
Abstract
The dive response is well documented for marine mammals, and includes a significant reduction in heart rate (fH) during submersion as compared while breathing at the surface. In the current study we assessed the influence of the Respiratory Sinus Arrhythmia (RSA) while estimating the resting fH while breathing. Using transthoracic echocardiography we measured fH, and stroke volume (SV) during voluntary surface apneas at rest up to 255 s, and during recovery from apnea in 11 adult bottlenose dolphins (Tursiops truncatus, 9 males and 2 females, body mass range: 140–235 kg). The dolphins exhibited a significant post-respiratory tachycardia and increased SV. Therefore, only data after this RSA had stabilized were used for analysis and comparison. The average (±s.d.) fH, SV, and cardiac output (CO) after spontaneous breaths while resting at the surface were 44 ± 6 beats min−1, 179 ± 31 ml, and 7909 ± 1814 l min−1, respectively. During the apnea the fH, SV, and CO decreased proportionally with the breath-hold duration, and after 255 s they, respectively, had decreased by an average of 18%, 1–21%, and 12–37%. During recovery, the fH, SV, and CO rapidly increased by as much as 117%, 34%, and 190%, respectively. Next, fH, SV and CO rapidly decreased to resting values between 90–110 s following the surface apnea. These data highlight the necessity to define how the resting fH is estimated at the surface, and separating it from the RSA associated with each breath to evaluate the significance of cardiorespiratory matching during diving.
Similar content being viewed by others

Introduction
The cardiovascular responses in marine mammals were initially studied during forced dives by Per Scholander1 who concluded that during a breath-hold there is a decrease in heart-rate (fH), and presumably a decrease in cardiac output (CO), and an increase in peripheral resistance (vasoconstriction)2. Scholander later coined these cardiovascular responses the “master switch of life” and proposed that this was a reflex that was tightly linked to diving2. Following this initial work, numerous studies have measured fH during restrained, semi-restrained, and voluntary diving and swimming in a range of breath-hold diving vertebrates1,3,4,5,6,7,8,9,10,11,12,13,14,15,16,17. It has been concluded that there are marked differences in cardiac responses in forced vs. voluntary/natural diving18. However, the minimum fH during diving in free ranging pinnipeds is similar to those during forced diving experiments, and there was no clear correlation between diving capacity and the level of bradycardia19. Based on these differences some have argued that the dive response is not a reflex adjustment20, but a more generalized response that is not a specific trait for diving19. In addition, some studies have provided evidence that diving mammals possess cognitive control over cardiac function16,21,22,23, which may allow management of gases during diving by altering the ventilation-perfusion matching24.
Still, it is universally agreed that diving vertebrates experience a diving related bradycardia while submerged20,25,26. However, there is disagreement whether there are changes in stroke volume (SV) during diving5,9,12,13,15,22,27,28,29,30, and to our knowledge, no study has assessed SV in diving cetaceans. Thus, current estimates of CO in cetaceans, and how they may be modified during diving, are based on estimated values from pinnipeds and terrestrial species. These direct extrapolations could significantly bias our understanding of the eco-physiology of cetaceans and the results of gas dynamics modeling regarding diving performance31,32,33,34.
In cetaceans, most studies have largely been focused on cardiac frequencies7,16,17,21,35, and no published data exists on cardiac contractile responses during forced or voluntary dives. Despite anatomical difficulties obtaining images of the heart, a non-invasive study on cardiac performance using transthoracic echocardiography have previously been published in bottlenose dolphins (Tursiops truncatus) before and after high intensity exercise36. In cetaceans, the respiratory sinus arrhythmia (RSA) possibly results in a biased average resting fH depending on the breathing frequency (fR). For example, in the bottlenose dolphin the fH immediately following a spontaneous breath during rest may be as high as 80–100 beats · min−1, but rapidly declines to a stable values around 40–50 beats · min−1 in about 8–20 s following the breath16,17,36,37. The respiratory phase in cetaceans begins with a rapid expiration, followed by an inspiration and a respiratory pause that can last for up to a minute38. Consequently, if the resting fH at the surface is estimated over a pre-determined time interval, the RSA may significantly alter the value, and it will vary with the fR and the duration of the time interval chosen.
Most studies evaluating the cardiac responses during diving and/or exercise in diving mammals have included the fH changes caused by the RSA to estimate the resting fH at the surface7,10,16,17,21,35,39. It has long been recognized that stress significantly affects physiological responses, and researchers have indicated that cardiac responses during forced dives likely affects any extrapolations to normal cardiac function1,16,40. Similar to the effect stress have on fH, the confounding influence of respiration should be separated from the fH before conclusions can be made about the cardiovascular changes during a breath-hold1,20. Consequently, conclusions from past studies on either forced or freely diving marine mammals may be confounded by the RSA. For this reason, Miedler et al.36 proposed that the fH should be evaluated once the instantaneous fH (ifH) had stabilized, maintaining a more or less constant level during at least 5–7 s, which occurs approximately 8–20 s after the breath (see Figs 1 and 2 in37, and see Supplementary Material).
