
Abstract
Sources of particulate organic carbon (POC) play important roles in aqueous carbon cycling because internal production can provide labile material that can easily be turned into CO2. On the other hand, more recalcitrant external POC inputs can cause increased loads to sedimentary organic matter that may ultimately cause CH4 release. In order to differentiate sources, stable isotopes offer a useful tool. We present a study on the Itupararanga Reservoir (Brazil) where origins of POC were explored by comparing its isotope ratios (δ13CPOC) to those of dissolved inorganic carbon (δ13CDIC). The δ13CPOC averaged around − 25.1‰ in near-surface waters, which indicates higher primary production inferred from a fractionation model that takes into account carbon transfer with a combined evaluation of δ13CPOC, δ13CDIC and aqueous CO2. However, δ13CPOC values for water depths from 3 to 15 m decreased to − 35.6‰ and indicated different carbon sources. Accordingly, the δ13CDIC values of the reservoir averaged around + 0.6‰ in the top 3 m of the water column. This indicates CO2 degassing and photosynthesis. Below this depth, DIC isotope values of as low as − 10.1‰ showed stronger influences of respiration. A fractionation model with both isotope parameters revealed that 24% of the POC in the reservoir originated from detritus outside the reservoir and 76% of it was produced internally by aqueous CO2 fixation.
Similar content being viewed by others


Introduction
Reservoirs and their tributaries are active components of landscapes. They receive, transport, process and store inorganic and organic carbon1. Although lakes and reservoirs cover only 2.2% of the global continental area, they may play so far poorly accounted roles in cycling continental carbon2,3,4. In addition, regional investigations of carbon budgets become increasingly important for managing water resources because they define ecosystem functions and services5,6,7,8.
Spatiotemporal changes of physicochemical and biological parameters in subtropical reservoirs, can increase our knowledge of carbon turnover in water bodies. For instance, excessive precipitation, can increase terrestrial nutrients, contaminants, soil leaching and additions of untreated sewage input9. Rainfall events can also cause near-surface turbulences in lakes that has been shown to enhance gas exchange by increasing transfer velocities of CO210,11. Precipitation can also lead to increased input from external sources (i.e. allochthonous matter for instance from plant debris or soils) that may become trapped in lakes and reservoirs. This can, in turn, change biogeochemical dynamics of the water column12. Such processes include carbon uptake and release of carbon within the water column that may also be controlled by seasonal changes of temperature, weather patterns or light availability. These may in turn have important influences on carbon fluxes inside open water bodies such as for instance sedimentation, or gross primary production, ecosystem respiration and external carbon inputs13. For the latter, reservoir catchments may offer large but unknown contributions that may exceed internal (i.e. autochthonous) primary production14. On the other hand, autochthonous POC and sediment mineralization are strongly constrained15 and excess of this type of organic carbon can increase the emission of greenhouse gases by 20–70% (according to the mineralization rate), mainly during periods of eutrophication16.
Because of the important role of POC as a carbon source or sink, an essential task is to determine the origin of this carbon fraction in aquatic ecosystems. For instance, changes in terrestrial and in-lake sources of POC by photosynthesis can affect the internal organic carbon (OC) cycling in the water column17,18,19,20. Better knowledge of these carbon sources and sinks may also help to establish important decision tools for water quality management. This includes prevention and control of eutrophication and pollution, watershed degradation, and landscape-related processes including transportation, transformation and deposition of terrestrial material in aquatic ecosystems21,22.
Aquatic life relies on allochthonous carbon sources from terrestrial detritus and soils, but can also depend on autochthonous sources such as macrophytes and algae. Patterns of processing these sources of organic carbon are different. In general, allochthonous OC is more recalcitrant and has lower degradation rates and thus often becomes preferentially sequestered in sediments23,24,25,26,27. In contrast, autochthonous OC from primary production usually undergoes rapid turnover24. This shows the importance to understand the extents of these endmembers28,29. This is also reflected by the recent scientific literature that has shown increasing interest in POC dynamics, especially by means of stable isotopes of carbon fractions21,30,31,32,33. This demand to better understand POC dynamics also relates to the importance of integral tools to study the production and consumption of dissolved inorganic carbon (DIC) and CO2 through microbial decomposition, degassing, photosynthesis and respiration in aquatic systems17,34,35,36,37. Such approaches have hardly been applied to subtropical reservoirs (and so far nowhere in Brazil), where carbon processing may be particularly intense due to warm temperatures.
This case study was carried out in the Itupararanga Reservoir that is an important freshwater body in the state of São Paulo (Brazil) and has been monitored since 1998 by the State of São Paulo Environmental Company38 [https://cetesb.sp.gov.br/aguas-interiores/publicacoes-e-relatorios/—in Portuguese]. This monitoring revealed that this subtropical reservoir faces progressive deterioration in water quality over the past three decades (Fig. 1). Such processes and related carbon cycling were investigated by POC/chl-a ratios and also by combined application carbon isotope ratios (δ13CPOC and δ13CDIC) and aqueous CO2 contents.

Location map of the Itupararanga Reservoir. Sampling sites P1 to P7 were sampled between December 2016 and December 2018 and are indicated by red stars. The reservoir boundaries were outlined using RapidEye images (5-m spatial resolution from 17 July and 30 August 2014) provided by Ministry of Environment and their drainage systems (1: 25.000) were downloaded from DataGeo (https://datageo.ambiente.sp.gov.br/app/#). All data processing operations were carried out using the software ArcGis 10.2.2 (ESRI, USA) and the maps were projected onto a common coordinate system (World Geodetic System 1984 [WGS 84]).
