strategies: correlated evolution in pines. Oikos 94, 326–336 (2001).
Beven II, J. L., Berg, R. & Hagen, A. National hurricane center tropical cyclone report: Hurricane Michael (AL142018). Natl. Weather Serv. 111 (2019). https://www.nhc.noaa.gov/data/tcr/AL142018_Michael.pdf. Accessed 12 October 2019.
Zampieri, N. E., Pau, S. & Okamoto, D. K. The impact of Hurricane Michael on longleaf pine habitats in Florida. bioRxiv https://doi.org/10.1101/736629 (2019).
Walls, S. C. et al. Seeking shelter from the storm: conservation and management of imperiled species in a changing climate. Ecol. Evol. 9, 7122–7133 (2019).
Congdon, V. M., Bonsell, C., Cuddy, M. R. & Dunton, K. H. In the wake of a major hurricane: differential effects on early vs. late successional seagrass species. Limnol. Oceanogr. Lett. 4, 155–163 (2019).
Edmiston, H. L. et al. Tropical storm and hurricane impacts on a Gulf Coast Estuary: Apalachicola Bay, Florida. J. Coast. Res. 10055, 38–49 (2008).
Livingston, R. J., Hackney, C. T., Adams, S. M. & Martin, W. Medium-sized rivers of the Gulf Coastal Plain in biodiversity of the Southeastern United States. In Biodiversity of the Southeastern United States (eds Hackney, C. T. et al.) 351–385 (Wiley, New York, 1992).
Couch, C. A., Hopkins, E. H. & Hardy, P. S. Influences of environmental settings on aquatic ecosystems in the Chattahoochee-Flint River Basin. Water Resour. Investig. Rep. 95–4278, 65 (1996).
Blake, E. S., Rappaport, E. N., Landsea, C. W. & Miami, N. The deadliest, costliest, and most intense United States tropical cyclones from 1851 to 2006 (and other frequently requested hurricane facts)-technical memorandum NWS TPC-5. Natl. Ocean. Atmos. Adm. (NOAA)—National Hurric. Cent. (NHC), US Dep. Commer. Memo. NWS TPC-5 54, 43 (2007).
Censky, E. J., Hodge, K. & Dudley, J. Over-water dispersal of lizards due to hurricanes [7]. Nature 395, 556 (1998).
Jones, M. T. & Sievert, P. R. Effects of stochastic flood disturbance on adult wood turtles, Glyptemys insculpta, in Massachusetts. Can. Field Nat. 123, 313–322 (2009).
Massie, J. A. et al. Going Downriver: Patterns and cues in hurricane-driven movements of common snook in a subtropical coastal river. Estuaries Coasts https://doi.org/10.1007/s12237-019-00617-y (2019).
Strickland, B. A. et al. Movements of juvenile bull sharks in response to a major hurricane within a tropical estuarine nursery area. Estuaries Coasts https://doi.org/10.1007/s12237-019-00600-7 (2019).
NMFS & USFWS. Recovery Plan for the Northwest Atlantic Population of the Loggerhead Sea Turtle (Second Revision). National Marine Fisheries Service (2008).
National Marine Fisheries Service, U.S. Fish and Wildlife Service & SEMARNAT. Bi-National Recovery Plan for the Kemp’s Ridley Sea Turtle (Lepidochelys kempii), Second Revision. Fisheries (Bethesda) (2011).
Seminoff, J. A. et al. Status review of the green turtle (Chelonia mydas) under the Endangered Species Act. (2015).
Convention on International Trade in Endangered Species of Wild Fauna and Flora. Draft proposal of prohibiting the commercial trade of diamondback terrapin. (2013).
Enge, K. M. et al. What the world needs is a better gopher tortoise trap. Herpetol. Rev. 48, 543–576 (2012).
Powell, R. & Henderson, R. W. A new species of Gonatodes (Squamata: Gekkonidae) from the West Indies. Caribb. J. Sci. 41, 709–715 (2005).
