
Abstract
Reports that aging slows down in space prompted this investigation of anti-aging effects in humans by analyzing astronauts’ heart rate variability (HRV). Ambulatory 48-hour electrocardiograms from 7 astronauts (42.1 ± 6.8 years; 6 men) 20.6 ± 2.7 days (ISS01) and 138.6 ± 21.8 days (ISS02) after launch were divided into 24-hour spans of relative lower or higher magnetic disturbance, based on geomagnetic measures in Tromso, Norway. Magnetic disturbances were significantly higher on disturbed than on quiet days (ISS01: 72.01 ± 33.82 versus 33.96 ± 17.90 nT, P = 0.0307; ISS02: 71.06 ± 51.52 versus 32.53 ± 27.27 nT, P = 0.0308). SDNNIDX was increased on disturbed days (by 5.5% during ISS01, P = 0.0110), as were other HRV indices during ISS02 (SDANN, 12.5%, P = 0.0243; Triangular Index, 8.4%, P = 0.0469; and TF-component, 17.2%, P = 0.0054), suggesting the action of an anti-aging or longevity effect. The effect on TF was stronger during light (12:00–17:00) than during darkness (0:00–05:00) (P = 0.0268). The brain default mode network (DMN) was activated, gauged by increases in the LF-band (9.7%, P = 0.0730) and MF1-band (9.9%, P = 0.0281). Magnetic changes in the magnetosphere can affect and enhance HRV indices in space, involving an anti-aging or longevity effect, probably in association with the brain DMN, in a light-dependent manner and/or with help from the circadian clock.
Similar content being viewed by others


Introduction
Aging is the inevitable time-dependent decline in physiological organ function and a major risk factor for cardiovascular disease, cancer and Alzheimer’s disease. Novel treatments and translational approaches have been tried to prevent, delay, alleviate or even reverse age-related diseases. Maximal longevity, however, appears unchanged. Space research is bringing insight on how to directly intervene against the aging hallmarks and prevent age-related diseases. This possibility may exist because spaceflight has complex effects on the physiology of organisms, such as reduced gravity and different electromagnetic surroundings, which induce changes to almost every system in the body.
The Sun markedly affects most activities on Earth, including its solid, liquid and gas states. Whether life started on Earth or in the cosmos (panspermia hypothesis), it evolved following long physical/chemical processes, so that life on Earth matches what this planet requires1,2. Now, humans plan to live on the moon or Mars3,4. Spaceflight opens new opportunities to learn about capabilities of the human body that could not be studied on Earth. Spaceflight is known to dramatically alter cardiovascular dynamics5,6,7,8,9,10,11, also posing significant risks, such as an overall greater than 10-fold faster onset and time course of muscle and bone atrophy12,13,14,15. Microgravity is one of the crucial contributors to these observed physiological changes.
How microgravitational space environments affect aging is not well understood16,17. Honda et al.18 reported that spaceflight in Caenorhabditis elegans suppressed the formation of transgenically-expressed polyglutamine aggregates, which normally accumulate with increasing age. Inactivation of each of the seven genes that were down-regulated in space was found to extend lifespan on the ground. Aging in Caenorhabditis elegans seemed to be slowed through neuronal and endocrine responses to space environmental cues.
Whether similar anti-aging effects apply to astronauts during long-term missions in space is investigated herein by analyzing heart rate variability (HRV) on quiet versus magnetically-disturbed days. Exposure to weak geomagnetic fields is associated with biological effects1,19,20,21,22. Previously we showed that magnetic disturbances suppress HRV primarily in the very-low frequency region, which is clinically important since its reduction is a predictor of morbidity and mortality from cardiovascular disease23,24. HRV in the minutes to hours (very-low to ultra-low frequency) range is also a powerful predictor of longevity in clinically healthy people.
Methods
Experimental conditions on the ISS
Universal Time Coordinated (UTC) is used aboard the ISS. The windows are covered during night hours to give the impression of darkness because the station experiences 16 sunrises and sunsets per day.
A typical day for astronauts begins with awakening at 06:00, followed by post-sleep activities and a morning inspection of the station. The crew then eats breakfast and takes part in a daily planning conference with Mission Control before starting work at around 08:10. The first scheduled exercise of the day follows, after which the crew continues work until 13:05. Following a one-hour lunch break, the afternoon consists of more exercise and work before astronauts carry out their pre-sleep activities beginning at 19:30, including dinner and a crew conference. The scheduled sleep span begins at 21:3025.
Subjects
Of the 10 healthy astronauts who participated in ISS JAXA investigation named “Biological Rhythms 48 Hrs”, 3 were excluded because their data were insufficient for the purpose of this study. There was partial shortage of their pre- and post-flight ECG data probably due to poor contact of the electrodes. The mean (±SD) age of the 7 healthy astronauts (6 men, 1 woman) included in this investigation was 42.1 ± 6.8 years. Their mean stay in space was 151.3 ± 21.8 days. Astronauts had passed class III physical examinations from the National Aeronautics and Space Administration (NASA). The study was approved by the Institutional Review Boards of NASA, ESA (European Space Agency) and JAXA (Japan Aerospace Exploration Agency). Informed consent was obtained from all subjects. A detailed explanation of the study protocol was given to the subjects before they gave written, informed consent, according to the Declaration of Helsinki Principles. All methods were performed in accordance with the JAXA/ESA/NASA guidelines and regulations.
Experimental protocol
Ambulatory around-the-clock 48-hour electrocardiographic (ECG) records were obtained by using a two-channel Holter recorder (FM-180; Fukuda Denshi). Measurements were made four times: once before flight (Pre); two times during flight on the International Space Station (ISS): ISS01 (20.6 ± 2.7, 18–25 days), and ISS02 (138.6 ± 21.8, 101–159 days) after launch; and once after return to Earth (Post). The 48-hour records during ISS01 and ISS02 were divided into 24-hour spans of relative lower or higher magnetic disturbance for comparison of HRV endpoints between “quiet” and “disturbed” conditions.
Assessment of space magnetics
The ISS is protected from the space environment by Earth’s magnetic field. The ISS orbits the Earth every 90 min at an altitude of 330 to 480 km. Izumi et al.26 reported that the geomagnetic strength at 400 km is about 4/5 or 3/4 that at sea level. Lacking direct measurement on the ISS, we used geomagnetic measurements at 1-min intervals from the Auroral Observatory of the University of Tromsø, in Tromsø, Norway (69°39′N, 18°56′E): total intensity (F in nT), declination (D, angle between geographic and magnetic north, in degrees), inclination (I, angle between horizontal plane and magnetic direction, in degrees), horizontal intensity (H in nT), and vertical intensity (Z in nT).
