
Abstract
Invasive species continue to proliferate and detrimentally impact ecosystems on a global scale. Whilst impacts are well-documented for many invaders, we lack tools to predict biotic resistance and invasion success. Biotic resistance from communities may be a particularly important determinant of the success of invaders. The present study develops traditional ecological concepts to better understand and quantify biotic resistance. We quantified predation towards the highly invasive Asian tiger mosquito Aedes albopictus and a representative native mosquito Culex pipiens by three native and widespread cyclopoid copepods, using functional response and prey switching experiments. All copepods demonstrated higher magnitude type II functional responses towards the invasive prey over the analogous native prey, aligned with higher attack and maximum feeding rates. All predators exhibited significant, frequency-independent prey preferences for the invader. With these results, we developed a novel metric for biotic resistance which integrates predator numerical response proxies, revealing differential biotic resistance potential among predators. Our results are consistent with field patterns of biotic resistance and invasion success, illustrating the predictive capacity of our methods. We thus propose the further development of traditional ecological concepts, such as functional responses, numerical responses and prey switching, in the evaluation of biotic resistance and invasion success.
Similar content being viewed by others

Introduction
Invasive alien species continue to threaten biodiversity globally and disrupt ecosystem structure and functioning1,2. Identifying and understanding ecological processes that contribute to invasion success are essential to predicting and remediating invader impacts in communities3,4. In particular, species diversity in recipient communities can drive ‘biotic resistance’ towards invasive species as a result of competition, predation, parasitism and other antagonisms, thus potentially limiting or preventing invasion success5,6. Predation is an especially pervasive force which can shape ecosystem structuring through both trait- and density-mediated interactions with prey7,8. However, the relationship between the invasibility of communities and levels of biotic resistance (e.g. predation) therein remain unclear, and the development and validation of methodologies to predict invasion success or failure are urgently required4. Indeed, the identification of universal species traits which reliably predict invader impact and success across taxonomic and trophic groups has thus far been largely unsuccessful9. In turn, this has acted as a hindrance to practical assessments of invasion risk and the associated development of measures to remediate invader impacts, and particularly for invasive species with no known invasion history3.
Invasion science has been slow to integrate some key ecological concepts which are classically predictive in consumer-resource systems3. Traditionally, the functional response, defined as the relationship between resource availability and resource use, has been applied by ecologists to quantify interaction strengths between consumers and resources (e.g. predators and prey)10,11,12. Indeed, both functional response form and magnitude have been identified as robust measures of ecological impact from consumers towards resources, including from existing and emerging invasive species towards native resources4,9,13,14. In particular, the attack rate (search coefficient) and handling time parameters of functional response models align closely with the magnitude of ecological impacts across resource densities9. Furthermore, whilst three broad forms of functional response have generally been categorised (linear type I, saturating type II, sigmoidal type III), type II functional responses have been identified as particularly impactful towards target resources due to the destabilising pressures they impart on low densities of resources such as prey species13. In contrast, type III functional responses may enable population stability of resources due to low density refuge effects15.
Classically, the consumer functional response has also been combined with the numerical response, which describes the consumer population response as resource densities change. In turn, this can quantify the overall impact of consumers on resource populations, termed the total response or offtake rate10,11,12. Recently, the functional response approach in invasion science has been combined with proxies for the numerical response (e.g. abundance, fecundity) to develop population-level metrics which have been proven to correlate tightly with known ecological impacts (i.e. Relative Impact Potential3; Relative Invasion Risk16). However, whilst functional and numerical responses have been applied to examine the ecological impacts of invasive consumers (e.g. predators) towards native resources (e.g. prey), there has been relatively little application of these approaches to quantify biotic resistance potential towards invasive resources (e.g. prey) in the context of invasion success6,17,18.
Aside from functional and numerical responses, integrations of switching propensities between invasive and native resources have been neglected by invasion scientists4,19,20,21. This is despite the importance of consumer switching patterns for the persistence of species. Characteristically, if a consumer exhibits a switching propensity, disproportionately fewer rare resources are consumed whilst, simultaneously, disproportionately more abundant resources are consumed15. Importantly, switching (i.e. a form of frequency-dependent predation) may facilitate coexistence between resources (e.g. prey) by imparting stability through low-density refugia concomitant with type III functional responses22. Therefore, in the context of invasion science, switching propensities by resident consumers have the capacity to directly influence biotic resistance towards invasive species and could thus be used in combination with functional and numerical responses of consumers in predictive approaches for invasion success. Indeed, if a resident consumer exhibits high magnitude per capita (functional response) and population-level (numerical response) effects, coupled with consumptive preferences towards invasive resources, offtake rates towards invaders will be high and it is thus theoretically less likely that the invader will succeed and subsequently have impacts on ecosystems. Although mechanistic interpretation must be cautioned in the absence of field experiments, comparative laboratory experiments may be used to rapidly quantify these effects (i.e. functional/numerical responses and preferences), and such studies have proven highly informative in the context of invader impact and success14,20.
The present study develops a novel approach to quantify and compare levels of biotic resistance by resident consumers towards invasive species using the aforementioned concepts. We have three key objectives: (1) to quantify per capita effects among natural enemies towards separate invasive and native prey; (2) to examine ratio-dependent prey preferences of the natural enemies where invasive and native prey coexist, and; (3) to use population-level responses alongside per capita effects and prey preferences to quantify and compare levels of biotic resistance among agents. These results help to inform management responses for the biocontrol of target species by providing a novel means of comparing agent efficacies. Further, our results help to inform predictions of invasion success, given that high levels of biotic resistance towards invasive over native species may limit invasive species success likelihoods5,6.
