
Abstract
Animals can gain significant advantages from abilities to detect cues from predators, assess risks, and respond adaptively to reduce the likelihood of injurious interactions. In contrast, predator cue-induced changes in behaviour may interfere with fitness-associated activities such as exploration, foraging and reproduction. Despite the ecological importance of predator-prey interactions in insects, remarkably little is known about the abilities of insects to detect and respond to olfactory cues from predators, or the potential costs of such responses. We here demonstrate that a tephritid fruit fly, the Queensland fruit fly Bactrocera tryoni, is able to detect and respond differentially to volatile olfactory cues from four potential predators (three spiders and an ant) that vary in prevalence and diurnal activity. Male and female flies increased or decreased motility (velocity, active time, distance moved), or exhibited no change in motility, depending on which predator volatiles they encountered. Further, flies significantly reduced foraging, oviposition and mating propensity in the presence of volatiles from any of the predators. This study is the first report of predator-specific responses to olfactory cues in a tephritid fruit fly, and highlights that such anti-predator responses can impose costs on general activity and reproductive behaviour.
Similar content being viewed by others


Introduction
Direct physical encounters between predators and prey are the most apparent elements of predator-prey interactions. However, there are also myriad less visible ways in which predators and prey affect each other, modifying their behaviour in response to predatory opportunities and predation risks, respectively. Animals commonly emit or deposit characteristic chemical signatures that can be exploited as valuable sources of information by their predators. Diverse predators use olfactory cues to locate and identify prey. For example, the German wasp, Vespula germanica, uses the pheromone produced by male Mediterranean fruit flies as a cue to locate its prey1,2. Similarly, a Zodariid spider Habronestes bradleyi is attracted to alarm pheromones of its ant prey, Iridomyrmex purpureus3. On the other hand, prey species may detect olfactory cues from predators, and use this information to modify their behaviour to reduce risks of attack4,5,6. Such ‘non-consumptive effects’ of predators on prey behaviour are crucial for understanding ecological structure of predator-prey interactions7. Although numerous studies exist for olfaction-mediated indirect effects of predators on prey in vertebrates and aquatic systems8,9,10,11,12, much less is known about terrestrial insect systems13,14.
Responses of prey species to cues from their predators have been investigated in several insect systems. Aphid colonization is decreased in the presence of contact chemical cues from Coccinella septempunctata ladybirds15. Similarly, Colorado potato beetle larvae, Leptinotarsa decemlineata, reduce feeding in the presence of predator cues14. Striped cucumber beetles, Acalymma vittatum, reduce feeding and movement in the presence of tactile and visual cues from wolf spiders16. Most studies of non-consumptive effects in insect systems have focused on sensory modalities other than olfaction and have reported effects of cues from a single predator. However, insects may be attacked by diverse predators, each of which may present different risks and may have a characteristic chemical signature17. There are numerous contexts within which responses to olfactory cues from predators, and trade-offs, might be anticipated. Motility is important for most insects18, and inhibition of motility can have deleterious impacts on fitness. Because of differences amongst predators in sensory abilities and behaviour, prey species might move more quickly, or slower, or freeze, depending on the type of predator they encounter4.
Foraging is vital for survival and reproduction, but animals may be confronted with the challenge of acquiring food under increased risk of predation. Such conflicts in fitness currencies result in a trade-off to which prey may respond to predator cues by reducing foraging activities19,20. Mating is commonly associated with increased predation risk21. Sexual signals not only broadcast an individual’s location or quality to potential mates, they can also alert predators to the location of potential prey1,22,23. Oviposition in insects is often a complex process that involves active site-selection and egg laying that may make females more vulnerable to detection or attack. Insect eggs are easy targets for predators, and so female insects may appraise the suitability of an oviposition site in order to reduce predation risks for their offspring24. Although there are many studies investigating oviposition behaviour in the presence of predators or other dangers25,26, the effects of olfactory cues from predators on oviposition by insects are little known. Given differences among predators in the risks posed to prey species, and in prey responses that might mitigate risk, understanding the effects of olfactory cues from diverse predators is essential to progress beyond understanding prey response in single predator-prey systems.