The aim of this study was to provide estimates of cardiac function in the bottlenose dolphin before, during, and following a voluntary surface apnea at rest. These data provide the first voluntary, non-invasive, and semi-continuous measurements of fH, SV and CO before, during, and after a surface apnea of up to 255 s using transthoracic ultrasound in 11 adult bottlenose dolphins. The species-specific data provided in this study clearly show that the magnitude and importance of the cardiac responses during voluntary diving may have been significantly overestimated in previous studies, and show that the effect of RSA may have biased the results from previous studies. In addition, we show that both fH and SV decrease temporally during the apneic period, and show that cardiovascular recovery times are between 70–100 s following voluntary surface apneas at rest. We conclude that the results presented in this study will enhance our ability to more accurately understand the physiological limitations and gas exchange dynamics in cetacean diving physiology, and provide methods to more readily compare results within and between different studies and species.
Results
Heart rate, stroke volume and cardiac output during rest
During rest, the average resting instantaneous fH (ifH, 44 ± 6 beats min−1, range: 27–67 beats min−1, n = 13, P > 0.1), SV (iSV, 179 ± 31 ml, P > 0.3, n = 11), and CO (iCO, 7909 ± 1814 ml min−1, P > 0.9) were not significantly affected by Mb (Table 1). The average mass-specific fH (sfH), SV (sSV), and CO (sCO) were 0.26 ± 0.04 beats min−1 kg−1, 1.06 ± 0.24 ml kg−1, and 47 ± 11 ml min−1 kg−1, respectively.
Heart rate, stroke volume and cardiac output during surface apnea
Of the 13 dolphins that participated in the study, the diameter of the aortic valve orifice could be measured in 11 animals (Table 1). These 11 dolphins participated in a total of 173 voluntary surface apneas at rest, ranging in duration from 20 s to 255 s (Table 2). For ifH, the best model to describe the effect of surface apnea included the maximal duration of the dive, and the time during the dive (Fig. 1A, Table 3). The best models to describe the effect of apnea on iSV included Mb and apnea duration (Fig. 1B, Table 3). For iCO the best model included Mb, time during the dive, and the maximal duration of each breath-hold (Fig. 1C, Table 3).

Instantaneous (A) heart rate (ifH, n = 13), (B) stroke volume (iSV, n = 11), and (C) cardiac output (iCO, n = 11) versus time during dive (s) in bottlenose dolphin. Solid and dotted red lines are the resting average and 95% confidence limits, respectively, while breathing after correcting for respiratory sinus arrhythmia. Gray solid and dotted lines are model predicted regressions for (A) maximal dive duration of 30 s or 300 s (B) and (C) for a 150 kg and 230 kg dolphin (for a 120 s maximal dive duration in (C), and cyan solid and dotted lines are model predictions for a maximal dive duration of 30 s or 300 s for a 200 kg animal. Note for fH, panel A, the model did not warrant inclusion of Mb. The legend in panel A give the symbol for each individual animal in panels A–C.

Instantaneous (A) heart rate (ifH, n = 13), (B) stroke volume (iSV, n = 11), and (C) cardiac output (iCO, n = 11) versus duration following a voluntary surface apnea (s) at rest in bottlenose dolphin. Solid and dotted red lines are the resting average and 95% confidence limits, respectively while breathing after correcting for respiratory sinus arrhythmia. Gray solid line is model predicted regressions. The legend in panel A give the symbol for each individual animal in panels A–C.
Heart rate, stroke volume and cardiac output during recovery
The average (n = 10) ifH, iSV, and iCO after the 10 first seconds following the first breath from a voluntary surface apnea at rest was 78 ± 8 beats · min−1 (t-test compared to pre-dive rest, t-value = 11.1, df = 21 P < 0.01), 225 ± 41 ml (t-test compared to pre-dive rest, t-value = 2.9, df = 19 P < 0.01), and 17612 ± 3047 ml · min−1 (t-test compared to pre-dive rest, t-value = 9.0, df = 19 P < 0.01), respectively. For ifH (Fig. 2A), iSV (Fig. 2B), and iCO (Fig. 2C, Table 3) the best models to describe the recovery from surface apnea included recovery time. Neither the duration of the surface apnea, nor Mb warranted inclusion in the model (P > 0.3 for all).
Discussion
Given the importance to understand the effects of O2 delivery during exercise and submersion, determining the cardiovascular responses are important to understand physiological function in cetaceans. The objective with the current study was to evaluate ifH, iSV and iCO before, during and following voluntary surface apneas at rest to help improve our understanding of cardiorespiratory responses associated with diving. Similar to previous studies, we report a significant RSA10,16,17,36,37, which alters how the resting fH is evaluated and thereby the magnitude of the apparent dive response. As RSA is known to be affected by both tidal volume (VT) and fR in humans and dolphins37,41, we propose that studies that are aimed at evaluating the cardiovascular responses associated with diving and exercise need to evaluate the effect of RSA when estimating the surface fH that is made as a comparison. For example, if ifH increases to approximately 120 beats · min−1 immediately following a dive, what proportion of this increase is merely due to elevated fR? To answer this, the cardiorespiratory coupling should be determined for each species to accurately determine the effect of RSA (see Fig. 1 in37, and Supplemental Material).