Historically, magnetic records obtained from the ground have widely been used to study physical processes that occur in the near-Earth environment. Kamide et al.27 proposed various models of the estimation of ionospheric electric field, ionospheric currents and field-aligned currents from ground magnetic records, along with incoherent scatter radars, satellite measurements of X-ray and UV aurorae. This type of study over the past two hundred years about the structure and temporal changes of the Sun–Earth space, now called space weather, has now become of interest to space science. However, it is still extremely difficult to specify the state of geomagnetics in space2,28,29,30,31. Humans are not consciously aware of changes in space weather. Except for the presence of aurorae, which occur when magnetic disturbances exceed those encountered in the current study, external cues are also lacking.
Analysis of HRV
Data collection and measurement procedures were conducted as previously reported8,9,10,11. Briefly, for HRV measurements, QRS waveforms were read from continuous ECG records. The RR intervals between normal QRS waveforms were extracted as normal-to-normal (NN) intervals, which were A/D converted (125-Hz) with 8-ms time resolution. After the authors confirmed that all artifacts were actually removed and that the data excluded supraventricular or ventricular arrhythmia, time-domain HRV indices (SDNN, SDANN, SDNNIDX and Triangular Index, TI), and conventional frequency-domain measures (HF: 0.15–0.40 Hz, LF: 0.04–0.15 Hz, and VLF: 0.003–0.04 Hz)32 were obtained with the MemCalc/CHIRAM (Suwa Trust GMS, Tokyo, Japan) software33. Time series of NN intervals were also processed consecutively in 180-min intervals, progressively displaced by 5 min, to estimate TF (0.0001–0.50 Hz), ULF (0.0001–0.003 Hz), and the 1/fβ-type scaling in HRV. Focus was placed on the frequency range of 0.0001–0.01 Hz (periods of 2.8 hours to 1.6 minutes).
Time series of NN intervals covering 5-min intervals were processed consecutively, and spectral power in four frequency regions were computed using the Maximum Entropy Method (MEM): LF-band (0.01–0.05 Hz), MF1-band (0.05–0.10 Hz), MF2-band (0.10–0.15 Hz), and HF-band (0.15–0.20 Hz) according to Baria et al.11,34. A positive response in these bands is hypothesized to indicate how astronauts adapt to the space environment: the LF- and MF1-bands reflect an activation of the DMN’s medial prefrontal cortex (mPFC), posterior parietal cortex, posterior portion of precuneus and posterior cingulate cortex (PCC), while the MF2- and HF-bands show an activation of the orbitofrontal and temporal cortex parts of the DMN34.
Circadian differences in HRV response to magnetic disturbance in space
Circadian stage-dependent HRV responses to magnetic disturbances in space were assessed by subdividing the 24-hour day into 4 time spans of 5 hours each: morning (around 08:30, 06:00–11:00); daytime (around 14:30, 12:00–17:00); evening (around 20:30, 18:00–23:00); and night (around 02:30, 00:00–05:00).
Statistical analyses
Data were expressed as mean ± standard deviation (SD). HRV endpoints were compared among the 4 sessions by 1-way ANOVA, with contrasts. Of particular interest were comparisons of ISS01 vs. Pre and ISS02 vs. Pre, which reflect the extent of astronauts’ adaptation to the space environment.
Comparisons between magnetically quiet and disturbed days used the two-sided paired-t test. Effects of magnetic activity on HRV endpoints were tested in 4 spans (morning, daytime, evening and night) for each astronaut by Student t-test, adjusted for unequal variances and unequal sample sizes according to Welch’s correction. Subsequently, the number of statistically significant individual responses during morning, daytime, or evening versus night was compared by χ2 or Fisher exact test.
The Stat Flex (Ver. 6) software (Artec Co., Ltd., Osaka, Japan) was used. P-values less than 0.05 were considered to indicate statistical significance.
Results
Assessment of space magnetics on the ISS
Geomagnetic indices from the Auroral Observatory of the University of Tromso are compared between quiet and disturbed days in Table 1. The 24-hour SD of Z (magnetic field’s vertical component) was about twice as large on disturbed than on quiet days (ISS01: 72.01 ± 33.82 versus 33.96 ± 17.90 nT, P = 0.0307; ISS02: 71.06 ± 51.52 versus 32.53 ± 27.27 nT, P = 0.0308), and so was the 24-hour SD of the declination. Variations in H (magnetic field’s horizontal component) and inclination were larger on disturbed than on quiet days during ISS01, while during ISS02 those in total intensity were larger. No differences were found in the 24-hour averages (Table 1).
Effects of magnetics on astronauts’ HRV in space
Alterations of HRV associated with geomagnetic activity are summarized in Table 2. A 4.0% decrease in the 24-hour average of HR (P = 0.0392) was observed during ISS01. Parasympathetic activity estimated by r-MSSD and pNN50 showed no statistically significant changes, whereas sympathetic activity estimated by LF/HF increased by 13.7% during ISS02 (Table 2, top). No difference in β was found, suggesting that magnetic disturbances of magnitude similar to that taking place during ISS01 and ISS02 may not influence the intrinsic cardiovascular autonomic regulatory system.
Heart rate variability related to possible anti-aging or longevity
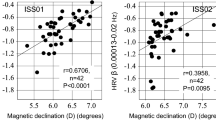
HRV indices suggestive of anti-aging or longevity effects include SDNN and SDANN. Although the cardiovascular autonomic regulatory function, reflected by β, was statistically significantly suppressed (Fig. 1, left), SDNN (Fig. 1, middle) and SDANN (Fig. 1, right), computed over 24 hours, show HRV plasticity already after about 3 weeks as well as after 6 months in space.

HRV plasticity observed in SDNN and SDANN, as indices of anti-aging or longevity. Although the cardiovascular autonomic regulatory function, reflected by β, was statistically significantly suppressed (left), SDNN (middle) and SDANN (right), computed over 24 hours, show a quality of HRV plasticity after about 6 months in space. Symbols were assigned to individual astronauts.
SDNNIDX is statistically significantly higher on disturbed than on quiet days during ISS01 (by 5.5%, P = 0.0110) and ISS02 (by 6.5%, P = 0.0405). During ISS02, 3 other HRV indices are elevated on disturbed days: SDANN (12.5%, P = 0.0243), Triangular Index (8.4%, P = 0.0469) and TF (17.2%, P = 0.0054) (Table 2, upper middle); SDNN shows a similar trend (10.0%, P = 0.0806).
