
Abstract
The formation and subsequent breakup of the supercontinent Pangaea has dominated Earth’s evolution for the last 320 million years. Although its configuration at the time of breakup is widely accepted, there remains uncertainty about its configuration at the time of its amalgamation. The classic Pangaea-A model, widely known as “Wegenerian” configuration, implies that Pangaea did not deform internally between amalgamation and breakup. Palaeomagnetic studies suggest the possibility of a Pangaea-B configuration, in which Gondwana was located about 3000 km farther east relative to Laurasia compared its location in Pangaea-A. Here, we provide firm evidence of an Iberian-Appalachian connection in the Late Pennsylvanian (307–299 Ma) which confirms a Pangaea-A configuration for the relative locations of Gondwana and Laurasia in the late Palaeozoic, negating the possibility of Pangaea-B at that time. This evidence is based on palaeobotanical and biostratigraphic findings recently documented in the Carboniferous successions of Iberia (Douro Basin, Portugal). These new findings also precisely constrain the timing of uplift of the Appalachian and Iberian (Variscan) orogens and climatic changes during the amalgamation of Pangaea and final closure of the Rheic Ocean.
Similar content being viewed by others


Introduction
Over the past 30 years, a broad consensus has emerged that repeated cycles of supercontinent amalgamation and dispersal have occurred since the end of the Archean, and these cycles have profoundly affected the Earth’s evolution1,2. Less clear is whether the supercontinent changes its configuration during its existence due to internal stresses.
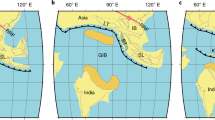
Although the classical “Wegenerian” configuration of Pangaea immediately prior to its Early Mesozoic breakup is well constrained, there remains uncertainty about its late Palaeozoic configuration. Two end member models have emerged; Pangaea-A, which is essentially the “Wegenerian” fit (A-13; A-24), and Pangaea-B, based on palaeomagnetic data5, in which Gondwana was located about 3000 km farther east relative to Laurasia, compared to the Pangaea-A configuration (Fig. 1). A late Palaeozoic Pangaea-B configuration (Fig. 2) would require substantial lateral (dextral) shear along major faults, inferred by Irving5 to have occurred between the middle Carboniferous and Late Triassic, in order to obtain the Wegenerian configuration before Pangaea breakup. More recent palaeomagnetic data have been used to support the transition from a Pangaea-B to a Pangaea-A configuration during the Permian6,7, and in the most recent model8, the transition occurred between 275 and 260 Ma. However, geologic evidence that would distinguish between these hypotheses is lacking. Moreover, the validity of the palaeomagnetic data purported to support the Pangaea-B configuration has recently been challenged9.

Idealized Pangaea-A (“Wegenerian”) configuration based on continental connection between eastern Laurentia (Laurasia) and Iberia (northwestern Gondwana20,21,22,23,24,25,26,27,28,29,30,31,32,33) in the late Palaeozoic (adapted from9). Colour legend for the image: blue: Oceans; light brown: Gondwana; dark brown: Laurasia; grey: shallow seas and coastal/flooded areas.

Late Palaeozoic Pangaea-B configuration in which Gondwana is located about 3000 km farther east relative to Laurasia (adapted from9).
The collision between Laurasia and Gondwana during the Late Devonian-early Permian was a key event in the amalgamation of Pangaea and resulted in the destruction of the Rheic Ocean and the formation of the Appalachian and Variscan (Hercynian) orogens in the interior of Pangaea10,11. A key element in reconstructing palaeogeographic environments is to examine the first appearance of shared flora between continents. For example, the occurrence of the Permian flora Glossopteris has been crucial in understanding the configuration of Gondwana12,13. The confinement of this flora to Gondwana and its absence from Laurasia has been attributed to the presence of physical barriers (e.g. distance, mountain ranges, climate/latitude) that may have restricted its migration13.