The cardiovascular changes associated with diving and exercise while submerged have long fascinated eco-physiologists trying to understand how marine vertebrates manage gases during diving. Most initial studies were performed in a laboratory setting during forced submersions, but the development of bio -logging tools, or use of medical technology have opened up new avenues for research and understanding the cardiorespiratory changes while submerged under natural conditions, or in studies on animals under voluntary control7,10,17,21,35,37,39,42. The changes in fH associated with diving have been studied intensively in numerous species26. Only a few studies have measured CO in live marine mammals, and most of these studies have been made in pinnipeds5,9,15,22,27,28,29,30. Of these, only three have been performed in unrestrained and/or free swimming animals9,15,22. In the sea lion, SV and fH were not reported15, but in the seal, CO and fH increased while SV decreased as the animal was swimming at the surface9. A similar increase in CO was observed during exercise in the submerged harbor seal, while SV also increased slightly9. We are only aware of one study that has measured SV in cetaceans either at rest or following high intensity exercise36. In a study looking at the development of the dive response in bottlenose dolphins, it was reported that there were no differences with age in resting fH at the surface, but the fH during submersions was reduced to lower values in older animals7. In the harbor porpoise, the fH before and during diving was altered depending on the anticipated dive duration, suggestive of cognitive control of fH21. In exercising dolphins there was a linear increase in fH with increasing metabolic rate10. A similar complex relationship was observed in freely swimming and exercising harbor porpoises where the degree of diving bradycardia was altered by the duration and activity level of the submersion35. An important detail in the current study is whether the post-breath period that cause variation in fH is included in calculating the resting fH as this response is pronounced in this taxonomic group and depends on the fR37.
In our past work36,37, and based on the result in the current study, we propose that for better interpretation of the magnitude of the cardiac adjustment during a breath-hold, the comparison should be made with surface values that separate the confounding influence of the RSA. There could be different ways to account for this, and one would be to allow for a long enough resting period that assured that the animals fR was normal, or at least report the fR so that comparisons could be made between studies. However, as that would still involve confounding the autonomic cardiac response with the voluntary process of breathing, an alternative would be to measure the resting heart rate with minimal influence of the RSA. Thus, we propose that it can be done by measuring the ifH as we have done in the current study, or continuous ECG measurements that allows the changes in ifH to be modeled and the stabilized fH predicted37. We propose to call this value the RSA-corrected resting fH, to differentiate this with the resting fH reported in most studies.
The average RSA-corrected resting fH in the current study was not different from those reported in past studies using the same individual dolphins and the same methodology, or continuously measuring the ECG (fH = 41 ± 9 beats min−1 n = 13, 2-tailed t-test, P = 0.33, t-value = 1.0, df = 24)36,37, but was considerably lower and less variable as compared with resting fH’s reported in previous studies in the bottlenose dolphin (ranging from 60–105 beats min−1)7,17,39. The RSA-corrected resting fH, SV and CO, where estimated to exclude the changes in fH, SV and CO associated with respiration36,37. As both fH and SV increase following a breath, our average readings were lower as compared with an average obtained if the fH was estimated over the whole inter-breath cycle, which included the RSA (see Fig. 1 in37, or Supplemental Material). Conversely, estimating resting fH over a determined period of time includes the variation associated with the RSA, and the average will therefore depend on the duration of the measurement and the fR within this period.
Thus, the higher fH reported in previous studies may be confounded through inclusion of RSA, and without reporting the fR a comparison is difficult. Consequently, estimating resting fH without accounting for the RSA result in a higher resting value, and if used to assess the dive response it will result in higher values than when using the RSA-corrected resting fH. For example, in one study in the bottlenose dolphin, the estimated resting fH while at the surface was 105 ± 8 beats min−1, and decreased to 40 ± 6 beats min−1, while submerged at 15 m for an average dive duration of 85 ± 51 s (range 14–160 s)39. The RSA-corrected resting fH during a voluntary surface apnea up to 160 s in the current study was similar to the resting diving fH reported in the past study39, and similar to the resting fH for spontaneous breathing at the surface after accounting for the RSA (27 to 67 beats min−1, see refs 36,37). Thus, our results suggest that the changes in fH associated with diving, after accounting for RSA, are less pronounced than previously reported, and only become significantly lower than resting surface values during apneas exceeding 2.5–3 min (Fig. 1A). Thus, the results presented here provide an interesting perspective that when accounting for the RSA dolphins do not exhibit the pronounced diving bradycardia associated with the stress during forced dives or those that include the RSA when estimating the resting fH, except during extended apneas. Whether this is a universal trait in other marine mammals, or how exercise or depth influence these changes remains to be determined once past and future studies evaluate the RSA-corrected resting fH in voluntary diving animals. Interestingly, similar results were found in elephant seals where the RSA minimum and fH during apnea appeared similar43. In addition, the RSA developed with age, and its magnitude correlated with dive duration43. Thus, the RSA may be an important physiological index that is more correlated with diving ability in addition to cognitive ability to alter fH and SV to accurately match ventilation and perfusion to improve gas exchange24.