The larger TF power on disturbed days (ISS02) was contributed by higher ULF (31.0%, P = 0.0141) and VLF (12.7%, P = 0.0320) power; changes in LF (9.0%, P = 0.0737) and HF (−7.7%, P = 0.2825) power are not significant. SDNNIDX is known as a surrogate for VLF, which was already higher on disturbed days during ISS01 (14.1%, P = 0.0033) (Table 2, lower middle). The frequency-domain HRV endpoints of a 2-day record from one astronaut, split between the quiet and disturbed day, are illustrated in Fig. 2 together with the magnetic declination index for those two days. In this case, not only TF (P < 0.0001), ULF-(P = 0.0002) and VLF (P < 0.0001), but also LF (P = 0.0284) power is higher on the disturbed day, as shown by Student’s t-test.

An astronaut’s circadian profiles of frequency-domain measures of HRV during ISS02 are compared between the quiet (left) and magnetically disturbed (right) day. HRV endpoints were computed by MEM spectra over 5-min or 180-min intervals and compared between the 2 days by Student’s t test. Values shown in individual graphs indicate spectral power cumulated over 24 hours. The larger values during disturbed vs. quiet conditions indicate a statistically significant response to magnetic disturbance in space. Not only TF (P < 0.0001), ULF (P = 0.0002) and VLF (P < 0.0001), but also LF (P = 0.0284) power (in msec2) were found to be statistically significantly higher on disturbed than on quiet days.
Involvement of the default mode network (DMN) in humans’ magnetoreception in space
Examining the 4 HRV bands reflecting the brain’s DMN activity, MF1-band is statistically significantly higher on disturbed than on quiet days during ISS02 (9.9%, P = 0.0281). LF-band also shows a similar trend (9.7%, P = 0.0730) (Table 2, bottom). No differences in MF2- and HF-bands are found, however. Figure 3 illustrates the simultaneous records of the geomagnetic declination and vertical intensity together with HRV’s LF-, MF1-, MF2- and HF-bands of one astronaut. In this case, all 4 HRV bands reflecting the brain’s DMN activity are higher on the disturbed than on the quiet day (P < 0.0001), as shown by Student’s t-test.

An astronaut’s circadian profiles of HRV measures related to DMN activity during ISS02 are compared between the quiet (left) and magnetically disturbed (right) day. HRV endpoints were computed by MEM spectra over 5-min or 180-min intervals and compared between the 2 days by Student’s t test. Values shown in individual graphs indicate spectral power cumulated over 24 hours. Spectral power (in msec2) of all 4 HRV bands (LF, MF1, MF2, and HF) were found to be higher on the magnetically disturbed than on the quiet day (P < 0.0001).
Circadian stage-dependent HRV response to magnetic disturbance in space
When subdividing the 24-hour day into 4 time spans of 5 hours each, centered around 08:30 (morning), 14:30 (daytime), 20:30 (evening) and 02:30 (night), the effect of magnetic activity in space was found to be circadian stage-dependent during ISS02. More astronauts had higher TF power on disturbed than on quiet days during the daytime and evening than during the night (P = 0.0308 by χ2 test). During ISS01, more astronauts also tended to have a higher VLF power and higher SDNNIDX on disturbed days during the daytime than during the night (P = 0.0943 by χ2 test), but they had less HF power in the evening than at night (P = 0.0962 by Fisher exact test) (Table 3).
Discussion
To examine whether the space environment may convey anti-aging effects in humans, HRV indices of astronauts on long-term missions were compared between days with relative lower or higher magnetic activity. Tromso’s geomagnetic measures served as surrogate since the ISS is protected by Earth’s magnetosphere. The fact that already during ISS01 SDNNIDX, a surrogate of the VLF-component, is increased on disturbed days suggests an anti-aging effect, as do similar increases during ISS02 in SDNN (a surrogate for the TF-component), SDANN, the Triangular Index and TF-component. The effect of magnetic activity on TF, suggesting an anti-aging effect, was found to be circadian stage-dependent, being more effective during light (12:00–17:00) than during dark (0:00–05:00) (P = 0.0268). The magnetic field in space also induced an activation of the brain’s DMN, gauged by HRV (increase in LF- and MF1-bands). An anti-aging effect in space may thus be mediated by the DMN in a light-dependent manner or/and with help from the circadian clock.
An anti-aging effect of the space environment was reported by Honda et al.18 on Caenorhabditis elegans. While their results indicate that space-flown worms age more slowly compared with ground control worms, the authors note that male Drosophila lived shorter on the ground after spaceflight compared to controls maintained on the ground through their lifetimes18. Caenorhabditis elegans is a classic animal model used to help understand human health concerns due to its short lifespan and ease of culture35,36. Simulated microgravity reportedly could affect early embryogenesis, reproduction and locomotion behavior, and potentially cause the oxidative stress and DNA damage in nematodes37,38,39,40,41.
In the study by Honda et al.18, the inactivation of each of seven genes that were down-regulated in space extended lifespan on the ground. These genes encode proteins that are likely related to neuronal or endocrine signaling: acetylcholine receptor, acetylcholine transporter, choline acetyltransferase, rhodopsin-like receptor, glutamate-gated chloride channel, shaker family of potassium channel, and insulin-like peptide. One of the genes identified encodes insulin, which is associated with metabolic control. In humans, insulin is also associated with modulation of lifespan. Li et al.42 showed that simulated microgravity could significantly increase the expression of p38 MAPK signaling in the intestine, which may mediate a protection mechanism for animals against the adverse effects from simulated microgravity. Studies in space may provide information about human life that cannot be learned on Earth, including lessons about aging.
The recently published NASA Twins Study describing a multidimensional analysis of a year-long human spaceflight43 also reported that telomeres predominantly lengthened on the ISS, suggesting an anti-aging effect of the space environment. This intriguing change at the chromosomal level, however, could also be interpreted as indicating an increased cancer risk, related to the enhanced exposure to ionizing radiation in space, warranting further investigation of how space weather may affect long-term missions in space.