However, determination of the palaeogeography of Laurasia relative to Gondwana during the late Palaeozoic is hindered by the lack of palaeobiogeographic evidence linking both continents. In this paper, we draw on recent discoveries in Carboniferous successions in the Iberian Massif (Douro Basin, Portugal)14,15 that, for the first time, provide linkages between the ancient landmasses Laurentia and Iberia (located along the northern margin of Gondwana) along the palaeoequatorial belt during the Late Pennsylvanian (307–299 Ma) (Fig. 1). In so doing, we provide palaeobotanical and biostratigraphic evidence that the Pangaea-A configuration was in place at that time, negating the possibility of Pangaea-B configuration in the late Palaeozoic.
Iberian-Gondwanan Connection
Models for Variscan orogenesis and Pangaea amalgamation rely on ca. 420–320 Ma continental reconstructions. At ca. 420 Ma, reconstructions primarily influenced by palaeomagnetic data16,17 show Gondwanan terranes, including Iberia rifted from the northern Gondwanan margin thereby forming the Palaeotethys Ocean18,19. Other reconstructions, however, based on a wealth of faunal, lithological, stratigraphic, detrital zircon and palaeoclimatic data20,21,22,23,24,25, imply that these terranes remained along the Gondwanan margin for the entirety of the Palaeozoic26,27,28,29. In the latter scenario, Rheic Ocean closure resulted from continental collision of Laurasia with the northern Gondwanan margin, which began ca. 380 Ma25,30. Iberia preserves a continuous Early Ordovician to Late Devonian passive margin sequence31,32 including typically Gondwanan Late Ordovician glaciomarine deposits33, and lacks ca. 420 Ma rift-drift deposits predicted by the formation of the Palaeotethys Ocean. On the basis of this evidence, we adopt the second scenario and our reconstructions showing a unified Iberia and Gondwana throughout the Palaeozoic.
Palaeofloral and Biostratigraphic Evidence
Abundant Carboniferous-Permian floras and palaeoenvironmental/climatic distribution data have been identified in Laurasia13,15,34,35,36. Detailed studies15,34,37 of flora that demonstrate significant affinities between the Pennsylvanian (late Moscovian and Gzhelian) floras of North America and Iberian Massif are interpreted to reflect a proximal palaeobiogeography between Laurentia and Iberia within the palaeoequatorial belt. Biostratigraphic studies34,38 identify the existence of a macrofloral biostratigraphic gap for the Kasimovian stage in the Appalachian region in West Virginia Basin (USA) correlated with the Upper Pennsylvanian of Portugal15 (Fig. 3). This gap is documented in parts of the palaeoequatorial belt during the Kasimovian and is attributed to a lowstand reflecting a major glaciation event in southern Gondwana15,34.

Biostratigraphic constraints between Laurentia and Iberia including a macrofloral biostratigraphic gap correlated between the Upper Pennsylvanian successions of Appalachian region in West Virginia and Iberia in Portugal. Diagram modified from15. Abbreviations: R.: regional; Moscov.: Moscovian; West.: Westphalian.
Carboniferous-Permian floras, restricted to same type of palaeoenvironments shared by Laurentia and Iberia, are key elements to determine the palaeogeography of Pangaea as it amalgamated. Determination of land bridges linking Laurentia and Iberia for floral exchange attests to the importance of constraining the palaeoenvironmental and palaeoclimatic conditions between these continental lands in the interior of Pangaea. Such constraints are provided by the floras that were restricted to “dryland” environments located in the tropical regions of central Pangaea and lived in both Laurentia and Iberia. Cycadopsid Lesleya, a rare Carboniferous-early Permian seed-plant of the Euramerican realm, was a dry-climate adapted flora (known as “dryland flora”) restricted to tropical dryland environments of central Pangaea14,39,40,41,42,43 (Fig. 4).

Palaeoenvironmental and palaeoclimatic constraints and floral migration between Laurentia and Iberia within Pangaea-A. Enlarged view of central Pangaea in Fig. 1 (white rectangular box area) showing the emergence of “dryland” environments at varying spatial and temporal scales and diachronous migration of dry-climate adapted flora like Lesleya between the Laurentian and Iberian landmasses. Lesleya-fossil record data for the floral migration route are from14,40,41,42,43,51,52.
