
Abstract
Herbivores modulate the structure and composition of plant communities, including plant invasions. This is conditioned by plant palatability which can be reduced by its chemical or physical traits. The effects that ungulates browsing has on pine invasions are variable and the empirical evidence on the causes of this variability is scarce. We experimentally explored how sheep browsing preference varies between seedlings of pine species with different invasiveness; Pinus contorta (high invasiveness), P. ponderosa (medium invasiveness), P. radiata (medium invasiveness) and P. jeffreyi (low invasiveness). Secondly, we quantified anti-herbivory chemical compounds and physical traits of these species and related them with sheep preference observed. The browsing incidence of P. contorta was 68%, P. ponderosa 58%, P. radiata 29%, and P. jeffreyi 84%. Among anti-herbivory traits analyzed, α-pinene concentration had a negative effect on the probability of a terminal bud being browsed and on browsing intensity. Meanwhile, foliar toughness was negatively related to browsing intensity and water concentration was positively related to browsing intensity. Also, the most invasive species, P. contorta, was highly damaged. Thus, sheep herbivory could be slowing pine invasion rate; suggesting that could be considered a tool to control early invasions, especially for this particular species.
Similar content being viewed by others


Introduction
Herbivory is a strong modulator of the structure and dynamic of the plant community, both for its direct and indirect effects on plants (e.g. changes in the chemical and physical composition of the soil)1, 2. In particular, the effect of herbivory on plant invasions has been widely debated in the context of the Natural Enemies Hypothesis, Biotic Resistance, and Enemy Release Hypothesis3,4,5,6,7,8. Herbivores can avoid some species, promoting its invasion, or can limit invasion by consuming them9, 10. The role of introduced ungulates in facilitating plant invasions has been demonstrated by many studies11,12,13,14,15,16,17. Nevertheless, the impact of herbivory on the establishment of non-native species depends on many factors, including plant tolerance, resistance, and palatability.
In the Southern Hemisphere, the invasion by species of the Pinaceae family is well recorded18. The family is native to the northern hemisphere and has been introduced in this region mainly for forestry purposes, but also with the aim of control soil erosion or as ornamentals19. However, the invasiveness among pine species differs. It was suggested that three pine traits mainly contribute to increasing their invasive capacity: low seed mass, short juvenile period, and short reproduction intervals with large seed production20. However, there could be many other factors behind pine invasions, such as abiotic factors (i.e. climatic factors, wind, light availability), the propagule pressure in the area, the presence of mycorrhizal fungi, the biotic resistance of the community where it is introduced, and the presence of herbivores13, 21,22,23.
The effects of ungulates on pine invasions are variable across studies: some studies suggest that they promoted the invasion22, 24,25,26 and others suggest that they hinder the pine invasion27,28,29,30. For example, it was proposed that sheep and cows could favor the Pinus sylvestris and Pinus mugo invasion process in Spain, while Abies alba could be controlled by these ungulates in the same areas, possibly, because of its higher sensitivity to herbivory damage31. Other studies observed that exotic invasive deer preferred native species over the exotic Pinaceae species Pseudotsuga menziesii and Pinus ponderosa in Argentina22, 24. Lastly, in another study, it was observed that feral horses could have promoted the establishment of Pinus halepensis seedlings after a long period of grazing in Argentinean Pampean grasslands26. Among the studies that suggest that ungulates could be controlling pine invasions; it was observed that semi-feral cattle and sheep prevent the regeneration of Pinus nigra and Pinus sylvestris in France28, 29. Finally, it was suggested that exotic vertebrates herbivores negatively affected Pinus radiata survival in Mediterranean areas of Chile30. These opposing results may be due to distinctive characteristics of ungulates; such as its behavior, physiology, habits, and origin; as well as intrinsic characteristics of the plant such as phenology, plant morphology, and palatability32,33,34,35,36.
Regarding plant palatability, the plant nutritional quality (e.g. proportion of fiber and water content), its physical characteristics (e.g. leaf toughness and trichomes), and their secondary metabolites (e.g. alkaloids and monoterpenes) are important traits that affect the ungulate preference32,33,34, 37. Particularly for mammalian herbivores, in several studies, the preference between conifer species was related to the type and concentrations of secondary compounds38,39,40,41,42,43,44. For example, total foliar monoterpenes concentration influenced negatively the browsing behavior of Red deer (Cervus elaphus)38 and Black-tailed deer (Odocoileus hemionus columbianus)39. However, chemical defenses vary greatly between different conifer species and even between genotypes within a species, causing palatability to differ45, 46.
Among ungulates, it is known that sheep exhibit high plasticity in feeding preference, and even though they are considered grazers herbivores, they have been used to successfully control weeds because they can consume plants with higher fiber content than can other domestic ungulates47. Although studies are scarce, it has been suggested that regular sheep browsing could limit the spread of species in the Pinaceae family. Ledgard48 observed that in New Zealand, even low stocking rates of sheep reduced the successful establishment of Pinus contorta seedlings younger than two years old. However, for efficient control, it is necessary more intensive browsing to impede the establishment of new seedlings49. Crozier and Ledgard50 assessed the palatability of seven pine species to sheep in New Zealand. They found a gradient of consumption, with Pinus radiata and Pinus contorta, being the most browsed, and in less degree, Pinus sylvestris, Pinus ponderosa, Larix decidua, Pseudotsuga menziesii, and Pinus nigra. Thus, differential palatability may hinder the control of certain species1, 9, 51, 52. Palatability is a relative concept dependent on plant chemical and physical characteristics and ontogeny32, 53. Herbivore preference could also depend on the relative presence of palatable plants in the community and the nutritional needs of the herbivore54, 55. For example, Cingolani et al.56 observed that characteristics that indicate high structural quality for sheep, were in fact avoided and the authors suggest that sheep select plants for their chemicals compounds also nutritionally necessary at the time of the ingest.
The empirical evidence for the control of pine invasion by sheep is scarce at global level48 and nonexistent for Patagonia. In this study, we take an experimental approach to explore the potential of sheep herbivory to control pine invasion. First, we experimentally explored how sheep browsing differs across seedlings of four pine species with varying invasiveness: P. contorta, P. ponderosa, P. radiata, and P jeffreyi. Second, we quantified anti-herbivore chemical compounds and physical traits in seedlings of these pine species to understand the factors behind the observed preferences.