HRV is defined as the beat-to-beat alterations to the sinus rhythm which result from the interactions between sympathetic and parasympathetic activity32. But HRV is probably more than an indicator of probable disturbances in the autonomous system44. HRV has been considered as a surrogate parameter on complex interactions within biological systems. The relation between reduced HRV and mortality risk was first shown by Wolf et al. in 197845. High HRV is usually associated with good health, whereas low HRV might signify pathological changes, in agreement with the relation between lower HRV parameters and adverse cardiovascular outcomes46,47,48. HRV has recently been considered to reflect the state of the brain as much as the state of the heart49,50,51,52, and is viewed to also reflect the state of health and well-being53,54,55,56, a feature increasingly recognized as a hallmark of anti-aging. Whereas reduced HRV is associated with major risk factors of cognitive impairment, increased HRV indices (including SDNN, SDANN, Triangular Index and TF-, ULF- and VLF-components) are associated with better executive function in the middle aged and elderly57,58,59,60.
The atmosphere gradually ends a hundred kilometers above Earth’s surface. The Earth resides in a vast cavity called the magnetosphere, created by the interaction between Earth’s magnetic field and the solar wind, a gas of charged particles flowing continuously from the Sun. The ISS, which orbits the Earth at an altitude of 330 to 480 km, is also protected from the space environment by Earth’s magnetic field2,26. While conditions in the magnetosphere and on the Sun can potentially influence human health1,2,19, very little is known about their role in the adaptive response of humans to the space environment. The present study is the first to investigate this question.
Time- and frequency-domain HRV endpoints have served as potential markers of stress in organismic functions associated with adaptability and health. For instance, several recent reviews report significantly longer overall survival in cancer patients with higher versus lower HRV55,61,62,63. SDNNIDX and the VLF-component both increased in space in response to magnetic activity after just about one month on the ISS, meaning an activation of HRV in the 0.003–0.04 Hz frequency range. Definite increases in VLF, SDANN, TI, TF, and ULF were later observed on disturbed days during ISS02. SDNN also showed an increasing trend during ISS02. Exposure to variable magnetic fields in space resulted in an increase in several HRV endpoints, suggesting that aging may also be slowed down in astronauts in space, as it was for Caenorhabditis elegans18. The fact that VLF showed a strong response may point to anti-aging effects since VLF power is an indicator of health, lower VLF power being associated with arrhythmic death46 and inflammation64,65.
Why does the VLF-component act most prominently on the autonomous system? Historically, the physiological mechanisms involved in the long-term regulation mechanisms and autonomic nervous activity were linked to thermoregulation, the renin-angiotensin system and other hormonal factors66,67. Studying the autonomic nerves’ re-innervation in the transplanted heart, however, Kember et al.68,69 suggested that the VLF component was generated by the stimulation of afferent sensory neurons in the heart, which in turn activated various levels of the feedback and feed-forward loops in the heart’s intrinsic cardiac nervous system70. The VLF rhythm is now understood to be intrinsically generated by the heart itself, and that the amplitude and frequency of these oscillations are modulated by efferent sympathetic activity. As such, it should be fundamental to health and well-being. An enhanced VLF appearing soon after arrival at the ISS, persisting throughout the space mission may thus represent the most fundamental response to maintain health in space.
During the 5-year experimental span in solar cycle 24, an increased response of HRV was observed in space: 10% in SDNN, 12.5% in SDANN, 8.4% in TI, 17.2% in TF, 31.0% in ULF, 12.7% in VLF, and 9.0% in LF power (Table 2). By contrast, our previous 1998–2000 studies (during solar cycle 23) in a subarctic area indicated that magnetic storms, which involved larger magnetic disturbances than those observed during ISS01 and ISS02, suppressed HRV indices in 19 clinically healthy subjects24. The decrease in HRV was statistically validated for TF (−18.6%, P = 0.00009), primarily contributed to by VLF (−21.9%, P < 0.000001) in conjunction with ULF (−15.5%, P = 0.00865) and LF (−14.2%, P = 0.00187). Solar activity undergoes an about 11-year cycle. HRV decreases associated with geomagnetic storms, which occur more frequently during solar maxima, may increase the cardiovascular disease risk of susceptible individuals. Such an outcome was witnessed in data on mortality from myocardial infarction in Minnesota, which show a 5% increased mortality during years of maximal solar activity (P = 0.023)71. Factors underlying the difference in response observed in space or subarctic areas will need further investigation based on concomitant longitudinal geomagnetic and biomedical monitoring in order to better understand any anti-aging benefits of the space environment, which could then serve to design countermeasures to prevent adverse vascular events on Earth.
Our previous investigation showed a graded response of HRV depending on the degree of geomagnetic activity (gauged by the geomagnetic index ap) in healthy young men living above the arctic circle72,73. Specifically, as compared to days when ap < 7, on days when 7 < ap < 20 or days when 20 < ap < 45, TF was decreased by 18.1% and 31.6%, ULF by 18.1% and 27.5%, and VLF by 12.9% and 28.6%, respectively. A graded decrease of HRV to geomagnetic activity suggests the existence of human magnetoreceptors. Phillips et al.74 proposed a light-dependent magnetoreception mechanism and established a link between magnetic field sensitivity and the visual system in eastern red-spotted newts. A magnetoreception mechanism may thus also exist in humans.
Cryptochromes are blue-light absorbing flavoproteins that in animals have an important function in the circadian clock75,76,77. Furthermore, it has been suggested that cryptochromes act as receptor molecules with magnetically sensitive radical pair reactions in the light-dependent magnetic compass sense of life on Earth78,79,80,81,82. Many animals, including birds, flies, bats, turtles, ants, mole rats, foxes, cows and deer, use the Earth’s magnetic field for orientation and navigation. Nießner et al.83 also localized Cry1 in the retina of Canidae, Mustelidae, Ursidae and some Primates. While humans are not consciously aware of changes in magnetic field and while human magnetoreception has been rarely tested, yielding inconclusive results, a new study84 reports a strong and reproducible human brain response to geomagnetic stimulation, involving a drop in amplitude of EEG alpha oscillations (8–13 Hz) in association with horizontal rotations when the static vertical field is directed downwards. Such alpha event-related desynchronization has previously been linked to sensory and cognitive processing of external stimuli, including vision, auditory and somatosensory cues. Humans are also known to have two cryptochromes (CRY1 and CRY2), which act as clock proteins coordinating the circadian system. Foley et al.85 found that human cryptochrome is sensitive to blue light. Using a transgenic approach, they showed that human CRY2 can act as a magnetic sensor.
Mutant flies lacking the gene for cryptochrome did not respond to magnetic fields, but by transfecting them with two different cryptochrome genes from monarch butterflies they responded to magnetic fields under full-spectrum light78,80. These deletion and replacement experiments showed that the cryptochrome is an essential feature of the light-dependent magnetic sensing system in animals. These animals, however, were exposed to magnetic fields 8 to 10 times stronger than the geomagnetic field on Earth, and similar experiments cannot be conducted in humans, of course.
