
Abstract
Nitrogen species such as ammonia and nitrite are considered as major stressors in modern aquaculture practices. We developed enrichments of ammonia oxidising bacteria (AOB) and nitrite oxidising bacteria (NOB) for effective mitigation of nitrogenous wastes in the shrimp culture operations. The objective of this study was to understand the microbial community composition of AOB and NOB enrichments using the V3-V4 region of the 16S rDNA gene by Illumina MiSeq sequencing. The analysis revealed 2948 and 1069 OTUs at 97% similarity index and Shannon alpha diversity index of 7.64 and 4.85 for AOB and NOB enrichments, respectively. Comparative analysis showed that a total of 887 OTUs were common among AOB and NOB enrichments. The AOB and NOB enrichment were dominated by Eubacteria at 96% and 99.7% respectively. Proteobacterial phylum constituted 31.46% (AOB) and 39.75% (NOB) and dominated by α-Proteobacteria (20%) in AOB and γ-Proteobacteria (16%) in NOB. Among the species in AOB enrichment (2,948) two sequences were assigned to ammonia oxidising bacterial group belonging to Nitrosomonas, and Nitrosococcus genera and two belonged to archaeon group comprising Nitrosopumilus and Candidatus Nitrososphaeraea genera. The NOB enrichment was predominated by Nitrospiraceae and Thermodesulfovibrionaceae. Further, the data revealed the presence of heterotrophic bacteria contributing to the process of nitrification and form microcosm with the AOB and NOB. PICRUSt analysis predicted the presence of 24 different nitrogen cycling genes involved in nitrification, denitrification, ammonia and nitrogen transporter family, nitrate reduction and ammonia assimilation. The study confirms the presence of many lesser known nitrifying bacteria along with well characterised nitrifiers.
Similar content being viewed by others


Introduction
Aquaculture is an important economic activity supplying quality animal protein, generating employment and providing foreign exchange. Fish and fishery products are the most traded food items in the world, and an estimated 45% of the fish produced enters the international market. In terms of value, shrimp/prawn is the second most traded item next only to salmon in the USD 152 billion global seafood market1. In the modern-day intensive and semi-intensive shrimp aquaculture, management of accumulating metabolic wastes, especially in zero water exchange systems has been a major challenge. Accumulation of nitrogenous wastes generated by animal excreta and degradation of uneaten feed leads to deterioration of culture environment and stress to farmed animals2,3,4. Ammonia is the primary end product of protein metabolism in most aquatic animals5 and is also produced following microbial decomposition of organic wastes. Increase in the levels of nitrogenous species in the shrimp haemolymph leads to reduced food intake, increased oxygen consumption, increased excretion of nitrogen, and altered protein concentrations cause moderate to high mortality6. Further, the ammonia (>5 ppm) and nitrite (>0.35 ppm) are known to affect immunity and enhance susceptibility to diseases. Nitrite, an intermediary in the nitrogen metabolism, transforms the haemocyanin to meta-haemocyanin, reducing its ability to transfer oxygen to the tissues leading to hypoxia and death7.
Nitrification is the biological oxidation of ammonia (NH3) to nitrite (NO2−) and then to nitrate (NO3−), is a major oxidative process involved in maintenance of global nitrogen cycle in aquatic systems8 and soil9. This process is predominantly carried out by two different classes of lithoautotrophic microbes, namely the ammonia-oxidising bacteria (AOB) (e.g., Nitrosomonas and Nitrosospira) and the ammonia-oxidising archaea (AOA) (e.g., Nitrososphaerea, Nitrosopumilus) as one guild that convert ammonia to nitrite. Key enzymes involved in the process are ammonia monooxygenase encoded by the amoCAB genes and hydroxylamine dehydrogenase encoded by the haoAB genes10. The nitrite-oxidising bacteria (NOB) (e.g., Nitrospira and Nitrobacter) convert nitrite to nitrate mediated by the enzyme nitrite oxidoreductase encoded by nxrAB genes11. Recent studies have shown that nitrification is also carried out by heterotrophic bacteria which are commonly called as heterotrophic nitrifying bacteria12,13,14; however, they are lesser efficient than the autotrophs15.
In the intensive aquaculture system, sustainable management of toxic nitrogenous metabolites plays a crucial role in assured production. Generally, these metabolites are controlled by the application of adsorbents like Yucca extract and zeolite16,17 or by microbial interventions. The utility of adsorbents is restricted to physical removal of the ammonia build up in the system and needs repeated application. On the other hand, a recent study doubts the efficiency of mixotrophic microbes in effective mitigation of nitrogenous wastes in aquaculture18. However, factors like microbial diversity, salinity tolerance, conversion efficiency, growth rate and enrichment could influence the performance of formulated microbial consortia. Though AOB and NOB are efficient in nitrification, their slow-growing nature has been a hindrance in formulating a microbial product for bioremediation19,20. We have developed a microbial enrichment with nitrification property and the efficacy was tested in the laboratory and brackishwater aquaculture farms. The objective of this paper is to understand the microbial diversity of the enrichments developed in the laboratory using 16S rDNA sequencing analysis.
Results and Discussion
AOB and NOB cultures were enriched from the pond sediments under autotrophic conditions with ammonia and nitrite as a sole energy source. For determining in vitro activity of the consortia, the established enrichments were spiked with known concertation of ammonia (19 ppm) and nitrite (16 ppm) as described in methods. The AOB enrichment efficiently oxidised ammonia to nitrite in 10 days while NOB consortium oxidized nitrite to nitrate in 14 days (Fig. 1). Though non-specific isolation media is available, it is a challenging effort to isolate slow-growing nitrifying bacterial species in pure form and are maintained as enrichments. Our enrichments being highly efficient in the oxidation of ammonia and nitrite, in the present study, 16S rRNA high throughput sequence methods were used for knowing the bacterial diversity in the AOB and NOB consortia, and the results revealed the presence of several unclassified lineages in these enrichments.