Methods
Study area
This research was conducted in NW Patagonia in the Fortín Chacabuco ranch, located in Neuquén province, Argentina (41° 0′ 16.67″ S and 71° 10′ 46.62″ W). The landscape is mountainous, and it is crossed by many rivers and streams. The climate is Mediterranean, with 60% of precipitation occurring in autumn and winter. The annual rainfall ranges between 300 and 700 mm per year and the average annual temperature does not exceed 10 °C57. The vegetation corresponds to a semiarid Patagonian steppe in the Andean piedmont and forms a mosaic depending on exposure and soil. The dominant vegetation is the steppe of tussock grasses, Pappostipa speciosa in the lower sectors, and Festuca pallescens in higher areas, with scattered shrubs of Acaena splendens, Senecio bracteolatus, and Mulinum spinosum. There are many flood meadows with Juncus balticus, Poa pratensis, and Festuca pallescens associated with shrub of Ochetophila trinervis, Discaria chacaye, Escallonia virgata, Berberis microphylla and Maytenus boaria57.
The ranch also contains plantations of P. contorta and P. ponderosa, and pines have invaded some areas. Many pines that have escaped from plantations show signs of herbivory. Besides domestic herbivores (cattle, sheep, and horses), invasive non-native herbivores are present, including red deer (Cervus elaphus) and European hare (Lepus europaeus), as well as native herbivores such as guanaco (Lama guanicoe).
Even though sheep production in Patagonia has decreased in recent decades, it remains the main agricultural activity in Patagonia, and the main production is the Merino breed58. As such, sheep are among the most abundant and influential ungulate herbivores in several Patagonian ecosystem processes, including plant invasions59, 60.
Experimental design
To assess sheep preference between four invasive pine species and to estimate anti-herbivores chemical compounds and physical traits of these pine species; we obtained 480 individuals (120 of each species) from a plant nursery that produces seedlings for forestry purposes. We used 400 individuals to perform the preference experiment and we separated 80 individuals from the same population to assess the anti-herbivory chemical compounds and physical traits. The plant nursery is located near the site where we performed the experiment. The seedlings were younger than two years old. The mean initial height of P. contorta seedlings was 6.8 cm (SD 2.4), of P. ponderosa 9.4 cm (SD 2.4), of P. radiata 12.9 cm (SD 3.9) and of P. jeffreyi 10.6 cm (SD 2.5).
Sheep browsing preference
To evaluate the differential preference of sheep (Ovis aries) among non-native pines with different invasive capacity, we exposed seedlings of four species to Merino wethers herbivory (castrated male sheep) under a recommended stocking rate on that site. Pine species were P. contorta (high invasiveness), P. ponderosa and P. radiata (moderate invasiveness) and P. jeffreyi (low invasiveness).
Within the study area, we installed five experimental enclosures of 25 × 25 m with a 1.50 m tall fence (each enclosure constituted a replicate). Enclosures had hardware cloth 0.6 m tall buried in the soil to prevent access by other herbivores, such as European hare (L. europaeus). Each enclosure was stocked with sheep at a density typically recommended for the study area (0.2–0.3 wethers/ha/year for grassland)61, 62. This density was based on the assessment of the forage condition of the sites and the area of the enclosure, according to this, the density estimated corresponded to two wethers in each enclosure for three days. More details about the estimation of the stocking rate can be found as Supplementary Methods S1 online. The experiment was conducted in October 2015, during the austral spring, the season with the maximum availability and quality of forage in the community, minimizing the risk of overgrazing. Also, we established the enclosures adjacent to each other, and no sheep were alone, so as not to alter their gregarious behavior. A similar design was used successfully in previous studies in the region49, 60. Environmental conditions and forage availability between enclosures corresponding to a replicate were similar. In each enclosure (a replicate) we planted 20 seedlings of each pine species (100 individuals of each species in total), alternating species so that there were not two consecutive individuals of the same species.
The number of browsed seedlings, maximum seedling height before and after the treatment, number of terminal buds damaged, number of browsed branches, the number of defoliated seedlings (i.e. whether the seedling was only the needles damaged, not branches) and the probability of seedling survival immediately after the treatment were recorded for all individuals (100 individuals of each of the four species). The probability of seedling survival was estimated based on the conservative assumption that only seedlings with all branches browsed would die. All seedlings were removed at the end of the experiment.
Because our research was conducted on non-regulated animals in our country, we did not require any ethical approval. However is important to mention that all animals were treated under general animal welfare criteria following the guidelines of National Institutes of Animal production63, 64, with constant clean water and food availability (besides the pine seedlings presented during the experiment).
Anti-herbivores chemical compounds and physical traits
To examine whether seedlings of the four Pinus species studied (Pinus contorta, P. ponderosa, P. radiata, and P. jeffreyi) differ in their chemical defenses, physical characteristics and plant quality; we measure toughness and monoterpene content in needles of 20 seedlings per species, and 15 out of those 20 seedlings were used to measured non-volatile resin and water content of the needles and branches. It was not possible to perform the sheep preference experiment and these measurements on the same individuals for two reasons. Firstly, since we were studying seedlings (not juveniles or adults), the amount of biomass left after sheep browsed was not enough to measure the chemical defenses and physical traits in the same individual. Secondly, it is known that pine trees have induced defenses65, 66, so if we measured the chemistry of the seedling after the sheep fed on them, the chemistry would have changed in response to the damage, and we wanted to relate the browsing preference of sheep to the chemistry and physical traits for the four pine species to tissues that were not previously damaged.
