
Abstract
Recent work has disclosed the critical role played by enamel peptides in sex classification of old skeletal remains. In particular, protein AMELY (amelogenin isoform Y) is present in the enamel dental tissue of male individuals only, while AMELX (isoform X) can be found in both sexes. AMELY can be easily detected by LC-MS/MS in the ion extracted chromatograms of the SM(ox)IRPPY peptide (monoisotopic [M + 2 H]+2 mass = 440.2233 m/z). In this paper, we exploited the dimorphic features of the amelogenin protein to determine the sex of the so-called ‘Lovers of Modena’, two Late Antique individuals whose skeletons were intentionally buried hand-in-hand. Upon discovery, mass media had immediately assumed they were a male-female couple, even if bad preservation of the bones did not allow an effective sex classification. We were able to extract proteins from the dental enamel of both individuals (~1600 years old) and to confidently classify them as males. Results were compared to 14 modern and archaeological control samples, confirming the reliability of the ion chromatogram method for sex determination. Although we currently have no information on the actual relationship between the ‘Lovers of Modena’ (affective? Kin-based?), the discovery of two adult males intentionally buried hand-in-hand may have profound implications for our understanding of funerary practices in Late Antique Italy.
Similar content being viewed by others


Introduction
Sex, together with age-at-death and height, is one of the critical pieces of information needed to define the biological profile of skeletal remains. Although some bones are sexually dimorphic (e.g., cranium and os coxae)1,2, the bad state of preservation of some archaeological remains might alter or totally compromise the readability of dimorphic traits in an individual, even if the appropriate osteological techniques are applied. Similarly, taxonomy, context and/or age may also influence the correct determination of sex3,4,5. For example, it is usually straightforward to determine the sex of a buried adult human through the macroscopic examination of dimorphic districts, while it can be difficult to ascertain the sex of faunal remains or of pre-pubertal individuals4. In these contexts, DNA may be a valid alternative for sex determination even if the analytical costs and the survival of the DNA itself may strongly limit the use of genetic markers6.
Recently, a paper by Stewart et al.7 has revolutionized the way to achieve sex determination of skeletons from archaeological and forensic contexts, thanks to the enamel proteins. One of these proteins is amelogenin, whose gene is translated into two isoforms linked to sexual chromosomes8, namely AMELX (present in both sexes) and AMELY (restricted to the male sex only), which constitute 90% of the entire enamel proteome9. The homonymous genes are expressed as amelogenins, the proteins within tooth enamel10. These proteins are released by ameloblasts (i.e., enamel growth cells) during the secretory stage of enamel development9, helping the crystal structure organization.
By liquid-chromatography mass spectrometry (LC-MS/MS), a relatively rapid and cost-effective technique, it is possible to check the occurrence of AMELX and/or AMELY isoforms within enamel samples in order to rapidly estimate the sex of an individual. Specifically, the peptide SM(ox)IRPPY (monoisotopic [M + 2 H]+2 mass 440.2233 m/z), is present only in the AMELY isoform, unambiguously characterizing male sex. Conversely, the absence of the AMELY peptides can be due to female sex or to a partial lack of sequence coverage11. For this reason, the probability of a female true positive sex estimation is rarely near 100%.