This subtropical reservoir is economically and environmentally important for the metropolitan region of São Paulo. The general deterioration of environmental and water quality of the Itupararanga system has attracted the interest of several researchers39,40,41,42,43. These studies considered aspects of nutrients and cyanobacteria communities, sedimentary macroinvertebrates, metal toxicity and land-use in relation to water quality. This work has shown that the Itupararanga Reservoir receives substantial amounts of nutrients, mostly because of agricultural land use in the surrounding catchment42. Another important source of nutrients is sewage discharge from urban areas. They mainly reach the reservoir untreated at the entrance39,41. These inputs can stimulate primary production and may also cause eutrophication. This terrestrial nutrient input also suggests that organic matter (OM) and detrital material may enter the reservoir by streams and overland flow via an extensive shoreline. However, less attention has been paid to other carbon sources in this system. Therefore, the aim of this study is to advance knowledge on the reservoir-internal origin of POC by exploring spatiotemporal variations of this carbon fraction together with stable isotope ratios of DIC and POC. This also helps to outline terrestrial inputs. This technique has so far hardly been applied to the Itupararanga Reservoir that can be seen as a representative for a subtropical water body and we introduce the approach as a tool to further understand carbon cycling water systems.
Results
Increased concentrations of both, POC and chl-a were observed at all surface sampling locations at depths above 3 m in December 2017, with maximum values of 2.2 mg L−1 and 42.8 μg L−1 for POC and chl-a (Fig. 2). After this season, a decrease of chl-a and POC concentrations were found in all seven sampling locations between December 2017 and March 2018. The average decrease was 67% for chl-a and 62% for POC and a maximum chl-a decrease of 87% was found at P2. The maximum decrease for POC was 80% at P7. POC and chl-a values at the seven sampling locations in the Itupararanga Reservoir are displayed in Fig. 2.

Concentration patterns of chl-a (white symbols, left axis) and POC (black symbols, right axis) at the seven sampling locations in the Itupararanga Reservoir. Samples presented in this graph were only collected in the top 3 m of the water column. Error bars represent 1σ standard deviations as determined from selected triplicate analyses.
In a first attempt to outline sources, we plotted POC/chl-a ratios against \({\updelta }^{13} {\text{C}}_{{{\text{POC}}}}\) values (Fig. 3). Here we used a threshold of 100 for POC/chl-a ratios to define dominance of photosynthesis over detrital input44. According to this approach, the primary production dominates in only 26% of the surface water samples. These samples are from P1 in March 2017, December 2017 and March 2018, from P2 in December 2017, from P3 in March 2017, from P5 in December 2017 and from P7 in March 2018.

POC/chl-a ratios versus\({\updelta }^{13} {\text{C}}_{{{\text{POC}}}}\) at the seven sampling locations from December 2016 to March 2018 in the top 3 m water depth. Symbols below the POC/chl-a ratio of 100 indicate dominance of photosynthetic sources of POC, the group with POC/chl-a ratios above 100 indicate samples with dominance of detrital sources of organic carbon. Error bars represent 1σ -standard deviations as determined from triplicate analyses of selected samples.
In order to test how carbon might be exchanged between DIC and POC, we investigated their stable isotope values in a Spearman correlation. It indicated positive and significant trends with r values between 0.79 and 0.98 and p values between less than 0.0001 and 0.0182 (Table 1).
Samples from depths below 3 m showed lower values for \({\updelta }^{13} {\text{C}}_{{{\text{DIC}}}}\) that ranged from—10.4‰ to—7.0‰ for all campaigns. Representative values from this group are—8.8‰ for P1 in December 2017 at 3 m,—7.5‰ for P2 in December 2017 at 5 m,—9.2‰ for P3 in March 2017 at 11 m,—9.2‰ for P4 in December 2017 at 8 m,—8.4‰ for P5 in December 2017 at 11 m,—10.4‰ for P6 in December 2017 at 15 m, and—10.1‰ for P7 in December 2017 at 14 m depth (Fig. 4). For the same sampling locations representative samples from the surface had more positive \({\updelta }^{13} {\text{C}}_{{{\text{DIC}}}}\) values with + 1.5‰ for P2 in December 2017 at 1 m, + 0.6‰ for P3 in December 2016 at 1 m, + 0.3‰ for P4 in December 2017 at 1 m and + 0.04‰ for P6 in March 2018 at 1 m depth (Fig. 4).
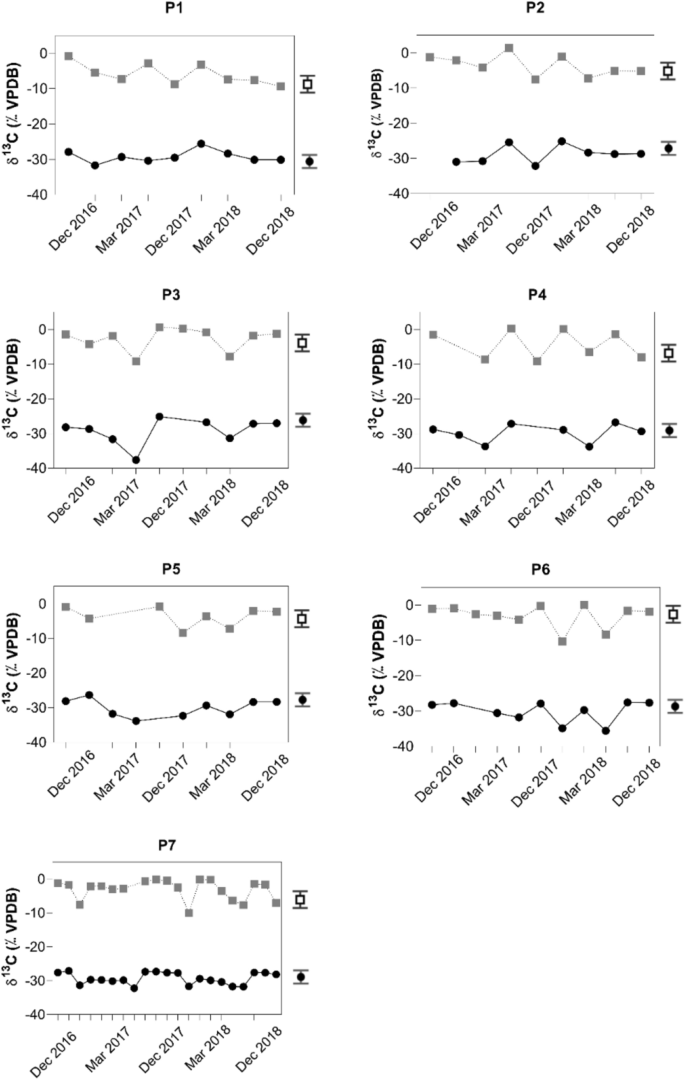
Temporal distributions of δ13CDIC (upper curves) and δ13CPOC (lower curves) for the seven sampling locations in the Itupararanga Reservoir. Each point in the graphs refers to a sampling depth in the water column, which is shown in the Tables S1–S7 (supplementary material) The 1 − σ precisions δ13CPOC and δ13CDIC were ± 0.3‰ and ± 0.1‰, respectively. These are shown by a symbol with error bars on the right side of the graphs.