Wilson, M., Tucker, A. D., Beedholm, K. & Mann, D. A. Changes of loggerhead turtle (Caretta caretta) dive behavior associated with tropical storm passage during the inter-nesting period. J. Exp. Biol. 220, 3432–3441 (2017).
Matley, J. K., Eanes, S., Nemeth, R. S. & Jobsis, P. D. Vulnerability of sea turtles and fishes in response to two catastrophic Caribbean hurricanes, Irma and Maria. Sci. Rep. 9, 1–15 (2019).
Crowe, L. M., Hatch, J. M., Patel, S. H., Smolowitz, R. J. & Haas, H. L. Riders on the storm: loggerhead sea turtles detect and respond to a major hurricane in the Northwest Atlantic Ocean. Mov. Ecol. 8, 1–13 (2020).
Strickland, B. A. et al. Variation in movement behavior of alligators after a major hurricane. Anim. Biotelem. 8, 1–10 (2020).
Hart, K. M., Lamont, M. M., Fujisaki, I., Tucker, A. D. & Carthy, R. R. Common coastal foraging areas for loggerheads in the Gulf of Mexico: opportunities for marine conservation. Biol. Conserv. 145, 185–194 (2012).
Swadling, D. S. et al. Seagrass canopies and the performance of acoustic telemetry: implications for the interpretation of fish movements. Anim. Biotelem. 8, 1–12 (2020).
Sergio, F., Blas, J. & Hiraldo, F. Animal responses to natural disturbance and climate extremes: a review. Glob. Planet. Change 161, 28–40 (2018).
Hays, G. C., Bradshaw, C. J. A., James, M. C., Lovell, P. & Sims, D. W. Why do Argos satellite tags deployed on marine animals stop transmitting?. J. Exp. Mar. Bio. Ecol. 349, 52–60 (2007).
Foley, A. M. et al. Assessing Karenia brevis red tide as a mortality factor of sea turtles in Florida, USA. Dis. Aquat. Organ. 132, 109–124 (2019).
Schmid, J. R. & Witzell, W. N. Seasonal migrations of immature Kemp’s ridley turtles along the west coast of Florida. Gulf Mex. Sci 24, 27–40 (2006).
Seney, E. E. & Landry, A. M. Movement patterns of immature and adult female Kemp’s ridley sea turtles in the northwestern Gulf of Mexico. Mar. Ecol. Prog. Ser. 440, 241–254 (2011).
Coleman, A. T., Pitchford, J. L., Bailey, H. & Solangi, M. Seasonal movements of immature Kemp’s ridley sea turtles (Lepidochelys kempii) in the northern Gulf of Mexico. Aquat. Conserv. Mar. Freshw. Ecosyst. 27, 253–267 (2017).
Fauquier, D. A. et al. Brevetoxin in blood, biological fluids, and tissues of sea turtles naturally exposed to Karenia brevis blooms in central West Florida. J. Zoo Wildl. Med. 44, 364–375 (2013).
Hays, G. C., Lusci, P., Papi, F., Seppia, P. & Marsh, R. Changes in behaviour during the inter-nesting and post-nesting migration for Ascension Island green turtles. Mar. Ecol. Prog. Ser. 189, 263–273 (1999).
Schofield, G. et al. Microhabitat selection by sea turtles in a dynamic thermal marine environment. J. Anim. Ecol. 78, 14–21 (2009).
Schofield, G. et al. Inter-annual variability in the home range of breeding turtles: Implications for current and future conservation management. Biol. Conserv. 143, 722–730 (2010).
Pike, D. A. Environmental correlates of nesting in loggerhead turtles, Caretta caretta. Anim. Behav. 76, 603–610 (2008).
Lamont, M. M. & Iverson, A. R. Shared habitat use by juveniles of three sea turtle species. Mar. Ecol. Prog. Ser. 606, 187–200 (2018).
Hochscheid, S., Bentivegna, F. & Hays, G. C. First records of dive durations for a hibernating sea turtle. Biol. Lett. 1, 82–86 (2005).