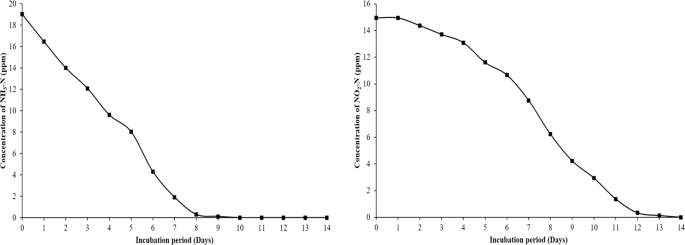
In-vitro Ammonia oxidation rate (a) and Nitrite oxidation rate (b) by AOB and NOB enrichments.
Diversity of microbial communities
In total, approximately 488,402 bacterial sequence tags from AOB enrichments and 384,579 bacterial sequence tags from NOB enrichments for 16S rDNA with an average length of 2 × 300 bp were obtained. Following sub-sampling, a total of 2,948 OTUs for AOB and 1,069 OTUs for NOB consortia respectively were obtained at 97% similarity index. Shannon alpha diversity index was found to be 7.64 in AOB enriched culture and 4.85 in NOB consortia confirming rich microbial diversity among the enrichments (Table 1). Comparative analysis showed that a total of 887 (21.92%) OTUs were common among AOB and NOB consortia (Figs. 2, S1).

Venn diagram showing number of common OTUs between AOB and NOB consortia.
Relative abundance of bacterial community
With 16S rDNA high throughput sequencing, nearly 4,000 OTUs were recovered from the two-distinct nitrifying bacterial enrichments. Altogether a total of 47 bacterial phyla were observed in both the enrichments. Eubacteria dominated both AOB and NOB consortia (96% and 99% respectively) with remaining taxa belonging to Archaea. The proteobacterial phylum was found to be dominant in both the enrichments, with 31.46% and 39.75% in AOB and NOB consortia respectively. Additionally, Acidobacteria, Actinobacteria, Bacteroidetes, Chloroflexi, Cyanobacteria, Firmicutes, Nitrospira and Planctomyceteswere were the other phyla present in both AOB and NOB enrichments. Among the proteobacterial phylum, α-Proteobacteria dominated in AOB (20%) and NOB (18%) consortia while γ proteobacteria constituted 16% and 8% in NOB and AOB consortia respectively (Figs. 3a, 4).

Phylum (a) and genus (b) level distribution among AOB and NOB enrichments.

Dominant taxa present in AOB and NOB consortia at the phylum level.
Taxonomic composition of microbial enrichments
Microbial communities in ammonia oxidising enrichment
A total of 2,948 species were observed in AOB consortia, out of which two sequences were assigned to AOB group belonging to Nitrosomonas, and Nitrosococcus genera and two belong to archaeon group comprising Nitrosopumilus and Nitrososphaera genera. Of the OTUs belonging to AOB guild, 20 OTUs were identified as Nitrosomonas marina. The enrichments also contained 107 OTU s of Nitrosococcus genera (Fig. 5), represented under unclassified species clade. Among the ammonia oxidising archaea group, eight OTUs belong to Nitrosopumilus comprising unclassified species and four OTUs belonging to Nitrososphaeraea comprising the Candidatus Nitrososphaeraea gargensis (Fig. 5). The genus Stanieria belonging to the group of methyl ammonia oxidiser community were also found (four OTUs). Similarly, 269 OTUs showing similarity to Nitrospira-like AOB organisms were observed in the consortia. Genera with lesser abundance belonging to Psychrobacter, Bacillus (two OTUs), Hyphomicrobium (two OTUs), Methanosaeta (two OTUs), Sediminibacterium (one OTU), Pseudomonas (four OTUs) and Candidatus species (six OTUs) were recorded (Fig. 3b, Table S1).
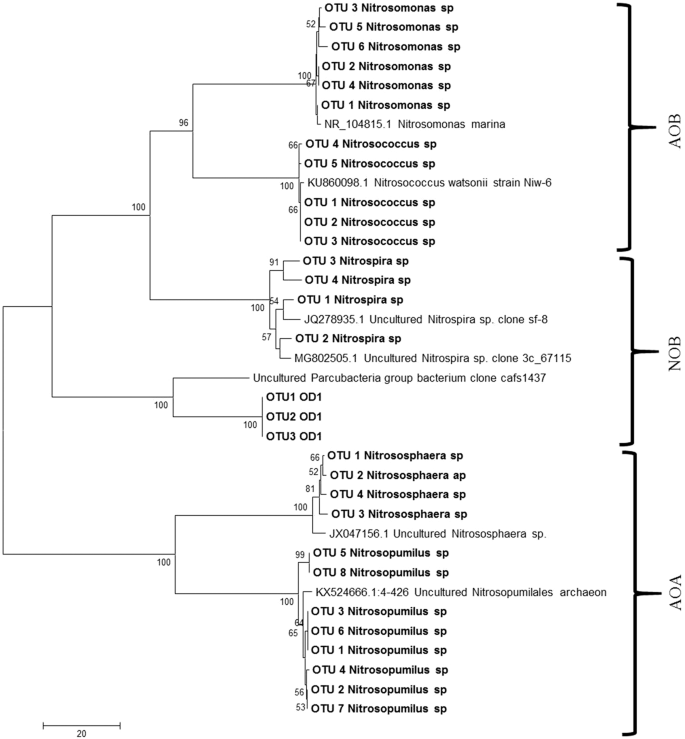
Phylogenetic analysis of representative sequences of Ammonia oxidising bacteria, archaea and nitrite oxidising bacteria in AOB and NOB enrichments.