We use a well-documented field pattern, whereby an invasive mosquito species is a superior competitor compared to natives within the same trophic level, but is known to coexist, perhaps through differential biotic resistance from indigenous predators23,24,25,26. We develop our approach using a model invader/native system based on the invasive Asian tiger mosquito Aedes albopictus and native common house mosquito Culex pipiens. Aedes albopictus is a highly invasive species that can vector pathogens that cause disease, and is known to outcompete native mosquitoes27,28,29. Yet, this invader has been documented to coexist with native species in aquatic habitats despite its superior competitive ability24. Whilst C. pipiens is also regarded as an invasive vector species in certain parts of the world30, in our study system (United Kingdom) it represents a widespread and abundant native species31, and is thus an appropriate candidate for invader-native comparative purposes. We quantify and compare biotic resistance towards these mosquito prey by three species of cyclopoid copepods. Predatory copepods are abundant and widespread crustaceans which are capable of thriving in most aquatic habitat types32. Given their tolerance to ephemeral environments via dormant life stages, and potential for both human-mediated and zoochorous dispersal between aquatic habitats33,34,35,36, there is high potential for copepod-mosquito overlap in various aquatic habitat types and thus predatory interactions. Furthermore, copepods are known to regulate mosquito populations in aquatic systems36. Informed by empirical field-patterns, we hypothesise that: (1) copepod functional response magnitudes will be significantly greater towards invasive compared to native mosquito prey; (2) predators will display a ratio-independent preference for invasive mosquito prey over native mosquitoes; (3) biotic resistance from native natural enemies will differ among predator species according to their per capita, selectivity and population-level effects.
Results
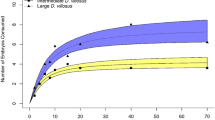
In all experiments, 100% of control prey survived and thus experimental deaths were directly attributable to predation by copepods, which was also evidenced by partially consumed prey remaining post-experiment. In the functional response experiment, support for raw consumption models containing prey species, predator species and prey density received substantial support (Table S1a). Significantly greater numbers of invasive A. albopictus were consumed as compared to native C. pipiens overall (χ2 = 9.14, df = 1, p = 0.003; Fig. 1). Consumption differed significantly across predator species (χ2 = 16.11, df = 2, p < 0.001), owing to significantly greater consumption by M. fuscus than M. albidus (p < 0.001) and M. viridis (p = 0.02). However, consumption towards A. albopictus was higher for all predators given a statistically unclear ‘prey species × predator species’ interaction (χ2 = 4.19, df = 2, p = 0.12). Consumption was also significantly greater under increasing prey densities (χ2 = 73.66, df = 4, p < 0.001).

Functional responses of Macrocyclops albidus (a), Macrocyclops fuscus (b) and Megacyclops viridis (c) towards Aedes albopictus (red triangles) and Culex pipiens (blue circles) prey. Shaded areas are bootstrapped (n = 2000) 95% confidence intervals and points are underlying consumption data.
All predatory copepods exhibited type II functional responses towards both C. pipiens and A. albopictus, owing to significantly negative first order linear coefficients (Fig. 1; Table 1). Attack rates tended to be higher towards A. albopictus than C. pipiens prey by M. albidus and M. viridis, whilst handling times were generally shorter for A. albopictus across all predator species (Table 1; Fig. 1). Maximum feeding rates were thus higher towards invasive A. albopictus prey than native C. pipiens for all three predators (Fig. 1; Table 1). Accordingly, in all instances, overall per capita predatory impacts as quantified by the functional response ratio (FRR) were considerably higher towards the invasive A. albopictus over the native C. pipiens (Table 1). Differential per capita predatory impacts between prey were, however, particularly pronounced for M. viridis (Table 1), and further evidenced by a divergence in confidence intervals under higher prey densities (Fig. 1c), as compared to confidence interval convergence in M. albidus (Fig. 1a) and M. fuscus treatments (Fig. 1b).
Raw consumption in the prey switching experiments was also substantially influenced by prey species, predator species and proportional prey availability (Table S1b). Again, significantly greater numbers of invasive A. albopictus were consumed than native C. pipiens (χ2 = 17.14, df = 1, p < 0.001; Fig. 2). Consumption was significantly influenced by predator species (χ2 = 7.48, df = 2, p = 0.02), owing to significantly greater consumption by M. fuscus as compared to M. albidus (p = 0.04), and was greater where a particular prey species was present in higher proportions (χ2 = 70.08, df = 4, p < 0.001).

Prey switching propensities of Macrocyclops albidus (a), Macrocyclops fuscus (b) and Megacyclops viridis (c) towards different proportions of Aedes albopictus (red triangles) and Culex pipiens (blue circles) prey. The solid line indicates the expected value in the case of no preference between prey types. Means are ± 1 SE.