Our previous investigation showing HRV suppression on days of high geomagnetic disturbance24 found a decrease in HRV only when daylight alternated with darkness (above the arctic circle, there are days of complete light or darkness), suggesting a mechanism sensitive to the alternation of light (L) and darkness (D). The hypothesis of a light-dark-influenced magnetoreception was supported by cross-spectral analysis. Group-averaged coherence at frequencies coincident with the geomagnetic Pc6 pulsations (periods ranging from 10 minutes to 5 hours) differed among the three natural lighting conditions, the association being weaker during L/L or D/D than during D/L (P < 0.000001). A light-dark-influenced magnetoreception mechanism in humans may thus involve the Pc6 band of the magnetic field. Only the magnetoreception in HRV VLF-component was equal throughout the seasons, irrespective of the three natural lighting conditions, suggesting that biological signals involving VLF, which is vital for life, are set-up to be highly sensitive to geomagnetics.
A circadian stage-dependent response of HRV to magnetic activity in space was also found in the present study, the most sensitive response being observed during the daytime (12:00–17:00) and evening (18:00–23:00) (Table 3), when the intensity of illumination on board was around 700 lux. This result is in agreement with the possibility that humans have a light-dependent magnetoreception mechanism that remains functional in space.
Recently we reported that adaptation to microgravity occurs during long-duration spaceflight primarily according to two primary processes, one involving the dynamics of large-scale brain networks, initiated by the default mode network (DMN), and another coordinated by the circadian system11. The adaptation process proceeded even in the absence of consciousness. Herein, we also observed an acceleration of the DMN activity gauged by HRV (Table 2). Our results lead to the hypothesis that exposure to magnetic variation in space, via the retina, or the brain directly, may activate the dynamics of large-scale brain networks, including the DMN. The resulting HRV changes involved an anti-aging effect, enhanced after 6 months in space, together with an adaptation to the novel space environment.
The idea that space-magnetics may directly affect DMN activity, perhaps bringing about an anti-aging effect, is not surprising in view of recent documentation of transcranial magnetic stimulation (TMS) results now being introduced into clinical practice86,87,88. Alterations in DMN activity patterns strongly correlated with depression. In depressed people, disordered activity in the DMN correlates with negative rumination. Liston et al.89 reported that TMS seemed to create a shift in the relationship between two nodes in the DMN, the medial prefrontal cortex and the dorsolateral prefrontal cortex. The medial prefrontal cortex is involved in threat assessment and emotional decision making. The dorsolateral prefrontal cortex, which is where TMS stimulation takes place, seems to be involved in more thoughtful decision making, holding multiple ideas in mind and trying to understand before judging. This shift in functional brain activity could account for the mechanism of action of TMS. It should be noted, however, that magnetic variations in space, of the order of 54000 nT with a SD of 35 to 70 nT, are more than 10,000 times weaker than TMS (1–2 Tesla).
Limitations
This is the first report investigating the effect of magnetic activity on humans’ HRV in space. Whereas it may seem that differences in geomagnetic indices were stronger during ISS01 than during ISS02, these differences may stem mostly from smaller SDs rather than from larger differences per se. Accordingly, it may no longer be surprising that some differences in HRV endpoints are more strongly expressed during ISS02 than during ISS01, a fact that may also be accounted for by the longer adaptation of astronauts to the space environment (see e.g., trajectories of LF/HF, TF, ULF and LF components, and LF-band).
The study was limited by the fact that geomagnetic measures in Tromso, Norway, were used instead of space weather measurements on the ISS. Magnetic activity during ISS01 and ISS02 did not reach levels found during magnetic storms, as it did in our subarctic study. Further studies are needed to assess the relative benefits and risks associated with the full spectrum of space weather conditions on the ISS.
The relatively short ECG records of only 48 hours from only 7 astronauts, monitored twice during a 6-month space mission is another limitation. These records were obtained during a span shorter than the solar activity cycle. Future work is needed to determine whether similar results can be obtained on additional astronauts, preferably based on longer ECG records, so that graded HRV responses to space weather can also be explored. Although larger and longer studies will also be needed to decide whether the physiological changes observed herein actually reflect an anti-aging effect, our results contributed novel information to better understand aging.
Conclusion
Life on Earth is protected by the atmosphere and the magnetic field. The Earth’s magnetic field can affect humans’ life, especially HRV indices1,19,20,21,22,23,24,71,72,73. We showed that the magnetic field can affect and enhance HRV indices involved in longevity, notably during the daytime and evening, probably in association with an elevated activity of brain’s DMN with help from the circadian clock. Anti-aging HRV indices were significantly increased on days of higher magnetic activity. The difference between our results herein and those of our previous observation in a subarctic area may relate to a difference in extent of disturbance of the magnetic field between the two studies.
Spaceflight produces changes that induce a state of frailty akin to aging, such as muscle and bone atrophy, and balance and coordination problems, which can mostly be reversed upon re-adaptation to life on Earth. As proposed by Vernikos et al.17,90, the aged human may reflect a figure of life as a long-term adaptation for avoiding gravity effects on Earth in the course of aging. Our results provide a start on a catalog of measurable components of aging, and indicate what may be effective ways to slow down or even reverse the aging process.
References
Halberg, F. et al. Cycles tipping the scale between death and survival (=life). Prog. Theor. Phys. Suppl. 173, 153–181 (2008).
Kamide, Y. Our life is protected by the Earth’s atmosphere and magnetic field: what aurora research tells us. Biomed. Pharmacother. 55, 21–24 (2001).
Newman, D. J. Life in extreme environments: how will humans perform on Mars? Gravit. Space Biol. Bull. 13, 35–47 (2000).
Charles, J. B. A year on the International Space Station: preparation for and implementation of a long-duration biomedical research mission. Aerosp. Med. Hum. Perform; in press (2018).
Hughson, R. L. et al. Cardiovascular regulation during long-duration spaceflights to the International Space Station. J. Appl. Physiol. 112, 719–727 (2012).
Xu, D. et al. Reduced heart rate variability during sleep in long-duration spaceflight. Am. J. Physiol. Regul. Integr. Comp. Physiol. 305, R164–R170 (2013).
Norsk, P., Asmar, A., Damgaard, M. & Christensen, N. J. Fluid shifts, vasodilatation and ambulatory blood pressure reduction during long duration spaceflight. J. Physiol. 593, 573–584 (2015).