In our study, two types of aerobic autotrophic microorganisms were observed in the AOB enrichments such as Nitrospira and Nitrosomonas that are known to be major players of ammonia oxidation. Autotrophic microbes can grow till exhaustion of nutrients and remain dormant for a certain period of time. Though heterotrophic microbes can grow five times faster than autotrophs, the latter group of microbes have significantly higher nitrification efficiency21. The AOB strains obtained in our study were assigned to the Nitrosomonas lineage (Fig. 5), confirming with the previous studies that these bacteria in this lineage can survive in low ammonia concentration, in soils9, lake sediments22 and biofilters23. Apart from ammonia oxidising bacterial population, archaea bacterial population were also detected in the developed enrichments. The majority of AOA classified till now were found to be oligotrophic and grow under low ammonia concentration. Gao et al.24 showed that AOB were more competitive than AOA under high concentration of ammonia. Similarly, substrate inhibition of archaeal nitrification under high concentration of ammonia has been reported25,26,27,28. Distribution of AOA is also ecosystem dependent, for example, Ca. Nitrosopumilus is widely present in seawater29 while Ca. Nitrososphaera more abundant in soil habitats with a wide range of pH and also in different aquatic ecosystems30. The results obtained in this study also showed lower diversity of AOA than AOB confirming with the previous studies. Phylogenetic analysis of archaea bacterial sequences were clustered into Nitrosopumilus and Nitrosophaera group and showed a distinct lineage towards ammonia oxidising bacterial groups (Fig. 5).
From our findings, the majority of microbes identified were from Proteobacteria phylum that is widely present in natural environments and plays a vital role in nutrient cycling and mineralisation of organic compounds31. Nearly 62% of the identified sequences were found to be unclassified genera (Fig. S2) indicating there might be novel microbial genera involved in ammonia oxidation or involved in aiding ammonia oxidation process. The dominant genera of ammonia-oxidising nitrifiers such as Nitrosomonas sp., Nitrosomonaseuropea and Nitrospira sp. have been documented in this study are responsible for efficient nitrification. Members of the group Nitrospira which is known to be a complete ammonia oxidizer32 is also found in our enrichments.
The Limnohabitans spp. present in AOB consortia is well-studied opportunistic bacterial group, with rapid generation time, acting on low molecular weight dissolved organic matter33. A recent study showed Limnohabitans species, strains Rim28 and Rim47 had great metabolic versatility, including photosynthesis, autotrophic carbon fixation, and ammonium and sulphur oxidisation34. Fitzgerald et al.35 reported the involvement of Luteibacter spp. of Xanthomonadaceae family in autotrophic utilisation of ammonia as a sole source of nitrogen in the low DO-nitrification process. Along with Nitrosomonas, Nitrosopumilus and Limnohabitans in AOB consortia these groups can also play a role in oxidising ammonia into nitrite.
Heterotrophic bacterial cultures like Bacillus, Arthrobacter, Pseudomonas and Exiguobacterium belonging to various phyla such as Firmicutes, Actinobacteria and Proteobacteria were observed in the enrichments and may not specifically be ammonia-oxidising nitrifiers; probably from the source water used for raising the enrichment. The results obtained in our study show that the enrichments consist of bacteria having mixotrophic ammonia oxidation and are well corroborated with the previous studies36 (Fig. S3).
Microbial communities in nitrite-oxidising enrichments
A total of 1,069 OTUs were observed in the NOB enrichments of which 184 OTUs belonging to phylum Nitrospirae were detected predominantly, two families belonging to Nitrospiraceae and Thermodesulfovibrionaceae. Under Nitrospiraceae 56 OTUs belong to unclassified Nitrospira genera whereas, 36 OTUs of Thermodesulfovibrionaceae families were distributed into three unclassified genera. NOB enrichments had 60% of unclassified sequences (Fig. S2). One of the interesting findings in NOB consortia is the presence of superphylum OD1. A total of 17,961 sequences belonging to the phylum OD1 has been classified as superphylum Candidate Phyla Radiation (CPR) Parcubacteria (Fig. 5) group which was identified as a major player in marine nitrogen cycle37.
In this study, we have documented a diverse lineage of Nitrospirae phylum known to play a major role in the nitrite oxidising process. To the best of our knowledge, a few chemolithoautotrophic microbes with nitrite oxidation have been documented. Most of these groups were previously documented to play a major role in nitrite oxidation and also involved in the denitrification process with key genes involved (Table S1). For the nitrite oxidisers, eight OTUs related to Nitrospira were detected in the enrichment. Nitrospira (Fig. 4) is identified as the most abundant nitrite oxidisers in low-nitrite environments38,39. A poorly characterised taxon, 0319-6A21 (Table S1) belongs to Nitrospira was first reported from lava caves than in surface soils40. Organic compounds produced by both AOB and NOB serve as a substrate for heterotrophic microbial growth41 whereas, a Nitrospira population can feed on the dead heterotrophic community during the enrichment in mineral nitrite medium42. NOB may also benefit from the presence of heterotrophs, for example, via heterotrophic nitrate reduction.
The previous study has demonstrated that members of Proteobacteria can utilise reduced sulphur compounds to obtain energy and also through carbon assimilation via the reductive tricarboxylic acid (rTCA) cycle43. Interestingly rare taxa <1% belong to Armatimonadetes, AC1, NC10, NKB19, OP8 and GN04 (Table S1) were detected in both the enrichments. Presence of NC10 group in both the samples suggests their involvement in methane oxidation coupled with nitrification process44,45 (Fig. S3). Although these groups were found to be present in less number, previous reports have also predicted their involvement in nitrogen cycle coupled to some other biogeochemical pathways.