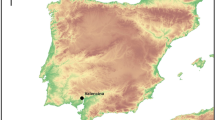
In this work, we examined the enamel proteome of two skeletons exhumed from a peculiar archaeological context, exploiting the sex-specificity of AMELY. In particular, we determined the sex of the so-called ‘Lovers of Modena’, two adult individuals intentionally buried hand-in-hand (Fig. 1; see Figs S17 and12). They were found in an Italian Late Antiquity cemetery (4th‒6th century, Ciro Menotti, Modena)12, together with other eleven individuals. Some of these skeletons showed signs of trauma, likely related to their violent death during war conflicts12. Immediately after the discovery of the ‘Lovers’ tomb, news about this peculiar finding spread all over the world and mass media rumored that the skeletons belonged to a man and a woman who had fallen in love13. However, due to the bad preservation of the skeletal dimorphic districts, sex determination based on the canonical osteological methods was not possible14. Moreover, preliminary genetic analyses were not consistent, due to diagenetic alterations of bone tissue, visible as a black-reddish coloration (see Fig. 1c)15, and low collagen content. Hence, we investigated the sex classification of the ‘Lovers’ using enamel peptides, as this tissue is more resistant to diagenetic modifications in both its organic16 and inorganic17,18 components (see also19). Our results were compared to control samples from the same site (n = 2) and from other Italian funerary contexts (n = 10). Two modern deciduous teeth were also used to further check the reliability of the method. To strengthen the method for sex estimation by ion chromatograms7, we propose the use of three-peptide peaks, with the aim of confidently identifying AMELY.

(a) Map of Italy with the location of the city of Modena (from NASA Visible Earth project ‒ credits to Jacques Descloitres, MODIS Rapid Response Team, NASA/GSFC); (b) Photograph of the Ciro Menotti necropolis (4th–6th century; Modena) during the archaeological excavation; the ‘Lovers of Modena’ are depicted within the inset; (c) Teeth and maxilla of individual 7_CM13; (d) Roman town of Mutina (Latin name of Modena) with the necropolis plan (modified from12).
Mass Spectrometry Results
In total, sixteen human teeth (Table 1) were analyzed during this study. Mascot database search and identification of the extracted peptides revealed that the most detectable species originated from the major tooth enamel proteins amelogenin, enamelin, and ameloblastin (Table S1). Three specimens (4_CM11; 5_FC176 and 17_MdT-2) showed the presence of CO1A2 (human collagen alpha-2(I) chain), likely related to residual dentine tissue. The high resolution (ion tolerance 5 ppm) extracted ion chromatograms for all the teeth revealed the presence of peptides derived from enamel proteins (see Figs S1–S16), as described in20: SIRPPYPSY (AMELX; [M + 2 H]+2 540.2796 m/z); SYEVLTPLK (AMELX,Y; [M + 2 H]+2 525.2975 m/z); PYFGYFGYH (ENAM; [M + 2 H]+2 575.7533 m/z); YEVLTPLKWY (AMELX,Y; [M + 2 H]+2 656.3528 m/z). A subset of the specimens also showed the presence of peptide SM(ox)IRPPY (AMELY; [M + 2 H]+2 440.2233 m/z), related to male sex7. We identified other peptides containing methionine and present only in AMELY, namely M(ox)IRPPY ([M + 2 H]+2 396.7073 m/z), SMIRPPY ([M + 2 H]+2 432.2258 m/z), SM(ox)RPPYS ([M + 2 H]+2 483.7393 m/z) and VLTPLKWYESM(ox)IRPPY ([M + 3 H]+3 670.3570 m/z) (see Fig. S18 and Table S2). For all the individuals but one (3_CM12-2), sex estimation based on the osteology agrees with the peptide fingerprint. Individual 3_CM12-2 was initially classified as a young woman through the ox coxae and the skull, but the presence of AMELY in its enamel likely suggests that the sex was misclassified. The two modern deciduous teeth were correctly classified as belonging to a male (16_MdT-1) and a female (17_MdT-2) through their enamel peptides (Table 1). Moreover, the presence of AMELY isoform in the enamel of the two ‘Lovers of Modena’ indicates that they were both males (Fig. 2). The occurrence of peptides SMIRPPY, M(ox)IRPPY and SM(ox)IRPPY in ion chromatograms of individuals 1_CM16-5 and 2_CM16-6 confirms the presence of AMELY (Fig. 3a). The fragmentation spectrum of peptide SM(ox)IRPPY from individual 1_CM16-5 is reported as an example (Fig. 3b). For this individual, protein identification through database search of MS2 spectra yielded a strong match for AMELY, with a significant protein sequence coverage of 51% (Fig. 3c), unambiguously supporting our hypothesis. Other MS2 spectra are reported within the Supplementary Information (see Figs S22–S32).