None of the focal copepod predators exhibited a prey switching propensity between invasive A. albopictus and native C. pipiens (Table 2; Fig. 2). Instead, preferential selection towards the invader was exhibited across all proportional availabilities, as evidenced by recurrently high preference indices (ai > 0.5; Table 2). Preference index type (predicted/observed) and proportional availability were identified as important model components (Table S1c). Preference indices towards A. albopictus were significantly higher than expected under conditions of no preference (χ2 = 16.33, df = 1, p < 0.001). However, the strength of preference towards A. albopictus interacted with its proportional availability (χ2 = 17.10, df = 4, p = 0.002). Here, overall, preferences towards A. albopictus were particularly stronger than expected under higher proportional availabilities (0.5, 0.75, 0.9) (all p ≤ 0.01).
Owing to greater relative FRR between invasive/native prey species, higher fecundity and strong preferences towards invasive A. albopictus prey, Relative Biotic Resistance (RBR) was greater by M. albidus as compared to M. fuscus (Table 3). However, in turn, RBR scores of both M. albidus and M. fuscus were lower than M. viridis. This was driven by particularly marked relative FRR of M. viridis towards A. albopictus prey, coupled with higher levels of fecundity in this copepod (Table 3; Fig. 3). Accordingly, M. viridis is expected to exert the greatest degree of biotic resistance towards the invasive prey.

Triplot illustrating differential Biotic Resistance (BR) of Macrocyclops albidus (circle), Macrocyclops fuscus (triangle) and Megacyclops viridis (diamond) towards invasive Aedes albopictus prey. Estimations include relative functional response ratios (FRRs) between invasive and native prey, predator fecundities and invasive prey preferences. Increasing levels of BR are read from bottom left to top right, and raw BR scores are displayed in parentheses. Colours are ramped with increasing BR towards red colouration.
Discussion
Invasive alien species continue to spread, establish and reproduce in novel environments1, yet we have a distinct lack of methodologies to predict invasion success3,4. Biotic resistance may be a key mechanism which controls the success of invasive species. In the present study, we develop novel measures of biotic resistance driven by resident consumers towards invasive prey through the integration of functional responses, numerical response proxies and prey switching propensities. Our model species Aedes albopictus is a highly invasive vector mosquito, known to have superior competitive abilities for resources over analogous native mosquitoes, however often fails to displace these natives24,27,29,37. Culex pipiens is a native and widespread mosquito in our study region (United Kingdom), whilst A. albopictus has only recently been detected in the United Kingdom, following numerous successful invasions across Europe38. However, our results suggest that differential biotic resistance may limit the invasion success of this species, and cyclopoid copepods are known to be important mosquito predators35,36. Indeed, given that all native copepods exhibited greater interaction strengths and strong preferences towards the invasive A. albopictus over the native C. pipiens, resident predator communities may limit levels of invader success and impact. Copepods are particularly well-adapted to thrive in ephemeral aquatic habitats which these mosquitoes colonise, owing to their ability to enter dormant life history stages and spread via zoochorous dispersal by vectors such as birds and insects, or by wind33,34,39,40. Whilst human-mediated copepod introductions into minute container-style habitats have been required for effective mosquito control at community-scales35, even in these instances, our results provide evidence for the efficacy of copepods in reducing target invader populations whilst alleviating native species from predatory impact. Nevertheless, mechanistic interpretation of our laboratory experimental results should be cautioned in the context of invasion success, with further field-based validation required that incorporates additional context-dependencies, such as emergent effects from interactions with other predator types and habitat complexities.
In the present study, irrespective of copepod species, we consistently demonstrate higher magnitude per capita ecological impacts towards invasive A. albopictus prey as compared to analogous native C. pipiens prey using a comparative functional response approach. We subsequently demonstrate clear consumptive preferences towards invasive over native mosquito prey by all focal consumers regardless of proportional prey species availability, conducive with a lack of prey switching. Then, we integrate fecundity estimations as a proxy for the numerical response to quantify and compare the potential biotic resistance of resident consumers towards this invasive prey species. Whilst drawing parallels between laboratory-based studies and field observations should viewed with caution, our results align with field patterns of coexistence, wherein invasive Aedes mosquitoes have repeatedly been shown to coexist with native competitors, despite their clear competitive advantage23,24,25. We propose that the combination of functional and numerical responses, alongside examinations of prey switching propensities, may help to predict the occurrence of such field patterns in relation to biotic resistance and invasion success. Although we applied our metrics to a model copepod-mosquito predator-prey system, our approaches are equally applicable to other consumer-resource systems where an invasive species suffers from biotic resistance by resident consumers (e.g. predators). Thus, our metrics can, at least theoretically, be applied across multiple habitat types and taxonomic groups for predictions of invasion success and quantifications of biotic resistance. Yet, given numerous additional context-dependencies are known to alter levels of biotic resistance (e.g. habitat complexity6,18), these effects should be considered in future studies to better-reflect real systems.
Macrocyclops albidus, M. fuscus and M. viridis exhibited type II functional responses towards both prey types, characterised by high rates of mosquito prey consumption at low densities. This finding aligns with copepod functional responses forms reported in other studies18. However, functional response attack rates tended to be considerably higher, and handling times lower, towards invasive A. albopictus as compared to native C. pipiens. Given attack rates correspond to impacts at low prey densities whilst handling times reciprocate asymptotic maximum feeding rates, per capita predatory impacts towards A. albopictus remain higher than analogous native prey irrespective of prey density. The particularly high per capita impact of copepods towards A. albopictus also aligns with the documented ability of copepod biocontrol agents to be especially efficacious in the suppression of Aedes mosquitoes as compared to Culex36. Furthermore, our holistic derivations of per capita impact through coupling of attack rate and handling time into the functional response ratio (FRR: a/h) demonstrate greater predatory impacts by all focal copepods towards the invasive mosquito prey as compared to the native. This differential impact was particularly pronounced for M. viridis. The FRR metric has recently been developed for invasion scientists and practitioners and balances information from both key functional response parameters41. Invasive consumers have been shown to exhibit consistently higher FRRs as compared to native comparators across multiple study systems and taxonomic groups41. In a similar vein, we suggest that the FRR metric can be applied in quantifications of biotic resistance across study systems, as it can negate contradictory impact predictions for natural enemies based on one functional response parameter over the other (i.e. attack rate, handling time).