Carbon isotope fractionation
The data were also plotted according to a fractionation model by Rau et al.55. In this approach the aqueous CO2 contents are plotted versus isotope differences between \({\updelta }^{13} {\text{C}}_{{{\text{CO}}2}}\) and \({\updelta }^{13} {\text{C}}_{{{\text{POC}}}}\) according to Eqs. (2)–(5) (Fig. 5). The same was repeated for surface samples in order to afford the best comparion with the POC/chl-a ratio data (Fig. S1).

Entire set of samples compared to the model by Rau et al.55. (A) Theoretical (solid lines) and measured fractionation values (symbols) (ϵ), between CO2(aq) and phytoplanktonic POC with varying growth rates (μ). (B) Zoom-in. Samples that plot between the curves are of autochthonous origin. Samples below the green solid line (μ = 2.0) do not follow the model by Rau et al.55 and this group rather indicates samples of allochthonous origin.
Figure 5 shows that 80% of the samples collected during the period from December 2016 to December 2018 agree with the fractionation model by primary producers. In contrast, the signal of possibly detrital POC (samples that did not fit the Rau fractionation model) appeared in the warm seasons of December 2017 and March 2018, respectively (Table S10—supplementary material). When treating only the surface samples with this approach, 95% of all samples fitted the model (Fig S1). The ones that did not agree with the fractionation model are samples from P1 in December 2017 and March 2018, both at a water depth of 3 m.
Discussion
The possible significance of subtropical reservoirs in global and regional carbon budgets is still vague. One reason for this is the complexity and variability of external carbon inputs to these systems. Here, we investigated seasonal and spatial variations of POC and chl-a. These parameters can be combined to POC/chl-a ratios. However, this approach only offers a subjective indicator for dominance of external or internal POC sources. Combined investigation of δ13CPOC and δ13CDIC values seem more promising to help separating POC sources, because this technique also maps transformations of carbon. Therefore, the application of a carbon stable isotope fractionation model should be able to provide distinct information on POC sources.
When looking at concentrations of POC and chl-a, most samples in Fig. 2 showed similar spatio-temporal distributions of both parameters. Note that P1 showed no considerable changes in chl-a in December 2016 and March 2017 (i.e. the warm season with precipitation of 148 and 146 mm, respectively). This observation was made even though the corresponding POC concentrations decreased. However, March 2017 and December 2017 had similar precipitation patterns with averages of 148 and 157 mm, respectively, but both chl-a and POC increased. Despite similarities of rainfall averages in both sampling periods, individual intense events may have increased the flushing of nutrients into the reservoir. This may in turn have caused POC generation by primary production. These trends agree with the findings of other studies44,45,46.
Sampling locations P2, P3, P5, P6 and P7 showed decreasing trends in POC from December 2016 to March 2017. These were not matched by increasing chl-a concentrations until the end of December 2017. This trend might be related to declining inputs of detrital material, organic matter consumption, or sedimentary deposition. On the other hand, increases in chl-a contents in the same period was likely caused by supply of phytoplanktonic POC. This was confirmed by an observed algae bloom in the same period, according to the Inland Water Quality Report, produced by CETESB (São Paulo State Environmental Company) in 2017. Higher concentrations of both parameters shown in Fig. 2 may also reflect algae blooms in the reservoir in December 2017.
Low POC/chl-a values in Fig. 3 indicated a dominance of photosynthesis (phytoplankton) in the POC pool. Similar observations were also made by other authors in other open aqueous systems44,47. Even though it is not possible to detect clear seasonal and spatial patterns from this approach, more than half of the samples were classified with origin of detrital dominance (Fig. 3). The other group of samples belong to the warm periods of March 2017, December 2017, and March 2018. They all seem to be characterised by a dominance of photosynthesis. Nevertheless, some sample points of these sampling events also plotted in the detrital field of Fig. 3.
Even though all samples for the chl-a measurements belong to the surface (\(\le\) 3 m depth), their POC/chl-a ratios suggest that 74% of the POC in the Itupararanga Reservoir originate from detritus outside the reservoir. One explanation is that the POC/chl-a threshold ratio of 100 may only serve as a rough and arbitrary indicator for algae versus detrital carbon input. Moreover, this threshold value may vary over time and space. Overall, one would expect much more samples to plot in the field of photosynthesis when considering the known nutrient inputs to the reservoir and the observed algae blooms. One intermediate conclusion is that the POC pool is more complex and cannot be explained with chl-a and POC concentration data alone. This would be plausible, because primary production in complex communities impact aquatic environments via numerous metabolic interactions48.