Ion chromatograms representing selected peptides of the ‘Lovers’ enamel proteome20. Chromatograms search was performed using Xcalibur software (Thermo Scientific) with a mass tolerance of 5 ppm. Peptide sequences, protein names and retention times are reported in the graphs. The presence of peptide SM(ox)IRPPY (AMELY; [M + 2 H]+2 440.2233 m/z) in both the specimens suggests that the two individuals were males.

(a) Ion chromatogram representing peptides SM(ox)IRPPY, M(ox)IRPPY and SMIRPPY of individual 1_CM16-5 and 2_CM16-6; the occurrence of these three specific peaks confirms the presence of AMELY; (c) fragmentation spectrum of peptide SM(ox)IRPPY (monoisotopic mass [M + 2 H]+2 440.2233 m/z); y-axis indicates ion intensities; (c) sequence coverage (51%) of AMELY isoform 2 for individual 1_CM16-5.
To speculate on the age and the post-depositional preservation of enamel samples, we checked the rate of glutamine deamidation, a post-translational modification which involves the conversion of a glutamine (Gln) amino acid residue into glutamic acid, with an expected mass shift of +0.984 Da. Extreme environmental conditions (e.g., high temperature, pH and humidity) may accelerate protein deamidation, even though the strong mineral binding allows better preservation and survival of the protein itself 11,19. Previous work on collagen, clearly indicates that Gln deamidation strongly correlates with the thermal age of the specimens (which accounts for the burial environmental conditions) rather than the absolute chronological age21. The Gln deamidation rate of our samples was checked on those peptides containing the sequence YQSIRPPYP, namely Q57 (see11; Fig. 4). We chose to focus on Q57 to possibly compare our data with the previous study of Parker et al.11; here they also observed that the rate of deamidation at Q57 is slower than in other residue positions11. As expected, modern teeth showed a lower degree of %deQ (~0.6) while older samples (up to ~2000 years old) had a higher degree of deamidation (>0.8). Some specimens presented a %deQ close to 1 (completely deamidated), as in the case of 3_CM12-2 (4th‒6th cent.) and 12_S554 (3rd‒4th cent.). When the %deQ is plotted against the age of the sample (years before present), the dataset seems to fit a logarithmic model (R2 = 0.87, p < 0.01; Fig. 4). The only sample presenting a higher degree of deamidation in relation to its age is 5_FC176 (18th cent.), with a %deQ of 0.93 ± 0.03 (residual = 0.11; z-score = 3.0; Fig. S19). Parker et al.11 state that the rate of deamidation cannot be used to properly check the age of the sample, mostly due to the high rate of variations they observed within samples of the same age. Our samples showed a lower degree of deamidation variability compared to Parker et al.11, even if specimen 5_FC176 seems to be an outlier. High temperature and low pH may affect deamidation rate, in post-depositional environment, but also during sample pre-treatment11. Most studies (see7,11 and16) use HCl (3–5%) to extract enamel proteins, possibly causing Gln deamidation. The main difference between our study and Parker et al.11 is the fact that we extracted enamel proteins at room temperature, while Parker et al. incubated the samples at 56 °C for 60 min. Thus, to avoid an increase of the Gln deamidation rate, the extraction of enamel proteins at room or even lower temperature with diluted acid may be the best approach22. Moreover, calculating the rate of Gln deamidation by integrated peak areas rather than measured ion intensities (as11) may yield better results in terms of estimation22. However, further work on amelogenin Gln deamidation is required to properly address the abovementioned issues.

(a) Deamidation rate of Gln calculated as reported in the Method section; error bars are 1σ. Sample 5_FC176 seems to be an outlier of the regression; (b) example of ion peak integrated area (AA) for peptide YEVLTPLKWYQSIRPPYP, employed for the calculation of %deQ.