None of the copepod predators examined in the present study exhibited a prey switching propensity away from the invasive prey. That is, relative to proportional abundances, invasive A. albopictus were disproportionately selected over native C. pipiens across all availabilities. Prey switching facilitates patterns of coexistence in ecosystems through a form of frequency-dependent predation characterised by low density refuge effects4,15. If our laboratory-based results persist in the wild, it is likely that the sustained preferential selection towards A. albopictus would permit patterns of coexistence between these prey species even in light of the superior competitive capability of A. albopictus over analogous native mosquito species24,27,28,29,37,42,43. As such, this consumptive preference may offset competitive replacement of the native by the invader. Our results exemplify the potential power of our metrics for quantifications of biotic resistance which may mediate levels of invasion success, and future work should ground-truth these concepts across other invasive species study systems using field-based observations.
Behavioural responses to predator cues are likely key drivers of such differential biotic resistance between invasive and native prey. In particular, naïveté to unfamiliar predators in novel ecosystems can influence interaction strengths and further impede invasion success20,44. Invasive Aedes mosquitoes have been shown to be less responsive to predation risk and exhibit higher incidences of behaviours which make them more apparent and vulnerable to predators (e.g. thrashing, browsing)24,25. Indeed, whilst Culex mosquitoes are filter feeders which spend most time at the water surface, Aedes are browsers which spend more time thrashing below the surface45. Given these substantial behavioural differences, it is plausible that the predatory patterns exhibited by copepods in the present study extend to other aquatic predator groups, owing to potentially higher encounter rates with the more motile Aedes prey. Furthermore, Aedes mosquitoes have been shown to be attracted to predatory copepods when ovipositing, whilst Culex mosquitoes are evasive of these cues18,46. Such behavioural factors could be major drivers in limiting the success of invasive species via biotic resistance and, in our system, may help to regulate disease risk in the context of invasive vector mosquito species.
The present study integrated estimates of fecundity as proxies for numerical responses of copepods47. The use of such proxies has proven robust in derivations of ecological impacts of invasive species and biocontrol agents3,48, and, here, high fecundity could facilitate rapid population-level responses to increases in resource availability following natural enemy inoculation. However, importantly, the present study did not compare fecundities of predators fed on the focal invasive/native prey, which may have altered estimates given differences in nutritional values between prey species. Nonetheless, whilst M. fuscus exhibited high magnitude functional responses towards invasive prey and strong selective tendencies, the fecundity of this copepod is substantially lower than both M. albidus and M. viridis47. Accordingly, in this study, our novel metric identified M. viridis as a particularly efficacious predator towards invasive A. albopictus prey, owing to high per capita impacts towards the invader, strong selectivity traits and relatively marked fecundity. For management, our predictive metrics suggest augmentative releases of native copepods such as M. viridis for biocontrol could be especially efficacious in the suppression of invasive mosquito species, in light of favourable consumptive and reproductive traits.
In conclusion, we propose that the integration of traditional ecological concepts that have been neglected by invasion scientists could enhance predictions of biotic resistance and help to inform invasion success. In turn, such predictions of biotic resistance directly inform management strategies for pests, vectors and invasive species via biocontrol. Biotic resistance from predators can be a key mechanism which controls invasion success, and these predator-prey interactions can be robustly quantified in controlled laboratory conditions. We show that the assimilation of functional responses, numerical response proxies and prey switching propensities enables more holistic derivations of potential biotic resistance towards invasive species at the population-level. Our results are consistent with empirical patterns, whereby the invasive mosquito A. albopictus is capable of outcompeting native mosquito species in a laboratory setting27,37, but has not been able to displace native mosquitoes in a similar fashion in the field24,49. Yet, further field-based studies are required to validate the predation patterns documented in the present study, and link them to invasion success. Nevertheless, we propose that biotic resistance is an important factor in regulating the invasion process, and can be quantified using metrics grounded in classical ecological concepts. For practitioners, use of these concepts could enable relatively rapid comparisons of biological control agents prior to release, by quantifying and comparing per capita agent effects and preferences alongside population-level responses. In turn, this could improve the efficiencies associated with natural enemy introductions. Future research should also seek to ascertain the context-dependency of these approaches in predicting the success or failure of invasions across a multitude of study systems, alongside implications for the efficacy of biocontrol agents. Moreover, quantifications of predatory efficacies across a full spectrum of life history stages would provide a more holistic account of biotic resistance levels.