In this study, we assess the ability of predator-naïve Queensland fruit flies, Bactrocera tryoni (Froggatt) (Tephritidae), to detect and respond to olfactory cues from four taxonomically and ecologically diverse cursorial predators and highlight the impact of their responses on fitness-related behaviours, including motility, feeding, and oviposition. Bactrocera tryoni is highly polyphagous, developing in more than 100 different fruits, and is widespread in Eastern Australia, having dispersed far beyond its pre-European range over the past century and also having established in some Pacific islands27,28. Tephritid fruit flies are ecologically and economically important in most tropical and warm temperate regions of the globe, and yet very little is known about their relationships with predators or their abilities to detect and respond to predation risk. The present study is an important step in advancing understanding of predator-prey interactions in tephritid flies, as well as in insects more broadly.
Results
Flies alter motility when exposed to olfactory cues from predators
In the presence of olfactory cues from predators, non-predators, and filtered air controls female and male flies exhibited significant variation in (i) mean velocity (Females χ2 = 149.3, df = 29, P < 0.0001; Males χ2 = 120.5, df = 29, P < 0.0001; Fig. 2A,B), (ii) active time (Females χ2 = 80.2, df = 29, P < 0.0001; Males χ2 = 81.4, df = 29, P < 0.0001; Fig. 2C,D), and (iii) distance moved (Females χ2 = 153.6, df = 29, P < 0.0001; Males χ2 = 140.4, df = 29, P < 0.0001; Fig. 2E,F). Compared to flies exposed only to filtered air, female and male flies significantly increased velocity, active time, and distance moved in the presence of olfactory cues from H. minitabunda and O. smaragdina, but showed significantly reduced velocity, active time and distance moved when exposed to olfactory cues from C. robusta (Fig. 2). In contrast, velocity, active time and distance moved of female and male flies in the presence of olfactory cues from O. quadratarius and a non-predator, P. affinis, were not significantly different from control (Fig. 2).

Distribution and diurnal activity of predators, prey and non-predator. (A) Helpis minitabunda a tropical/sub-tropical spider and is mostly found in the north eastern region of Australia. It is a diurnal jumping spider (depicted with yellow sun). (B) Opisthoncus quadratarius is a temperate spider found towards the south eastern region of Australia and is a diurnal jumping spider. (C) Clubiona robusta is a cosmopolitan clubionid spider with a wide range. It is present throughout Australia and is nocturnal (depicted with a blue moon). (D) Oecophylla smaragdina is commonly known as the green tree ant and is mostly present in the northern regions of tropical Australia. Workers are active in the day as well as night (depicted with both yellow sun and blue moon). (E) Plautia affinis, the green stink bug is common in gardens and orchards of eastern Australia. It is an herbivorous sap-feeding non-predator and is diurnal. (F) Bactrocera tryoni is a fruit fly native to eastern Australia and is also known as the Queensland fruit fly. Previously its range was limited to north eastern Australia, but its range has expanded since European colonisation. Distribution data are from atlas of life, Australia (atlasoflife.org.au). Illustrations are by V.K.

Motility of B. tryoni exposed to predator olfactory cues. Flies either increased or decreased motility, measured by flies (A,B) velocity, (C,D) active time and (D,E) distance moved, depending on the predator that cues were from. Differences across the set of treatments was analysed by Kruskal-Wallis test (P < 0.0001, see text) followed by Dunn’s multiple comparison test against filtered air control. Asterisk denotes significant difference from control and whiskers denote the max and min. FA = Filtered Air; PA = Plautia affinis; HM = Helpis minitabunda; OQ = Opisthoncus quadratarius; CR = Clubiona robusta; OS = Oecophylla smaragdina.
Flies decrease foraging when exposed to olfactory cues from predators
Number of visits to a food zone by female and male flies varied significantly in the presence of olfactory cues from predators, non-predators, and filtered air controls (Females χ2 = 111.3, df = 29, P < 0.0001; Males χ2 = 113.5, df = 29, P < 0.0001; Fig. 3A,B). Both female and male flies made fewer visits to the food zone when exposed to olfactory cues from the predators H. minitabunda, O. quadratarius, C. robusta, and O. smaragdina compared to flies exposed only to filtered air. However, number of visits to food zones in the presence of olfactory cues of non-predator P. affinis was not significantly different from control. Time spent in the food zone by female and male flies varied significantly in the presence of olfactory cues from predators, non-predators and filtered air controls (Females; χ2 = 124.1, df = 29, P < 0.0001; Males χ2 = 131.7, df = 29, P < 0.0001; Fig. 3C,D). Reflecting results for number of visits to the food zone, both female and male flies spent less time in the food zone when exposed to olfactory cues from H. minitabunda, O. quadratarius, C. robusta, and O. smaragdina compared to flies exposed only to filtered air. However, amount of time in the food zone in the presence of olfactory cues of non-predator P. affinis was not significantly different from control.