Discussion
We confidently identified the sex of all the individuals considered in this study, using enamel peptides. Individual 3_CM12-2, classified as a female by osteology, revealed the presence of AMELY within his enamel proteome, a robust marker of the male sex. Given that this individual was a young adult (~20 years) at the time of death, we suggest that the skeletal misclassification was due to his age and the related low degree of sexualization of the dimorphic districts. Such evidence clearly supports the need of alternative cost-effective methods for sex classification for doubtful skeletal materials. Compared to Stewart et al.7, we propose the use of at least two additional peptides/peaks together with m/z 440.2233 (SM(ox)IRPPY) to confidently identify AMELY as, for instance, m/z 432.2258 (SMIRPPY) and 396.7073 (M(ox)IRPPY) or other AMELY unique peptides (see Table S2). The sequence of the peptides has been further confirmed by MS2 spectra annotation.
The ‘Lovers of Modena’ have been recognized both as males by the presence of AMELY. The literature lacks of any comparable evidence (namely, hand-in-hand males) in terms of analogous geo-chronological contexts. Furthermore, to our knowledge, such a gesture was uncommon if not totally unrepresented in the art of Late Antiquity or, in general, before modern times. However, other archaeological sites, both geographically and temporally dispersed, yielded burials containing embraced or hand-in-hand individuals (see23 and references therein). This is the case, for instance, of the ‘Lovers of Valdaro’, a young female-male couple found embraced within a Neolithic tomb (San Giorgio, Mantua, Italy; ~6000 years BP). Of similar age (~5800 years BP) are the two embraced individuals discovered within the Greek site of Alepotrypa Cave. Similarly, two skeletons (male and female) were found embraced within a Turkish archaeological site (Diyarbakir, Turkey) dated to Neolithic times (~8000 years BP). The Bronze Age site of Staryi Tartas (Novosibirsk, Siberia) yielded several tombs containing female-male couples or, in some cases, even an entire family (two adults and two juveniles). The archaeologists suggest that these Bronze Age burials could be related to ritual sacrifices or to after-death beliefs23. In Romania, two skeletons dated to the Middle Ages (15th–16th century AD) were found buried hand-in-hand (Cluj-Napoca, Romania) within the cemetery of a Dominican convent23. In all cases, the burials seemed to contain female-male couples, even though a more robust sex determination by enamel proteins or DNA might be necessary to confidently classify the individuals.
We suggest that the ‘Lovers of Modena’ burial represents a voluntary expression of commitment between two individuals, rather than a recurring cult practice of the Late Antiquity; their position may reflect such relationship. The presence of several injured individuals within the Ciro Menotti necropolis let us suppose the destination of this place as war-cemetery12. In this sense, the two ‘Lovers’ could have been war comrades or friends, died together during a skirmish and, thus, buried within the same grave. Alternatively, the two individuals were relatives, possibly cousins or brothers given their similar ages, sharing the same grave due to their family bond. Although we cannot exclude that these two individuals were actually in love, it is unlikely that people who buried them decided to show such bond by positioning their bodies hand in hand. Particularly, Late Antique social attitudes and Christian religious restrictions lead to the rejection of any hypothesis of deliberate manifestation of homosexual relationship. In fact, since 390, male passivity was frowned upon by law24 and, during the reign of Justinian (527–565), sex between males was fully considered a crime24.
With this paper, we demonstrated that LC-MS/MS analysis of enamel peptides represents a robust method to estimate the sex of both ancient and modern human skeletal remains, ultimately providing an excellent contribution to paleoanthropology, bioarchaeology, and forensic anthropology. We also strengthened the method of Stewart et al.7, by proposing the use of three extracted ion chromatograms, corresponding to different AMELY unique peptides, to confidently identify protein AMELY. We applied this approach to a peculiar archaeological context, determining the sex of the ‘Lovers of Modena’, which turned out to be both males. These individuals are, thus, a unique representation of commitment between two men during the Italian Late Antiquity.