Materials and Methods
Animal collection and husbandry
Eggs of Aedes albopictus were obtained through the infraVec2 project and originated from Montpellier, France. Eggs of Culex pipiens complex mosquitoes were obtained from a colony maintained at Queen’s Marine Laboratory (QML), Portaferry, Northern Ireland, originating from The Pirbright Institute, Surrey, England48. Both mosquito species were maintained in a laboratory at QML, at 25 ± 2 °C and under a 16:8 light and dark photoperiod. The focal predators, Macrocyclops albidus, Macrocyclops fuscus and Megacyclops viridis were obtained from Glastry Clay Pit Ponds, Northern Ireland (54°29′18.5″N, 5°28′19.9″W) and cultured in the same laboratory (25 ± 2 °C,16:8 light and dark) on a diet of Paramecium caudatum and Chilomonas paramecium ad libitum until maturation.
Experimental design
Adult female predatory M. albidus, M. fuscus and M. viridis (respective mean total lengths excluding caudal setae ± SD: 1.70 ± 0.09 mm; 1.81 ± 0.11 mm; 1.83 ± 0.17 mm) were selected for experiments and separately starved for 24 h prior to feeding. Recently hatched, size-matched first instar A. albopictus (mean ± SD: 1.40 ± 0.12 mm) and C. pipiens (mean ± SD: 1.32 ± 0.11 mm) larvae were used as prey. Copepods are known to be most efficient in consumption of early instar mosquito prey36. Experiments were undertaken in 20 mL arenas of 42 mm dia. containing dechlorinated tapwater from an aerated source during daylight. We employed a phenomenological experimental approach to compare biotic resistance towards mosquitoes factorially in a replicated laboratory design. Accordingly, our design does not seek to mechanistically replicate natural systems (see9). Indeed, mechanistic interpretation of such experiments must be approached with caution, or supported with further empirical parameter estimates50,51,52. Nevertheless, phenomenological designs, such as ours, are useful for comparative purposes in factorial experiments to examine differences in predator-prey interactions under controlled conditions53.
For the functional response experiment, prey species were introduced separately at each of five densities into arenas (2, 4, 7, 10 or 15; n = 5 per experimental group). For the prey switching experiment, prey species were introduced in combination at each of five ratios (2:18, 5:15, 10:10, 15:5 or 18:2; n = 3 per experimental group). Experiments were conducted in a completely randomised array to eliminate positional effects. After addition, prey were allowed to settle for 2 h prior to the beginning of the experiments via predator introduction. Once individual copepod predators were introduced, they were allowed to feed for 6 h, after which the predators were removed and remaining live larval mosquito prey counted and identified to quantify numbers killed. Controls in each experiment consisted of a replicate of each prey treatment in the absence of predators.
Statistical analyses
Data were analysed using R v 3.5.154. In the functional response experiment, raw numbers of prey consumed were examined with respect to prey species, predator species and starting prey densities in a factorial generalised linear model (GLM). All interaction terms were included in the initial model. A Poisson error distribution with log link was employed. We used second-order derivations of Akaike’s Information Criterion (AICc) and model averaging to identify the best-supported model using the ‘MuMIn’ package55,56. Here, all possible models were identified and ranked based on AICc (lower values indicate a better fit). Model comparisons used ∆AICc, comprising the difference between the AICc of candidate models and the best-supported model. Akaike model weights (wi) were additionally used to probabilistically identify the best model, wherein predictor variables with good support yielded high cumulative wi values (near 1). Post-hoc Tukey tests were performed using ‘lsmeans’ where a factor yielded significance at the 95% confidence interval57.
The ‘frair’ package was used to perform functional response analyses58. Logistic regression considering the proportion of prey consumed with respect to initial prey density was used to identify functional response types. Categorically, a type II functional response is inferred where a significantly negative first-order term results, whilst a significantly positive first order term followed by a significantly negative second-order term indicates a type III functional response59. To account for prey depletion over the experimental period, we fit Rogers’ random predator equation for the non-replacement of prey59,60:
where Ne is the number of prey eaten, N0 is the initial density of prey, a is the attack rate, h is the handling time and T is the total experimental period. The random predator equation was fit for each predator and prey treatment group using maximum likelihood estimation, with the Lambert W function implemented to make the equation solvable61. Functional response models were non-parametrically bootstrapped 2000 times to generate 95% confidence intervals around starting estimations. Using the handling time (h) parameter, maximum feeding rate estimates (1/h) were additionally calculated.
We subsequently applied a new overall measure of per capita impact towards both prey types for each predator, by combining attack rates (a, functional response initial slope) and handling times (h, functional response asymptote) into the functional response ratio, which captures both parameters41:
where FRR is the attack rate a divided by the handling time h. This solves the problem of which parameter to choose for comparisons, as a large a combined with a small h gives a large value (and hence quantifies a large per capita effect), while a low a and a high h gives a low value (and hence quantifies a low per capita effect). We denote FRRi as towards invasive A. albopictus and FRRn as towards native C. pipiens.
In the prey switching experiment, numbers of prey consumed were analysed using generalised linear mixed models (GLMM) with Poisson error distribution and log link using the ‘lme4’ package62. Here, consumption was modelled with prey species, predator species and proportion available, alongside their interactions, as fixed effects, and with a random effects structure to account for repeated measures of prey types within each experimental replicate. Model averaging based on AICc was, again, implemented to select predictors which minimised information loss55, and post-hoc comparisons were performed using pairwise Tukey tests57.