The Spearman correlation in Table 1 indicates strong relationships between \({\updelta }^{13} {\text{C}}_{{{\text{POC}}}}\) and \({\updelta }^{13} {\text{C}}_{{{\text{DIC}}}}\). This is a good indicator for the possibility that both variables may have been affected by similar processes with biological productivity being most plausible. This means that processes affecting DIC isotope fractionation result from transformation of this carbon source via organic carbon production (photosynthesis) and utilization (respiration).
More 12C-enriched values of \({\updelta }^{13} {\text{C}}_{{{\text{DIC}}}}\) ranged from—10.4‰ to—7.0‰ and suggest respiratory signals that were most obvious at sampling location P1 (Fig. 4). Such 12C-enriched values of \({\updelta }^{13} {\text{C}}_{{{\text{DIC}}}}\) have also been observed in other studies49,50. The fact that these values were observed only for samples from water depths of three meters and below is plausible, because beyond this depth limited penetration of light hampers photosynthesis. This process usually enriches the remaining DIC in 13C. Additionally, increased rainfall may also have flushed 13C-depleted DIC and POC from the surrounding catchment17. The observed more positive δ13CDIC values near the water surface likely result from equilibration with atmospheric CO2 and photosynthesis that both enrich the remaining DIC in 13C.
For a more detailed investigation, we plotted the data according to a model described by Rau et al.55. Overall, the majority of the samples collected between December 2016 and December 2018 revealed a dominance of reservoir-internally produced POC. This may indicate a typical pattern for tropical and subtropical reservoirs with higher temperatures, critical nutrient inputs, and generally enhanced biological activities. The sample group that conforms to the model by Rau et al.55 covers all sampling depths across all seasons. This is a good indication that POC was predominantly produced autochthonously with low spatiotemporal variability. When photosynthetic POC was found outside the photic zone, it likely was produced near the surface and afterwards moved downwards through the water column. The samples that do not fit this model (Fig. 5B), indicate a dominance of allochthonous POC that do not follow the same carbon transfer patterns between aquatic CO2 and POC. This confirms other studies that also showed allochthonous inputs to produce isotope differences between POC and DIC (ε) that are below the modelled lines by Rau et al.44,51.
For a better comparison to the alternative method of POC/chl-a rations, we only considered samples from within 3 m water depth in the Rau Model (Fig. S1 in supplementary material). This shows that only 5% of the sub-data set did not agree with the model (i.e. P1 in December 2017 and P1 in March 2018, both at 3 m depth). The agreement with the model of the rest of the surface water samples in the photosynthetically active zone is a good indicator of biological in situ POC generation by photosynthesis. This model interpretation contrasts with results of POC/chl-a ratios (Fig. 3), that revealed different contributions for autochthonous and allochthonous sources of carbon with 26 and 74%, respectively. This difference is surprising, because the samples for chl-a were also collected in the photic zone of the top 3 m of the water column. One reason for this discrepancy may be that more detrital POC occurs in the reservoir after strong rainfall and storms. These could also mobilize chl-a from the landscape from plant residues. In such a process POC/chl-a ratios could be different to those of freshly produced algae material and mask the effects of in situ photosynthesis. Similar masking effects might exist after re-suspension of sedimentary POC. However, the arbitrary definition of 100 of POC/chl-a ratios for the separation of photosynthesis and detrital is likely the strongest reason for the disagreement between both approaches. For instance, setting this threshold to 150 would result in about half of the samples resulting from terrestrial input.
The fact that many data fit the model at different specific growth rates (μ) may also indicate a seasonal sequence of primary producer types such as Bacillariophyceae, Achnanthidium minutissimum, F. delicatissima var. delicatissima. Such varieties of different algae were also observed in previous studies52,53. This observation is also supported by seasonal patterns of δ13CDIC and δ13CPOC in the data set. Because the investigated seasonality intervals span several months, autochthonous POC can continually be produced in surfaces water and likely has more persistent effects on the reservoir ecosystem than terrestrial POC34.
Overall, simultaneous considerations of δ13CDIC and δ13CPOC together with the availability of CO2(aq) in the Rau Model, marks the transfer of carbon from the DIC to the POC phase. It should therefore be a better indicator of biological use of carbon in the form of algae growth. It also shows how carbon stable isotope fractionation depends on rate-limiting sources and cell size. Thus, this approach seems better in capturing complexities of carbon transfer by algae growth54. Nonetheless, the Rau Model can only predict assimilation of aqueous CO2 and does not account for direct uptake of aqueous HCO3- from the water column. This may ignore a significant part of an algae community that operates in this way55. Moreover, at sites close to the shore, active macrophytes may alter the results by influencing the δ13CDIC that also serves as an input parameter to the Rau Model. Even if macrophytes contribute to the POC pool by plant fragments, they may follow different isotope enrichment patterns that were not investigated here.
The fractionation model by Rau et al.55 was tested for all data and all periods of sampling. It successfully narrowed down the complexity of carbon assimilation to specific fractionation, growth-rates (μ) and extracellular CO2(aq) concentrations. Because it revealed that the majority of samples from less than 3 m water depth (i.e. the photic zone) agreed with this model, photosynthesis and CO2(aq) consumption seems to be the dominating process for POC production. Also, for some samples collected at deeper water depths than 3 m, the model produced plausible results, thus indicating vertical transport of biologically produced POC in the water column.
When considering all data points of this study, about twenty percent of POC samples did not fit the model and plotted below the theoretical lines. These samples most likely correspond to detrital input or re-suspension of sedimentary matter. The latter may either originate from upwelling during storms that in addition can also import more POC from outside the reservoir. These findings can also extend to other freshwater systems and therefore offer a tool to separate POC input pathways that in turn may have important controls on cycling of CO2(aq). This would for instance be the case, if more autochthonous POC is present. On the other hand, predominant POC from terrestrial sources may imply more generation of methane from sediments. This is because more recalcitrant allochthonous material is usually less readily consumed in the water column and becomes more likely deposited in sediments. Here anoxic conditions may produce methane over longer time periods than residence times in the water column.