Yamamoto, N. et al. Effects of long-term microgravity exposure in space on circadian rhythms of heart rate variability. Chronobiol. Int. 32, 327–340 (2015).
Otsuka, K. et al. Intrinsic cardiovascular autonomic regulatory system of astronauts exposed long-term to microgravity in space: observational study. npj Microgravity 1, 15018; https://doi.org/10.1038/npjmgrav.2015.18 (2015). Erratum in: npj Microgravity 8, 12; https://doi.org/10.1038/npjmgrav.2016.31 (2016).
Otsuka, K. et al. Long-term exposure to space’s microgravity alters the time structure of heart rate variability of astronauts. Heliyon 2, e00211, https://doi.org/10.1016/j.heliyon.2016.e00211 (2016).
Otsuka, K. et al. Circadian challenge of astronauts’ unconscious mind adapting to microgravity in space, estimated by heart rate variability. Sci. Rep. 8, 10381, https://doi.org/10.1038/s41598-018-28740-z (2018).
Miyamoto, A. et al. Medical baseline data collection on bone and muscle change with space flight. Bone 22(5 Suppl.), 79S–82S (1998).
Goodship, A. E. et al. Bone loss during long term space flight is prevented by the application of a short term impulsive mechanical stimulus Acta Astronaut. 43, 65–75 (1998). Erratum in: Acta Astronaut. 45, 59 (1999).
Leblanc, A. et al. Bisphosphonates as a supplement to exercise to protect bone during long-duration spaceflight. Osteoporos. Int. 24, 2105–2114 (2013).
Gopalakrishnan, R. et al. Muscle volume, strength, endurance, and exercise loads during 6-month missions in space. Aviat. Space Environ. Med. 81, 91–102 (2010).
Honda, Y., Honda, S., Narici, M. & Szewczyk, N. J. Spaceflight and ageing: reflecting on Caenorhabditis elegans in space. Gerontology 60, 138–142 (2014).
Vernikos, J. & Schneider, V. S. Space, gravity and the physiology of aging: parallel or convergent disciplines? A mini-review. Gerontology 56, 157–166 (2010).
Honda, Y. et al. Genes down-regulated in spaceflight are involved in the control of longevity in Caenorhabditis elegans. Sci. Rep. 2, 487, https://doi.org/10.1038/srep00487 (2012).
Halberg, F. et al. Chronoastrobiology: proposal, nine conferences, heliogeomagnetics, transyears, near-weeks, near-decades, phylogenetic and ontogenetic memories. Biomed. Pharmacother. 58(Suppl 1), 150s–187s (2004).
Mitsutake, G. et al. Does exposure to an artificial ULF magnetic field affect blood pressure, heart rate variability and mood? Biomed. Pharmacother. 58(Suppl 1), 20s–27s (2004).
Stoupel, E. et al. Acute myocardial infarction (AMI) (n-11026) on days of zero geomagnetic activity (GMA) and the following week: Differences at months of maximal and minimal solar activity (SA) in solar cycles 23 and 24. J. Basic Clin. Physiol. Pharmacol. 23, 5–9 (2012).
Alabdulgader, A. et al. Long-term study of heart rate variability responses to changes in the solar and geomagnetic environment. Sci. Rep. 8, 2663, https://doi.org/10.1038/s41598-018-20932-x (2018).
Otsuka, K. et al. Geomagnetic disturbance associated with decrease in heart rate variability in a subarctic area. Biomed. Pharmacother. 55(Suppl 1), 51s–56s (2001).
Otsuka, K. et al. Alternating-light-darkness-influenced human electrocardiographic magnetoreception in association with geomagnetic pulsations. Biomed. Pharmacother. 55(Suppl 1), 63s–75s (2001).
Ohshima, H. Healthy life learned from space medicine. Auton. Nerv. Syst. (Tokyo) 54, 73–76 (2017).
Izumi, R., Ishioka, N., Mizuno, K. & Goka, T. Space environment, electromagnetic fields, and circadian rhythm. Biomed. Pharmacother. 55(Suppl 1), 25s–31s (2001).
Kamide, Y., Richmond, A. D. & Matsushita, S. Estimation of ionospheric electric field, ionospheric currents and field-aligned currents from ground magnetic records. J. Geophys. Res. 86, 801–813 (1981).
Kamide, Y. Estimate of electromagnetic quantities in space from ground magnetic records. Science 241, 328 (1988).
Kamide, Y., Shue, J.-H., Hausman, B. A. & Freeman, J. W. Toward real-time mapping of ionospheric electric fields and currents. Adv. Space Res. 26, 213–222 (2000).
Kamide, Y. & Balan, N. The importance of ground magnetic data in specifying the state of magnetosphere–ionosphere coupling: a personal view. Geosci. Lett. 3, 10, https://doi.org/10.1186/s40562-016-0042-7 (2016).
Engebretson, M. & Zesta, E. The future of ground magnetometer arrays in support of space weather monitoring and research. Space Weather 15, 1433–1441 (2017).
Task, F. of the European Society of Cardiology and the North American Society of Pacing and Electrophysiology Heart rate variability: standards of measurement, physiological interpretation, and clinical use. Circulation 93, 1043–1065 (1996).
Saito, K., Koyama, A., Yoneyama, K., Sawada, Y. & Ohtomo, N. ed., A recent advances in time series analysis by maximum entropy method. Hokkaido University Press (Sapporo, 1994).
Baria, A. T., Baliki, M. N., Parrish, T. & Apkarian, A. V. Anatomical and functional assemblies of brain BOLD oscillations. J. Neurosci. 31, 7910–7919 (2011).
Brenner, S. Genetics of Caenorhabditis elegans. Genetics 77, 71–94 (1974).
Adenle, A. A., Johnsen, B. & Szewczyk, N. J. Review of the results from the International C. elegans first experiment (ICE-FIRST). Adv. Space Res. 44, 210–216 (2009).
Sasagawa, Y. et al. Effects of gravity on early embryogenesis in Caenorhabditis elegans. Biol. Sci. Space 17, 217–218 (2003).
Higashitani, A. et al. Checkpoint and physiological apoptosis in germ cells proceeds normally in spaceflown Caenorhabditis elegans. Apoptosis 10, 949–954 (2005).
Higashibata, A. et al. Decreased expression of myogenic transcriptional factors and myosin heavy chains in Caenorhabditis elegans muscles developed during spaceflight. J. Exp. Biol. 209, 3209–3218 (2006).
Adachi, R. et al. Spaceflight results in increase of thick filament but bot thin filament proteins in the paramyosin mutant of Caenorhabditis elegans. Adv. Space Res. 41, 816–823 (2008).