To measure needle toughness, 60 fresh needles from 20 seedlings per Pinus species were removed. Needle toughness (g/cm2) was measured with a force gauge penetrometer (type 516; Chatillon, Largo, FL, USA) and three measurements were averaged per seedling. The remaining plant material was placed individually in plastic bags and stored in a − 80 ºC freezer for later analyses. For monoterpene extraction, needles were ground with liquid nitrogen in a chilled mortar and pestle to minimize the possible loss of monoterpene. For each sample, the frozen needle powder was put in a separate borosilicate glass vial, and weighted exact weights (between 0.5 and 0.6 g). After recording the weights, 2 mL of GCgrade n-hexane (Merck, Germany) with 0.1 μL mL-1 ( +)-fenchone (Sigma-Aldrich) as an internal standard was added to each vial. Vials were immediately closed with bakelite caps and parafilm to avoid evaporation and mixed with a vortex. The ground needles were left to soak for 7 days at ambient temperature. After a week, 2 mL aliquots of each sample were placed into a clear chromatography autosampler vials for GC analysis. A 1 μL subsample of each aliquot was then injected on a PerkinElmer Clarus 680 Gas Chromatograph with PerkinElmer Clarus 600 T Mass Spectrometer (GC–MS) fitted with an Elite-5 MS column (60 m × 0.25 mm × 0.25 μm; Perkin Elmer) and coupled with a mass spectrometer (PerkinElmer Clarus 600 T Mass Spectrometer) used as detector. The carrier gas used was Helium at a flow rate of 1 mL/min for 20 mL/min split mode. The injector temperature was set at 250 °C to volatilize the liquid sample and the time–temperature history of the oven enclosing the column consisted of three steps: an initial heating at 60 °C during 1 min, a temperature ramp of at 5 °C/min from 60 to 130 °C, and a final holding at 130 °C during 15 min. We could identify four monoterpenes (α-pinene, β-pinene, 3-carene and β-phellandrene) by comparing retention times of known standards and mass spectra using the NIST Mass Spectral Search Program for the NIST/EPA/NIH Mass Spectral Library (version 2.0f) and TurboMass V 5.4.2 software to calculate the area under the absorbance curve for each peak. Since we were unable to obtain a pure standard of the monoterpene β-phellandrene, this monoterpene was measured using the α-phellandrene pure standard being α and β-phellandrenes similar in retention times and mass spectra. Concentrations for each compound were calculated using 9-point calibration curves with injections of known amounts of pure standards and fenchone (the internal standard). All standards were purchased from Sigma Aldrich (Saint Louis, MO).
In order to measure the non-volatile resin content, needles and branches were analyzed together. We cut 15 cm long branches with its needles in pieces, weighted to the nearest 0.0001 g and placed in labeled borosilicate glass tubes. Then each tube was filled with hexane until the material was covered, mixed with a vortex and placed in an ultrasonic bath at 45 °C for 20 min. The tubes were closed and the plant material soaked for 24 h at room temperature. The entire extraction was repeated twice to maximize extraction outputs. The solution was then filtered with paper filters in pre-weighted borosilicate glass tubes and left to evaporate under the fume hood. Each tube has a residual content that corresponded to the non-volatile resin and was weighted to the nearest 0.0001 g. Since non-volatile resin content was obtained from wet weights, a separate subset of 15 cm branches from each sample was dried (oven dry 60 °C until constant weight) and weighed to obtain dry mass conversion factors. The dry weight (DW) of the sample was calculated using the wet weights of the samples and multiplied by each conversion factor. The resin content of the sample was expressed as mg of non-volatile resin /g of stem DW. To assess plant quality, percent water content was calculated as [(wet weight − dry weight)/wet weight] × 100, from the same 15 cm long branches used to measure total non-volatile resin content.
Data analysis
Sheep browsing preference
We used generalized linear mixed models (GLMM)67, 68 to test the browsing preference of sheep for four species of pines. The models were fitted with the enclosure as a random effect to control for the possible variability between replicates in adjacent enclosures. The predictor variable was pine species, a factor with four levels. We modeled the following six response variables: (a) the probability of a seedling being browsed (browsing incidence: number of browsed seedlings/total number of seedlings), for this variable, we consider that a seedling was browsed if the terminal bud, and/or any branches, and/or the needles were damaged, (b) the relative reduction in height (initial height − final height/initial height), (c) the probability of a terminal bud being browsed, (d) the proportion of browsed branches per seedling (browsing intensity: number of browsed branches/total number of branches), (e) the probability of a seedling being defoliated (whether only the needles were damaged and it was not damaged the stem or branches), and (f) the probability of seedling survival. Models a, c, d, e, and f were fitted with a logit link function and binomial error distribution and model b was fitted with a logit link function and beta error distribution.
Because in the preference trial the four species were presented to sheep at the same time, we performed post-hoc comparisons that compared one species with the other three in turn and not a paired comparison. For that analysis, the lsmeans function from “lsmeans” R package69 was applied and the “Scheffe” adjust method was used.
The function glmer from the R package “lme4”70 was used to fit the browsing incidence, the probability of a terminal bud being browsed, the browsing intensity, the probability of a seedling being defoliated, and the probability of seedling survival models. The function glmmadmb from the “glmmADMB” package71 was used to fit the relative reduction in height (initial height − final height/initial height) model.
Anti-herbivores chemical compounds and physical traits
To identify differences in total monoterpene between species (that follow a normal distribution) we performed an ANOVA and pairwise post-hoc comparisons of the coefficients were tested using the function glht from the “multcomp” package72. The variables α-pinene, β-pinene, 3-carene, β-phellandrene, resins, toughness, and water content did not follow a normal distribution (Shapiro–Wilks Normality Test) so, to test differences in these variables between species, the non- parametric Kruskal–Wallis test was applied, and pairwise post-hoc comparisons of the coefficients were tested using the function kruskalmc from the “pgirmess” R package73.
Relation between sheep preference and pines’ anti-herbivores chemical compounds and physical traits
With the aim to have an overview of the relation between sheep herbivory and the anti-herbivores compounds assessed we explore two aspects of the experiments detailed before. First, to understand how anti-herbivore traits drive the first approach of sheep and pines (i.e. as browsing repellents), we assessed the relation between the concentrations of chemical compounds and physical traits with the probability of a sheep browse only the terminal bud of the seedlings. On the other hand, to understand how anti-herbivore traits drive sheep browsing intensity on pines (i.e. sheep continues to eat the pine after the first approach), we assessed the relationship between these anti-herbivore defenses and browsing intensity. To identify the relation between the chemical and physical variables (predictor variables) with the probability of a terminal bud being browsed or the browsing intensity (response variables) we performed three generalized linear mixed models (GLMM) for each response variable: (1) total monoterpenes and total resins as predictor variables; (2) monoterpenes separately, i.e. α-pinene, β-pinene, 3-carene and β-phellandrene as predictor variables and (3) toughness and water content as predictor variables. The models were fitted with a logit link function and binomial error distribution. Before performing these regressions we checked for collinearity and, following Dormann et al.74, we only include variables with a correlation <|0.7|. Only β-pinene and 3-carene were highly correlated (0.77), so for model 2, we do not include β-pinene. Because the number of individuals tested for sheep preferences (20 seedlings of each species in five enclosures, 100 seedlings of each species in total) was greater than individuals tested for chemical and physical content (15 seedlings of each species to test resins and water, and 20 seedlings of each species to test monoterpenes and foliar toughness), we performed the regressions considering the median of each predictor variable in relation to the individuals tested in the preference experiments. We are aware that this analysis has limitations, but it is appropriate to give a general idea of the principal relations between the anti-herbivory compounds and the sheep preference observed. Also, this approach is used in other biochemical studies due to the high cost and difficulties to assess these traits in larger sampler sizes75. Predictor variables were standardized. The models were nested, fitting species as random effects to consider the possible variability between them. In the cases where there was no variation between species, we fitted a generalized linear model (GLM) without random effects. The function glmer from the R package “lme4”70 was used. Because the predictor variables were standardized, the model estimates correspond to the magnitude of change in the response variable if the predictor variable changes by one deviation standard. Inferences were made from parameters with P < 0.05. The R package ggplot276 was used for visualization of the models. R version 3.5.0 was used77.