Overall, our results suggest that stable isotopes can help to trace sources of POC. Such combined applications of stable carbon isotope and physicochemical parameters are promising to provide sensitive differentiations of carbon sources in natural waters.
Methods
Study site
The Itupararanga Reservoir is located in a subtropical climate that belongs to the Cwb-type, according to Köppen classification56. It is situated in the upper Sorocaba River catchment in the state of São Paulo, Brazil (Fig. 1). The climate is characterised by a dry season from April to September and a wet season from October to March. The monthly average temperature is above 18° C for all months, and reaches 22° C in December57. Table S8 (supplementary material) shows monthly precipitation data from 2016 to 2018 of several meteorological stations near the Itupararanga Reservoir from the Brazilian National Institute of Meteorology (INMET) and the Department of Water and Energy (DAAE). The reservoir was built in 1912 with the original purpose of hydroelectric power generation. Nowadays it is predominantly used as a source for drinking water for approximately one million people as well as for irrigation and leisure purposes58. The reservoir has maximum and mean depths of 21 and 7.8 m, respectively. The main channel has a length of about 26 km. The water residence time varies between 95 and 270 days and the reservoir has a maximum storage volume of 286 million m341,58. The most important contributing streams are the Sorocabaçu, Sorocamirim and Una rivers next to numerous small streams (e.g. Paruru, Ressaca, and Campo Verde)59.
Seven locations (P1–P7) inside the Itupararanga Reservoir were sampled between December 2016 and December 2018 at depths between 0 and 15 m below the water surface (Fig. 1). Depending on the depth of the water column sampled intervals were between 1 and 5 m (cf. Tables S1–S7 in the supplementary material).
Land use in the vicinity of sampling locations P1 and P2 is dominated by grass and pasture, however P1 also receives inputs from sewage discharge. Sampling locations P3 and P4 are in the vicinity to urban areas, whereas P5 to P7 are located in parts of the catchment that are dominated by forests, agriculture and silviculture42.
On-site and laboratory procedures
Temperature and pH profiles were measured at all sampling sites using a portable multiparameter probe (Horiba U-50)42. Samples for isotope measurements were filtered via nylon disk filters with a pore size of 0.45 μm into 40-mL amber glass vials according to standards of the U.S. Environmental Protection Agency (EPA vials). All sample vials for isotope measurements were preserved with 0.05 mL of a saturated mercuric chloride (HgCl2) solution in order to avoid secondary biological activities after sampling33.
DIC concentrations and their respective carbon isotope values (\({\updelta }^{13} {\text{C}}_{{{\text{DIC}}}}\)) were determined in continuous flow mode by an Aurora 1030 W TIC-TOC analyser (OI Analytical, College Station, Texas, USA) that was connected to a ThermoFisher Delta V Plus isotope ratio mass spectrometer (IRMS). The 1 − σ precision for DIC concentration measurements were better than 5‰ relative standard deviation (s.d.). The 1 − σ precision for \({\updelta }^{13} {\text{C}}_{{{\text{DIC}}}}\) analyses was determined by at least triplicate measurements of selected control samples and better than ± 0.1‰. Details of coupling the OI analyser to the IRMS and measurement techniques are described in St-Jean60 and van Geldern61.
POC samples were filtered on a glass fibre filter paper with a pore size of 0.4 μm (MN GF-5, Macherey–Nagel, Germany). Prior to sampling, filter papers were heated for 6 h at 500 °C to remove trace amounts of carbon. After filtration in the field, they were dried at 60 °C for 24 h and then pulverized in a carbon-free mortar. Pulverized samples were dried and fumigated by concentrated HCl in a desiccator for 24 h to remove possible carbonate particles. This prepared material was then weighed into tin capsules and analysed on a Costech Elemental Analyser (model ECS 4,010) linked to a ThermoFisher Delta V Plus IRMS for the analysis of the POC carbon isotope ratios (expressed as \({\updelta }^{13} {\text{C}}_{{{\text{POC}}}}\)).
All isotope values are expressed in per mille (‰) against the Vienna Pee Dee Belemnite standard according to:
where R is the ratio of the heavy to the light carbon isotope (i.e. 13C/12C)62. All isotope data were corrected for instrumental drift and linearity. All 1 − σ standard deviations for δ13CPOC at least triplicate measurements of selected samples were better than ± 0.3‰.
Chlorophyll-a (chl-a) was determined according to method of Wetzel and Likens (2000) using a 90% alkaline acetone solution for extraction of the chl-a from the 0.4 μm pore size filters. The absorbance was measured at 665 and 750 nm in a spectrophotometer for determinations of chl-a concentrations per litre. Chl-a was measured only in selected samples that were collected from surface water (1–3 m).
In order to apply the model by Rau et al.55 for evaluation of POC, DIC and CO2 relationships for algal growth the partial pressure of CO2 (pCO2) had to be calculated. This was done as outlined in Marx et al. (2018) by using pH and HCO3- with the following Eq. 63.
where HCO3− is the concentration of bicarbonate, H+ is 10−pH, K1 is the temperature-dependent first dissociation constant for the dissociation of H2CO3 (mol L−1), and KH is the Henry’s law constant in mol L−1 atm−1.