Both resting sSV, and sCO were significantly higher as compared with a previous study assessing cardiovascular changes following a bout of exercise (sSV = 0.79 ± 0.14 ml · kg−1, 2-tailed t-test, P < 0.01, t-value = 3.4, df = 22; sCO = 32 ± 9 ml · min−1 · kg−1, 2-tailed t-test, P < 0.01, t-value = 3.6, df = 22)36. However, both sSV and sCO were considerably lower as compared with those measured at rest in the harbor seal (Phoca vitulina, sSv = 1.8–3.1 mL · kg−1, sCO = 102–394 mL · min−1 · kg−1)9, and California sea lion (Zalophus californianus, sSv = 2.0 mL · kg−1, sCO = 150–180 mL · min−1 · kg−1)15 using thermodilution techniques. We believe that these differences between species could in part be explained by the lower fR, breath duration, and higher flow-rates, and VT’s in the cetaceans as compared with the pinnipeds38,44. Thus, if respiration significantly alters cardiac function (fH and SV), it is vital that these variables are compared during periods between breaths to minimize the impact of respiration on cardiac function. By standardizing these measurements to prevent the confounding effect of RSA, it would allow inter- and intra-species comparisons by reducing the variability caused by the RSA.
It has been suggested that the extreme changes in fH associated with forced submergence, when an animal does not know or has no control over the duration of the apnea, reflect an animal that prepares for a maximal asphyxic challenge20. For this reason, Blix20 proposed that studies that assess the dive response should clearly distinguish between forced and voluntary diving. An additional issue that may significantly alter variation in fH, especially in studies on trained animals, is the suggestion that marine mammals have cognitive control of fH20,21,22,23. As repeated measurements were performed for each trial, the time during the dive allowed us to investigate temporal changes during a breath-hold. The maximal dive duration was the total duration of the apnea, which allowed us to assess whether the initial cardiovascular changes were different for a long or short breath-hold, possibly indicating a learning effect or cognitive ability to alter fH and SV. Our results showed a greater initial drop in fH during longer dives (Fig. 1A), which provide additional support of voluntary control of the diving related changes in fH20,21,22,23. Previously, it has been proposed that by fine tuning the alveolar ventilation (\({\dot{V}}_{A}\)) and lung perfusion, marine vertebrates are able to selectively exchange O2 and CO2 during diving, while minimizing N2 exchange24. This mechanism relies in part on voluntary control of pulmonary and systemic perfusion, and the ability to selectively perfuse collapsed regions of the lung24. Disruption of such refined control of pulmonary blood flow could have severe consequences in gas management, and may explain how species that are normally able to avoid diving related problems experience gas bubble disease when exposed to stressful situations while diving32,45,46,47,48,49,50. While our study does not provide direct evidence of voluntary control of perfusion, we show indirect evidence that the dolphins alter fH and SV depending on the length of the dive, similar to the results in the harbor porpoise21. As proposed by Mottishaw et al.19, the cardiovascular changes during diving may be a complex physiological response altered by a number of factors such as voluntary/anticipatory adjustment, submersion, exercise, stress, and fear. Thus, the cardiovascular responses during diving may be a much more plastic physiological trait rather than a purely autonomic response, or potentially merely an extension of the RSA.
Following a voluntary surface apnea up to 255 s, fH, SV, and CO increased by 78% (maximal value: 164%), 26% (maximal value: 73%), and 123% (maximal value: 229%), respectively (Figs 1 vs. 2). Similar changes in fH at maximal exercise are difficult to assess as they generally do not account for the RSA10,51. However, maximal fH, SV, and CO 10 s following a single bout of high intensity exercise increased by as much as 307%, 294%, and 727%, respectively36. Consequently, the cardiovascular changes during recovery from a voluntary surface apnea appear less extreme than measured following a high intensity bout of exercise. There are several possible reasons for the more moderate cardiovascular changes during recovery in the current study. First, the dolphins did not perform high intensity exercise and therefore did not have elevated metabolic rate. The elevated fH, SV and CO following the breath-hold in the current study was only used to replenish the O2 stores and remove any CO2 produced caused by the surface apnea. During exercise, the dolphins incur an O2 debt that may be exacerbated by the elevated rate of O2 consumption. This may require greater cardiovascular recruitment to rapidly repay the greater O2 debt following exercise and may extend recovery as compared with voluntary apnea at rest. Increased aerobic work and greater O2 debt may explain why ifH, iSV and iCO took at least 4 min to recover following a high intensity bout of exercise36, while in the current study these variables had returned to baseline after approximately 90–110 s (Fig. 2A–C). This agrees with a previous study showing that the O2 stores had recovered after approximately 1.2 min following a surface apnea of up to 5 min52. Consequently, for an actively diving dolphin we would expect the recovery fH, SV and CO to reach higher values and/or increase for a longer duration as higher activity would increase the O2 debt. In grey seals, the surface interval increased with dive duration for short dives (<7 min) but not for long dives53. In extreme divers, such as Weddell and elephant seals there does not appear to be a clear relationship between dive duration, even those exceeding the aerobic dive limit, and surface interval duration54,55. One reason could be that seals partially recover while diving55. However, the surface fH increased with dive duration in the elephant seal54, but there was no clear relationship in the grey seal53. Similarly, the recovery fH during single dives correlated with dive duration and activity in the Steller sea lion14. Thus, these data provide evidence for excellent control of cardiovascular function to rapidly and efficiently manage metabolic gases.