Results
Sheep browsing preference
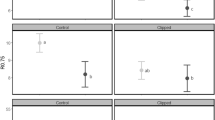
In general terms, the browsing incidence on P. contorta, P. ponderosa, P. radiata, and P. jeffreyi seedlings, as estimated by the model, was 68%, 58%, 29%, and 84% respectively (Table 1). Pinus radiata seedlings were significantly less browsed than the seedlings of the other three pine species, while P. jeffreyi seedlings were significantly more browsed than the seedlings of the other species (Fig. 1a, results of post-hoc comparisons can be found as Supplementary Table S2.1 online).

Herbivory on four pine species with different invasiveness: C: P. contorta (high invasiveness), P: P. ponderosa and R: P. radiata (moderate invasiveness), and J: P. jeffreyi (low invasiveness). (a) Estimated probability of browsing incidence. (b) Relative reduction in seedling height. (c) Estimated probability of a terminal bud being browsed. (d) Estimated probability of browsing intensity. (e) Estimated probability of a seedling being defoliated (only the needles damaged). (f) Estimated probability of seedling survival. Black point refers to the mean and black line to the median, whiskers correspond to SE. Different letters above boxplots indicate significant differences (Scheffe method).
The relative reduction in P. contorta, P. ponderosa, P. radiata, and P. jeffreyi seedling height estimated by the model was 27%, 24%, 23%, and 18% respectively (Table 1) with the reduction in height of P. contorta seedlings significantly higher than the reductions in height of P. radiata and P. jeffreyi seedlings; and reductions in height of P. ponderosa seedlings were significantly greater than the reduction in height of P. jeffreyi seedlings (Fig. 1b; results of post-hoc comparisons can be found as Supplementary Table S2.1 online).
The probability that a terminal bud of a P. contorta, a P. ponderosa, a P. radiata and a P. jeffreyi seedling being damaged, as estimated by the model, was 44%, 29%, 31%, and 19% respectively (Table 1); with the proportion of terminal bud browsed of P. contorta seedlings significantly higher than the proportion of terminal bud browsed of P. jeffreyi (Fig. 1c, results of post-hoc comparisons can be found as Supplementary Table S2.1 online).
The browsing intensity of P. contorta, P. ponderosa, P. radiata and P. jeffreyi seedlings estimated by the model was 28%, 29%, 27%, and 18%, respectively (Table 1); with the browsing intensity of P. jeffreyi significantly lower than the browsing intensity of P. contorta and P. ponderosa (Fig. 1d, results of post-hoc comparisons can be found as Supplementary Table S2.1 online).
In terms of the probability of a seedling having only the needles damaged, i.e., being defoliated, the probability estimated by the model for P. contorta, P. ponderosa, P. radiata and P. jeffreyi seedling was 10%, 30%, 1.7%, and 56% respectively (Table 1). These percentages differed significantly for the four species (Fig. 1e, results of post-hoc comparisons can be found as Supplementary Table S2.1 online).
The probability of survival of a P. contorta, a P. ponderosa, a P. radiata and a P. jeffreyi seedling immediately after the treatment was 93%, 90%, 91% and 99% respectively (Table 1), with the probability of survival of P. jeffreyi significantly higher than the probability of survival of P. ponderosa and P. radiata seedlings (Fig. 1f; results of post-hoc comparisons can be found as Supplementary Table S2.1 online).
Anti-herbivores chemical compounds and physical traits
The mean concentration of α-pinene in seedlings of P. contorta, P. ponderosa, P. radiata, and P. jeffreyi was 0.44, 0.98, 0.40 and 3.02 μL/g respectively; with the concentration of P jeffreyi seedlings significantly higher than the other three species, and the concentration of P. ponderosa seedlings significantly higher than the concentration of P. contorta and P. radiata seedlings (Kruskal–Wallis chi-squared = 61.177, df = 3, p value = 3.294e−13; Fig. 2a, results of post-hoc comparisons can be found as Supplementary Table S3.2 online).

Chemical compounds and physical traits of four pine species with different invasiveness: C: P. contorta (high invasiveness), P: P. ponderosa and R: P. radiata (moderate invasiveness), and J: P. jeffreyi (low invasiveness). (a) α-pinene, (b) Toughness, (c) Water content, (d) Total monoterpenes, (e) β-pinene, (f) 3-carene, (g) β-phellandrene, (h) non-volatile resins. Black point refers to the mean and black line to the median, whiskers correspond to SE. Different letters above boxplots indicate significant differences. Data of total monoterpenes, α-pinene β-pinene, β-phellandrene, 3-carene, and toughness was taken from the needles and data of non-volatile resins content and water content was taken from branches and needles together.
The mean foliar toughness of P. contorta, P. ponderosa, P. radiata and P. jeffreyi seedlings was 98.25, 173.25, 47.75 and 209 g/cm2 respectively; with the toughness of P. ponderosa and P. jeffreyi significantly higher than the toughness of P. contorta and P. radiata seedlings (Kruskal–Wallis chi-squared = 67.654, df = 3, p value = 1.357e−14; Fig. 2b, results of post-hoc comparisons can be found as Supplementary Table S3.2 online).
The percent water content of P. contorta, P. ponderosa, P. radiata and P. jeffreyi seedlings was 0.56, 0.61, 0.58 and 0.46% respectively; with the percent water content of P. ponderosa significantly higher than the percent water content of P. contorta and P. jeffreyi and the percent water content of P. radiata significantly higher than the percent water content of P. jeffreyi (Kruskal–Wallis chi-squared = 54.388, df = 3, p value = 9.274e−12; Fig. 2c, results of post-hoc comparisons can be found as Supplementary Table S3.2 online).