The isotope composition of CO2 (\({\updelta }^{13} {\text{C}}_{{{\text{CO}}_{2} }}\)) was calculated from DIC and the corresponding \({\updelta }^{13} {\text{C}}_{{{\text{DIC}}}}\), pH, and temperature using carbonate equilibrium constants that were linked with isotope equilibrium fractionation factors62,64. This led to the following equation:
where \({\updelta }^{13} {\text{C}}_{{{\text{CO}}2}}\) is the \({\updelta }^{13} {\text{C}}\) of the ambient CO2(aq), \(\epsilon_{P}\) is the temperature-dependent equilibrium isotope fractionation between HCO3− and CO2(aq) with the following equation64,65,66,67.
where b = 9.866, c = − 24.12 and TK is the temperature in Kelvin62. The latter equation was also confirmed by other studies66,67. The photosynthetic fractionation factor between the isotope composition of CO2 and POC was calculated following the model by Rau et al.55:
where \(\epsilon_{p}\) is the temperature-dependent equilibrium stable isotope difference between CO2(aq) and POC. Here \({\updelta }^{13} {\text{C}}_{{{\text{CO}}2\left( {{\text{aq}}} \right)}}\) was derived from Eq. (3) and \({\updelta }^{13} {\text{C}}_{{{\text{POC}}}}\) is the measured stable carbon isotope composition of the POC.
References
Tanentzap, A. J. et al. Bridging between litterbags and whole-ecosystem experiments: a new approach for studying lake sediments. J. Limnol.76, 431–437 (2017).
Mendonça, R. et al. Organic carbon burial in global lakes and reservoirs. Nat. Commun.8, 1–7 (2017).
Raymond, P. A. et al. Global carbon dioxide emissions from inland waters. Nature503, 355–359 (2013).
Tranvik, L. J. et al. Lakes and reservoirs as regulators of carbon cycling and climate. Limnol. Oceanogr.54, 2298–2314 (2009).
Serra-Pompei, C., Hagstrom, G. I., Visser, A. W. & Andersen, K. H. Resource limitation determines temperature response of unicellular plankton communities. Limnol. Oceanogr.64, 1627–1640 (2019).
Webb, J. R., Santos, I. R., Maher, D. T. & Finlay, K. The importance of aquatic carbon fluxes in net ecosystem carbon budgets: a catchment-scale review. Ecosystems22, 508–527 (2019).
Hanson, P. C., Pace, M. L., Carpenter, S. R., Cole, J. J. & Stanley, E. H. Integrating landscape carbon cycling: research needs for resolving organic carbon budgets of lakes. Ecosystems18, 363–375 (2015).
Stets, E. G., Striegl, R. G., Aiken, G. R., Rosenberry, D. O. & Winter, T. C. Hydrologic support of carbon dioxide flux revealed by whole-lake carbon budgets. J. Geophys. Res.114, G01008 (2009).
Ho, D. T. et al. Influence of current velocity and wind speed on air-water gas exchange in a mangrove estuary. Geophys. Res. Lett.43, 3813–3821 (2016).
Read, J. S. et al. Lake-size dependency of wind shear and convection as controlson gas exchange. Geophys. Res. Lett.39, L09405 (2012).
Kokic, J., Wallin, M. B., Chmiel, H. E., Denfeld, B. A. & Sobek, S. Carbon dioxide evasion from headwater systems strongly contributes to the total export of carbon from a small boreal lake catchment. J. Geophys. Res. Biogeosci.120, 16 (2015).
Lapierre, J. F., Seekell, D. A. & Giorgio, P. A. Climate and landscape influence on indicators of lake carbon cycling through spatial patterns in dissolved organic carbon. Glob. Change Biol.21, 4425–4435 (2015).
Chiu, C. Y. et al. Terrestrial loads of dissolved organic matter drive inter-annual carbon flux in subtropical lakes during times of drought. Sci. Total Environ.717, 137052 (2020).
Cole, J. J. & Caraco, N. F. Carbon in catchments: connecting terrestrial carbon losses with aquatic metabolism. Mar. Freshw. Res.52, 101–110 (2001).
Gudasz, C., Bastviken, D., Premke, K., Steger, K. & Tranvik, L. J. Constrained microbial processing of allochthonous organic carbon in boreal lake sediments. Limnol. Oceanogr.57, 163–175 (2012).
Huang, C. et al. Enhanced mineralization of sedimentary organic carbon induced by excess carbon from phytoplankton in a eutrophic plateau lake. J. Soils Sedim.19 (2019).
Kanduč, T., Szramek, K., Ogrinc, N. & Walter, L. M. Origin and cycling of riverine inorganic carbon in the Sava river watershed (Slovenia)inferred from major solutes and stable carbon isotopes. Biogeochemistry86, 137–154 (2007).
Savoye, N. et al. Dynamics of particulate organic matter d15N and d13C during spring phytoplankton blooms in a macrotidal ecosystem (Bay of Seine, France). Mar. Ecol. Prog. Ser.255, 27–41 (2003).
Qin, B., Hu, W., Gao, G., Luo, L. & Zhang, J. Dynamics of sediment resuspension and the conceptual schema of nutrient release in the large shallow Lake Taihu, China. Chin. Sci. Bull.49, 54–64 (2004).
Niemistö, J., Tamminen, P., Ekholm, P. & Horppila, J. Sediment resuspension: rescue or downfall of a thermally stratified eutrophic lake?. Hydrobiologia686, 267–276 (2012).
Chen, D., Liu, Q., Xu, J. & Wang, K. Model-based evaluation of hydroelectric Dam’s impact on the seasonal variabilities of POC in coastal ocean: a case study of three gorges project. J. Mar. Sci. Eng.7, 320 (2019).
Marcé, R. et al. Carbonate weathering as a driver of CO2 supersaturation in lakes. Nat. Geosci.8, 107 (2015).
Quadra, G. R., Sobek, S., Paranaíba, J. R. & Isidorova, A. High organic carbon burial but high potential for methane ebullition in the sediments of an Amazonian hydroelectric reservoir. Biogeosciences17, 1495–1505 (2020).
Guillemette, F., von Wachenfeldt, E., Kothawala, D. N., Bastviken, D. & Tranvik, L. J. Preferential sequestration of terrestrial organic matter in boreal lake sediments. J. Geophys. Res. Biogeosci.122, 863–874 (2017).