Manly’s selectivity index was then used to quantify preferences for invasive A. albopictus prey by each predator species across proportions available, with adjustments for non-replacement of prey63,64:
where ai is Manly’s selectivity index for invasive A. albopictus, ni0 is the number of the invader available at the start of the experiment, ri is the number of the invader consumed, m the number of prey types, nn0 the number of native C. pipiens available at the start of the experiment and rn is the number of native prey consumed. Resulting indices range from 0 to 1, wherein 0 indicates complete avoidance and 1 indicates complete preference. In our two-prey system, values of 0.5 are indicative of neutral selectivity by predators between prey types. Prior to formal analysis, we transformed resulting ai values to account for extreme data points (0, 1)65:
where αt is the transformation and n is the sample size. Beta regression using the ‘betareg’ package was used to compare indices towards A. albopictus with those expected under null preference (0.5) with respect to predator species and prey proportion available66. Model averaging based on AICc was used in model selection as before, and post-hoc comparisons were undertaken using Tukey tests55,57.
Combining the above results, we then quantified Biotic Resistance (BR) towards invasive prey using relative FRRs between invasive A. albopictus and native C. pipiens prey (FRRi/FRRn; Eq. 2), reproductive effort as a numerical response proxy (clutch weight produced per female body weight per day47) and mean invasive prey preferences (Eq. 3) for each predator species:
where the Biotic Resistance (BR) of a predator towards invasive prey is a product of the relative FRR between invasive and native prey (FRRi/FRRn), predator numerical response proxy reproductive effort (fecundity, FE) and the mean preference index towards the invasive prey (αi). We selected fecundity as a suitable numerical response proxy given its importance for the proliferation of natural enemies following changes in resource availability, and because reproductive effort estimates for the focal predator species were readily available in the literature. Relative Biotic Resistance (RBR) was then developed and used to compare among the three different predator species:
where BR1 and BR2 are Biotic Resistance for predator 1 and predator 2, respectively. Here, values of 1 indicate equivalence in biotic resistance between the two predators and values >1 indicate greater biotic resistance by predator 1 as compared to predator 2. Conversely, RBR values <1 indicate lesser biotic resistance by predator 1 compared to predator 2. We produced triplots to further illustrate differences16.
We thus first quantified and compared functional responses by three native predators towards native and invasive prey when presented separately. Second, we examined prey preferences of the same predator species towards the two prey species when both are present simultaneously at different relative proportions. Thirdly, we used a predator numerical response proxy (fecundity), alongside functional responses and prey preferences, to predict which resident predator is likely to exert the greatest degree of biotic resistance towards the focal invasive species.
Data availability
Underlying functional response and prey switching data are available in the online supporting information.
References
Seebens, H. et al. No saturation in the accumulation of alien species worldwide. Nat. Commun. 8, 14435 (2017).
Seebens, H. et al. Global rise in emerging alien species results from increased accessibility of new source pools. Proc. Nat. Acad. Sci. 115, E2264–E2273 (2018).
Dick, J. T. A. et al. Invader relative impact potential: a new metric to understand and predict the ecological impacts of existing, emerging and future invasive alien species. J. Appl. Ecol. 54, 1259–1267 (2017).
Cuthbert, R. N., Dickey, J. W. E., McMorrow, C., Laverty, C. & Dick, J. T. A. Resistance is futile: lack of predator switching and a preference for native prey predict the success of an invasive prey species. R. Soc. Open Sci. 5, 180339 (2018).
Elton, C. S. The ecology of invasions by animals and plants (Methuen, 1958).
MacNeil, C., Dick, J. T. A., Alexander, M. E., Dodd, J. A. & Ricciardi, A. Predators vs. alien: differential biotic resistance to an invasive species by two resident predators. NeoBiota 19, 1–19 (2011).
Paine, R. T. Food webs: linkage, interaction strength and community infrastructure. J. Anim. Ecol. 49, 667–685 (1980).
Abrams, P. A., Menge, B. A., Mittelbach, G. G., Spiller, D. A. & Yodzis, P. The role of indirect effects in food webs in Food webs: integration of patterns and dynamics (eds Polis, G. A. & Winemiller, K.) 371–395 (Chapman and Hall, 1996).
Dick, J. T. A. et al. Advancing impact prediction and hypothesis testing in invasion ecology using a comparative functional response approach. Biol. Invasions 16, 735–753 (2014).
Solomon, M. E. The natural control of animal populations. J. Anim. Ecol. 18, 1–35 (1949).
Holling, C. S. The components of predation as revealed by a study of small mammal predation of the European pine sawfly. Can. Entomol 91, 293–320 (1959).
Holling, C. S. Some characteristics of simple types of predation and parasitism. Can. Entomol. 91, 385–398 (1959).
Dick, J. T. A. et al. Ecological impacts of an invasive predator explained and predicted by comparative functional responses. Biol. Invasions 15, 837–846 (2013).
Alexander, M. E., Dick, J. T. A., Weyl, O. L. F., Robinson, T. B. & Richardson, D. M. Existing and emerging high impact invasive species are characterized by higher functional responses than natives. Biol. Lett. 10, 20130946 (2014).
Murdoch, W. W. Switching in general predators—experiments on predator specificity and stability of prey populations. Ecol. Monogr. 39, 335–354 (1969).
Dickey, J. W. E. et al. Assessing the relative potential ecological impacts and invasion risks of emerging invasive species. NeoBiota 40, 1–24 (2018).