Pangaean tropical regions experienced major cyclic environmental changes during the Pennsylvanian-early Permian interval, with significant modifications to ecosystems and biotic communities (biotic stress) resulting from alternation of wetland and dryland floras. Such changes were a result of glacial and interglacial cycles, and their effects were especially felt in the tropical regions of central Pangaea during this interval35,40,41,43,44,45,46,47,48,49. The dryland environments occupied part of the tropical landscapes of central Pangaea during the Pennsylvanian14,35,40,41,42,43 (Fig. 4). The emergence of these environments is intricately linked to a warmer or drier climate during interglacial periods46,49,50. These interglacial periods led to significant changes in climate and therefore the overall composition of resident floral assemblages in the tropical regions of central Pangaea in the late Palaeozoic34,35,40,41,43,47,49.
Fossils of Lesleya have been widely documented in Early-Middle Pennsylvanian-age dryland basins of North America14,40,41,42,43,51,52. Recent discoveries in the Upper Pennsylvanian of Portugal have documented the first occurrence of Lesleya in Iberian Massif14. The Portuguese Lesleya specimens were found in lower Gzhelian strata of the Douro Basin and occur in intramontane deposits that preserve evidence of dry climate14. Dry climate is characterized by the moisture-deficient (dryness) and well-drained conditions14,15. The appearance of Lesleya in Iberia (Fig. 4) coincided with the onset of an interglacial interval in the Kasimovian-Gzhelian (304 Ma) after the waning of a major glaciation in southern Gondwana35,40,53,54. As a result, parts of palaeoequatorial belt especially of central Pangaea, where eastern Laurentia and Iberia were located, became drier and less humid during the Gzhelian (Late Pennsylvanian, 304–299 Ma)34,35,55 (Fig. 4).
Other typical dryland floras such as the walchian conifers Walchia and Ernestiodendron, cordaitalean Cordaites, callipterid peltasperms Autunia conferta and Rhachiphyllum, and the dicranophyllalean Dicranophyllum also flourished at various places in Laurentia (e.g. West Virginia) and Iberia. Such dryland biomes were more abundant during periods of warm or dry climate in the Late Pennsylvanian and early Permian14,15,34,37,38,40,49,50,56. These palaeobotanical data provide palaeogeographic constraints on the proximity of Laurentia and Iberia and are key to distinguishing between the competing Pangaea configurations.
The ‘Missing link’: Resolving Pangaea-A versus Pangaea-B Controversy
The Pangaea-A versus Pangaea-B controversy underscores large uncertainties about the palaeogeographic position of Gondwana relative to Laurasia in the Late Devonian-early Permian interval. Recent palaeobotanical and biostratigraphic studies14,15 indicate a proximal Iberian-Appalachian palaeogeography in the Late Pennsylvanian. Such evidence provides significant constraints in the palaeogeography, palaeoclimate and palaeotopography in both the Appalachian and Iberian (Variscan) orogens (Fig. 5).

Palaeogeographic and palaeotopographic constraints within Pangaea-A showing the continental linkage between eastern Laurentia and Iberia and uplift of the Appalachian and Variscan orogens in the late Gzhelian-early Permian. Topography adapted from North American 300 MA © 2013 Colorado Plateau Geosystems Inc (https://deeptimemaps.com/). Data for the legend are from57,58,59,60,61. Abbreviations: WV: West Virginia; IM: Iberian Massif; Aq: Aquitaine; AM: Armorican Massif; MC: French Central Massif; RH: Rheno-Hercynian terrane; ST: Saxo-Thuringian terrane; BM: Bohemian Massif; Sd: Sardinia (Italian island); Co: Corsica (French Mediterranean island); NI: Variscan basement of northern Italy.
Because they are indicators for climatic and environmental conditions14,39,40,41,42,43, the occurrence of dryland floras typical from North America such as Lesleya in the Upper Pennsylvanian strata of Portugal is evidence of migration of dry-climate adapted floras between the Laurasian and Gondwanan continents. This floral migration suggests that eastern Laurentia and Iberia were connected or geographically very close, sharing the same tropical dryland environment within central Pangaea in the Late Pennsylvanian (Fig. 4). Moreover, the appearance of Lesleya in the early Gzhelian (Late Pennsylvanian, 304–301 Ma) of Iberia, immediately after a transition from glacial to interglacial conditions in the Kasimovian-Gzhelian interval (304 Ma)40,53,54, indicates that this flora migrated from Laurentia to Iberia, possibly when new dryland habitats appeared (Fig. 4). In this proximal configuration, Iberia probably acted as a migratory option or refuge to the many dry-climate adapted floras of Laurentia, perhaps because conditions of greater dryness had prevailed in Iberia in the early Gzhelian14. During that time interval, new dryland species such as Lesleya iberiensis emerged in the Iberia in well-drained, moisture-deficient environments14.