Heterotrophic nitrate-reducing bacterial communities
The traditional theory of biological nitrogen removal makes a rigid difference between nitrification and denitrification process based on distinct growth conditions of nitrifiers and denitrifiers; however, heterotrophic nitrification and aerobic denitrification processes can simultaneously occur46,47. The potential for nitrification and denitrification was detected in both autotroph and heterotroph microbial lineages, suggesting the involvement of a diverse range of nitrogen metabolic pathway. Heterotrophic nitrifying bacteria produce hydroxylamine, nitrite and nitrate by nitrification process using organic carbon as a source for their growth. Most of these bacteria are capable of converting nitrification products directly to nitrogen gas through the process of aerobic denitrification48. In this study other than autotrophic microbes, heterotrophic microbes were also predominantly present in both the enrichments. Proteobacteria were the most critical contributors of all genes involved in denitrification pathway, besides the presence of other groups like Nitrospirae, Bacteroidetes, and uncultured microorganisms were shown to play essential roles in denitrification. The dominant denitrifiers in our enrichments mainly include Alcaligenes, Pseudoxanthomonas, Pseudomonas, Marinobacter, Shewanella, Thalassospira and Rhodobacter (Table. S1), most of which are taxonomically affiliated with ß or γ proteobacteria and bacteroidetes49. Proteobacterial groups, found in both the enrichments, were closely related to Hyphomicrobium, Paracoccus, Pseudomonas and Comamonas spp. (Table S1, Fig. S3) and known to be denitrifiers50. Studies by Feng et al.51 showed that bacteria belong to Bacillus, P. putida, P. stutzeri, Hydrogenophaga, and Achromobacter have been shown to have nitrification and aerobic denitrification abilities
The heterotrophic genera identified in AOB enrichment belong to Burkholderia, Exiguobacterium, Methanoregula and Methanosaeta while, groups belonging to Aequorivita, Candidatus Entotheonella, Erythrobacter, Flavobacterium, Idiomarina, Mesorhizobium, Oleibacter, Parvibaculum were present in NOB enrichments. Groups belonging to Alcanivorax, Anaerospora, Arenibacter, Bacillus, Bradyrhizobium, Brevibacterium, Brevundimonas, Candidatus Solibacter, Desulfovibrio, Devosia, Halorhodospira, Hyphomicrobium, Lewinella, Planctomyces, and Sphingobium were present in both AOB and NOB enrichments (Table S1, Fig. S3). These groups present in enrichments were classified based on their predicted functional properties such as denitrification, nitrification and nitrate reduction using KEGG orthology database (Fig. 6, Table S1). In this study, a broader diversity of metabolic genes involved in nitrogen metabolism, viz. nitrification, denitrification, nitrogen fixation, and dissimilatory nitrate reduction to ammonia (DNRA) were predicted using PICRUSt analysis in both the AOB and NOB enrichments.

Heat map showing the distribution of genes involved in nitrogen cycle based on KEGG orthologous gene identification by PICRUSt analysis.
Many OTU’s belong to yet to be classified group described as Candidatus species were also found to be predominant in both AOB and NOB consortia. Based on recent findings, AOB and NOB enrichments contain bacterial isolates belonging to super-phylum Verrucomicrobia, which are involved in methane regulated pathway52 were documented in both the enrichments. Some of the notable groups, such as Natronococcus, known for nitrate reduction were present in both the enrichments. Other Candidatus groups belong to unclassified Lautropia, phylum BRC2, Koribacter, Solibacter, Aquiluna, and Entotheonella (Table S1)12,53 were also found to be denitrifier strains. Few groups belong to Methanoregula, and Candidatus Hydrogenedens54 were found to convert nitrate to ammonia through dissimilatory nitrate reduction pathway.
16S rDNA high throughput gene sequencing indicated that the nitrifying populations in the enrichment involve a Nitrosomonas-like Nitrospira-like AOB, Nitrosopumilus, Candidatus Nitrososphaera and Nitrospira-like NOB that are adapted to ammonia and nitrite condition, respectively. Previous studies have also reported the heterotrophic nitrification and ammonification pathway present in these groups, but their mode of function remains less known. Based on preferential enrichment, and the reports from existing literature, organisms belong to Pseudomonas, members of the family Xanthomonadaceae, Limnohabitans, and Sphingomonas, CPR OD1, Candidatus species of Opitutus, Staneria, and Exiguobacterium group have the potential to participate in ammonia as well as nitrite oxidation, can function either as heterotrophic nitrifiers, or via autotrophic nitrification through yet uncharacterized pathways33,35,37. The potential for nitrification and denitrification was detected in a diverse lineage of both autotroph and heterotroph microbial communities, suggesting a diverse range of potential involved in nitrogen metabolism. One such group observed in the enrichment is Sulfurimonas, which reduces nitrate to dinitrogen gas coupling sulphur oxidation to denitrification pathway has been previously documented by Cerqueira et al.55.
Presumptive functional profile of microbes in the enrichments using PICRUSt analysis
PICRUSt was used to predict the function of enrichments based on 16S rDNA and the functionalities obtained are predictive. The analysis predicted 24 different nitrogen cycling genes that are involved in nitrification, denitrification, ammonia and nitrogen transporter family, nitrate reduction and ammonia assimilation (Fig. 6, Table S1). Genes coding for the enzyme involved in nitronate monooxygenase, nitrile hydratase, nitrate reductase, nitrilase, nitric oxide dioxygenase, nitric oxide reductase, nitric-oxide synthase, nitrite reductase, nitric-oxide reductase, nitrogenase, nitric nitrogen fixation protein, nitroreductase/dihydropteridine reductase, nitrous-oxide reductase, nitroreductase, nitrate reductase, nitrogenase, nitric oxide reductase, which are distributed among several phyla were predicted. Most dominant gene cluster involved in nitrification, denitrification, DNRA and assimilatory nitrogen reduction is given in Table (S1). Other than nitrogen metabolism, metabolic pathways related to sulphur, carbon, propionate, TCA cycle, amino acid and sugar metabolism, biodegradation and carbon fixation pathways and pathways associated with the degradation of aromatic compounds, nitrotoluene degradation and polyaromatic hydrocarbon degrading groups were also predicted to be present in both the enrichments (Figs. 7 and S4). However, it is to be noted that use of function prediction tools relies mainly on the availability of reference genomes included in the algorithm56 and the predicted functions are indicative, which needs further confirmation.