Hawkes, L. A., Broderick, A. C., Coyne, M. S., Godfrey, M. H. & Godley, B. J. Only some like it hot—quantifying the environmental niche of the loggerhead sea turtle. Divers. Distrib. 13, 447–457 (2007).
Johnston, G. R., Thomas, T. M., Suarez, E., Lau, A. & Mitchell, J. C. Population structure and body size of the suwannee alligator snapping turtle (Macrochelys suwanniensis) in Northern Florida. Chelonian Conserv. Biol. 14, 73–81 (2015).
Freeman, A. B., Eisemberg, C. C. & Stoetzel, H. Habitat use and movements in an upland population of Johnstone River Snapping Turtles, Elseya irwini. Herpetol. Conserv. Biol. 13, 464–472 (2018).
Jackson, D. C. & Ultsch, G. R. Physiology of hibernation under the ice by turtles and frogs. J. Exp. Zool. Part A Ecol. Genet. Physiol. 313 A, 311–327 (2010).
Williard, A. S. & Harden, L. A. Seasonal changes in thermal environment and metabolic enzyme activity in the diamondback terrapin (Malaclemys terrapin). Comp. Biochem. Physiol. A Mol. Integr. Physiol. 158, 477–484 (2011).
Mealey, B. K., Baldwin, J. D., Parks-Mealey, G. B., Bossart, G. D. & Forstner, M. R. J. Characteristics of mangrove diamondback terrapins (Malaclemys Terrapin Rhizophorarum) inhabiting altered and natural mangrove islands. J. N. Am. Herpetol. 2014, 76–80 (2014).
Lamont, M. M., Fujisaki, I., Stephens, B. S. & Hackett, C. Home range and habitat use of juvenile green turtles (Chelonia mydas) in the northern Gulf of Mexico. Anim. Biotelem. 3, 53 (2015).
Ozgul, A., Coulson, T., Reynolds, A., Cameron, T. C. & Benton, T. G. Population responses to perturbations: the importance of trait-based analysis illustrated through a microcosm experiment. Am. Nat. 179, 582–594 (2012).
Mendonça, M. T. Movements and feeding ecology of immature green turtles (Chelonia mydas) in a Florida Lagoon. Copeia 1983, 1013–1023 (1983).
Lamont, M. M., Seay, D. R. & Gault, K. Overwintering behavior of juvenile sea turtles at a temperate foraging ground. Ecology 99, 2621–2624 (2018).
Waide, R. Summary of the response of animal populations to hurricanes in the Caribbean. Biotropica 23, 508–512 (1991).
Selman, W. & Qualls, C. The impacts of Hurricane Katrina on a population of yellow-blotched sawbacks (Graptemys flavimaculata) in the Lower Pascagoula River. Herpetol. Conserv. Biol. 3, 224–230 (2008).
Fourqurean, J. W. & Rutten, L. M. The impact of Hurricane Georges on soft-bottom, back reef communities: site- and species-specific effects in south Florida seagrass beds. Bull. Mar. Sci. 75, 239–257 (2004).
Challener, R., McClintock, J. B., Czaja, R. & Pomory, C. Rapid assessment of post-Hurricane Michael impacts on a population of the sea urchin Lytechinus variegatus in seagrass beds of Eagle Harbor, Port Saint Joseph Bay, Florida. Gulf Caribb. Res. 30, SC11–SC16 (2019).
Jergenson, A. M., Miller, D. A. W., Neuman-Lee, L. A., Warner, D. A. & Janzen, F. J. Swimming against the tide: resilience of a riverine turtle to recurrent extreme environmental events. Biol. Lett. 10, 20130782 (2014).
Miller, D. A. W., Janzen, F. J., Fellers, G. M., Kleeman, P. M. & Bronikowski, A. M. Biodemography of ectothermic tetrapods provides insights into the evolution and plasticity of mortality patterns. Soc. Hierarchy Heal. Comp. Biodemogr. A Collect. Pap. 295, 314. https://doi.org/10.17226/18822 (2014).