The migration routes of dryland flora between Laurentia and Iberia (Fig. 4) provide insights into the location and timing of uplift of the Appalachian and Variscan orogens during continental collision between Laurasia and Gondwana during the amalgamation of Pangaea (Figs. 1 and 5). These migration routes were influenced by climate and tectonically-induced topographic changes. As mountain ranges acted as physical barriers to the floral exchanges13 between Laurentia and Iberia within central Pangaea, this migration occurred before uplift of the Appalachian and Variscan orogens, i.e. during the early Gzhelian (Late Pennsylvanian, 304–301 Ma) (Fig. 4). This palaeobiogeographic connection records early stages of uplift during the assembly and amalgamation of Pangaea and implies a connection along the palaeoequatorial belt between the Appalachian orogen and the Variscan orogen in Iberia (Fig. 5). A macrofloral biostratigraphic gap correlated between the Upper Pennsylvanian successions of Appalachian region in West Virginia and Portugal15 (Fig. 3) supports an Iberian-Appalachian connection at that time. The timing of this connection implies that uplift of the Appalachian and Variscan orogens occurred during the late Gzhelian (Late Pennsylvanian) to Asselian (early Permian) (301–295 Ma).
Our data provide the ‘missing link’ between Gondwana and Laurasia during the final amalgamation of the supercontinent Pangaea in the Late Pennsylvanian and confirms a Pangaea-A (“Wegenerian”) configuration at that time (Fig. 1). Consequently, these results indicate that the palaeomagnetic data used to support a Pangaea-B configuration (Fig. 2) in the late Palaezoic5 represent an artifact of data quality, geometrical fits used to restore the Atlantic-bordering continents to one another, and processes such as inclination shallowing in clastic rocks, as suggested by Domeier et al.9.
References
Worsley, T. R., Nance, R. D. & Moody, J. B. Global tectonics and eustasy for the past 2 billion years. Marine Geology 58, 373–400 (1984).
Nance, R. D., Murphy, J. B. & Santosh, M. The supercontinent cycle: A retrospective essay. Gondwana Research 25, 4–29 (2014).
Hallam, A. Atlas of Paleobiogeography. (Elsevier, Amsterdam, 1973).
Van der Voo, R. Paleomagnetism, Continental drift, and Plate Tectonics. Reviews of Geophysics and Space Physics 17, 227–235 (1979).
Irving, E. Drift of the major continental blocks since the Devonian. Nature 270, 304–309 (1977).
Muttoni, G. et al. Early Permian Pangea ‘B’ to Late Permian Pangea ‘A’. Earth and Planetary Science Letters 215(3), 379–394 (2003).
Muttoni, G. et al. Opening of the Neo-Tethys Ocean and the Pangea B to Pangea A transformation during the Permian. GeoArabia 14(48), 17–48 (2009).
Muttoni, G. & Kent, D. V. Adria as promontory of Africa and its conceptual role in the Tethys Twist and Pangea B to Pangea A Transformation in the Permian. Rivista Italiana di Paleontologia e Stratigrafia 125, 249–269 (2019).
Domeier, M., Van der Voo, R. & Torsvik, T. H. Review Article: Paleomagnetism and Pangea. The Road to reconciliation. Tectonophysics 514-517, 14–43 (2012).
Murphy, J. B. & Nance, R. D. The Pangea conundrum. Geology 36, 703–706 (2008).
Gutiérrez-Alonso, G. et al. Self-subduction of the Pangean global plate. Nature (Geoscience) 1, 549–553 (2008).
Wegener, A. Die Entstehung der Kontinente. Geologische Rundschau 3, 276–292 (1912).