Twardochleb, L. A., Novak, M. & Moore, J. W. Using the functional response of a consumer to predict biotic resistance to invasive prey. Ecol. Appl. 22, 1162–1171 (2012).
Cuthbert, R. N., Callaghan, A. & Dick, J. T. A. Interspecific variation, habitat complexity and ovipositional responses modulate the efficacy of cyclopoid copepods in disease vector control. Biol. Control 121, 89–87 (2018).
Taylor, N. G. & Dunn, A. M. Predatory impacts of alien decapod Crustacea are predicted by functional responses and explained by differences in metabolic rate. Biol. Invasions 20, 2821–2837 (2018).
Cuthbert, R. N. et al. Intermediate predator naïveté and sex-skewed vulnerability predict the impact of an invasive higher predator. Sci. Rep. 8, 14282 (2018).
Joyce, P. W. S., Dickey, J. W. E., Cuthbert, R. N., Dick, J. T. A. & Kregting, L. Using functional responses and prey switching to quantify invasion success of the Pacific oyster, Crassostrea gigas. Mar. Environ. Res. 145, 66–72 (2019).
Hughes, R. N. & Croy, M. I. An experimental analysis of frequency-dependent predation (switching) in the 15-spined stickleback, Spinachia spinachia. J. Anim. Ecol. 62, 341–352 (1993).
Griswold, M. W. & Lounibos, L. P. Does differential predation permit invasive and native mosquito larvae to coexist in Florida? Ecol. Entomol. 30, 122–127 (2005).
Kesavaraju, B., Damal, K. & Juliano, S. A. Do natural container habitats impede invader dominance? Predator-mediated coexistence of invasive and native container-dwelling mosquitoes. Oecologia 155, 631–639 (2008).
Zuharah, W. F. & Lester, P. J. Are exotic invaders less susceptible to native predators? A test using native and exotic mosquito species in New Zealand. Popul. Ecol. 53, 307–317 (2011).
Juliano, S. A., Westby, K. M. & Ower, G. D. Know your enemy: effects of a predator on native and invasive container mosquitoes. J. Med. Entomol. 56, 320–328 (2019).
Aliabadi, B. K. & Juliano, S. A. Escape from gregarine parasites affects the competitive impact of an invasive mosquito. Biol. Invasions. 4, 283–297 (2002).
Carrieri, M., Bacchi, M., Bellini, R. & Maini, S. On the competition occurring between Aedes albopictus and Culex pipiens (Diptera: Culicidae) in Italy. Ecol. Entomol. 32, 1313–1321.
Marini, G. et al. The effect of interspecific competition on the temporal dynamics of Aedes albopictus and Culex pipiens. Parasit. Vectors 10, 102 (2017).
Lounibos, P. Invasions by insect vectors of human disease. Annu. Rev. Entomol. 47, 233–266 (2002).
Townroe, S. & Callaghan, A. Morphological and fecundity traits of Culex mosquitoes caught in gravid traps in urban and rural Berkshire, UK. Bull. Entomol. Res. 105, 615–620 (2015).
Williamson, C. E. & Reid, J. W. Copepoda in Ecology and Classification of North American Freshwater Invertebrates (eds Thorp, J. H. & Covich, A. P.) 915–954 (Academic Press, 2001).
Havel, J. E. & Shurin, J. B. Mechanisms, effects, and scales of dispersal in freshwater zooplankton. Limnol. Oceanogr. 49, 1229–1238 (2004).
Green, A. J. & Figuerola, J. Recent advances in the study of long-distance dispersal of aquatic invertebrates via birds. Divers. Distrib. 11, 149–156 (2005).
Kay, B. H. & Nam, V. S. New strategy against Aedes aegypti in Vietnam. Lancet 365, 613–617 (2005).
Marten, G. G. & Reid, J. W. Cyclopoid copepods. J. Am. Mosq. Control Assoc. 23, 65–92 (2007).
Teng, H. J. & Apperson, C. S. Development and survival of immatures Aedes albopictus and Aedes triseriatus (Diptera: Culicidae) in the laboratory: effects of density, food, and competition on response to temperature. J. Med. Entomol. 37, 40–52 (2000).
Medlock, J. M. et al. Detection of the invasive mosquito species Aedes albopictus in southern England. Lancet Infect. Dis. 17, 140.
Figuerola, J. & Green, A. J. Dispersal of aquatic organisms by waterbirds: a review of past research and priorities for future studies. Freshwater Biol. 47, 483–494 (2002).
Hessen, D. O., Jensen, T. C. & Walsberg, B. Zooplankton diversity and dispersal by birds; insights from different geographical scales. Front. Ecol. Evol. 7, 74 (2019).
Cuthbert, R. N., Dickey, J. W. E., Coughlan, N. E., Joyce, P. W. S. & Dick, J. T. A. The Functional Response Ratio (FRR): advancing comparative metrics for predicting the ecological impacts of invasive alien species. Biol. Invasions 21, 2543–2547 (2019).
Costanzo, K. S., Mormann, K. & Juliano, S. A. Asymmetrical competition and patterns of abundance of Aedes albopictus and Culex pipiens (Diptera: Culicidae). J. Med. Entomol. 42, 559–570 (2005).
Murrell, E. G. & Juliano, S. A. Competitive abilities in experimental microcosms are accurately predicted by a demographic index for R*. PLoS One 7, e43458 (2012).