The concentrations of total monoterpenes, β-pinene, 3-carene, β-phellandrene, and non-volatile resins differed between pine species (Fig. 2d, e, f, g, and h, respectively). Mean values and results of statistical analyses are detailed in Tables 2 and 3 (Results of post-hoc comparisons can be found as Supplementary Table S3.2 online).
Relation between sheep preference and pines’ anti-herbivores chemical compounds and physical traits
Here, we report the general inferences made from parameters with p < 0.05. Details about final models, parameter estimates, significances, and confidence intervals can be found in Tables 4 and 5. Out of all anti-herbivores defenses analyzed, only α-pinene had a significant negative effect on the probability of a terminal bud being browsed (Table 4). The concentrations of α-pinene and foliar toughness were negatively related to browsing intensity (Table 5). Conversely, water concentration was positively related to browsing intensity (Table 5).
Discussion
We show that all pine species were browsed by sheep; however, the magnitude and the pattern of damage differed. Regarding the browsing incidence, P. jeffreyi was the most preferred species, followed by P. contorta, and P. ponderosa, and lastly P. radiata. That is, sheep mostly preferred the least invasive and the most invasive species. But, regarding browsing intensity, sheep browsed more intensely the most invasive pine species. Since the four species were damaged, sheep browsing at this stocking rate could be affecting the growth rate and retarding seedlings from growing into adults78, slowing down the invasion rate, although not stopping it.
Regarding the chemical compounds, we observed that the total concentration of monoterpenes does not influence the attractiveness of pines to sheep, nor the subsequent damage as we did not find a significant effect on the probability of browse on terminal buds not browsing intensity. But looking specifically across monoterpenes, the concentration of α-pinene was the major deterrent to browsing. Therefore, in general terms, the concentration of total monoterpenes does not prevent the browsing of pines by sheep but the presence of certain types of terpenes could be conditioning their pattern of herbivory. The negative effects of α-pinene concentration on sheep browsing observed in our study were also observed in another study79. Estell et al.79 performed an experiment assessing intake by sheep of alfalfa pellets treated with six chemical compounds (camphor, limonene, cis-jasmone, β-caryophyllene, borneol, or α-pinene), each one at five different concentrations. They observed a negative effect of chemical concentration on the intake of camphor and α-pinene. The general lack of anti-herbivore effect by monoterpenes concentration (except α-pinene) observed in our study could be due to the sheep domestic condition, probably individuals with high reaction to adverse stimuli were not selected during domestication80,81,82.
We found that foliar toughness negatively influenced browsing intensity. The differential browsing observed between the four species (terminal bud, lateral branches or needle damage) may be due to the differences in morphology and/or degree of lignification of each species (all seedlings were younger than two years old, thus these differences were not due to age, but rather intrinsic differences of the species). Pinus jeffreyi and P. ponderosa seedlings presented the highest foliar toughness and also, they were the most defoliated. Cingolani et al.56 measured plant functional traits and estimated sheep selectivity and plant grazing response, they observed that characteristics that indicate high structural quality for herbivores, between them low toughness, were in fact avoided suggesting that chemical defenses could be playing a more important role for this herbivore. In fact, Villalba and Provenza83 posed the hypothesis that food structure and nutritional composition interact to determine the lambs' preferences, so, when a particular nutrient is needed, the biochemical composition is more important than the structure in determining the preference. This pattern was also observed by Evju et al.84 in a study of sheep selectivity, they found that sheep select large, late-flowering herbs that have higher nutritional qualities (low leaf C/N ratio). In our study, P. jeffreyi and P. ponderosa, species with high browsing incidence, could have higher nutritional qualities for sheep, even when the physical structure was not the ideal for a grazer herbivore (high toughness). Regarding nutritional qualities, in our study, we only assessed seedlings’ water content. We observed that this variable had a significant positive influence on browsing intensity. It would be important to complement this work with studies of the influence of other nutritional compounds on sheep’s preference, like carbon, nitrogen, and phosphorus content.
It is important to clarify that due to the well-known induced response of Pinaceae to herbivory65, 66, the preference experiments and the anti-herbivory traits were measured in different plant material. Measuring these different variables in different plants might create differences that are not possible to account for. Even though all the seedlings came from the same source and were from the same pool, it is possible that other factors influenced the preference observed besides the traits assessed in our work, like differential microsite growth condition of each individual or morphological differences that were not possible to includes in the analyses, like plant height. However, we think that the performed analyses were the best possible approach to further our understanding of the relations between the anti-herbivory compounds and the sheep preference observed.
Some studies suggest that plant invasions could be controlled with mammal herbivory in other systems28,29,30, 85, 86. In Patagonia, Sarasola et al.27 suggested that domestic ungulates could be one of the factors that are controlling pines invasions. However, in that study, the authors did not detail the species of ungulate, and for Patagonia, no empirical evidence about sheep herbivory on pines has been recorded to date. We observed that sheep damaged pine seedlings according to this decreasing gradient of preference: Pinus jeffreyi, Pinus contorta, Pinus ponderosa, and Pinus radiata. This suggests that the most invasive species, P. contorta, could potentially be controlled with sheep herbivory. In another study in New Zealand, Crozier and Ledgard50 found that Pinus radiata and Pinus contorta were the most consumed species by sheep, followed by Pinus ponderosa. Our results correspond partially with this gradient of consumption since P. radiata was the least preferred species. In another study done in Puerto Patriada, Argentina, it was found that P. radiata individuals from different cohorts growing outside of plantations had varying non-volatile resin content of needles and branches (Dimarco personal communication). The difference in the sheep preference gradient found in our study and the one found by Crozier and Ledgard in New Zealand could be explained by the distinctive levels in chemical defenses that different genotypes of the same plant species could possess46, 87, and also by the growing conditions of the pine seedlings used by Crozier and Ledgard and in our study.
Even though P. contorta was preferred by sheep in the present study, we observed in a previous study that the stocking rate implemented will determine the effectiveness of the invasion control49. The stocking rate must be estimated taking into account the palatability, the pine age, the regeneration rate, and the degree of invasion of the focal species. If the species to control are unpalatable, it could be necessary stock management concentrating high stocking rates in key seasons (i.e. spring)88. Regarding the age, previous studies observed that the concentration of anti-herbivores compounds changes with the ontogeny89. So, we suggest performing an herbivory invasion control over seedling younger than 2 years old50 to guarantee high browse intensity of the seedling in order to avoid branch resprouting and ensure mortality. Lastly, Ledgard48, suggested that P. contorta is among the planted pine species with the highest regeneration rate. Thus, taking into account all the variables mentioned, for an effective herbivory control of P. contorta invasion (one of the most invasive species in the Southern Hemisphere) we suggest apply a high stocking rate in news fronts of invasion during short periods but with an annual frequency.