Methods
Sampling and pretreatment
The entire analytical protocol is summarized in Fig. S20. All reagents employed were of analytical grade, with the exception of hydrochloric acid being suprapur grade. Ultrapure water (18.2 MΩcm−1) was obtained from a Millipore water purification system (MilliQ Merck Millipore, Italy). From each skeletonized individual (n = 14), a tooth was extracted from the mandible or the maxilla using pliers. Two modern deciduous teeth were from personal collections and belonged to two of the authors of this paper. To improve the protein yield from the enamel tissue, we sampled small enamel chunks (ca. 10 mg) using a dentist drill (cleaned with 70% ethanol after each sampling), rather than extracting proteins by acid etch7,20. By employing this protocol, we were able to properly clean the sample surface before the actual extraction (see below) and to analyze the samples by LC-MS/MS. In comparison with the method proposed by Stewart et al.7, our protocol uses a traditional UPLC-MS/MS set up (advantage), but requires, at this stage, more material for the analysis (disadvantage). Enamel chunks were rinsed with MilliQ water in an ultrasonic bath, leached for 5 minutes with 200 μL of 5% HCl and then re-washed with MilliQ.
Enamel protein extraction and purification
To extract enamel proteins, each specimen was soaked in 200 μL of 5% HCl for 1 h in an Eppendorf tube at room temperature. After 1 h, none of the samples (ca. 10 mg of enamel) was completely digested. The supernatant was collected in a new Eppendorf tube and peptides were extracted by HyperSep SpinTips (Thermo Scientific) with C18 functionalized silica, according to the manufacturer’s protocol. Briefly, tips were conditioned three times with 50 μL of 100% acetonitrile and three times with 50 μL of 0.1% formic acid. Samples (200 μL of HCl) were loaded on the SpinTips plug in two steps of 100 μL each. Tips were then washed three times using 50 μL of formic acid 0.1%. Resin-bounded proteins were finally eluted using 10 μL of 60% acetonitrile in 0.1% formic acid, repeating the elution step twice. Samples were dried down at room temperature under a laminar flow hood (class 100). All the previously described protocols were performed at the Department of Chemical and Geological Sciences (Cleanroom facility, class 1000) and the Department of Life Sciences (Bioinorganic Chemistry and Bioelectrochemistry Lab) of the University of Modena and Reggio Emilia.
Tandem mass spectrometry
For UHPLC–HRMS analysis, dry extracted peptides were resuspended in 50 µL of a mixture of water:acetonitrile:formic acid 95:3:2, sonicated for 10 minutes at room temperature and centrifuged at 12100 r.c.f. for 10 minutes. A Thermo Scientific Dionex Ultimate 3000 UHPLC coupled to a Thermo high-resolution Q Exactive mass spectrometer (Thermo Scientific, Bremen, Germany) was used for the analyses. The column (Zorbax SB-C18 RRHT, 2.1 × 50 mm, 1.8 μ particle size, Agilent Technologies), thermostatted at 25 °C, was equilibrated with 0.3 mL/min of water 0.1% formic acid (A) with 2% acetonitrile (B); after sample injection (15 µl), B% was kept constant at 2% for 2′, then linearly increased from 2 to 28% in 64 minutes; B% was then brought to 95% in 4 minutes and kept at 95% B for 5 minutes, before the reconditioning step. Each sample required a total run time of 90 minutes. Centroided MS and MS2 spectra were recorded from 200 to 2000 m/z in Full MS/dd-MS² (TOP2) mode, at a resolution of 35000 and 17500, respectively. The two most intense multi-charged ions (TOP2) were selected for MS2 nitrogen-promoted collision-induced dissociation (NCE = 28). Precursor dynamic exclusion (6 seconds) and apex triggering (1 to 5 s) were set; peptide-like isotope pattern ions were preferred. The mass spectrometer was calibrated before the start of the analyses; an initial segment (0.1–0.7 minutes) with a lock mass (391.28429, corresponding to the [MH]+ of diisooctyl phtalate, an ubiquitous contaminant) was included in the MS method. More details are reported within the Supplementary Information.