In summary, this study provides the first non-invasive measurements of fH and SV before, during, and after voluntary surface apneas at rest in cetaceans. To account for the RSA, we measured the RSA-corrected resting ifH and iSV only during inter-breath periods when fH and SV had stabilized following the breath. We therefore report lower average fH while resting at the surface, and propose that these results have lower variability and provide improved comparable values across individuals or species with varying respiratory effort. Further work should be done to define the significance of the RSA, its role in managing gas exchange, and the temporal changes to allow the RSA-corrected resting fH and SV to be defined within and between species. The ifH, iSV and iCO decreased slightly with breath-hold duration as compared with the RSA-corrected resting ifH. However, compared to past studies, the level of the diving bradycardia was considerably reduced. Following the apnea, the ifH, iSV and iCO immediately increased as much as 164%, 73%, and 229%, respectively even during the stabilization period, but rapidly returned to resting levels within 50–60 s after several respirations. Based on our results, we propose that future studies that assess the diving related changes in cardiac function have to do similar correction to avoid the confounding effect of RSA, and past studies may have to be re-evaluated to tease apart the contradictory effects of submergence and exercise. Our data show that the initial diving ifH is lower during longer breath-hold, which provides additional support that cetaceans have partial cognitive control over their diving bradycardia. This, provides additional support for the hypothesis that cetaceans can voluntarily alter gas exchange during diving.
Material and Methods
Animals
The study protocols were approved by the Animal Care and Welfare Committee of the Oceanogràfic Foundation (OCE-17-16 and amendment OCE-29-18), and all experiments were performed in accordance with relevant guidelines and regulations. Ultrasound continuous flow Doppler was used to measure fH and SV at the level of the left ventricular outflow tract (at the level of the aortic valve orifice), during and following a surface apnea (Table 1) from 10 adult male and 3 adult female Atlantic bottlenose dolphins, 4 to over 33 years old, housed at the Oceanogràfic-Valencia and Dolphin Quest-Oahu, between 2016 and 2018 (Table 1).
The animal ID, body mass, and age (known or estimated) at the time of the study are summarized in Table 1.
Experimental trials
All experiments were performed using operant conditioning. Participation by the dolphins was voluntary, and the animals were not restrained and could refuse to participate or withdraw at any point during the experimental trial. Each experiment (trial) consisted of an animal staying stationary in the water in left lateral recumbence with the blow-hole out of the water, allowing ultrasound probe placement to find the left ventricle. To evaluate the effect of a surface apnea, the animals were conditioned to hold their breaths voluntarily as much as possible up to a maximum of 255 s (Table 2). The dolphins were asked by the trainer to turn on their side with the blowhole submerged until they decided to end the breath-hold. The ultrasound examination continued throughout the apnea and recovery period to assess the cardiovascular changes during a breath-hold and recovery. For measurements of fH, and SV during apnea and recovery, the breath-hold and recovery durations were recorded. Out of the 13 animals, we were only able to measure the aortic diameter in 11 and SV and CO were therefore only estimated in 11 dolphins (Table 1). For that reason, only 11 animals participated in the voluntary breath-hold (Table 2).
Ultrasound data acquisition
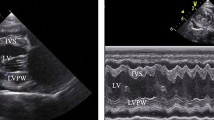
The ultrasound machine (Vivid-I, General Electric) with a 1–3 MHz phased array probe was used to obtain left ventricular ifH and iSV, as previously detailed36. CO was estimated as: iCO = ifH × iSV. The ifH was estimated from the flow traces, and the iSV calculated from the surface area of the aortic valve orifice multiplied by the velocity time integral of the systolic blood flow along the left ventricular outflow tract. The cross-sectional area of the aortic valve was calculated from the aortic valve orifice diameter as: surface area = radius2 × π. In order to obtain left ventricular systolic blood flow, the “apical” long axis view of the left ventricular outflow tract and the aortic root was used to place the continuous flow Doppler parallel to the blood flow in the left ventricle and through the middle of the aortic valve orifice. We measured the aortic valve orifice diameter at the level of valve insertion at the different cardiac phases to confirm that its diameter was circular and constant for all flow-rates.
Our pas studies and initial assessments indicated significant changes in both ifH and iSV following respiration36,37. Because of this significant respiratory sinus arrhythmia (RSA), at least 3 repeated measurements of fH and ventricular flow were made for each measurement and the average used as an estimate of the fH following a breath. To obtain resting data that were not affected by the RSA, we only analysed fH at least 12–20 s following a spontaneous breath and after the ifH had stabilized for 5–7 s following the breath (see Supplementary Material)36,37. At this point, the ifH and iSV were averaged for at least 3 R-R intervals for each measurement. These were then averaged for each individual. The ultrasound data were saved either as 8–16 s movies or 2-dimensional images and later analysed using manufacturer specific software (See S1 in Supplementary Material).