Foraging by B. tryoni exposed to predator olfactory cues. Flies decreased foraging, measured by (A,B) visits and (C,D) time spent. Differences across the set of treatments was analysed by Kruskal-Wallis test (P < 0.0001, see text) followed by Dunn’s multiple comparison test against filtered air control. Asterisk denotes significant difference from control and whiskers denote the max and min. FA = Filtered Air; PA = Plautia affinis; HM = Helpis minitabunda; OQ = Opisthoncus quadratarius; CR = Clubiona robusta; OS = Oecophylla smaragdina.
Flies reduce oviposition in the presence of olfactory cues from predators
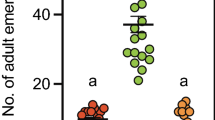
Differences were found in the number of eggs laid by B. tryoni in the presence of olfactory cues of predators, a non-predator, and filtered air (χ2 = 116.1, df = 29, P < 0.0001). Female B. tryoni laid significantly fewer eggs in the presence of olfactory cues from all tested predators compared to control (Fig. 4). Female B. tryoni laid 106.6 ± 12.30 eggs (mean ± s.e.m) in the presence of filtered air (control), but laid significantly fewer eggs in the presence of olfactory cues from H. minitabunda (9.1 ± 3.6 eggs), O. quadratarius (23.60 ± 5.04 eggs), C. robusta (7.96 ± 2.31 eggs), O. smaragdina (2.76 ± 1.61 eggs); however, number of eggs laid by flies in the presence of cues from non-predator P. affinis (78.90 ± 10.24 eggs) was not significantly different from control.

Oviposition by B. tryoni exposed to predator olfactory cues. Flies decreased overall oviposition in the presence of predator cues. Differences across the set of treatments was analysed by Kruskal-Wallis test (P < 0.0001, see text) followed by Dunn’s multiple comparison test against filtered air control. Asterisk denotes significant difference from control and whiskers denote the max and min. FA = Filtered Air; PA = Plautia affinis; HM = Helpis minitabunda; OQ = Opisthoncus quadratarius; CR = Clubiona robusta; OS = Oecophylla smaragdina.
Flies do not mate when exposed to predator cues
When exposed only to filtered air all 30 tested pairs mated, attempting copulation 4.23 ± 0.23 times before success and copulating for 63.72 ± 10.23 min (mean ± s.e.m). In the presence of non-predator olfactory cues, 20 of the 30 pairs mated (67%, a significant reduction compared with filtered air; Fishers exact test P < 0.001), attempting copulation 6.34 ± 1.34 times before success (not significantly different from filtered air; Welch ANOVA F1,31.99 = 3.285, P = 0.079) and copulating for 43.45 ± 7.34 min (not significantly different from filtered air; Welch ANOVA F1,47.29 = 2.061, P = 0.158). In contrast, no flies were observed to attempt copulation in the presence of olfactory cues from any of the predators (a significant reduction compared with both filtered air and non-predator olfactory cues; for both Fishers exact test P < 0.001 in all comparisons) and all flies spent the vast majority of time standing still. Although not quantified, male calling behaviour (rapid wing fanning)29 was commonly observed in the presence of filtered air or non-predator olfactory cues, but was never observed in the presence of predator olfactory cues.
Discussion
This study demonstrates very substantial effects of predator olfactory cues on motility, foraging, oviposition and mating activity of B. tryoni, as well as comparatively modest effects of non-predator cues on oviposition and mating. Olfactory cues from predators increased or decreased fly motility, depending on predator species. Specifically, B. tryoni did not modify motility level in the presence of olfactory cues from the spider O. quadratarius (or from the non-predator, P. affinis) but showed substantial changes in motility in response to cues from other spiders, H. minitabunda (increase), C. robusta (decrease), and an ant, O. smaragdina (increase). The lack of motility response to olfactory cues from O. quadratarius appears not to reflect a lack of detection, as cues from this species significantly inhibited foraging, oviposition and mating. The strong motility response to cues from H. minitabunda, C. robusta and O. smaragdina may reflect that these species are much more prevalent as leaf-dwelling predators in the ancestral range and habitat of B. tryoni, whereas O. quadratarius is less common in the ancestral range and is usually found on exposed tree trunks and branches of trees rather than on leaves (Fig. 1). Comparable ability to discriminate and respond differentially to cues from diverse predators has been reported in wood crickets and wolf spiders4,30, but not in tephritid fruit flies. Some vertebrate and insect species are known to respond to various predators based on particular predator traits, such as size, and commonness4,31. However, the present study appears to be the first to show that an insect, B. tryoni, can respond to olfactory cues of various predators with either an increase or decrease in motility. Previous studies have consistently found decreases in prey motility after detection of predator cues. This may be because these studies focused on nocturnal predators30,32,33,34,35. Nocturnal predators tend to rely less on vision, and more on substrate vibrations generated by moving prey to discern prey presence and location and this may have promoted a response of reduced motility when exposed to cues from nocturnal predators4. When in the presence of a nocturnal predator, decreased motility may increase the chances of remaining undetected36. However, decreased motility may be a less appropriate response in daytime by increasing vulnerability to visually orienting diurnal predators37,38. During the day, when the flies are able to rapidly decamp by visually oriented locomotion or flight, increased motility may be a more effective strategy to avoid non-flying predators31,39,40.