Vai, G. B. Development of the palaeogeography of Pangaea from Late Carboniferous to Early Permian. Palaeogeography, Palaeoclimatology, Palaeoecology 196, 125–155 (2003).
Correia, P., Sá, A. A., Murphy, J. B., Šimůnek, Z. & Flores, D. Lesleya Lesquereux from the Pennsylvanian of the Iberian Massif: part of a dryland megaflora from the Variscan orogen, northwestern Portugal. Canadian Journal of Earth Sciences 53, 883–895 (2016).
Correia, P., Šimůnek, Z., Sá, A. A. & Flores, D. A new Late Pennsylvanian floral assemblage from the Douro Basin, Portugal. Geological Journal 53, 2507–2531 (2018).
Van der Voo, R. Paleomagnetism of the Atlantic, Tethys and Iapetus Oceans. (Cambridge, New York, Port Chester, Melbourne, Sydney, 1993).
Tait, J., Schätz, M., Bachtadse, V. & Sojel, H. Palaeomagnetism and Palaeozoic palaeogeography of Gondwana and European terranes. In: Orogenic Processes: Quantification and Modelling in the Variscan Belt (eds. Franke, W., Haak, V., Oncken, O. & Tanner, D.). Geological Society of London Special Publication 179, 21–34 (2000).
Stampfli, G., Hochard, C., Vérard, C., Wilhem, C. & von Raumer, J. The formation of Pangea. Tectonophysics 593, 1–19 (2013).
Torsvik, T. H. & Cocks, L. R. M. Earth geography from 400 to 250 Ma: a palaeomagnetic, faunal and facies review. Journal of the Geological Society, London 161, 555–572 (2004).
Robardet, M. Alternative approach to the Variscan belt in southwestern Europe; proorogenic paleobiogeographical constraints. In: Variscan-Appalachian dynamics: The building of the Late Paleozoic basement (eds. Martínez-Catalán, J. R., Hatcher, R. D. Jr., Arenas, R. & Díaz García, F.). Geological Society of America Special Paper 364, 1–15 (2002).
Robardet, M. The Armorica “microplate”: fact or fiction? Critical review of the concept and contradictory palaeobiogeographical data. Palaeogeography, Palaeoclimatology, Palaeoecology 195, 125–148 (2003).
Linnemann, U. et al. West African provenance for Saxo-Thuringia (Bohemian Massif): did Armorica ever leave pre-Pangean Gondwana? – U/Pb- SHRIMP zircon evidence and the Nd isotopic record. International. Journal of Earth Sciences 93, 683–705 (2004).
Linnemann, U., Pereira, M. F., Jeffries, T., Drost, K. & Gerdes, A. Cadomian Orogeny and the opening of the Rheic Ocean: new insights in the diacrony of geotectonic processes constrained by LA–ICP–MS U–Pb zircon dating (Ossa-Morena and Saxo-Thuringian Zones, Iberian and Bohemian Massifs). Tectonophysics 461, 21–43 (2008).
Pastor-Galán, D., Gutiérrez-Alonso, G., Murphy, J. B., Fernández-Suárez, J. & Hoffmann, M. Provenance analysis of the Paleozoic sequences of the northern Gondwana margin in NW Iberia: Passive margin to Variscan collision and orocline development. Gondwana Research 23, 1089–1103 (2013).
Kroner, U. & Romer, R. L. Two plates—Many subduction zones: The Variscan orogeny reconsidered: Gondwana Research 24, 298–329 (2013).
Quesada, C. Geological constraints on the Paleozoic tectonic evolution of tectonostratigraphic terranes in the Iberian Massif. Tectonophysics 185, 225–245 (1991).
Quesada, C., Fonseca, P., Munhá, J., Oliveira, J. T. & Ribeiro, A. The Beja-Acebuches Ophiolite (Southern Iberia Variscan foldbelt): geological characterization and geodynamic significance. Boletín Geológico Minero 105, 3–49 (1994).
Winchester, J. A., Pharaoh, T. C. & Verniers, J. Paleozoic Amalgamation of Central Europe: an introduction and synthesis of new results from recent geological and geophysical investigations. In: Paleozoic Amalgamation of Central Europe (eds. Winchester, J. A., Pharaoh, T. C. & Verniers, J.). Geological Society of London, Special Publication 201, 1–18 (2002).