The duration to locate the aortic valve orifice differed between animals based on the size of the cardiac acoustic window, animal movement, and whether the animal was performing the surface apnea or breathing spontaneously. We standardized the recovery phase to begin immediately following the first breath, and cardiovascular measurements began as early as 10 s following this breath when the ifH again stabilized following the RSA.
Data assessment and statistical analysis
We separated the analysis into one of three conditions: pre-apnea, apnea, post-apnea. The relationship between a dependent variable (CO, SV and fH) and experimental covariates; body mass (Mb), time during dive, the total dive duration for a given dive (max duration), or time since end of apnea (recovery time), was analyzed using linear-mixed effects models (lme, R: A Language and Environment for Statistical Computing, R Foundation for Statistical Computing, version 3.3.3, 2016). We log10-transformed fH, SV, CO, time during dive, max dive duration, recovery time and Mb to generate linear functions that could be used with the lme function in R. The individual animal was treated as a random effect, which accounted for the correlation between repeated measurements on the same individual56. Initially, a univariate analysis on each independent variable was performed; only those variables with a P-value < 0.10 (Wald’s tests) were considered in a multivariate analysis. Best models of remaining variables were chosen by the log-likelihood (LL) ratio test, and the Akaike information criterion (AIC) and significant parameters assessed by the t-value between the estimate and its standard error. Acceptance of significance was set to the P < 0.05 level, while 0.05 < P < 0.1 was considered a trend. Data are presented as the mean ± standard deviation (s.d.), unless otherwise stated.
Data Availability
The data used in this study are freely available at the following link: osf.io/wdfmz.
References
Scholander, P. F. Experimental investigations on the respiratory function in diving mammals and birds. Hvalrådets Skrifter 22, 1–131 (1940).
Scholander, P. F. The Master Switch of Life. Sci. Am. 209, 92–106 (1963).
Andrews, R. D. et al. Heart rates of northern Elephant seals diving at sea and resting on the beach. J. Exp. Biol. 200, 2083–2095 (1997).
Berkson, H. Physiological adjustments to deep diving in the pacific green turtle (Chelonia mydas agassizii). Comp. Biochem. Physiol. 21, 507–524 (1967).
Blix, A. S. & Kjekshus, J. K. Cardiac output and its distribution through capillaries and A-V shunts in diving seals. Acta. Physiol. Scand. 118, 109–116 (1983).
Halsey, L. G. et al. How accurately can we estimate energetic costs in a marine top predator, the king penguin? Zool. 110, 81–92, https://doi.org/10.1016/j.zool.2006.09.001 (2007).
Noren, S. R., Cuccurullo, V. & Williams, T. M. The development of diving bradycardia in bottlenose dolphins (Tursiops truncatus). J. Comp. Physiol. B. 174, 139–147, https://doi.org/10.1007/S00360-003-0398-9 (2004).
Ponganis, P. J., Kooyman, G. L., Winter, L. M. & Starke, L. N. Heart rate and plasma lactate responses during submerged swimming and trained diving in California sea lions, Zalophus californianus. J. Comp. Physiol. B. 167, 9–16 (1997).
Ponganis, P. J., Kooyman, G. L., Zornow, M. H., Castellini, M. A. & Croll, D. A. Cardiac output and stroke volume in swimming harbor seals. J. Comp. Physiol. B. 160, 473–482 (1990).
Williams, T. M., Friedl, W. A. & Haun, J. E. The physiology of bottlenose dolphins (Tursiops truncatus): heart rate, metabolic rate and plasma lactate concentration during exercise. J. Exp. Biol. 179, 31–46 (1993).
Jones, D. R., Fisher, H. D., McTaggart, S. & West, N. H. Heart rate during breath-holding and diving in the unrestrained harbor seal. Can. J. Zool. 51, 671–680 (1973).
Jones, D. R. & Holeton, G. F. Cardiac output of ducks during diving. Comp. Biochem. Physiol. A. 41, 639–645 (1972).
Folkow, B., Nilsson, N. J. & Yonce, L. R. Effects of diving on cardic output in ducks. Acta. Physiol. Scand. 70, 347–361 (1967).
Hindle, A. G., Young, B. L., Rosen, D. A., Haulena, M. & Trites, A. W. Dive response differs between shallow- and deep-diving Steller sea lions (Eumetopias jubatus). J. Exp. Biol. Ecol. 394, 141–148 (2010).
Ponganis, P. J., Kooyman, G. L. & Zornow, M. H. Cardiac output in swimming California sea lions, Zalophus californianus. Physiol. Zool. 64, 1296–1306 (1991).
Elsner, R., Kenney, D. W. & Burgess, K. Diving Bradycardia in the Trained Dolphin. Nature 212, 407–408 (1966).
Houser, D. S., Dankiewicz-Talmadge, L. A., Stockard, T. K. & Ponganis, P. J. Investigation of the potential for vascular bubble formation in a repetitively diving dolphin. J. Exp. Biol. 213, 52–62, https://doi.org/10.1242/jeb.028365 (2010).