Database search
For protein identification, raw data, converted into mascot generic format using MsConvert (v. 3.0.10730, ProteoWizard tools25), were searched against Swiss-Prot (accessed Oct 2018; 20350 sequences for Homo Sapiens) for peptide sequences and cRAP (116 sequences) for contaminants with Mascot Server (Version 2.4, Matrix Science, London, UK). No proteolytic enzyme was selected, deamidated asparagines/glutamine (NQ) and oxidated methionine (M) were set as variable modifications in the search parameters. One missed cleavage was allowed. Mass tolerances were set at 10 ppm for the precursor ions (peak detection mismatch #13C = 1) and 0.05 Da for the product ions. An automatic decoy database search was used to estimate the false discovery rate; probability threshold was trimmed to get a FDR < 1%.
Glutamine deamidation rate
Deamidation rate of glutamine (deQ) was calculated from peptides of the AMELX protein containing sequence YQSIRPPYP, e.g.: VLTPLKWYQSIRPPYP (non-deamidated [M + 3 H]3+ 653.3681 m/z; deamidated [M + 3 H]3+ 653.6961 m/z); YEVLTPLKWYQSIRPPYP (non-deamidated [M + 3 H]3+ 750.7367 m/z; deamidated [M + 3 H]3+ 751.0648 m/z); YEVLTPLKWYQSIRPPYPS (non-deamidated [M + 3 H]3+ 834.1019 m/z; deamidated [M + 3 H]3+ 834.4299 m/z); YEVLTPLKWYQSIRPPYPSYG (non-deamidated [M + 3 H]3+ 853.1090 m/z; deamidated [M + 3 H]3+ 853.4370 m/z). The proportion of Gln deamidation (%deQ) was calculated as follows (see11):
where IPndeQ is the chromatogram integrated area of the peptide peak containing non-deamidated Gln and IPdeQ is the integrated area of the peptide peak containing deamidated Gln, namely Glu. Integrated peak areas were calculated using Qual Browser (Thermo Scientific Xcalibur). At last, an average value (±SD) was calculated for each sample, combining the different deamidation rates of Gln for the different peptides. A %deQ of 0 represents a not deamidated specimen, while a %deQ of 1 represents a completely deamidated specimen.
Blinding
Mass spectrometry experiments were blinded: sample selection was performed by one of the authors of the paper (GG), assigning random numbers to the specimens, and then employed as reference numbers for the sample preparation and MS analysis. The subsequent LC-MS/MS analyses and chromatogram interpretations were performed without any prior knowledge about the skeletal sex of the individual. Only at the end of the analysis, the results about the sex of the individuals were cross-checked.
Data Availability
All data generated or analyzed during this study are included in this paper (and its Supplementary Information Files). The mass spectrometry proteomics raw data have been deposited to the ProteomeXchange Consortium via the PRIDE26 partner repository with the dataset identifier PXD012587.
References
Giles, E. & Elliot, O. Sex determination by discriminant function analysis of crania. American Journal of Physical Anthropology 21(1), 53–68 (1963).
Bruzek, J. A method for visual determination of sex, using the human hip bone. Am. J. Phys. Anthropol. 117, 157–168 (2002).
Uerpmann, H. P. Animal bone finds and economic archaeology: a critical study of ‘osteo‐archaeological’method. World Archaeology 4(3), 307–322 (1973).
Schutkowski, H. Sex determination of infant and juvenile skeletons: I. Morphognostic features. American Journal of Physical Anthropology 90(2), 199–205 (1993).
Weinstock, J. Osteometry as a source of refined demographic information: sex-ratios of reindeer, hunting strategies, and herd control in the Late Glacial site of Stellmoor, Northern Germany. Journal of archaeological science 27(12), 1187–1195 (2000).
Skoglund, P., Storå, J., Götherström, A. & Jakobsson, M. Accurate sex identification of ancient human remains using DNA shotgun sequencing. Journal of Archaeological Science 40(12), 4477–4482 (2013).