Zhao, L., Rui, Q. & Wang, D.-Y. Molecular basis for oxidative stress induced by simulated microgravity in nematode Caenorhabditis elegans. Sci. Total Environ. 607–608, 1381–1390 (2017).
Li, W., Wang, S. & Wang, D. Regulation of the response of Caenorhabditis elegans to simulated microgravity by p38 mitogen-activated protein kinase signaling. Sci. Rep. 8, 857, https://doi.org/10.1038/s41598-018-19377-z (2018).
Garrett-Bakelman, F. E. et al. The NASA Twins Study: A multidimensional analysis of a year-long human spaceflight. Science 364, https://doi.org/10.1126/science.aau8650 (2019).
Ernst, G. Heart-Rate Variability-More than Heart Beats? Front. Public Health 5, 240, https://doi.org/10.3389/fpubh.2017.00240 (2017).
Wolf, M. M., Varigos, G. A., Hunt, D. & Sloman, J. G. Sinus arrhythmia in acute myocardial infarction. Med. J. Aust. 2, 52–53 (1978).
Bigger, J. T. Jr. et al. Frequency domain measures of heart period variability and mortality after myocardial infarction. Circulation 85, 164–171 (1992).
Tsuji, H. et al. Impact of reduced heart rate variability on risk for cardiac events. The Framingham Heart Study. Circulation 94, 2850–2855 (1996).
Liao, D. et al. Cardiac autonomic function and incident coronary heart disease: a population-based case-cohort study. The ARIC Study. Atherosclerosis Risk in Communities Study. Am. J. Epidemiol. 145, 696–706 (1997).
Thayer, J. F., Ahs, F., Fredrikson, M., Sollers, J. J. III. & Wager, T. D. A meta-analysis of heart rate variability and neuroimaging studies: implications for heart rate variability as a marker of stress and health. Neurosci. Biobehav. Rev. 36, 747–756 (2012).
Allen, B., Jennings, J. R., Gianaros, P. J., Thayer, J. F. & Manuck, S. B. Resting high-frequency heart rate variability is related to resting brain perfusion. Psychophysiology 52, 277–287 (2015).
Jennings, J. R., Sheu, L. K., Kuan, D. C., Manuck, S. B. & Gianaros, P. J. Resting state connectivity of the medial prefrontal cortex covaries with individual differences in high-frequency heart rate variability. Psychophysiology 53, 444–454 (2016).
Winkelmann, T. et al. Structural brain correlates of heart rate variability in a healthy young adult population. Brain Struct. Funct. 222, 1061–1068 (2017).
Zulfiqar, U., Jurivich, D.A., Gao, W. & Singer, D.H. Relation of high heart rate variability to healthy longevity. Am. J. Cardiol. 105, 1181–1185 (2010). Erratum in: Am. J. Cardiol. 106, 142 (2010).
Nicolini, P., Ciulla, M. M., De Asmundis, C., Magrini, F. & Brugada, P. The prognostic value of heart rate variability in the elderly, changing the perspective: from sympathovagal balance to chaos theory. Pacing Clin. Electrophysiol. 35, 622–638 (2012).
Guo, Y. et al. Prognostic value of heart rate variability in patients with cancer. J. Clin. Neurophysiol. 32, 516–520 (2015).
Grippo, A. J. Opinion: “Heart rate variability, health and well-being: A systems perspective” research topic. Front. Public Health 5, 246, https://doi.org/10.3389/fpubh.2017.00246 (2017).
Shah, A. J. et al. Is heart rate variability related to memory performance in middle-aged men? Psychosom. Med. 73, 475–482 (2011).
Zeki Al Hazzouri, A., Haan, M. N., Deng, Y., Neuhaus, J. & Yaffe, K. Reduced heart rate variability is associated with worse cognitive performance in elderly Mexican Americans. Hypertension 63, 181–187 (2014).
Zeki Al Hazzouri, A., Elfassy, T., Carnethon, M. R., Lloyd-Jones, D. M. & Yaffe, K. Heart rate variability and cognitive function in middle-age adults: The coronary artery risk development in young adults. Am. J. Hypertens. 31, 27–34 (2017).
Elias, M. F. & Torres, R. V. The renaissance of heart rate variability as a predictor of cognitive functioning. Am. J. Hypertens. 31, 21–23 (2018).
Arab, C. et al. Heart rate variability measure in breast cancer patients and survivors: a systematic review. Psychoneuroendocrinology 68, 57–68 (2016).
Zhou, X. et al. Heart rate variability in the prediction of survival in patients with cancer: a systematic review and meta-analysis. J. Psychosom. Res. 89, 20–25 (2016).
Kloter, E., Barrueto, K., Klein, S. D., Scholkmann, F. & Wolf, U. Heart rate variability as a prognostic factor for cancer survival - A systematic review. Front. Physiol. 9, 623, https://doi.org/10.3389/fphys.2018.00623 (2018).
Lampert, R. et al. Decreased heart rate variability is associated with higher levels of inflammation in middle-aged men. Am. Heart J. 156(759), e1–e7 (2008).
Carney, R. M. et al. Heart rate variability and markers of inflammation and coagulation in depressed patients with coronary heart disease. J. Psychosom. Res. 62, 463–467 (2007).
Akselrod, S. et al. Power spectrum analysis of heart rate fluctuation: a quantitative probe of beat-to-beat cardiovascular control. Science 213, 220–222 (1981).
Togo, F., Kiyono, K., Struzik, Z. R. & Yamamoto, Y. Unique very low-frequency heart rate variability during deep sleep in humans. IEEE Trans. Biomed. Eng. 53, 28–34 (2006).
Kember, G. C., Fenton, G. A., Armour, J. A. & Kalyaniwalla, N. Competition model for aperiodic stochastic resonance in a Fitzhugh-Nagumo model of cardiac sensory neurons. Phys. Rev. E 63, 041911, https://doi.org/10.1103/PhysRevE.63.041911 (2001).
Kember, G. C., Fenton, G. A., Collier, K. & Armour, J. A. Aperiodic stochastic resonance in a hysteretic population of cardiac neurons. Phys. Rev. E 61, 1816–1824 (2000).
Berntson, G. G. et al. Heart rate variability: origins, methods, and interpretive caveats. Psychophysiology 34, 623–648 (1997).
Cornelissen, G. et al. Chronomics, time structures, for chronobioengineering for ‘a full life’. Biomed. Instrum. Technol. 33, 152–187 (1999).