Kanwisher, J., Gabrielsen, G. & Kanwisher, N. Free and forced diving in birds. Sci. 211, 717–719, https://doi.org/10.1126/science.7192883 (1981).
Mottishaw, P. D., Thornton, S. J. & Hochachka, P. W. The diving response mechanism and its surprising evolutionary path in seals and sea lions. Am. Zool. 39, 434–450 (1999).
Blix, A. S. Diving Responses: Fact or Fiction. Physiology 2, 64–66, https://doi.org/10.1152/physiologyonline.1987.2.2.64 (1987).
Elmegaard, S. L., Johnson, M., Madsen, P. T. & McDonald, B. I. Cognitive control of heart rate in diving harbor porpoises. Curr. Biol. 26, R1167–R1176 (2016).
Elsner, R. W., Franklin, D. L. & Van Critters, R. L. Cardiac output during diving in an unrestrained sea lion. Nature 202, 809–810 (1964).
Ridgway, S. H., Carder, D. A. & Clark, W. Conditioned bardycardia in the sea lion Zalophus californianus. Nature 256, 37–38 (1975).
García-Párraga, D., Moore, M. & Fahlman, A. Pulmonary ventilation– perfusion mismatch: a novel hypothesis for how diving vertebrates may avoid the bends. Proc. Roy. Soc. B. 285, 20180482, https://doi.org/10.1098/rspb.2018.0482 (2018).
Butler, P. J. & Jones, D. R. Physiology of Diving Birds and Mammals. Physiol. Rev. 77, 837–899 (1997).
Ponganis, P. J. Diving Physiology of Marine Mammals and Seabirds. 333 (Cambridge University Press, 2015).
Zapol, W. M. et al. Regional blood flow during simulated diving in the conscioius weddell seal. J. Appl. Physiol. 47, 968–973 (1979).
Blix, A. S., Kjekshus, J. K., Enge, I. & Bergan, A. Myocardial blood flow in the diving seal. Acta. Physiol. Scand. 96, 277–280 (1976).
Murdaugh, H. V., Robin, E. D., Millen, J. E., Drewry, W. F. & Weiss, E. Adaptations to diving in the harbor seal: Cardiac output during diving. Am. J. Physiol. 210, 176–180 (1966).
Sinnett, E. E., Kooyman, G. L. & Wahrenbrock, E. A. Pulmonary circulation of the harbor seal. J. Appl. Physiol. 45, 718–727 (1978).
Fahlman, A., Jensen, F., Tyack, P. L. & Wells, R. Modeling tissue and blood gas kinetics in coastal and offshore common Bottlenose dolphins, Tursiops truncatus. Front. Physiol. 9, 1–13, https://doi.org/10.3389/fphys.2018.00838 (2018).
Fahlman, A., Tyack, P. L., Miller, P. J. & Kvadsheim, P. H. How man-made interference might cause gas bubble emboli in deep diving whales. Front. Physiol. 5, https://doi.org/10.3389/fphys.2014.00013 (2014).
Hooker, S. K., Baird, R. W. & Fahlman, A. Could beaked whales get the bends? Effect of diving behaviour and physiology on modelled gas exchange for three species: Ziphius cavirostris, Mesoplodon densirostris and Hyperoodon ampullatus. Resp. Physiol. Neurobiol. 167, 235–246 (2009).
Zimmer, W. M. X. & Tyack, P. L. Repetitive shallow dives pose decompression risk in deep-diving beaked whales. Mar. Mam. Sci. 23, 888–925 (2007).
McDonald, B. I., Johnson, M. & Madsen, P. T. Dive heart rate in harbour porpoises is influenced by exercise and expectations. J Exp Biol 221, jeb168740, https://doi.org/10.1242/jeb.168740 (2018).
Miedler, S., Fahlman, A., Valls Torres, M., Álvaro Álvarez, T. & Garcia-Parraga, D. Evaluating cardiac physiology through echocardiography in bottlenose dolphins: using stroke volume and cardiac output to estimate systolic left ventricular function during rest and following exercise. J. Exp. Biol. 218, 3604–3610, https://doi.org/10.1242/jeb.131532 (2015).
Cauture, F. et al. Using Respiratory Sinus Arrhythmia to Estimate Inspired Tidal Volume in the Bottlenose Dolphin (Tursiops truncatus). Front. Physiol. 10, https://doi.org/10.3389/fphys.2019.00128 (2019).
Fahlman, A., Moore, M. J. & Garcia-Parraga, D. Respiratory function and mechanics in pinnipeds and cetaceans. J. Exp. Biol. 220, 1761–1763, https://doi.org/10.1242/jeb.126870 (2017).
Noren, S. R., Kendall, T., Cuccurullo, V. & Williams, T. M. The dive response redefined: underwater behavior influences cardiac variability in freely diving dolphins. J. Exp. Biol. 215, 2735–2741, https://doi.org/10.1242/jeb.069583 (2012).
Irving, L., Scholander, P. F. & Grinnell, S. W. The respiration of the porpoise, Tursiops truncatus. J. Cell. Comp. Physiol. 17, 145–168 (1941).
Hirsch, J. A. & Bishop, B. Respiratory sinus arrhythmia in humans: how breathing pattern modulates heart rate. American Journal of Physiology - Heart and Circulatory Physiology 241, H620–H629 (1981).