Bactrocera tryoni exhibited reduced number of visits and time spent at a food source in the presence of olfactory cues from predators. Reduced foraging activity has an obvious fitness cost of reduced acquisition of nutritional resources41. The absence of response to olfactory cues from the tested non-predator indicates that the response to predator cues does not reflect simply a general change in foraging behaviour in the presence of any olfactory cues. Instead, the reductions in foraging behaviour appear to reflect the danger indicated by specific olfactory cues from predators.
Animals may accrue substantial fitness advantages when they choose to oviposit in sites that increase the survival and growth of their offspring24,26. There is abundant evidence that many aquatic insects make oviposition choices based on cues that enable them to estimate predation risk, thus decreasing mortality of their offspring42. However, few studies in terrestrial insects are available. As predicted, female flies decreased oviposition in the presence of predator cues. Effects of both ant and spider volatiles likely reflect direct risks to ovipositing female flies. Oviposition requires a female to spend time at an oviposition site, likely increasing the chances of detection by a predator. Also, female flies may be less able to detect or escape from predators while ovipositing. Responses to olfactory cues from the ant O. smaragdina may also reflect that ants forage on fruit fly eggs and larvae in addition to adults43.
Olfactory cues from predators completely eliminated mating activity of B. tryoni. In some wolf spiders, males are known to wait longer to initiate mating in the presence of predator cues, thereby decreasing predation risk44. While this change in mating behaviour reduces predation risk, males employing this strategy often suffer reduced mating rates23. Only a handful of previous studies have reported differential responses of prey species to odors from different predators14,45 with no previous studies in tephritid flies. Chemical cues from predators that drive non-consumptive effects are poorly understood in insects (For an exception, see25). Some studies have reported that prey species respond to predator cues associated with consumed conspecifics, rather than endogenous predator cues10,19,46,47. In our experiments, predators were not fed B. tryoni, and hence the olfactory cues appear to be from the predators themselves. The flies used in our experiments were predator-naïve, and so the observed responses must be innate rather than learned.
This is the first detailed study of predator-specific responses to olfactory cues in a tephritid fly and is one of few such studies in terrestrial insects more broadly. Our findings indicate olfaction-mediated behaviour as a key for understanding predator-prey relations in tephritid flies, as well as for how tephritid fruit flies manage conflicts and trade-offs between fitness currencies. We found distinct responses to olfactory cues from each of the predators tested with B. tryoni and anticipate that distinct responses would also be evident for many other untested predators. We also anticipate that other tephritid flies will have comparable abilities to detect and respond to olfactory cues from their similarly diverse predators. The present study indicates that substantial advances in understanding the behavioural and chemical ecology of tephritid flies can be achieved by a new focus on the role of olfaction in predator-prey relations.