Maavara, T. et al. River dam impacts on biogeochemical cycling. Nat. Rev. Earth Environ.1, 103–116 (2020).
Bravo, C., Millo, C., Covelli, S., Contin, M. & De Nobili, M. Terrestrial-marine continuum of sedimentary natural organic matter in a mid-latitude estuarine system. J. Soils Sedim.20, 1074–1086 (2020).
Klump, J. V., Edgington, D. N., Granina, L. & Remsen, C. C. Estimates of the remineralization and burial of organic carbon in Lake Baikal sediments. J. Great Lakes Res.46, 102–114 (2020).
Finlay, J. C. & Kendall, C. in Stable Isotopes in Ecology and Environmental Science 283–333 (2008).
Reitsema, R. E., Meire, P. & Schoelynck, J. The future of freshwater macrophytes in a changing world: dissolved organic carbon quantity and quality and its interactions with macrophytes. Front. Plant Sci.9, 629 (2018).
Attermeyer, K. et al. Organic carbon processing during transport through boreal inland waters: particles as important sites. J. Geophys. Res. Biogeosci.123, 2412–2428 (2018).
Bayer, T. K., Gustafsson, E., Brakebusch, M. & Beer, C. Future carbon emission from boreal and permafrost lakes are sensitive to catchment organic carbon loads. J. Geophys. Res. Biogeosci.124, 1827–1848 (2019).
Du, H. et al. Evaluation of eutrophication in freshwater lakes: a new non-equilibrium statistical approach. Ecol. Ind.102, 686–692 (2019).
Barth, J. A. C., Mader, M., Nenning, F., van Geldern, R. & Friese, K. Stable isotope mass balances versus concentration differences of dissolved inorganic carbon—implications for tracing carbon turnover in reservoirs. Isot. Environ. Health Stud.53, 413–426 (2017).
Chen, J. et al. Combined use of radiocarbon and stable carbon isotope to constrain the sources and cycling of particulate organic carbon in a large freshwater lake, China. Sci. Total Environ.625, 27–38 (2018).
Cole, J. J. et al. Plumbing the global carbon cycle: integrating inland waters into the terrestrial carbon budget. Ecosystems10, 172–185. https://doi.org/10.1007/s10021-006-9013-8 (2007).
Tranvik, L. J., Cole, J. J. & Prairie, Y. T. The study of carbon in inland waters—from isolated ecosystems to players in the global carbon cycle. Limnol. Oceanogr. Lett.3, 41–48 (2018).
van Breugel, Y., Schouten, S., Paetzel, M., Nordeide, R. & Sinninghe Damsté, J. S. The impact of recycling of organic carbon on the stable carbon isotopic composition of dissolved inorganic carbon in a stratified marine system (Kyllaren fjord, Norway). Org. Geochem. 36, 1163–1173 (2005).
CETESB, C. A. D. E. D. S. P. Relação de Áreas Contaminadas, 2013).
Beghelli, F. G. S., Frascareli, D., Pompêo, M. L. M. & Moschini-Carlos, V. Trophic state evolution over 15 years in a tropical reservoir with low nitrogen concentrations and cyanobacteria predominance. Water Air Soil Pollut.227, 95 (2016).
Beghelli, F. G. S., Santos, A. C. A., Urso-Guimarães, M. V. & Calijuri, M. C. Spatial and temporal heterogeneity in a subtropical reservoir and their effects over the benthic macroinvertebrate community. Acta Limnol. Bras.26, 306–317 (2014).
Frascareli, D. et al. Spatial distribution, bioavailability, and toxicity of metals in surface sediments of tropical reservoirs, Brazil. Environ. Monit. Assess.190, 199 (2018).
Melo, D. S. et al. Self-organizing maps for evaluation of biogeochemical processes and temporal variations in water quality of subtropical reservoirs. Water Resour. Res.55, 14p (2019).
Simonetti, V. C., Silva, D. C. C. & Rosa, A. H. Análise da influência das atividades antrópicas sobre a qualidade da água da APA Itupararanga (SP), Brasil. Geosul34, 1–27 (2019).
Barth, J. A. C., Veizer, J. & Mayer, B. Origin of particulate organic carbon in the upper Sr. Lawrence: isotopic constraints. Earth Planet. Sci. Lett.162, 112–121 (1998).
Wei, H., Sun, J., Moll, A. & Zhao, L. Phytoplankton dynamics in the Bohai Sea—observations and modelling. J. Mar. Syst.44, 233–251 (2004).
Zhao, C., Zang, J., Liu, J., Sun, T. & Ran, X. Distribution and budget of nitrogen and phosphorus and their influence on the ecosystem in the Bohai Sea and Yellow Sea, China. Environ. Sci.36, 2115–2127 (2016).
Suzuki, K. W., Ueda, H., Nakayama, K. & Tanaka, M. Spatiotemporal dynamics of stable carbon isotope ratios in two sympatric oligohaline copepods in relation to the estuarine turbidity maximum (Chikugo River, Japan): implications for food sources. J. Plankton Res.36, 461–474 (2014).
Topçuoğlu, B. D., Meydan, C., Nguyenc, T. B., Langc, S. Q. & Holdena, J. F. Growth kinetics, carbon isotope fractionation, and gene expression in the Hyperthermophile Methanocaldococcus jannaschii during hydrogen-limited growth and interspecies hydrogen transfer. Appl. Environ. Microbiol.18, 00180–00181 (2019).
Ravelo, A. C. & Hillaire-Marcel, C. in Proxies in Late Cenozoic Paleoceanography Developments in Marine Geology (eds C. Hillaire–Marcel & A. De Vernal) 735–764 (Elsevier, 2007).
Striegl, R. G., Aiken, G. R., Dornblaser, M. M., Raymond, P. & Wickland, K. P. A decrease in discharge-normalized DOC export by the Yukon River during summer through autumn. Geophys. Res. Lett. L21413 (2005).