Stewart, N. A., Gerlach, R. F., Gowland, R. L., Gron, K. J. & Montgomery, J. Sex determination of human remains from peptides in tooth enamel. Proceedings of the National Academy of Sciences 114(52), 13649–13654 (2017).
Faerman, M. et al. Sex identification of archaeological human remains based on amplification of the X and Y amelogenin alleles. Gene 167(1), 327–332 (1995).
Castiblanco, G. A. et al. Identification of proteins from human permanent erupted enamel. European journal of oral sciences 123(6), 390–395 (2015).
Bansal, A. K., Shetty, D. C., Bindal, R. & Pathak, A. Amelogenin: A novel protein with diverse applications in genetic and molecular profiling. Journal of oral and maxillofacial pathology: JOMFP 16(3), 395 (2012).
Parker, G. J. et al. Sex estimation using sexually dimorphic amelogenin protein fragments in human enamel. Journal of Archaeological Science 101, 169–180 (2019).
Vazzana, A. et al. A multianalytic investigation of weapon-related injuries in a Late Antiquity necropolis, Mutina, Italy. Journal of Archaeological Science: Reports 17, 550–559 (2018).
Vazzana, A. et al. Dal caso mediatico alla musealizzazione: l’esempio della cosiddetta “Tomba degli Amanti” di Modena. Atti e Mem. – Deput. di Stor. patria per le antiche Prov. Modenesi Serie XI, 277–283 (2015a).
Vazzana, A., Pellegrini, S., Traversari, M., Labate, D. & Gruppioni, G. La necropoli tardoantica di via Ciro Menotti-Modena, analisi antropologica preliminare. Atti e Mem. – Deput. di Stor. patria per le antiche Prov. Modenesi Serie XI, 285–292 (2015b).
Brown, E. L., Dixon, R. A. & Birkett, J. W. The discolouration of human teeth from archaeological contexts: elemental analysis of a black tooth from a roman cranium recovered from the River Witham, Lincoln, UK. Journal of Anthropology, 859153 (2014).
Cappellini, E., et al. Early Pleistocene enamel proteome sequences from Dmanisi resolve Stephanorhinus phylogeny. bioRxiv, 407692 (2018).
Budd, P., Montgomery, J., Barreiro, B. & Thomas, R. G. Differential diagenesis of strontium in archaeological human dental tissues. Applied Geochemistry 15(5), 687–694 (2000).
Lugli, F., Cipriani, A., Peretto, C., Mazzucchelli, M. & Brunelli, D. In situ high spatial resolution 87Sr/86Sr ratio determination of two Middle Pleistocene (ca 580 ka) Stephanorhinus hundsheimensis teeth by LA–MC–ICP–MS. International Journal of Mass Spectrometry 412, 38–48 (2017).
Demarchi, B. et al. Protein sequences bound to mineral surfaces persist into deep time. Elife 5, e17092 (2016).
Stewart, N. A. et al. The identification of peptides by nanoLC-MS/MS from human surface tooth enamel following a simple acid etch extraction. RSC Advances 6(66), 61673–61679 (2016).
Van Doorn, N. L., Wilson, J., Hollund, H., Soressi, M. & Collins, M. J. Site‐specific deamidation of glutamine: a new marker of bone collagen deterioration. Rapid communications in mass spectrometry 26(19), 2319–2327 (2012).
Pace, A. L., Wong, R. L., Zhang, Y. T., Kao, Y. H. & Wang, Y. J. Asparagine deamidation dependence on buffer type, pH, and temperature. Journal of pharmaceutical sciences 102(6), 1712–1723 (2013).
Geller, P. L. The bioarchaeology of socio-sexual lives. Springer, New York (2016).
Harper, K. From shame to sin. Harvard University Press (2013).
Chambers, M. C. et al. A cross-platform toolkit for mass spectrometry and proteomics. Nature Biotechnology 30(10), 918 (2012).