The general metabolic pathways of AOB and NOB consortiaby network enrichment analysis.
Due to the presence of many unclassified genera in the enrichment, this study suggests that heterotrophic bacterial group documented in the enrichment may play a role in nitrogen conversion. From the findings, we believe presence of heterotrophic bacteria along with autotrophic bacterial groups and form a microcosm57. This may be due to production of the secondary metabolites, including nitrogenous compounds by autotrophs facilitating survival of the heterotrophic bacteria in the enrichments. Recently nitrification activity has been observed in heterotrophic bacteria, which are capable of utilising ammonia and nitrite33,36,47. Although nitrifying bacteria may act as primary producers and play a significant role in the nitrogen cycle, little is known about its adaptability with heterotrophic microbial community and mode of interactions.
Conclusion
The AOB and NOB enrichments developed in our laboratory had efficient nitrification potential. The study has documented the microbes involved in the process of nitrification in the AOB and NOB enrichments. In addition to well-known autotrophs like Nitrosomonas, Nitrosococcus, Nitrospira, Nitrosopumilus, Nitrososphaera; many Candidatus species, heterotrophic bacteria such as Marinobacter, Paracoccus, Burkholderia, Exiguobacterium, Alcaligenes, Pesudomonas, Hyphomicrobium, Caldilinea, Thalassospira and Rhodobacter etc were observed in the enrichments. The present study also documented the presence of diverse lineage of phylum OD1 showing the importance of this group in the nitrogen cycle. The enrichments, beside containing ammonia and nitrite oxidising autotrophs were found to be associated with heterotrophic microbial groups involved in nitrification, denitrification and establishing a microcosm with DNRA and assimilatory nitrate reduction.
Materials and method
Microbial enrichments
Sediment samples (n = 66) were collected from shrimp aquaculture ponds (n = 15), having salinity 18–48 ppt from coastal regions of India. About 5 g from each sediment samples were aseptically inoculated into sterile Koops58 medium for AOB cultivation and Watson and Waterbury59 medium for NOB cultivation with pH 7.5–8.0 and incubated at 30 °C in an orbital shaker at 120 rpm for 24 days, and the culture has been continuously enriched for six months. Among the samples processed two enrichments were found to show promising activity and the efficient AOB and NOB enrichments were pooled separately. To understand the AOB and NOB oxidation rate, enrichments were spiked with a known concentration of ammonia (19 ppm) and nitrite (16 ppm). Daily sampling was conducted to analyse ammonia (by Nessler’s reagent60 at 405 nm) and nitrite (by Sulphanilamide, N-ethylene- di hydrochloride61 at 540 nm) oxidation rate.
Preparation of MiSeq library
DNA was extracted from the AOB and NOB consortia by phenol: chloroform extraction protocol from Ghosh et al.62. The amplicon libraries were prepared using Nextera XT index kit (Illumina Inc. USA) as per metagenomic sequencing library preparation protocol. Primers for amplification of the 16S rDNA gene-specific for bacterial V3–V4 were designed at Eurofins Genomics and Bioinformatics laboratory (Bangalore, India) with the sequence as 341F 3′-GCCTACGGGNGGCWGCAG-5′ and 806R 3′-ACTACHVGGGTATCTAATCC-5′. Samples from two batches of culture for each of AOB and NOB were analysed for confirmation of the data.
Data analysis
The raw sequencing data were processed using the QIIME pipeline v1.7.0 to obtain the high-quality tags and chimera checked using UCHIME algorithm against the reference database (Gold database) to generate clean data. Then the effective tags from all samples were clustered into operational taxonomic units (OTUs) using QIIME with 97% similarity. Taxonomy was assigned to the representative sequences using the RDP Classifier. The following diversity analyses were determined in QIIME (v1.7.0). The phylogenetic tree was constructed for representative sequences of predominant AOB and NOB enrichments using MEGA 7.
The sequences were also processed using a MOTHUR pipeline (v. 1.41.1) to filter reads for quality, create contigs and reduce noise63. Ambiguous sequences were discarded, and sequences longer than 466 base pairs (bp) were dislodged while those with a maximum homopolymer length of 6 bp were allowed. Sequences were aligned with SILVA database release 132, and the SILVA taxonomy was used for classification of representative sequences and operational taxonomic units (OTUs) at 97% similarity. Chimeras were identified and removed with the chimera.vsearch option. Alpha diversity (e.g., Chao, Shannon, Simpson, Fisher and ACE) for individual samples were estimated using MOTHUR and submitted to MicrobiomeAnalyst64. Samples analysed with MicrobiomeAnalyst were filtered for low abundance based on the mean abundance of OTUs, and for low variability using the inter-quantile range assessment.
Functional pridiction
Metagenomic inference and functional analysis were performed using a phylogenetic investigation of communities by reconstruction of unobserved states (PICRUSt). Pathway analysis was conducted using the Kyoto Encyclopaedia of Genes and Genomes (KEGG) orthologous pathways. The 16S rDNA data were analysed as indicated by the PICRUSt genome prediction software (http://picrust.github.io/picrust/)65 from raw sequence reads in the following environment: NumPy (1.7.1), biom-format (1.3.1), PICRUSt (1.0.0-dev), and PICRUSt script (1.0.0-dev).