Methods
Study system
Queensland fruit fly, Bactrocera tryoni (Diptera: Tephritidae) (Fig. 1F), were obtained from a colony originating from central coastal New South Wales (29 generations from wild, at ~5,000 flies per generation) and maintained in a controlled environment laboratory at Macquarie University (25 ± 0.5 °C, 65 ± 5% RH, photoperiod of 11.5:0.5:11.5:0.5 light: dusk: dark: dawn). Adult flies were fed yeast hydrolysate, sugar and water ad libitum. Four native cursorial predators - three spiders and an ant – and one native non-predator were selected as sources of olfactory cues. The distribution and diurnal activity of predators, prey and non-predator are presented in Fig. 1. Three spiders, Helpis minitabunda (Salticidae, Fig. 1A), Opisthoncus quadratarius (Salticidae, Fig. 1B), Clubiona robusta (Clubionidae, Fig. 1C), and an ant Oecophylla smaragdina (Fig. 1D), were used as sources of predator olfactory cues. Female spiders were collected from December 2017 to March 2018 from orange orchards of NSW Department of Primary Industries, Somersby, NSW, Australia, and workers of O. smaragdina were obtained from August to October 2018 from trees around the Department of Agriculture and Fisheries, Mareeba, QLD, Australia. Spiders were maintained in the controlled environment laboratory (25 ± 0.5 °C, 65 ± 5% RH, photoperiod of 11.5:0.5:11.5:0.5 light: dusk: dark: dawn) for at least 1 month and were fed small insects 4 times each week. Ants were collected freshly from natural colonies as required. We also included a non-predatory stinkbug, Plautia affinis (Fig. 1E), and these were obtained from orange orchards in Somersby, NSW. The non-predator was used in this study as a positive control against the possibility that the flies simply respond in a generic way to olfactory cues from any insect or spider source. Female spiders were used in our study because they were more abundant than male spiders, while worker ants were selected as they are the colony members that forage and attack prey.
Olfactory cues from predators and non-predators
Olfactory cues from predators or non-predators were obtained by blowing charcoal filtered air over predators or non-predators into arenas. Briefly, a 50 mL closed glass volatile collection chamber (Sigma-Aldrich, USA), with an inlet and outlet, containing a single spider, a group of 6 ants, a non-predator, or no insect (filtered air control) was set-up 30 min before each experiment to allow a buildup of olfactory cues within the chamber. After 30 min, charcoal filtered air was passed through the chamber to carry olfactory cues from the volatile collection chamber into the test arenas using a gas sampling pump (KNF Pumps, Model no. NMP850.1.2KNDCB, Switzerland) at a rate of 1 L/min.
Arenas and software
We designed two kind of arenas for this study. A behavioural arena (Fig. S1A) was used for analysis of motility, foraging and mating, while an oviposition arena was used to assess oviposition behaviour. The behavioural arena comprised of a closed, clear polystyrene Petri dish (145 mm dia. x 20 mm deep). The Petri dish was covered on all sides with white lamination paper (100 mm high) to mitigate possible positional biases caused by external visual stimuli. The arena had 2 holes (5 mm dia.) on the sides for inlet and outlet of olfactory cues from insects or filtered air. Video recordings were carried out with an overhead HD camera (Go Video, Digital 540TLV) at recording speed of 25 frames per second. The arena was placed 1 m below the camera and was lit by fluorescent lights, although recordings of mating, which occurs at dusk48, were enabled using infrared lighting. The camera was connected to a digital video recorder and each recording was for 10 min.
The oviposition arena (Fig. S1B) was a cylindrical clear polystyrene jar (150 mm high x 90 mm dia.). Two holes (5 mm dia.) on the sides of the jar served as inlet and outlet for olfactory cues or filtered air. An Eppendorf tube (5 mL) with 10–12 holes (<1 mm dia.) on the upper half served as oviposition device. To stimulate egg laying, the oviposition device contained 1 mL of mango juice, which contains the known B. tryoni oviposition stimulant γ‐octalactone49. After each trial, the arena was washed with warm water, wiped with 70% ethanol and air dried for 20 min. Motility (active time, mean velocity, distance moved), time spent in zones, number of visits to zones and x, y coordinates of flies was quantified using Lolitrack Ver 4 (Loligo Systems, Denmark). Mating behaviour was recorded using BORIS V6.3.450. Thirty replicates were conducted for each bioassay for each predator.
Bioassays
For motility assays, a 10-day old virgin male or female fly was placed in the arena and was allowed to acclimatize for 20 min, after which one of the olfactory cues or filtered air was pumped into the arena through the inlet. Fly movement was recorded for 10 min.
The foraging assay was conducted following Zaninovich et al.51 with minor changes. A single virgin 10-day old male or female fly that had been provided water, but no food for 24 hours was introduced into the arena and allowed to acclimatize for 20 min. Next, filtered air or air containing one of the olfactory cues from predators or non-predator was pumped through the inlet of the arena for 1 min before dispensing 100 µL of sugar solution (commercial cane sugar, 10% w/v) using a micropipette onto the centre of the arena demarcated as ‘food zone’. The food zone consisted of a Petri dish (50 mm dia.) containing the sugar solution. The number of visits made, and time spent by flies in the food zone was recorded for 10 min and analyzed.