Li, Y., Ke, Z., Wang, S., Smith, G. R. & Liu, X. An exotic species is the favourite prey of a native enemy. PLoS ONE 6, e24299 (2011).
Yee, D. A., Kesavaraju, B. & Juliano, S. A. Larval feeding behavior of three co-occurring species of container mosquitoes. J. Vector Ecol. 29, 315–322 (2004).
Torres-Estrada, J. L., Rodríguez, M. H., Cruz-López, L. & Arredondo-Jimenez, J. I. Selective oviposition by Aedes aegypti (Diptera:Culicidae) in response to Mesocyclops longisetus (Copepoda:Cyclopoidea) under laboratory and field conditions. J. Med. Entomol. 38, 188–192 (2001).
Maier, G. Patterns of life history among cyclopoid copepods of central Europe. Freshwater Biol. 31, 77–86 (1994).
Cuthbert, R. N., Dick, J. T. A., Callaghan, A. & Dickey, J. W. E. Biological control agent selection under environmental change using functional responses, abundances and fecundities; the Relative Control Potential (RCP) metric. Biol. Control 121, 50–57 (2018).
Lounibos, L. P. et al. Testing predictions of displacement of native Aedes by the invasive Asian Tiger Mosquito Aedes albopictus in Florida, USA. Biol. Invasions 3, 151–166 (2001).
Caldow, R. W. G. & Furness, R. W. Does Holling’s disc equation explain the functional response of a kelptoparasite? J. Anim. Ecol. 70, 650–662 (2001).
Jeschke, J. M. & Hohberg, K. Predicting and testing functional responses: an example from a tardigrade–nematode system. Basic Appl. Ecol. 9, 145–151 (2008).
Jeschke, J. M., Kopp, M. & Tollrian, R. Predator functional responses: discriminating between handling and digesting prey. Ecol. Monogr. 72, 95–112 (2002).
Alexander, M. E., Dick, J. T. A. & Callaghan, A. Trait-mediated indirect interactions in a marine intertidal system as quantified by functional responses. Oikos 122, 1521–1531 (2012).
R Core Development Team. R: A Language and Environment for Statistical Computing (R Foundation for Statistical Computing, 2018).
Barton, K. MuMIn: Multi-model inference. (R package, 2015).
Burnham, K. P. & Anderson, D.R. Model selection and multi-model inference: A practical Information-theoretic approach (Springer, 2002).
Lenth, R. V. Least-Squares Means: The R Package lsmeans. J. Stat. Softw. 69, 1–33 (2016).
Pritchard, D. W., Paterson, R. A., Bovy, H. C. & Barrios-O’Neill, D. Frair: an R package for fitting and comparing consumer functional responses. Meth. Ecol. Evol. 8, 1528–1534 (2017).
Juliano, S. A. Nonlinear curve fitting: predation and functional response curves in Design and Analysis of Ecological Experiments (eds Scheiner, S. M. & Gurevitch, J.) 159–182 (Oxford University Press, 2001).
Rogers, D. Random search and insect population models. J. Anim. Ecol. 41, 369–383 (1972).
Bolker, B. M. Ecological models and data in R (Princeton University Press, 2008).
Bates, D., Maechler, M., Bolker, B. & Walker, S. Fitting linear mixed-effects models using lme4. J. Stat. Softw. 67, 1–48 (2015).
Manly, B. F. J. A model for certain types of selection experiments. Biometrics 30, 281–294 (1974).
Chesson, J. The estimation and analysis of preference and its relationship to foraging models. Ecology 64, 1297–1304 (1983).
Smithson, M. & Verkuilen, J. A. Better lemon squeezer? Maximum-likelihood regression with beta-distributed dependent variables. Psych. Methods 11, 54–71 (2006).
Cribari-Neto, F. & Zeileis, A. Beta regression in R. J. Stat. Softw. 34, 1–24 (2010).
Acknowledgements
We acknowledge funding from the Department for the Economy, Northern Ireland and the Natural Environment Research Council. This publication was supported by the project, Research Infrastructures for the control of vector-borne diseases (lnfravec2), which has received funding from the European Union’s Horizon 2020 research and innovation programme under grant agreement No. 731060. We also acknowledge The Pirbright Institute under BBSRC project code BBS/E/I/00007039. Thanks to Marie Russell for stimulating discussion.
Author information
Authors and Affiliations
Contributions
Conceived and designed the experiments: R.C., A.C., J.D. Performed the experiments: R.C. Analysed the data: R.C. Contributed materials and analysis tools: R.C., J.D., A.C. Wrote the first draft of paper: R.C. Prepared revised versions: R.C., A.C., J.D.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Cuthbert, R.N., Callaghan, A. & Dick, J.T.A. A novel metric reveals biotic resistance potential and informs predictions of invasion success. Sci Rep 9, 15314 (2019). https://doi.org/10.1038/s41598-019-51705-9
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-019-51705-9
This article is cited by
-
Biological, ecological and trophic features of invasive mosquitoes and other hematophagous arthropods: What makes them successful?
Biological Invasions (2024)
-
Assessment of the occurrence and abundance of an exotic zooplankton species (Kellicottia bostiniensis) across a neotropical wetland over 12 years
Limnology (2023)
-
Predicting invasive consumer impact via the comparative functional response approach: linking application to ecological theory
Biological Invasions (2022)
-
Pushing the switch: functional responses and prey switching by invasive lionfish may mediate their ecological impact
Biological Invasions (2021)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