Our study suggests that sheep browsing could be a practical tool to control pine spread beyond the plantations. We observed that sheep consumed seedlings across our pine species at different intensities. Herbivory pine control could be more effective for pine species with low α-pinene concentration, low foliar toughness, and high nutritional qualities. Forestry practices accompanied by appropriate stock management plans could be an efficient method to control early pine invasions.
Data availability
The datasets generated during the current study are available from the corresponding author upon request.
References
Augustine, D. J. & McNaughton, S. J. Ungulate effects on the functional species composition of plant communities: herbivore selectivity and plant tolerance. J. Wildl. Manag. 62, 1165–1183 (1998).
Huntly, N. Herbivores and the dynamics of communities and ecosystems. Annu. Rev. Ecol. Syst. 22, 477–503 (1991).
Keane, R. M. & Crawley, M. J. Exotic plant invasions and the enemy release hypothesis. Trends Ecol. Evol. 17, 164–170 (2002).
Maron, J. L. & Vila, M. When do herbivores affect plant invasion? Evidence for the natural enemies and biotic resistance hypotheses. Oikos 95, 361–373 (2001).
Lockwood, J. L., Hoopes, M. F. & Marchetti, M. P. Establishment success: the influence of biotic interactions. In Invasion Ecology (eds Lockwood, J. L. et al.) 107–131 (Wiley, Hoboken, 2013).
Meijer, K., Schilthuizen, M., Beukeboom, L. & Smit, C. A review and meta-analysis of the enemy release hypothesis in plant-herbivorous insect systems. PeerJ https://doi.org/10.7717/peerj.2778 (2016).
Nunez-Mir, G. C. et al. Biotic resistance to exotic invasions: its role in forest ecosystems, confounding artifacts, and future directions. Biol. Invasions 19, 3287–3299 (2017).
Jeschke, J. M. & Heger, T. Invasion Biology. Hypotheses and Evidence (CABI, Wallingford, 2018).
Averill, K. M., Mortensen, D. A., Smithwick, E. A. H. & Post, E. Deer feeding selectivity for invasive plants. Biol. Invasions 18, 1247–1263 (2016).
Parker, J., Burkepile, D. & Hay, M. Opposing effects of native and exotic herbivores on plant invasions. Science 311, 1459–1461. https://doi.org/10.1126/science.1121407 (2006).
Hobbs, R. J. Synergisms among habitat fragmentation, livestock grazing, and biotic invasions in Southwestern Australia. Conserv. Biol. 15, 1522–1528 (2001).
Knight, T. M., Dunn, J. L., Smith, L. A., Davis, J. & Kalisz, S. Deer facilitate invasive plant success in a Pennsylvania forest understory. Nat. Areas J. 29, 110–116 (2009).
Nuñez, M. A. et al. Exotic mammals disperse exotic fungi that promote invasion by exotic trees. PLoS ONE 8, 1–6 (2013).
Oduor, A. M. O., Gómez, J. M. & Strauss, S. Y. Exotic vertebrate and invertebrate herbivores differ in their impacts on native and exotic plants: a meta-analysis. Biol. Invasions 12, 407–419 (2010).
Spear, D. & Chown, S. L. Non-indigenous ungulates as a threat to biodiversity. J. Zool. 279, 1–17 (2009).
Vavra, M., Parks, C. G. & Wisdom, M. J. Biodiversity, exotic plant species, and herbivory: the good, the bad, and the ungulate. For. Ecol. Manag. 246, 66–72 (2007).
Loydi, A. & Zalba, S. M. Feral horses dung piles as potential invasion windows for alien plant species in natural grasslands. Plant Ecol. 201, 471–480 (2009).
Richardson, D. M. & Rejmánek, M. Conifers as invasive aliens: a global survey and predictive framework. Divers. Distrib. 10, 321–331 (2004).
Simberloff, D. et al. Spread and impact of introduced conifers in South America: lessons from other southern hemisphere regions. Austral. Ecol. 35, 489–504 (2010).
Rejmánek, M. & Richardson, D. M. What attributes make some plant species more invasive?. Ecology 77, 1655–1661 (1996).
Nuñez, M. A., Simberloff, D. & Relva, M. A. Seed predation as a barrier to alien conifer invasions. Biol. Invasions 10, 1389–1398 (2008).
Nuñez, M. A., Relva, M. A. & Simberloff, D. Enemy release or invasional meltdown? Deer preference for exotic and native trees on Isla Victoria Argentina. Austral. Ecol. 33, 317–323 (2008).
Nuñez, M. A. & Medley, K. A. Pine invasions: climate predicts invasion success; something else predicts failure. Divers. Distrib. 17, 703–713 (2011).
Relva, M. A., Nuñez, M. A. & Simberloff, D. Introduced deer reduce native plant cover and facilitate invasion of non-native tree species: evidence for invasional meltdown. Biol. Invasions 12, 303–311 (2010).
Osem, Y., Lavi, A. & Rosenfeld, A. Colonization of Pinus halepensis in Mediterranean habitats: consequences of afforestation, grazing and fire. Biol. Invasions 13, 485–498 (2011).
de Villalobos, A., Zalba, S. M. & Peláez, D. V. Pinus halepensis invasion in mountain pampean grassland: effects of feral horses grazing on seedling establishment. Environ. Res. 111, 953–959 (2011).
Sarasola, M. M., Rusch, V. E., Schlichter, T. M. & Ghersa, C. M. Invasión de coníferas forestales en áreas de estepa y bosques de ciprés de la cordillera en la Región Andino Patagónica. Ecol. Austral. 16, 143–156 (2006).
Chauchard, S., Pille, G. & Carcaillet, C. Large herbivores control the invasive potential of nonnative Austrian black pine in a mixed deciduous Mediterranean forest. Can. J. For. Res. 36, 1047–1053 (2006).
Boulant, N., Kunstler, G., Rambal, S. & Lepart, J. Seed supply, drought, and grazing determine spatio-temporal patterns of recruitment for native and introduced invasive pines in grasslands. Divers. Distrib. 14, 862–874 (2008).
Becerra, P. I. & Bustamante, R. O. The effect of herbivory on seedling survival of the invasive exotic species Pinus radiata and Eucalyptus globulus in a Mediterranean ecosystem of Central Chile. For. Ecol. Manag. 256, 1573–1578 (2009).