Perez-Riverol, Y. et al. The PRIDE database and related tools and resources in 2019: improving support for quantification data. Nucleic Acids Res 47(D1), D442–D450 (2019).
Frigerio, M. & Vazzana, A. L’area sepolcrale rinvenuta in Piazza Grande, in: Mutina Splendidissima. La Città Romana e La Sua Eredità. De Luca Editori d’Arte srl, Roma, pp. 399–400 (2017).
Giorgi, E. & De Maria, S. Urbanistica e assetti monumentali di Suasa. Novità dalle ricerche recenti, in: Epigrafia e Archeologia. Atti Del Convegno Di Studi (Macerata, 22–23 Aprile 2013). pp. 79–142 (2013).
Acsádi, G. & Nemeskéri, J. History of human life span and mortality. Akadémiai Kiadó (1970).
Meindl, R. S. & Lovejoy, C. O. Ectocranial suture closure: A revised method for the determination of skeletal age at death based on the lateral-anterior sutures. Am. J. Phys. Anthropol. 68, 57–66 (1985).
Schmitt, A. Une nouvelle méthode pour estimer l’âge au décès des adultes à partir de la surface sacro-pelvienne iliaque. Bull. Mémoires la Société d’Anthropologie Paris 2, 89–101 (2005).
Brooks, S. & Suchey, J. M. Skeletal age determination based on the os pubis: a comparison of the Acsádi-Nemeskéri and Suchey-Brooks methods. Human evolution 5(3), 227–238 (1990).
Acknowledgements
This research was funded by the European Research Council (ERC) under the European Union’s Horizon 2020 Research and Innovation Programme (grant agreement No. 724046 – SUCCESS awarded to Prof. Stefano Benazzi – erc-success.eu). The authors acknowledge the ‘Fondazione Cassa di Risparmio di Modena’ for funding the UHPLC-ESI-Q Exactive system at the Centro Interdipartimentale Grandi Strumenti (CIGS). This research was conducted within the framework of the ‘Da San Geminiano alla cattedrale romanica’ project, one of the 2018–2020 goals of the UNESCO site ‘Modena: Cattedrale, Torre Civica e Piazza Grande’, coordinated by ‘Musei Civici di Modena’ and ‘Soprintendenza Archeologia, Belle Arti e Paesaggio per la Città Metropolitana di Bologna e le province di Modena Reggio Emilia e Ferrara’.
Author information
Authors and Affiliations
Contributions
F.L., G.D.R. and S.B. conceived the study and designed the experiments; F.L. and G.D.R. performed the protein extraction and purification; F.G. and D.P. performed the mass spectrometry analyses; F.L., G.D.R. and F.G. analyzed the data and ran the Mascot searches; A.V., G.G. and G.O. provided the samples and determined the sex and the age of the individuals; S.B. supervised the whole work; all the authors discussed the data and contributed to writing the manuscript.
Corresponding authors
Ethics declarations
Competing Interests
The authors declare no competing interests.
Additional information
Publisher’s note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Lugli, F., Di Rocco, G., Vazzana, A. et al. Enamel peptides reveal the sex of the Late Antique ‘Lovers of Modena’. Sci Rep 9, 13130 (2019). https://doi.org/10.1038/s41598-019-49562-7
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-019-49562-7
This article is cited by
-
A label-free quantification method for assessing sex from modern and ancient bovine tooth enamel
Scientific Reports (2024)
-
Windows into the past: recent scientific techniques in dental analysis
British Dental Journal (2024)
-
Amelogenin peptide analyses reveal female leadership in Copper Age Iberia (c. 2900–2650 BC)
Scientific Reports (2023)
-
Osteological, multi-isotope and proteomic analysis of poorly-preserved human remains from a Dutch East India Company burial ground in South Africa
Scientific Reports (2023)
-
Unbalanced sex-ratio in the Neolithic individuals from the Escoural Cave (Montemor-o-Novo, Portugal) revealed by peptide analysis
Scientific Reports (2023)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