Florida Department of Environmental Protection. St. Joseph Bay Aquatic Preserve Management Plan (2008).
National Oceanic and Atmospheric Administration. Southeast Fisheries Science Center Techniques Manual (2008).
Watson, K. P. & Granger, R. A. Hydrodynamic effect of a satellite transmitter on a juvenile green turtle (Chelonia mydas). J. Exp. Biol. 201, 2497–2505 (1998).
CLS. Argos user’s manual: worldwide tracking and environmental monitoring by satellite (2011).
Hays, G. C. et al. The importance of sand albedo for the thermal conditions on sea turtle nesting beaches. Oikos 93, 87–94 (2001).
Witt, M. J. et al. Assessing accuracy and utility of satellite-tracking data using Argos-linked Fastloc-GPS. Anim. Behav. 80, 571–581 (2010).
Lopez, R. & Malarde, J. Improving Argos doppler location using Kalman filtering (2011).
Breed, G. A., Jonsen, I. D., Myers, R. A., Don Bowen, W. & Leonard, M. L. Sex-specific, seasonal foraging tactics of adult grey seals (Halichoerus grypus) revealed by state-space analysis. Ecology 90, 3209–3221 (2009).
Jonsen, I. D., Flemming, J. M. & Myers, R. A. Robust state-space modeling of animal movement data. Ecology 86, 2874–2880 (2005).
Kalman, R. E. A new approach to linear filtering and prediction problems. J. Basic Eng. 82, 35–45 (1960).
Patterson, T., Thomas, L., Wilcox, C., Ovaskainen, O. & Matthiopoulos, J. State-space models of individual animal movement. Trends Ecol. Evol. 23, 87–94 (2008).
Calenge, C. The package adehabitat for the R software: tool for the analysis of space and habitat use by animals. Ecol. Modell. 197, 1035–1041 (2006).
Hart, K. M., Lamont, M. M., Sartain, A. R. & Fujisaki, I. Migration, foraging, and residency patterns for Northern Gulf loggerheads: implications of local threats and international movements. PLoS One 9, e103453 (2014).
Harden, L. A., Midway, S. R. & Williard, A. S. The blood biochemistry of overwintering diamondback terrapins (Malaclemys terrapin). J. Exp. Mar. Biol. Ecol. 466, 34–41 (2015).
Acknowledgements
We thank Joseph Alday and Carson Arends for field assistance. The research was conducted under State of Florida Marine Turtle Permit #094 and Special Use Permit #33447, and National Marine Fisheries Service Permit #17183. All turtle handling and sampling was performed according to the Institutional Animal Care Protocol USGS/WARC/GNV 2019-15 and USGS/WARC/GNV 2018-04. Maps throughout this paper were created using ArcGIS software by Esri. For more information about Esri software, please visit www.esri.com. Any use of trade, product, or firm names is for descriptive purposes only and does not imply endorsement by the US Government.
Author information
Authors and Affiliations
Contributions
M.M.L. developed the concept, assisted with field work and lead the writing of the manuscript. D.J. conducted statistical analyses, made figures, and assisted with manuscript writing. D.J.C. conducted field work and assisted with manuscript writing. All authors reviewed and approved the manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Disclosure: Any use of trade, product, or firm names is for descriptive purposes only and does not imply endorsement by the US Government. This draft manuscript is distributed solely for purposes of scientific peer review. Its content is deliberative and pre-decisional, so it must not be disclosed or released by reviewers. Because the manuscript has not yet been approved for publication by the US Geological Survey (USGS), it does not represent any official USGS finding or policy.
Supplementary Information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Lamont, M.M., Johnson, D. & Catizone, D.J. Movements of marine and estuarine turtles during Hurricane Michael. Sci Rep 11, 1577 (2021). https://doi.org/10.1038/s41598-021-81234-3
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-021-81234-3
This article is cited by
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