Williams, T. M., Blackwell, S. B., Richter, B., Sinding, M.-H. S. & Heide-Jørgensen, M. P. Paradoxical escape responses by narwhals (Monodon monoceros). Sci. 358, 1328–1331, https://doi.org/10.1126/science.aao2740 (2017).
Castellini, M. A., Rea, L. D., Sanders, J. L., Castellini, J. M. & Zenteno-Savin, T. Developmental changes in cardiorespiratory patterns of sleep-associated apnea in northern elephant seals. Am. J. Physiol.-Regul. Integr. Comp. Physiol. 267, R1294–R1301, https://doi.org/10.1152/ajpregu.1994.267.5.R1294 (1994).
Piscitelli, M. A., Raverty, S. A., Lillie, M. A. & Shadwick, R. E. A Review of Cetacean Lung Morphology and Mechanics. J. Morphol. 274, 1425–1440, https://doi.org/10.1002/jmor.20192 (2013).
Bernaldo de Quirós, Y. et al. Decompression versus decomposition: distribution, quantity and gas composition of bubbles in stranded marine mammals. Front. Physiol. 3, 1–19 (2012).
Hooker, S. K. et al. Deadly diving? Physiological and behavioural management of decompression stress in diving mammals. Proc. Roy. Soc. B. 279, 1041–1050, https://doi.org/10.1098/rspb.2011.2088 (2012).
Fernandez, A. et al. “Gas and fat embolic syndrome” involving a mass stranding of beaked whales (family Ziphiidae) exposed to anthropogenic sonar signals. Vet. Pathol. 42, 446–457 (2005).
Jepson, P. D. et al. Gas-bubble lesions in stranded cetaceans. Nature 425, 575–576, https://doi.org/10.1038/nature02528,2004 (2003).
García-Párraga, D. et al. Decompression Sickness (“the bends”) in Sea Turtles. Dis. Aquat. Org. 111, 191–205 (2014).
Fernández, A. et al. Deadly acute Decompression Sickness in Risso’s dolphins. Sci. Rep. 7, 13621, https://doi.org/10.1038/s41598-017-14038-z (2017).
Davis, R. W. & Williams, T. M. The marine mammal dive response is exercise modulated to maximize aerobic dive duration. J. Comp. Physiol. A. 198, 583–591 (2012).
Fahlman, A. et al. Ventilation and gas exchange before and after voluntary static surface breath-holds in clinically healthy bottlenose dolphins, Tursiops truncatus. J. Exp. Biol. 222, 1–9, https://doi.org/10.1242/jeb.192211 (2019).
Thompson, D. & Fedak, M. A. Cardiac responses of grey seals during diving at sea. J. Exp. Biol. 174, 139–154 (1993).
Hindell, M. A. & Lea, M.-A. Heart rate, swimming speed, and estimated oxygen consumption of a free-ranging Southern elephant seal. Physiol. Zool. 71, 74–84 (1998).
Kooyman, G., Wahrenbrock, E., Castellini, M., Davis, R. & Sinnett, E. Aerobic and anaerobic metabolism during voluntary diving in Weddell seals: Evidence of preferred pathways from blood chemsitry and behavior. J. Comp. Physiol. B. 138, 335–346 (1980).
Littell, R. C., Henry, P. R. & Ammerman, C. B. Statistical analysis of repeated measures data using SAS procedures. J. Anim. Sci. 76, 1216–1231 (1998).
Acknowledgements
A special thanks to all the trainers and staff at Oceanogràfic and Dolphin Quest, who made this study possible through their dedication and motivation. A special thanks to Lorrie Rea who helped guide us to the past data in elephant seals, and to Marianne Boström for reminding us about Järesta finten. We are grateful to the anonymous reviewers who provided constructive suggestions that helped improve the manuscript. Funding for this project was provided by the Office of Naval Research (ONR Award # N000141613088). Dolphin Quest and the Oceanogràfic provided in kind support of animals, crew and access to resources.
Author information
Authors and Affiliations
Contributions
A.F. and S.M. conceived of the study, designed the experiments, collected and analyzed the data; A.F. carried out the statistical analysis, wrote the paper, and obtained the research funding; J.R.-L., A.J., J.A. participated in the design of the study and was in charge of animal training; L.M.-B. and D.G.P. helped conceive the study, and revised the paper; F.C. helped with the data analysis and revised the paper. All authors gave final approval for publication.
Corresponding author
Ethics declarations
Competing Interests
The authors declare no competing interests.
Additional information
Publisher’s note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Fahlman, A., Miedler, S., Rocho-Levine, J. et al. Re-evaluating the significance of the dive response during voluntary surface apneas in the bottlenose dolphin, Tursiops truncatus. Sci Rep 9, 8613 (2019). https://doi.org/10.1038/s41598-019-45064-8
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-019-45064-8
This article is cited by
-
Doppler echocardiography in a healthy, non-sedated Southern Sea Lion (Otaria flavescens) - a preliminary approach about the feasibility and clinical utility
Veterinary Research Communications (2023)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