Mating assays were carried out at dusk, the normal mating time of B. tryoni. A pair of 15-day old virgin male and female flies were introduced into a behavioural arena 30 min before the onset of dusk. They were allowed to acclimatize for 20 min after which one of the olfactory cues or filtered air was pumped through the inlet of the arena. The activity of flies was recorded over-night and the video was analyzed using BORIS software. Number of copulation attempts and copula duration was recorded.
Oviposition assays were conducted using the oviposition arena. A single 15-day-old gravid female was introduced into the arena and was allowed to acclimatize for 20 min. An oviposition device was placed on the floor in the centre of the arena. Simultaneously, one of the olfactory cues or filtered air was pumped through the inlet of the arena. The flies were allowed to oviposit for ~16 h. The collected eggs were washed into a Petri dish and counted under a stereo microscope (MZ6, Leica Microsystems, Germany).
Statistics
Data for motility, foraging and oviposition assays did not conform to a normal distribution, and so were subjected to Kruskal-Wallis test followed by Dunn’s multiple comparision test. Statistics were calculated using GraphPad Prism version 7 for Mac (GraphPad Software, San Diego, CA, USA). Data for mating behaviour, including number of copulation attempts and copula duration, were subjected to Welch ANOVA. Data comparing number of flies that did and did not attempt copulation or mate in different treatment groups were subjected to Fishers Exact Tests.
Data availability
The datasets generated and analysed during this current study are available from the corresponding author on reasonable request.
References
Hendrichs, J., Katsoyannos, B. I., Wornoayporn, V. & Hendrichs, M. A. Odour-mediated foraging by yellowjacket wasps (Hymenoptera: Vespidae): predation on leks of pheromone-calling Mediterranean fruit fly males (Diptera: Tephritidae). Oecologia 99, 88–94 (1994).
Hendrichs, M. A. & Hendrichs, J. Perfumed to be killed: Interception of Mediterranean fruit fly (Diptera: Tephritidae) sexual signalling by predatory foraging wasps (Hymenoptera: Vespidae). Ann. Entomol. Soc. Am. 91, 228–234 (1998).
Allan, R. A., Elgar, M. & Capon, R. Exploitation of an ant chemical alarm signal by the Zodariid spider Habronestes bradleyi Walckenaer. Proc. R. Soc. B. 263, 69–73 (1996).
Binz, H., Bucher, R., Entling, M. H. & Menzel, F. Knowing the risk: Crickets distinguish between spider predators of different size and commonness. Ethology 120, 99–110 (2013).
Schmitz, O. Predator and prey functional traits: understanding the adaptive machinery driving predator–prey interactions. F1000Res. 6, 1767, https://doi.org/10.12688/f1000research.11813.1 (2017).
Poulin, R. X. et al. Chemical encoding of risk perception and predator detection among estuarine invertebrates. Proc. Natl. Acad. Sci. USA 115, 662–667 (2018).
Lima, S. L. Nonlethal effects in the ecology of predator-prey interactions. BioScience 48, 25–34 (1998).
Kats, L. B. & Dill, L. M. The scent of death: Chemosensory assessment of predation risk by prey animals. Écoscience 5, 361–394 (1998).
Dicke, M. & Grostal, P. Chemical detection of natural enemies by arthropods: An ecological perspective. Annu. Rev. Ecol. Syst. 32, 1–23 (2001).
Chivers, D. P. & Smith, R. J. F. Chemical alarm signalling in aquatic predator-prey systems: A review and prospectus. Écoscience 5, 338–352 (1998).
Preisser, E. L., Bolnick, D. I. & Benard, M. F. Scared To Death? The effects of intimidation and consumption in predator–prey interactions. Ecology 86, 501–509 (2005).
Werner, E. E. & Peacor, S. D. A Review of trait-mediated indirect interactions in ecological communities. Ecology 84, 1083–1100 (2003).
Ache, B. W. & Young, J. M. Olfaction: Diverse species, conserved principles. Neuron 48, 417–430 (1998).
Hermann, S. L. & Thaler, J. S. Prey perception of predation risk: volatile chemical cues mediate non-consumptive effects of a predator on a herbivorous insect. Oecologia 176, 669–676 (2014).
Ninkovic, V., Feng, Y., Olsson, U. & Pettersson, J. Ladybird footprints induce aphid avoidance behavior. Biol. Control 65, 63–71 (2013).