Scotese, C. R. A Continental Drift Flipbook. Journal of Geology 112, 729–741 (2004).
Arenas, R. et al. Two-stage collision: Exploring the birth of Pangea in the Variscan terranes. Gondwana Research 25, 756–763.
Robardet, M. & Gutiérrez-Marco, J. C. The Ordovician, Silurian and Devonian sedimentary rocks of the Ossa Morena Zone (SW Iberian Peninsula, Spain). Journal of Iberian Geology 30, 73–92 (2004).
Quesada, C. The Ossa Morena Zone of the Iberian Massif: a tectonostratigraphic approach to its evolution. Zeitschrift der Deutschen Gesellschaft für Geowissenschaften 157(4), 585–595 (2006).
Robardet, M. & Doré, F. The late Ordovician diamictic formations from Southwestern Europe: North-Gondwana glaciomarine deposits. Paleogeography, Paleoclimatology, Paleoecology 66, 19–31 (1988).
Wagner, R. H. & Lyons, P. C. A critical analysis of the higher Pennsylvanian megafloras of the Appalachian region. Review of Palaeobotany and Palynology 95, 255–283 (1997).
Wagner, R. H. Climatic changes as mirrored by Carboniferous and Permian floral distributions. Monografías del Jardín Botánico de Córdoba 39, 29–39 (2004).
Cocks, L. R. M. & Torsvik, T. H. The Palaeozoic geography of Laurentia and western Laurussia: A stable craton with mobile margins. Earth-Science Reviews 106, 1–51 (2011).
Wagner, R. H. & Álvarez-Vázquez, C. The Carboniferous floras of the Iberian Peninsula: A synthesis with geological connotations. Review of Palaeobotany and Palynology 162, 239–324 (2010).
Wagner, R. H. & Álvarez-Vázquez, C. A redescription of the Stephanian species Callipteridium virginianum (Fontaine & White, 1880) comb. nov. and Alethopteris leonensis Wagner, 1964. Callipteridium and Alethopteris. Scripta Geologica (Special Issue) 7, 93–139 (2010).
Falcon-Lang, H. J., Pendleton, J. L. & Wellman, C. H. Dryland plant communities in the Pennsylvanian (mid- to late Bolsovian) Winterbourne Formation of Bristol, southern Britain: further evidence for taphonomic megabias. Review of Palaeobotany and Palynology 166, 268–285 (2011).
DiMichele, W. A., Cecil, C. B., Montañez, I. P. & Falcon-Lang, H. J. Cyclic changes in Pennsylvanian paleoclimate and effects on floristic dynamics in tropical Pangaea. International Journal of Coal Geology 83, 329–344 (2010).
DiMichele, W. A., Bashforth, A. R., Eble, C. F. & Nelson, W. J. A Middle Pennsylvanian (early Asturian) tropical dry forest, Atokan-Desmoinesian boundary, Illinois Basin, USA Spanish. Journal of Palaeontology 31, 41–84 (2016).
Bashforth, A. R., Cleal, C. J., Gibling, M. R., Falcon-Lang, H. J. & Miller, R. F. Early Pennsylvanian plant communities on a seasonal tropical fluvial landscape (Tynemouth Creek Formation, New Brunswick, Canada). Review of Palaeobotany and Palynology 200, 229–263 (2014).
Bashforth, A. R., DiMichele, W. A., Eble, C. F. & Nelson, W. J. Dryland vegetation from the Middle Pennsylvanian of Indiana (Illinois Basin): the dryland biome in glacioeustatic, paleobiogeographic, and paleoecologic context. Journal of Paleontology 90, 785–814 (2016).
Gastaldo, R. A., DiMichele, W. A. & Pfefferkorn, H. W. Out of the icehouse into the greenhouse – A late Paleozoic analog for modern global vegetational change. GSA Today 6, 1–7 (1996).
Falcon-Lang, H. J. Pennsylvanian tropical rainforests responded to glacial-interglacial rhythms. Geology 32, 689–692 (2004).
DiMichele, W. A. Wetland-dryland vegetational dynamics in the Pennsylvanian Ice Age tropics. International Journal of Plant Sciences 175, 123–164 (2014).