Bartolomé, J., Boada, M., Saurí, D., Sánchez, S. & Plaixats, J. Conifer dispersion on subalpine pastures in Northeastern Spain: characteristics and implications for rangeland management. Rangel. Ecol. Manag. 61, 218–225 (2008).
Forbes, J. M. Learning about food: conditioned preferences and aversions. In Voluntary Food Intake and Diet Selection in Farm Animals (ed. Forbes, J. M.) 117–143 (CABI, Wallingford, 2007).
Danell, K., Bergström, R. & Edenius, L. Effects of large mammalian browsers on architecture, biomass, and nutrients of woody plants. Source J. Mammal. 75, 833–844 (1994).
McNaughton, S. J. Grazing as an optimization process: grass-ungulate relationships in the Serengeti. Am. Nat. 113, 691–703 (1979).
Canham, C. D., McAninch, J. B. & Wood, D. M. Effects of the frequency, timing, and intensity of simulated browsing on growth and mortality of tree seedlings. Can. J. For. Res. 24, 817–825 (1994).
Persson, I. L., Danell, K. & Bergström, R. Different moose densities and accompanied changes in tree morphology and browse production. Ecol. Appl. 15, 1296–1305 (2005).
Pollock, M. L., Lee, W. G., Walker, S. & Forrester, G. Ratite and ungulate preferences for woody New Zealand plants: influence of chemical and physical traits. N. Z. J. Ecol. 31, 68–78 (2007).
Duncan, A. J., Hartley, S. E. & Iason, G. R. The effect of monoterpene concentrations in Sitka Spruce (Picea sitchensis) on the browsing behavior of red deer (Cervus elaphus). Can. J. Zool. Can. Zool. 72, 1715–1720 (1994).
Kimball, B. A., Russell, J. H. & Ott, P. K. Phytochemical variation within a single plant species influences foraging behavior of deer. Oikos 121, 743–751 (2012).
Zhang, X. & States, J. S. Selective herbivory of Ponderosa pine by Abert squirrels: a re-examination of the role of terpenes. Biochem. Syst. Ecol. 19, 111–115 (1991).
Elliott, S. & Loudon, A. Effects of monoterpene odors on food selection by red deer calves (Cervus elaphus). J. Chem. Ecol. 13, 1343–1349 (1987).
Bryant, J. P. et al. Interactions between woody plants and browsing mammals mediated by secondary metabolites. Annu. Rev. Ecol. Syst. 22, 431–446 (1991).
Bryant, J. P., Reichardt, P. B. & Clausen, T. P. Chemically mediated interactions between woody plants and browsing mammals. J. Range Manag. 45, 18–24 (1992).
Baraza, E., Hódar, J. A. & Zamora, R. Consequences of plant-chemical diversity for domestic goat food preference in Mediterranean forests. Acta Oecol. 35, 117–127 (2009).
Moreira, X. et al. Trade-offs between constitutive and induced defences drive geographical and climatic clines in pine chemical defences. Ecol. Lett. 17, 537–546 (2014).
Radwan, M. A. & Crouch, G. L. Selected chemical constituents and deer browsing preference of Douglas Fir. J. Chem. Ecol. 4, 675–683 (1978).
Frost, R. A. & Launchbaugh, K. L. Grazing for Rangeland Weed Managenent: a new look at an old tool. Rangelands 25, 43–47 (2003).
Ledgard, N. J. The spread of lodgepole pine (Pinus contorta, Dougl.) in New Zealand. For. Ecol. Manag. 141, 43–57 (2001).
Zamora-Nasca, L. B., Relva, M. A. & Núñez, M. A. Ungulates can control tree invasions: experimental evidence from nonnative conifers and sheep herbivory. Biol. Invasions 20, 583–591 (2018).
Crozier, E. R. & Ledgard, N. J. Palatability of wilding conifers and control by simulated sheep browsing. In Alternatives to the Chemical Control of Weeds. Proceedings of International Conference, Rotorua, July 1989. Bulletin No. 155 (eds Basset, C. et al.) 139–143 (Rotorua, Ministry of Forestry, Forest Research Institute, 1990).
Mayle, B. Domestic Stock Grazing to Enhance Woodland Biodiversity (Forestry Commission, Edinburgh, 1999).
Westoby, M. The LHS Strategy Scheme in Relation to Grazing and Fire. VIth International Rangeland Congress (Australian Rangeland Society, Canberra, 1999).
Westoby, M. An analysis of diet selection by large generalist herbivores. Am. Nat. 108, 290–304 (1974).
Villalba, J. J., Burritt, E. A. & Clair, S. B. S. Aspen (Populus tremuloides Michx.) intake and preference by mammalian herbivores: the role of plant secondary compounds and nutritional context. J. Chem. Ecol. 40, 1135–1145 (2014).
Rhodes, A. C., Larsen, R. T., Maxwell, J. D. & St. Clair, S. B. Temporal patterns of ungulate herbivory and phenology of aspen regeneration and defense. Oecologia 188, 707–719 (2018).
Cingolani, A. M., Posse, G. & Collantes, M. B. Plant functional traits, herbivore selectivity and response to sheep grazing in Patagonian steppe grasslands. J. Appl. Ecol. 42, 50–59 (2005).
Bran, D., Ayesa, J. & Lopez, C. Áreas ecológicas de Neuquen (Instituto Nacional de Tecnología Agropecuaria - INTA, Buenos Aires, 2002).
Mueller, J. Producción ovina en Argentina, situación actual y perspectivas futuras. Boletín Inf. INTA 200, 19–21 (2001).
Aguiar, M. R. & Sala, O. E. Interactions among grasses, shrubs, and herbivores in Patagonian grass-shrub steppes. Ecol. Austral. 8, 201–210 (1998).
Zamora-Nasca, L. B., Relva, M. A. & Núñez, M. A. Ungulate browsing on introduced pines differs between plant communities: implications for invasion process and management. Austral. Ecol. 44, 973–982 (2019).
Bonvissuto, G. L., Somlo, R. C., Lanciotti, M. L., Carteau, A. G. & Busso, C. A. Guías de Condición para Pastizales Naturales de ‘Precordillera’, ‘Sierras y Mesetas’ y ‘Monte Austral’ de Patagonia (Instituto Nacional de Tecnología Agrpecuaria - INTA, Buenos Aires, 2008).
Siffredi, G. L. et al. Guía para la evaluación de Pastizales. Para las áreas ecológicas de Sierras y Mesetas Occidentales y de Monte de Patagonia Norte (INTA, Buenos Aires, 2013).