Williams, J. L. & Wise, D. H. Avoidance of wolf spiders (Araneae: Lycosidae) by striped cucumber beetles (Coleoptera: Chrysomelidae): Laboratory and field studies. Environ. Entomol. 32, 633–640 (2003).
Eklöv, P. Chemical cues from multiple predator-prey interactions induce changes in behavior and growth of anuran larvae. Oecologia 123, 192–199 (2000).
Nylin, S. Life history perspectives on pest insects: What’s the use? Austral Ecol. 26, 507–517 (2001).
Lima, S. L. & Dill, L. M. Behavioral decisions made under the risk of predation: a review and prospectus. Can. J. Zool. 68, 619–640 (1990).
Siepielski, A. M., Fallon, E. & Boersma, K. Predator olfactory cues generate a foraging–predation trade-off through prey apprehension. R. Soc. Open Sci. 3, 150537, https://doi.org/10.1098/rsos.150537 (2016).
Kemp, D. J. Costly copulation in the wild: mating increases the risk of parasitoid-mediated death in swarming locusts. Behav. Ecol. 23, 191–194 (2011).
Kemp, D. J. & Rutowski, R. L. A survival cost to mating in a polyandrous butterfly, Colias eurytheme. Oikos 105, 65–70 (2004).
Magnhagen, C. Predation risk as a cost of reproduction. Trends Ecol. Evol. 6, 183–186 (1991).
Damodaram, K. J. et al. Centuries of domestication has not impaired oviposition site-selection function in the silkmoth, Bombyx mori. Sci. Rep. 4, 7472, https://doi.org/10.1038/srep07472 (2014).
Ebrahim, S. A. M. et al. Drosophila avoids parasitoids by sensing their semiochemicals via a dedicated olfactory circuit. PLoS Biol. 13, e1002318, https://doi.org/10.1371/journal.pbio.1002318 (2015).
Stensmyr, M. C. et al. A conserved dedicated olfactory circuit for detecting harmful microbes in Drosophila. Cell 151, 1345–1357 (2012).
White, I. M. & Elson-Harris, M. M. 1994 Fruit flies of economic significance: their identification and bionomics (CAB International, 1994).
Clarke, A., Powell, K., Weldon, C. & Taylor, P. The ecology of Bactrocera tryoni (Diptera: Tephritidae): what do we know to assist pest management? Ann. Appl. Biol. 158, 26–54 (2010).
Mankin, R. W., Lemon, M., Harmer, A. M. T., Evans, C. S. & Taylor, P. W. Time-pattern and frequency analyses of sounds produced by irradiated and untreated male Bactrocera tryoni (Diptera: Tephritidae) during mating behaviour. Ann. Entomol. Soc. Am. 101, 664–674 (2008).
Persons, M. H., Walker, S. E. & Rypstra, A. L. Fitness costs and benefits of antipredator behavior mediated by chemotactile cues in the wolf spider Pardosa milvina (Araneae: Lycosidae). Behav. Ecol. 13, 386–392 (2002).
Caro, T. & Girling, S. Antipredator defenses in birds and mammals (University of Chicago Press, 2005)
Persons, M., Walker, S., Rypstra, A. & Marshall, S. Wolf spider predator avoidance tactics and survival in the presence of diet-associated predator cues (Araneae: Lycosidae). Anim. Behav. 61, 43–51 (2001).
Marshall, S. D., Pavuk, D. M. & Rypstra, A. L. A. Comparative study of phenology and daily activity patterns in the wolf spiders Pardosa Milvina and Hogna Helluo in soybean agroecosystems in Southwestern Ohio (Araneae, Lycosidae). J. Arachnol. 30, 503–510 (2002).
Storm, J. J. & Lima, S. L. Predator-naïve fall field crickets respond to the chemical cues of wolf spiders. Can. J. Zool. 86, 1259–1263 (2008).
Storm, J. J. & Lima, S. L. Mothers forewarn offspring about predators: A transgenerational maternal effect on behavior. Am. Nat. 175, 382–390 (2010).
Werner, E. E. & Anholt, B. R. Ecological consequences of the trade-off between growth and mortality rates mediated by foraging activity. Am. Nat. 142, 242–272 (1993).
Persons, M. H. & Uetz, G. W. The effect of prey movement on attack behavior and patch residence decision rules of wolf spiders (Araneae: Lycosidae). J. Insect Behav. 10, 737–752 (1997).
Clemente, C. J. et al. The visual system of the Australian wolf spider Lycosa leuckartii (Araneae: Lycosidae): visual acuity and the functional role of the eyes. J. Arachnol. 38, 398–406 (2010).