Functional predictions were assigned up to KO tier for metabolic pathways related to the nitrogen cycle, which are assumed to be more relevant for this study. Heat map clustering for genes involved in nitrogen cycle was done using heatmapper.ca66. Functional inference of microbiome was analysed from sequencing data through MOTHUR using the SILVA database, through Piphillin, a function prediction tool56. It uses the nearest-neighbour matching using global alignment between the 16S rRNA gene sequencing input OTUs representative sequences and abundance table with updated genomic databases to infer the metagenomics content of the samples. We used the web version of Piphillin incorporating the KEGG October 2018 version as a reference database, and applying 97% identity cut-off. The software uses USEARCH to generate genome abundance table which is then normalised by the copy number to get the genome content and is summed to get the KO relative abundance table. This output was analysed using MicrobiomeAnalyst tool for metabolic network visualisation and pathway mapping. The relative abundance of functional categories as Kyoto Encyclopaedia of Genes and Genomes (KEGG)67 pathways were generated from OTU table of assigned taxa and were integrated and interpreted in KEGG pathway for nitrogen metabolism and visualized using VANTED (v.2.2.1)68.
Data availability
The raw sequencing data of AOB (SRX5815865) and NOB (SRX5815867) consortia are available under the project ID PRJNA542210 in NCBI database.
References
FAO, IFAD, UNICEF, WFP and WHO. The State of Food Security and Nutrition in the World 2018. Building climate resilience for food security and nutrition. Rome, FAO. (2018).
Baldisserotto, B. Fish physiology applied to aquaculture.2. ed.Santa Maria: UFSM Publishing Housen (2009).
Lazzari, R. & Baldisserotto, B. Nitrogen and phosphorus waste in fish farming. Bull. Inst. Fish. 34(no. 4), 591–600 (2008).
Tomasso, J. R. Toxicity of nitrogenous Wastes to Aquaculture Animals. Rev. Fish. Sci. 2(no. 4), 291–314 (1994).
Hartenstein. Nitrogen Metabolism in Non-Insect Arthropods, In Comparative Biochemistry of Nitrogen Metabolism”, vol. 1, Campbell, J. W. ed., Academic Press, London (1970).
Romano, N. & Zeng, C. Toxic Effects of Ammonia, Nitrite, and Nitrate to Decapod Crustaceans: A Review on Factors Influencing their Toxicity, Physiological Consequences, and Coping Mechanisms. Rev. Fish. Sci. 21(1), 1–21 (2013).
Tomasso, J. R. Environmental nitrite and aquaculture: a perspective. Aquaculture Int. 20, 1107–1116 (2012).
Hou, J., Song, C., Cao, X. & Zhou, Y. Shifts between ammonia-oxidising bacteria and archaea in relation to nitrification potential across trophic gradients in two large Chinese lakes (Lake Taihu and Lake Chaohu). Water Res. 47, 2285–2296 (2013).
Wang, B. Z. et al. Differential contributions of ammonia oxidisers and nitrite oxidisers to nitrification in four paddy soils. ISME J. 9, 1062–1075 (2015).
Sedlacek, C. J. et al. Effects of bacterial community members on the proteome of the ammonia-oxidising bacterium Nitrosomonas sp. strain Is79. Appl. Environ. Microbiol. 82, 4776–4788 (2016).
Dionisi, H. M. et al. Quantification of Nitrosomonas oligotropha-like ammonia-oxidizing bacteria and Nitrospira spp. from full-scale wastewater treatment plants by competitive PCR. Appl. Environ. Microbiol. 68, 245–253 (2002).
Liu, F., Li, J., Feng, G. & Li, Z. New Genomic Insights into “Entotheonella” Symbionts in Theonellaswinhoei: Mixotrophy, Anaerobic Adaptation, Resilience, and Interaction. Front. Microbiol. 7, 1333 (2016).
Li, Y. T. et al. Aerobic-heterotrophic nitrogen removal through nitrate reduction and ammonium assimilation by marine bacterium Vibrio sp. Y1–5. Bioresour. Technol. 230, 103–111 (2017).
Rout, P. R., Bhunia, P. & Dash, R. R. Simultaneous removal of nitrogen and phosphorous from domestic wastewater using Bacilluscereus GS-5 strain exhibiting heterotrophic nitrification, aerobic denitrification and denitrifying phosphorous removal. Bioresour. Technol. 244, 484–495 (2017).
Joo, H. S., Hirai, M. & Shoda, M. Piggery wastewater treating using Alcaligenesfaecalis strain No. 4 with heterotrophic nitrification and aerobic denitrification. Water Res. 40, 3029–3036 (2006).
Reyes-Santacruz, A. R. & Chen, Y. H. The potential of Yucca schidigera extract to reduce the ammonia pollution from shrimp farming. Bioresour. Technol. 113, 311–314 (2012).
Zhou, L. & Boyd, C. E. Total ammonia nitrogen removal from aqueous solutions by the natural zeolite, mordenite: A laboratory test and experimental study. Aquaculture. 432, 252–257 (2014).
Yilin, L. & Boyd, C. E. Laboratory tests of bacterial amendments for accelerating oxidation ratesof ammonia, nitrite and organic matter in aquaculture pond water. Aquaculture. 460, 45–58 (2016).
Park, H. D. & Noguera, D. R. Characterization of two ammonia-oxidizing bacteria isolated from reactors operated with low dissolved oxygen concentrations. J. Appl. microbiology. 102(5), 1401–17 (2007).
Bock, E., Koops, H. P., Möller, U. C. & Rudert, M. A new facultatively nitrite oxidizing bacterium, Nitrobacter vulgaris sp. nov. Arch. microbiology 153(2), 105–10 (1990).