Cecil, C. B., DiMichele, W. A. & Elrick, S. D. Middle and Late Pennsylvanian cyclothems, American Midcontinent: Ice-age environmental changes and terrestrial biotic dynamics. Comptes Rendus Geoscience 346, 159–168 (2014).
DiMichele, W. A., Pfefferkorn, H. W. & Gastaldo, R. A. Response of Late Carboniferous and Early Permian plant communities to climate change. Annual Review of Earth and Planetary Sciences 29, 461–487 (2001).
DiMichele, W. A. et al. Geology of the Pennsylvanian-Permian in the Dunkard basin (ed. Harper, J. A.). Guidebook, 76th Annual Field Conference of Pennsylvania Geologists, 60–102 (Washington, PA, 2011).
DiMichele, W. A. et al. Callipterid peltasperms of the Dunkard Group, Central Appalachian Basin. International Journal of Coal Geology 119, 56–78 (2013).
Leary, R. L. & Pfefferkorn, H. W. An Early Pennsylvanian Flora with Megalopteris and Noeggerathiales from West-Central Illinois. (Illinois State Geological Survey Circular 500, 1977).
DiMichele, W. A., Kerp, H., Tabor, N. J. & Looy, C. V. The so-called “Paleophytic-Mesophytic” transition in equatorial Pangea – Multiple biomes and vegetational tracking of climate change through geological time. Palaeogeography, Palaeoclimatology, Palaeoecology 268, 152–163 (2008).
Isbell, J. L., Lenaker, P. A., Askin, R. A., Miller, M. F. & Babcock, L. E. Reevaluation of the timing and extent of late Paleozoic glaciation in Gondwana: role of the transantarctic mountains. Geology 31, 977–980 (2003).
Fielding, C. R. et al. Stratigraphic imprint of the Late Palaeozoic Ice Age in eastern Australia: a record of alternating glacial and nonglacial climate regime. Journal of the Geological Society 165, 129–140 (2008).
Cecil, C. B. An overview and interpretation of autocyclic and allocyclic processes and the accumulation of strata during the Pennsylvanian–Permian transition in the central Appalachian Basin, USA. International Journal of Coal Geology 119, 21–31 (2013).
Martino, R. L. Walchian conifers from the Mid-Late Pennsylvanian Conemaugh Group in the Appalachian Basin: Stratigraphic and depositional context, and paleoclimatic significance. International Journal of Coal Geology 171, 153–168 (2017).
von Raumer, J., Stampfli, G. M. & Bussy, F. Gondwana-derived microcontinents—the constituents of the Variscan and Alpine collisional orogens. Tectonophysics 365, 7–22 (2003).
Simancas, J. F. et al. The tectonic frame of the Variscan–Alleghanian orogen in Southern Europe and Northern Africa. Tectonophysics 398, 181–198 (2005).
Arenas, R. et al. The Galicia–Ossa-Morena Zone: Proposal for a new zone of the Iberian Massif. Variscan implications. Tectonophysics 681, 135–143 (2016).
Romer, R. L. & Kroner, U. First direct evidence for a contiguous Gondwana shelf to the south of the Rheic Ocean. Geology 47, 767–770 (2019).
van Staal, C. R., Whalen, J. B., Valverde-Vaquero, P., Zagorevski, A. & Rogers, N. Ancient orogens and modern analogues (eds. Murphy, J. B. et al.). Geological Society of London Special Publication 327, 271–316 (London, 2009).
Acknowledgements
We thank Trond Torsvik and an anonymous reviewer for their constructive comments. We are also grateful to scientific illustrator Nathan Rogers for the assistance on Figures drawing. JBM acknowledges the continuing support of N.S.E.R.C., Canada.
Author information
Authors and Affiliations
Contributions
P.C. and J.B.M. wrote the main manuscript text.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Correia, P., Murphy, J.B. Iberian-Appalachian connection is the missing link between Gondwana and Laurasia that confirms a Wegenerian Pangaea configuration. Sci Rep 10, 2498 (2020). https://doi.org/10.1038/s41598-020-59461-x
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-020-59461-x
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