Nordlund, D. A. & Morrison, R. Handling time, prey preference, and functional response for Chrysoperla rufilabris in the laboratory. Entomol. Exp. Appl. 57, 237–242 (1990).
Hasson, O. Pursuit-deterrent signals: communication between prey and predator. Trends Ecol. Evol. 6, 325–329 (1991).
Godin, J. G. J. & Smith, S. A. A. Fitness cost of foraging in the guppy. Nature 333, 69–71 (1988).
Albeny-Simões, D. et al. Attracted to the enemy: Aedes aegypti prefers oviposition sites with predator-killed conspecifics. Oecologia 175, 481–492 (2014).
Fernandes, W. D., Sant’Ana, M. V., Raizer, J. & Lange, D. Predation of fruit fly larvae Anastrepha (Diptera: Tephritidae) by ants in Grove. Psyche 2012, 108389, https://doi.org/10.1155/2012/108389 (2012).
Fowler-Finn, K. D. & Hebets, E. A. The degree of response to increased predation risk corresponds to male secondary sexual traits. Behav. Ecol. 22, 268–275 (2011).
Ferrari, M. C., Wisenden, B. D. & Chivers, D. P. Chemical ecology of predator–prey interactions in aquatic ecosystems: a review and prospectus. Can. J. Zool. 88, 698–724 (2010).
Wisenden, B. D., Chivers, D. P. & Smith, R. J. F. Learned recognition of predation risk by Enallagma damselfly larvae (Odonata, Zygoptera) on the basis of chemical cues. J. Chem. Ecol. 23, 137–151 (1997).
Chivers, D. P., Wisenden, B. D. & Smith, R. F. Damselfly larvae learn to recognize predators from chemical cues in the predators diet. Anim. Behav. 52, 315–320 (1996).
Pike, N., Wang, W. Y. S. & Meats, A. The likely fate of hybrids of Bactrocera tryoni and Bactrocera neohumeralis. Heredity 90, 365–370 (2003).
Kempraj, V., Park, S. J. & Taylor, P. W. γ‐Octalactone, an effective oviposition stimulant of Bactrocera tryoni. J. Appl. Entomol. 143, 1205–1209 (2019).
Friard, O. & Gamba, M. BORIS: a free, versatile open-source event-logging software for video/audio coding and live observations. Methods Ecol. Evol. 7, 1325–1330 (2016).
Zaninovich, O. A. et al. A single-fly assay for foraging behavior in Drosophila. J. of Vis. Exp. 81, 50801, https://doi.org/10.3791/50801 (2013).
Acknowledgements
The authors thank Dr. Stefano De Faveri, Department of Agriculture & Fisheries, Mareeba, Queensland for providing lab space and equipment for work with green tree ants. This research was supported by funds from Australian Research Council Industrial Transformation Training Centre for Fruit Fly Biosecurity Innovation (Project IC150100026), including a PhD scholarship for V.K. and a Research Fellowship for S.J.P. This research received additional support from the SITplus collaborative fruit fly program. Project Raising Q-fly Sterile Insect Technique to World Standard (HG14033) is funded by the Hort Frontiers Fruit Fly Fund, part of the Hort Frontiers strategic partnership initiative developed by Hort Innovation, with co-investment from Macquarie University and contributions from the Australian Government.
Author information
Authors and Affiliations
Contributions
V.K. and P.W.T. designed the study, V.K. and S.J.P. conducted bioassays and other experiments. V.K. wrote the first draft of the manuscript and all authors contributed to the final version of the manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplemetary information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Kempraj, V., Park, S.J. & Taylor, P.W. Forewarned is forearmed: Queensland fruit flies detect olfactory cues from predators and respond with predator-specific behaviour. Sci Rep 10, 7297 (2020). https://doi.org/10.1038/s41598-020-64138-6
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-020-64138-6
This article is cited by
-
The greater wax moth, Galleria mellonella (L.) uses two different sensory modalities to evaluate the suitability of potential oviposition sites
Scientific Reports (2023)
-
1-Octanol emitted by Oecophylla smaragdina weaver ants repels and deters oviposition in Queensland fruit fly
Scientific Reports (2022)
-
Chemical Cues from Entomopathogenic Nematodes Vary Across Three Species with Different Foraging Strategies, Triggering Different Behavioral Responses in Prey and Competitors
Journal of Chemical Ecology (2021)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
