
Abstract
CAPN5 Neovascular Inflammatory Vitreoretinopathy (CAPN5-NIV; OMIM 193235) is a poorly-understood rare, progressive inflammatory intraocular disease with limited therapeutic options. To profile disease effector proteins in CAPN5-NIV patient vitreous, liquid vitreous biopsies were collected from two groups: eyes from control subjects (n = 4) with idiopathic macular holes (IMH) and eyes from test subjects (n = 12) with different stages of CAPN5-NIV. Samples were analyzed by liquid chromatography-tandem mass spectrometry (LC-MS/MS). Protein expression changes were evaluated by principal component analysis, 1-way ANOVA (significant p-value < 0.05), hierarchical clustering, gene ontology, and pathway representation. There were 216 differentially-expressed proteins (between CAPN5-NIV and control vitreous), including those unique to and abundant in each clinical stage. Gene ontology analysis revealed decreased synaptic signaling proteins in CAPN5-NIV vitreous compared to controls. Pathway analysis revealed that inflammatory mediators of the acute phase response and the complement cascade were highly-represented. The CAPN5-NIV vitreous proteome displayed characteristic enrichment of proteins and pathways previously-associated with non-infectious posterior uveitis, rhegmatogenous retinal detachment (RRD), age-related macular degeneration (AMD), proliferative diabetic retinopathy (PDR), and proliferative vitreoretinopathy (PVR). This study expands our knowledge of affected molecular pathways in CAPN5-NIV using unbiased, shotgun proteomic analysis rather than targeted detection platforms. The high-levels and representation of acute phase response proteins suggests a functional role for the innate immune system in CAPN5-NIV pathogenesis.
Similar content being viewed by others


Introduction
CAPN5 Neovascular Inflammatory Vitreoretinopathy (CAPN5-NIV; OMIM 193235) is a rare, progressive inflammatory intraocular disease caused by mutations in the CAPN5 gene1. Before culminating in blindness, CAPN5-NIV disease progresses in a series of pathological stages, characterized by synaptic signaling defects (loss of b-wave on electroretinogram), inflammatory cell infiltration, neovascularization, and intraocular fibrosis (Fig. 1). These 5 stages each mimic common eye diseases that together account for a significant fraction of visual morbidity and blindness (e.g. uveitis, retinitis pigmentosa, proliferative diabetic retinopathy, and proliferative vitreoretinopathy)2. The CAPN5 gene encodes for calpain-5, a regulatory protease expressed in photoreceptors that modulates the biological function of its proteolytic targets3,4. CAPN5-NIV- mutations cause a gain-of-function, have been shown to hyper-activate the protease5,6,7. Although several CAPN5-NIV-causing mutations have been identified, it is not known how a hyperactive protease leads to uveitis1,7,8. Since the underlying mechanisms of CAPN5-NIV are poorly understood, these patients are left with few treatment options and fail conventional immunosuppressive therapy, such as oral corticosteroids and infliximab (anti-TNF-α)2.