SENASA. Manual de Bienestar Animal Un enfoqe práctico para el buen manejo de especies domésticas durante su tenencia, producción, concentración, transporte y faena (Servicio Nacional de Sanidad y Calidad Agroalimentaria, Buenos Aires, 2015).
Álvarez, J. M. et al. Bienestar animal Ovino (2005).
Mumm, R. & Hilker, M. Direct and indirect chemical defence of pine against folivorous insects. Trends Plant Sci. 11, 351–358 (2006).
Moreira, X. et al. Inducibility of chemical defences by two chewing insect herbivores in pine trees is specific to targeted plant tissue, particular herbivore and defensive trait. Phytochemistry 94, 113–122 (2013).
Crawley, M. J. Mixed-effect models. In The R Book 681–714 (Wiley, 2013).
Zuur, A. F., Ieno, E. N., Walker, N. J., Saveliev, A. A. & Smith, G. M. Mixed Effects Models and extensions in Ecology with R (Springer, Berlin, 2009).
Lenth, M. R. Package ‘lsmeans’. CRAN (2013).
Bates, D. M., Machler, M., Bolker, B. & Walker, S. Fitting linear mixed-effects models using lme4. J. Stat. Softw. 67, 1–48 (2015).
Skaug, H., Fournier, D., Nielsen, A., Magnusson, A. & Bolker, B. M. Generalized linear mixed models using ‘AD model builder’. Optim. Methods Softw. 27, 233–249 (2012).
Hothorn, T. et al. Package ‘multcomp’ - Simultaneous Inference in General Parametric Models. (2017). https://cran.r-project.org/web/packages/multcomp/multcomp.pdf
Giraudoux, P., Antonietti, J.-P., Beale, C., Pleydell, D. & Treglia, M. Package ‘pgirmess’: Spatial Analysis and Data Mining for Field Ecologists (2018). https://doi.org/10.1145/3097983.3098168doi:
Dormann, C. F. et al. Collinearity: a review of methods to deal with it and a simulation study evaluating their performance. Ecography (Cop.) 36, 027–046 (2013).
Peuke, A. D. Correlations in concentrations, xylem and phloem flows, and partitioning of elements and ions in intact plants. A summary and statistical re-evaluation of modelling experiments in Ricinus communis. J. Exp. Bot. 61, 635–655 (2010).
Wickham, H. ggplot2. Elegant Graphics for Data Analysis (2009). doi:10.1007/978-0-387-98141-3.
R Development Core Team. R: A language and environment for statistical computing. Vienna, Austria. https://www.R-project.org/ (2018).
Zamora, R., Gómez, J. M., Hódar, J. A., Castro, J. & García, D. Effect of browsing by ungulates on sapling growth of Scots pine in a mediterranean environment: consequences for forest regeneration. For. Ecol. Manag. 144, 33–42 (2001).
Estell, R. E. et al. Effects of volatile compounds on consumption of alfalfa pellets by sheep. J. Anim. Sci. 76, 228–233 (1998).
Kruska, D. & Stephan, H. Volumenvergleich allokortikaler Hirnzentren hei Wild- und Hausschweinen. Acta Anat. 84, 387–415 (1973).
Arnold, G. W., De Boer, E. S. & Boundy, C. A. P. The influence of odour and taste on the food preferences and food intake of sheep. Aust. J. Agric. Res. 31, 571–587 (1980).
Tribe, D. E. The importance of the sense of smell to the grazing sheep. J. Agric. Sci. 39, 309–312 (1949).
Villalba, J. J. & Provenza, F. D. Effects of food structure and nutritional quality and animal nutritional state on intake behaviour and food preferences of sheep. Appl. Anim. Behav. Sci. 63, 145–163 (1999).
Evju, M., Austrheim, G., Halvorsen, R. & Mysterud, A. Grazing responses in herbs in relation to herbivore selectivity and plant traits in an alpine ecosystem. Oecologia 161, 77–85 (2009).
Cadenasso, M. L., Pickett, S. T. A. & Morin, P. J. Experimental test of the role of mammalian herbivores on old field succession: Community structure and seedling survival. J. Torrey Bot. Soc. 129, 228–237 (2002).
Capó, E. A., Aguilar, R. & Renison, D. Livestock reduces juvenile tree growth of alien invasive species with a minimal effect on natives: a field experiment using exclosures. Biol. Invasions 18, 2943–2950 (2016).
Dimock, E. J., Silen, R. R. & Allen, V. E. Genetic resistance in Douglas -fir to damage by snowshoe hare and black-tailed deer. For. Sci. 22, 106–121 (1976).
Mobæk, R., Mysterud, A., Egil Loe, L., Holand, Ø & Austrheim, G. Density dependent and temporal variability in habitat selection by a large herbivore; an experimental approach. Oikos 118, 209–218 (2009).
Iason, G. R., O’Reilly-Wapstra, J. M., Brewer, M. J., Summers, R. W. & Moore, B. D. Do multiple herbivores maintain chemical diversity of Scots pine monoterpenes? Philos. Trans. R. Soc. Lond. B. Biol. Sci. 366, 1337–1345 (2011).
Acknowledgements
We are grateful to The Nature Conservancy and the Fortin Chacabuco ranch for allowing us to work in the area, especially to F. Montenegro and N. Rodríguez for their invaluable help. We also thank K. Stuble for the detailed English review and F. Tiribelli and A. di Virgilio for their suggestions that helped to improve the manuscript. This work was supported by a fellowship (LZN) and two grants from Agencia Nacional de Promoción Científica y Tecnológica of Argentina (PICT 2012-2283 and PICT 2016-0964). We also thank two anonymous reviewers for their very helpful comments that improved this manuscript.
Author information
Authors and Affiliations
Contributions
M.A.R., M.A.N., R.D.D., and L.Z.N. conceived and designed the work, discussed the results, and revised the manuscript. L.Z.N., R.D.D., P.A.A., and A.M. collected the data and supported all the related aspects of field acquisition of data. L.Z.N. analyzed the data. D.N. analyzed and helped with the interpretation of the chemical ecology results. L.Z.N. and R.D.D. wrote the paper.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Zamora-Nasca, L.B., Dimarco, R.D., Nassini, D. et al. Sheep feeding preference as a tool to control pine invasion in Patagonia: influence of foliar toughness, terpenoids and resin content. Sci Rep 10, 12113 (2020). https://doi.org/10.1038/s41598-020-68748-y
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-020-68748-y
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