Yoshioka, T. Phytoplanktonic carbon isotope fractionation: equations accounting for CO2-concentrating mechanisms. J. Plankton Res.19, 1455–1476 (1997).
Bottino, F., Calijuri, M. C. & Murphy, K. J. Temporal and spatial variation of limnological variables and biomass of different macrophyte species in a Neotropical reservoir (São Paulo—Brazil). Acta Limnol. Bras.25, 387–397 (2013).
Taniwaki, R. H. et al. Structure and dynamics of the community of periphytic algae in a subtropical reservoir (state of São Paulo, Brazil). Acta Botanica Brasilica27, 551–559 (2013).
Baird, M. E., Emsley, S. M. & McGlade, J. M. Using a phytoplankton growth model to predict the fractionantion of stable carbon isotopes. J. Plankton Res.23, 841–848 (2001).
Rau, G. H., Riebesell, U. & Wolf-Gladrow, D. A model of photosynthetic 13C fractionation by marine phytoplankton based on diffusive molecular CO2 uptake. Mar. Ecol. Prog. Ser.133, 275–285 (1996).
Kottek, M., Grieser, J., Beck, C., Rudolf, B. & Rubel, F. World Map of the Köppen–Geiger climate classification updated. Meteorol. Z.15, 259–263 (2006).
Pedrazzi, F. J. D. M., Conceição, F. T. D., Sardinha, D. D. S., Moschini-Carlos, V. & Pompêo, M. Spatial and temporal quality of water in the Itupararanga reservoir, Alto Sorocaba Basin (SP), Brazil. J. Water Resour. Prot.5, 64–71 (2013).
Ribeiro, A. R., Biagioni, R. C. & Smith, W. S. Estudo da dieta natural da ictiofauna de um reservatório centenário. Iheringia104, 404–412 (2014).
Smith, W. S. & Petrere, M. Jr. Spatial and temporal patterns and their influence on fish community at Itupararanga Reservoir, Brazil. Rev. Biol. Trop.56, 2005–2020 (2008).
St-Jean, G. Automated quantitative and isotopic (13C) analysis of dissolved inorganic carbon and dissolved organic carbon in continuous-flow using a total organic carbon analyser. Rapid Commun. Mass Spectrom.17, 419–428 (2003).
van Geldern, R. et al. Stable carbon isotope analysis of dissolved inorganic carbon (DIC) and dissolved organic carbon (DOC) in natural waters—results from a worldwide proficiency test. Rapid Commun. Mass Spectrom.27, 2099–2107 (2013).
Clark, I. D. & Fritz, P. Environmental Isotopes in Hydrogeology (CRC Press, London, 1997).
Marx, A. et al. Groundwater data improve modelling of headwater stream CO2 outgassing with a stable DIC isotope approach. Biogeosciences15, 3093–3106 (2018).
Mook, W. G., Bommerson, J. C. & Staverman, W. H. Carbon isotope fractionation between dissolved bicarbonate and gaseous carbon dioxide. Earth Planet. Sci. Lett.22, 169–176 (1974).
Mayer, B. et al. Assessing the usefulness of the isotopic composition of CO2 for leakage monitoring at CO2 storage sites: a review. Int. J. Greenhouse Gas Control37, 46–60 (2015).
Myrttinen, A., Becker, V. & Barth, J. A. C. Corrigendum to ‘A review of methods used for equilibrium isotope fractionation investigations between dissolved inorganic carbon and CO2’ [Earth Sci. Rev. 115(2012) [192–199]]. Earth-Sci. Rev. 141, 178 (2015).
Myrttinen, A., Becker, V. & Barth, J. A. C. A review of methods used for equilibrium isotope fractionation investigations between dissolved inorganic carbon and CO2. Earth Sci. Rev.115, 192–199 (2012).
Acknowledgements
This research was supported by Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES) and Deutscher Akademischer Austauschdienst (DAAD) through the Project “Organic carbon cycling in water reservoirs-ORCWAR” (DAAD ID 57414997; CAPES Grants 99999.008107/2015-07, 88887.122769/2016-00, 88887.303733/2018-00, 88887.141964/2017-00 and 88887.165060/2018-00) and also Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq Grant 158227/2018-2). We also acknowledge the help of Fundação de Amparo à Pesquisa do Estado de São Paulo (FAPESP, Grants 16/15397-1 and 18/20326-1) for scholarship and financial support. We thank Michael Herzog, Michael Mader, Anne Marx and Christian Hanke for assisting in laboratory analyses and field trips in Brazil.
Author information
Authors and Affiliations
Contributions
C.C. Bueno and J.A.C. Barth contributed to the conceptualisation of the study; D. Frascareli, E.S.J. Gontijo, R. van Geldern and K. Firese performed experiments and were responsible for investigation and validation of the results; C.C. Bueno and J.A.C. Barth were responsible for data interpretation, formal analysis and writing the original draft; A.H. Rosa, K. Friese and J.A.C. Barth were responsible for funding and reviewing and editing the manuscript; A.H. Rosa was responsible for funding acquisition and resources in Brazil. All authors reviewed and agree with the manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Bueno, C.d., Frascareli, D., Gontijo, E.S.J. et al. Dominance of in situ produced particulate organic carbon in a subtropical reservoir inferred from carbon stable isotopes. Sci Rep 10, 13187 (2020). https://doi.org/10.1038/s41598-020-69912-0
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-020-69912-0
This article is cited by
-
Isotopic Composition of Carbon and Nitrogen of Particulate Organic Matter in the Godavari Estuary: Seasonality in Sources and Processes
Estuaries and Coasts (2024)
-
Distribution and dynamics of particulate organic matter in Indian mangroves during dry period
Environmental Science and Pollution Research (2022)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