Oinuma, S. et al. Graded response of heart rate variability, associated with an alteration of geomagnetic activity in a subarctic area. Biomed. Pharmacother. 56(Suppl 2), 284s–288s (2002).
Otsuka, K. et al. Chronomics for chronoastrobiology with immediate spin-offs for life quality and longevity. Biomed. Pharmacother. 57(Suppl 1), 1s–18s (2003).
Phillips, J. B. & Borland, S. C. Behaviour evidence for use of a light-dependent magnetoreception mechanism by a vertebrate. Nature 359, 142–144 (1992).
Kume, K. et al. mCRY1 and mCRY2 are essential components of the negative limb of the circadian clock feedback loop. Cell 98, 193–205 (1999).
Griffin, E. A., Staknis, D. & Weitz, C. J. Light-independent role of CRY1 and CRY2 in the mammalian circadian clock. Science 286, 768–771 (1999).
Buhr, E. D. & Takahashi, J. S. Molecular components of the mammalian circadian clock. Handb. Exp. Pharmacol. 217, 3–27 (2013).
Gegear, R. J., Casselman, A., Waddell, S. & Reppert, S. M. Cryptochrome mediates light-dependent magnetosensitivity in Drosophila. Nature 454, 1014–1018 (2008).
Yoshii, U., Ahmad, M. & Helfrich-Forster, C. Cryptochrome mediates light-dependent magnetosensitivity of Drosophila’s circadian clock. PLoS Biol. 7, 813–819 (2009).
Gegear, R. J., Foley, L. E., Casselman, A. & Reppert, S. M. Animal cryptochromes mediate magnetoreception by an unconventional photochemical mechanism. Nature 463, 804–807 (2010).
Thoss, F., Bartsch, B., Fritzsche, B., Tellschaft, D. & Thoss, M. J. The magnetic field sensitivity of the human visual system shows resonance and compass characteristic. Comp. Physiol. A 186, 1007–1010 (2000).
Thoss, F., Bartsch, B., Tellschaft, D. & Thoss, D. The light sensitivity of human visual system depends on the direction of view. J. Comp. Physiol. A 188, 235–237 (2002).
Nießner, C. et al. Cryptochrome 1 in retinal cone photoreceptors suggests a novel functional role in mammals. Sci. Rep. 6, 21848, https://doi.org/10.1038/srep21848 (2016).
Wang, C. X. et al. Transduction of the geomagnetic field as evidenced from alpha-band activity in the human brain. eNeuro, https://doi.org/10.1523/ENEURO.0483-18.2019 (2019).
Foley, L. E., Gegear, R. J. & Reppert, S. M. Human cryptochrome exhibits light-dependent magnetosensitivity. Nat. Commun. 2, 356, https://doi.org/10.1038/ncomms1364 (2011).
Rounis, E. et al. Frequency specific changes in regional cerebral blood flow and motor system connectivity following rTMS to the primary motor cortex. Neuroimage. 26, 164–176 (2005).
Hampson, M. & Hoffman, R. E. Transcranial magnetic stimulation and connectivity mapping: Tools for studying the neural bases of brain disorders. Fron. Sys. Neur. 4, 1–8 (2010).
Eldaief, M. C., Halko, M. A., Buckner, R. L. & Pascual-Leone, A. Transcranial magnetic stimulation modulates the brain’s intrinsic activity in a frequency-dependent manner. Proc. Natl. Acad. Sci. USA 108, 21229–21234 (2011).
Liston, C. et al. Default mode network mechanisms of transcranial magnetic stimulation in depression. Biol. Psychiatry 76, 517–526 (2014).
Vernikos, J. & Hosie, R. The g-connection: harness gravity and reverse aging. iUniverse Inc, pp 294 (New York, 2004).
Acknowledgements
The authors thank Dr. I. Tayama, T. Aiba and S. Ishida from the Space Biomedical Research Group, Japan Aerospace Exploration Agency (JAXA), for cooperation in our study. The authors also acknowledge the cooperation of the astronauts, the engineers, staff and managers of JAXA and NASA. The help of Larry A. Beaty to improve the English language for greater clarity and readability is greatly appreciated. JAXA Chronobiology Project was supported by the Japan Aerospace Exploration Agency (K.M., H.O., S.F., C.M.) and Halberg Chronobiology Fund (G.C.).
The first two authors wish to dedicate this paper to the memory of Earl E Bakken and Franz Halberg, who early on envisioned brain-heart interactions and the importance of chronobiology for its investigation. In 1987, they wrote, “The chronobiologic component of blood pressure reflects but one end-result of the body’s coordination systems that link all the chronobiologic oscillators throughout the body into one harmonious symphony. These rhythms occur at all levels from the single cell to the whole organism. The synchronization of these oscillators is done via the brain in man and in influenced by the social regimen. Our getting up and going to bed at the times set by society once a day reset our free-running internal rhythm setters into step with world time. The brain (or mind) is therefore a vital link in the time region’s coordination of the heart, cardiovascular system and whole person”.
Author information
Authors and Affiliations
Contributions
K.O. and G.C. wrote the first draft of manuscript and prepared the figures. K.O., H.O. and C.M. designed the study, and K.M. and S.F. contributed to the acquisition of data. K.O., G.C. and Y.K. analyzed the data, and K.O., G.C., Y.K. and K.S. contributed to the writing and editing of the manuscript. All authors read and contributed to the final version of the manuscripts.
Corresponding author
Ethics declarations
Competing Interests
The authors declare no competing interests.
Additional information
Publisher’s note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Otsuka, K., Cornelissen, G., Kubo, Y. et al. Anti-aging effects of long-term space missions, estimated by heart rate variability. Sci Rep 9, 8995 (2019). https://doi.org/10.1038/s41598-019-45387-6
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-019-45387-6
This article is cited by
-
Advancing insights into microgravity induced muscle changes using Caenorhabditis elegans as a model organism
npj Microgravity (2024)
-
Methods for assessing change in brain plasticity at night and psychological resilience during daytime between repeated long-duration space missions
Scientific Reports (2023)
-
Prolonged microgravity induces reversible and persistent changes on human cerebral connectivity
Communications Biology (2023)
-
Comprehensive Analysis of Cellular Senescence-Related Genes in Prognosis, Molecular Characterization and Immunotherapy of Hepatocellular Carcinoma
Biological Procedures Online (2022)
-
Unconscious mind activates central cardiovascular network and promotes adaptation to microgravity possibly anti-aging during 1-year-long spaceflight
Scientific Reports (2022)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