Chaudhary, D. S., Vigneswaran, S., Ngom, H. H., Shim, W. G. & Moon, H. Bio-filter in water and wastewater treatment. Korean J. ChemEng 20(6), 1054–1065 (2003).
French, E., Kozlowski, J. A., Mukherjee, M., Bullerjahn, G. & Bollmann, A. Ecophysiological characterization of ammonia-oxidizing archaea and bacteria from freshwater. Appl. Environ. Microbiol. 78, 5773–5780 (2012).
Burrell, P. C., PhalennCM. & Hovanec, T. A. Identification of bacteria responsible for ammonia oxidation in freshwater aquaria. Appl. Environ. Microbiol. 67, 5791–5800 (2001).
Gao, J., Fan, X., Wu, G., Li, T. & Pan, K. Changes of abundance and diversity of ammonia-oxidizing archaea (AOA) and bacteria (AOB) in three nitrifying bioreactors with different ammonia concentrations. Desalination Water Treat. 57(no. 45), 21463–21475 (2016).
Di, H. J. et al. Ammonia-oxidizing bacteria and archaea grow under contrasting soil nitrogen conditions. FEMS Microbiol. Ecol. 72, 386–394 (2010).
Schleper, C. Ammonia oxidation: different niches for bacteria and archaea? ISME J. 4, 1092–1094 (2010).
Martens-Habbena, W. & Stahl, D. A. Nitrogen metabolism and kinetics of ammonia-oxidizing archaea. Methods Enzymol. 496, 465–487 (2011).
Verhamme, D. T., Prosser, J. I. & Nicol, G. W. Ammonia concentration determines differential growth of ammonia-oxidizing archaea and bacteria in soil microcosms. ISME J. 5, 1067–1071 (2011).
Könneke, M. et al. Isolation of an autotrophic ammonia-oxidizing marine archaeon. Nature 437, 543–546 (2005).
Alves, R. J. E., Minh, B. Q., Urich, T., von Haeseler, A. & Schleper, C. Unifying the global phylogeny and environmental distribution of ammonia-oxidising archaea based on amoA genes. Nat. Commun. 9(1), 1517–1533 (2018).
Cardona, E. et al. Bacterial community characterization of water and intestine of the shrimp Litopenaeusstylirostris in a biofloc system. J. BMC Microbiol. 16, 1–9 (2016).
Daims, H. et al. Complete nitrification by Nitrospira bacteria. Nature 528(7583), 504–9, https://doi.org/10.1038/nature16461 (2015).
Salcher, M. M., Posch, T. & Pernthaler, J. In situ substrate preferences of abundant bacterioplankton populations in a pre-alpine freshwater lake. ISME Journal. 7(5), 896–907 (2013).
Zeng, Y., Kasalický, V., Šimek, K. & Koblížek, M. Genome sequences of two freshwater beta-proteobacterial isolates, Limnohabitans species strains Rim28 and Rim47, indicate their capabilities as both photoautotrophs and ammonia oxidizers. J. Bacteriol. 194(22), 6302 (2012).
Fitzgerald, C. M., Camejo, P., Oshlag, J. Z. & Noguera, D. R. Ammonia-oxidizing microbial communities in reactors with efficient nitrification at low-dissolved oxygen. Water Res. 70, 38–51 (2015).
Kouki, S. et al. Isolation and characterization of facultative mixotrophic ammonia-oxidizing bacteria from constructed wetlands. J. Env. Sci. 23, 1699–1708 (2011).
Castelle, C. J., Brown, C. T., Thomas, B. C., Williams, K. H. & Banfield, J. F. Unusual respiratory capacity and nitrogen metabolism in a Parcubacterium (OD1) of the Candidate Phyla Radiation. Sci. Rep. 7, 40101 (2017).
Xia, X. Q. et al. Evaluating oligonucleotide properties for DNA microarray probe design. Nucleic Acids Res. 38, e121 (2010).
Regan, J. M., Harrington, G. W. & Noguera, D. R. Ammonia‐ and nitrite‐oxidizing bacterial communities in a pilot‐scale chloraminated drinking water distribution system. Appl. Env. Microbiol. 68, 73–81 (2002).
Holmes, A. J. et al. Phylogenetic structure of unusual aquatic microbial formations in Nullarbor caves, Australia. Env. Microbiol. 3, 256–264 (2001).
Rittmann, B. E. et al. Nitrification as a source of soluble organic substrate in biological treatment. Water Sci. Technol. 30, 1–8 (1994).
Dolinsek, J. et al. Interactions of nitrifying bacteria and heterotrophs: identification of a Micavibrio-like putative predator of Nitrospira spp. Appl. Environ. Microbiol. 79, 1534–1544 (2013).
Luo X. S., Han S., Lai S. S., Huang Q. Y. & Chen W. L. Long-term straw returning affects Nitrospira-like nitrite oxidizing bacterial community in a rapeseed-rice rotation soil. 57 309–315 (2017).
He, Z. et al. A novel denitrifying methanotroph of the NC10 phylum and its microcolony. Sci. Rep. 6, 32241 (2016).
Ettwig, K. F. et al. Nitrite-driven anaerobic methane oxidation by oxygenic bacteria. Nature. 464, 543–548 (2010).
Huang, X., Li, W., Zhang, D. & Qin, W. Ammonium removal by a novel oligotrophic Acinetobacter sp. Y16 capable of heterotrophic nitrification-aerobic denitrification at low temperature. Bioresour. Technol. 146, 44–50 (2013).
Li, C. et al. Removal of nitrogen by heterotrophic nitrification–aerobic denitrification of a phosphate accumulating bacterium Pseudomonas stutzeri YG-24. Bioresour. Technol. 182, 18–25 (2015).
Papen, H. & Von Berg, R. A. Most probable number method (MPN) for the estimation of cell numbers of heterotrophic nitrifying bacteria in soil. Plant. Soil. 199, 123–130 (1998).
Albertsen, M. et al. Genome sequences of rare, uncultured bacteria obtained by differential coverage binning of multiple metagenomes. Nat. Biotechnol. 31, 533–538 (2013).
Lu, H., Chandran, K. & Stensel, D. Microbial ecology of denitrification in biological wastewater treatment. Water Res. 64, 237–254 (2014).
Feng, C., Huang, L., Yu, H., Yi, X. & Wei, C. Simultaneous phenol removal, nitrification and denitrification using microbial fuel cell technology. Water Res. 76, 160–170 (2015).
Mohammadi, S. S., Pol, A., van Alen, T., Jetten, M. S. M. & Op den Camp, H. J. M. Ammonia oxidation and nitrite reduction in the Verrucomicrobial methanotroph Methylacidiphilum fumariolicum SolV. Front. Microbiol. 8, 1901 (2017).
Wilson, M. C. et al. An environmental bacterial taxon with a large and distinct metabolic repertoire. Nature 506, 58–62 (2014).
Momper, L., Aronson, H. S. & Amend, J. P. Genomic Description of ‘CandidatusAbyssubacteria,’ a Novel Subsurface Lineage Within the Candidate Phylum Hydrogenedentes. Front. Microbiol. 9, 1993 (2018).
Cerqueira, T. C., Barroso, H., Froufe, C. & Egas, R. Bettencourt. Metagenomic signatures of microbial communities in deep-sea hydrothermal sediments of Azores vent fields. Microb. Ecol. 76, 1–17 (2018).
Iwai, S. et al. Piphillin: Improved Prediction of Metagenomic Content by Direct Inference from Human Microbiomes. Plos one, 11(11) (2016).
Kindaichi, T., Ito, T. & Okabe, S. Ecophysiological interaction between nitrifying bacteria and heterotrophic bacteria in autotrophic nitrifying biofilms as determined by microautoradiography-fluorescence in situ hybridization. Appl. Environ. microbiology 70(3), 1641–1650 (2004).
Koops, H. P., Harms, H. & Wehrmann, H. Isolation of a moderate halophilic ammonia-oxidizing bacterium, Nitrosococcus mobiles nov.sp. Arch. Microbiol. 107, 277–282 (1976).
Watson, S. E. & Waterbury, J. B. Characteristics of two marine nitrite oxidizing bacteria, Nitrospinagracilisnov.gen. nov.sp. and Nitrococcusmobilisnov.gen. nov. sp. Arch. Microbiol. 77, 203–230 (1971).
Yuen, S. H. & Pollard, A. G. The determination of nitrogen in agricultural materials by the Nessler’s reagent: Preparation of the reagent. J. Sci. Food Agriculture 3, 441–447 (1952).
Griess-Romijn., van E. Physiological and chemical tests for drinking water. In NEN 1056. NederlandsNormalisatieInstitut Rijswijk (1966).
Ghosh, A. et al. Culture independent molecular analysis of bacterial communities in the mangrove sediment of Sundarbans India. Saline Syst. 6, 1 (2010).
Schloss, P. D. et al. Introducing Mothur: open-source. platform-independent, community-supported software for describing and comparing microbial communities. Appl. Environ. Microbiol. 75, 7537–7541 (2009).
Dhariwal, A. et al. MicrobiomeAnalyst - a web-based tool for comprehensive statistical, visual and meta-analysis of microbiome data. Nucleic Acids Res. 45, W180–188 (2017).
Langille, M. G. et al. Predictive functional profiling of microbial communities using 16S rRNA marker gene sequences. Nat. Biotechnol. 31(9), 814–821 (2013).
Babicki, S. et al. Heatmapper: web-enabled heat mapping for all. Nucleic Acids Res. (epub ahead of print) (2016).
Kanehisa, F. M., Tanabe, M., Sato, Y. & Morishima, K. KEGG: new perspectives on genomes, pathways, diseases and drugs. Nucleic Acids Res. 45, D353–D361 (2017).
Junker, B. H., Klukas, C. & Schreiber, F. VANTED: A system for advanced data analysis and visualization in the context of biological networks. BMC. Bioinformat. 7, 109 (2006).
Acknowledgements
This work was supported by ICAR-Consortium Research Platform on Vaccines and Diagnostics funded by the Ministry of Agriculture and Farmers Welfare, Government of India.
Author information
Authors and Affiliations
Contributions
P.K.P., S.V.A. and V.K.K. planning of the study and fund mobility. D.N. and M.L.A. culture enrichments and maintenance. V.T.N., S.D.G. and S.N., metagenomic analysis for the present study. V.B. and S.A. did metagenome sequencing, wrote the manuscript with input from V.T.N., S.D.G. and S.N. All authors have read and approved the final manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Baskaran, V., Patil, P.K., Antony, M.L. et al. Microbial community profiling of ammonia and nitrite oxidizing bacterial enrichments from brackishwater ecosystems for mitigating nitrogen species. Sci Rep 10, 5201 (2020). https://doi.org/10.1038/s41598-020-62183-9
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-020-62183-9
This article is cited by
-
Strategies for ammonia recovery from wastewater: a review
Environmental Chemistry Letters (2024)
-
Genetic variation in Zea mays influences microbial nitrification and denitrification in conventional agroecosystems
Plant and Soil (2024)
-
Microbial Communities in Standing Dead Trees in Ghost Forests are Largely Aerobic, Saprophytic, and Methanotrophic
Current Microbiology (2024)
-
Comparative analyses of gut microbiota reveal ammonia detoxification and nitrogen assimilation in Cyprinus carpio var. specularis
Folia Microbiologica (2024)
-
Physiological adaptation and population dynamics of a nitrifying sludge exposed to ampicillin
International Microbiology (2023)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