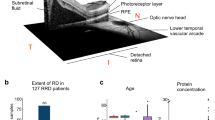
Clinical CAPN5-NIV phenotype: (A,B) Clusters of autoimmune reactive leukocytes in the vitreous cavity (inset, arrows). (C) Electroretinography reveals early synaptic signaling defects in CAPN5-NIV patients, detected as loss of the b-wave. (D) Fundus image of the normal retina. (E) Fundus image of CAPN5-NIV retina showing pigmentary degeneration (arrow). (F) Fluorescein angiography reveals cystoid macular edema at the fovea (arrow), a consequence of intraocular inflammation. (G) Intraocular fibrosis and pre-retinal scar tissue formation (arrow). (H) Vitreous hemorrhage (arrow) caused by retinal neovascularization. (I) Phthisis bulbi and involution of eye tissues at end-stage CAPN5-NIV disease. Images courtesy of Mahajan, et al. (2012).
To identify therapies for NIV, our group has previously taken a drug repositioning approach to treating CAPN5-NIV patients. Drug repositioning applies approved drugs towards new indications, which often are rare diseases with few therapeutic options2. To identify candidate drugs for repositioning, we have used a personalized proteomics approach2,9. Liquid vitreous biopsies from CAPN5-NIV patients undergoing surgery were analyzed by a multiplex ELISA array that simultaneously measured levels of 200 human cytokines. The levels of these cytokines were subsequently analyzed by heatmap clustering and pathway analysis to identify proteins that may be targeted by available drugs. This analysis showed that TNF-α levels were normal, explaining why infliximab therapy failed in these patients. It was further revealed that the CAPN5-NIV vitreous contained abundant levels of vascular endothelial growth factor (VEGF), T-cell proliferative markers, and interleukin-6 (IL-6). Based on this, we repositioned bevacizumab (anti-VEGF monoclonal antibody), intravitreal methotrexate (T-cell inhibitor), and tocilizumab (anti-IL-6 monoclonal antibody) and successfully mitigated neovascularization, inflammatory cell infiltration, and persistent fibrosis in these patients2.
Despite the success of our past personalized proteomics studies for CAPN5-NIV, the original analysis was limited to only 200 cytokines. Since different proteomic platforms can give different results, we sought to obtain a more global view of the CAPN5-NIV vitreous proteome at each stage. Liquid chromatography-tandem mass spectrometry (LC-MS/MS) is a powerful, unbiased analytical technique that ionizes molecular species in complex biochemical mixtures and sorts ions based on their mass-to-charge ratio (m/z)10,11. This technique is used in a “shotgun” approach to catalog and quantify the hundreds to thousands of proteins in a biological sample, which is often performed in tandem with liquid chromatography to separate peptides before they are ionized10. We have utilized LC-MS/MS techniques to catalogue proteins in the normal human retina, vitreous, and retinal pigmented epithelium (RPE)-choroid complex12,13,14. Here, we describe a similar shotgun proteomics study of CAPN5-NIV vitreous, using LC-MS/MS, and perform bioinformatic analysis to detect stage-specific changes and identify potential drug targets for further drug repositioning.
Results
Differential protein expression
Vitreous samples from 8 CAPN5-NIV patients (12 eyes) and 4 idiopathic macular hole (IMH) controls underwent trypsinization and liquid chromatography before analysis by tandem mass spectrometry (Table 1; Supplemental Fig. 1). In control IMH vitreous, we identified 373 ± 70 unique proteins (mean ± SD; n = 4). There were 428 ± 16 unique proteins in Stage II CAPN5-NIV (n = 2), 344 ± 55 in Stage III CAPN5-NIV (n = 5), and 392 ± 56 (n = 5) in Stage IV CAPN5-NIV (Table 1). The most abundant proteins shared among all samples were: serotransferrin (TF), alpha-1-antitrypsin (SERPINA1), and apolipoprotein A-I (APOA1). These proteins were also abundant in Stage II vitreous. The most abundant proteins in control samples were immunoglobulin heavy constant gamma 1 (IGHG1) and transthyretin (TTR). Immunoglobulin heavy constant gamma 1 (IGHG1), complement C3 (C3), and alpha-1-antichymotrypsin (SERPINA3) were the most abundant proteins in CAPN5-NIV samples. Proteomics data were then compared using principal component analysis (PCA). The score plot of PC1 and PC2 showed separation between the 12 CAPN5-NIV cases and 4 IMH controls based on protein intensities that were significantly different between the two groups (Fig. 2A). Protein intensities were further analyzed using 1-way ANOVA and hierarchical clustering. A total of 216 proteins were differentially-expressed among control and CAPN5-NIV samples (90 upregulated proteins in CAPN5-NIV samples and 126 downregulated proteins; p < 0.05; Fig. 2B,C).

Differentially-expressed proteins reveal differences between CAPN5-NIV cases and controls: (A) Principal component analysis (PCA) of the proteomics data. The score plot of PC1 and PC2 shows separation between CAPN5-NIV cases (orange) and IMH controls (green) based on protein intensities that were significantly different between the two groups. (B) Protein intensities were analyzed by 1-way ANOVA. Results are represented as a volcano plot. The horizontal axis (x-axis) displays the log2 fold-change value (CAPN5-NIV vs. controls) and the vertical axis (y-axis) displays the noise-adjusted signal as the -log10 (p-value) from the 1-way ANOVA analysis. (C) Protein intensities were analyzed by hierarchical clustering. A total of 216 proteins were differentially-expressed among control (IMH) and CAPN5-NIV samples (90 upregulated proteins in CAPN5-NIV samples and 126 downregulated proteins; p < 0.05). Results are represented as a heatmap and display protein expression levels on a logarithmic scale. Orange indicates high expression while dark green/black indicates low or no expression.
Gene ontology analysis
To obtain a global view of the protein classifications represented in CAPN5-NIV vitreous, a gene ontology (GO) analysis was performed. Lists of differentially-expressed proteins between CAPN5-NIV and controls were classified by their respective biological process, molecular function, and cellular compartment (Fig. 3A). When proteins were categorized by cellular compartment, we observed a significant fraction of proteins localized to the synapse in controls that decreased in CAPN5-NIV vitreous. Among these proteins were: neurexin-2 (NRXN2), glutamate receptor 4 (GluR4), neurofascin (NFASC), neuronal growth regulator 1 (NREG1), calsyntenin-1 (CLSTN1), and oligodendrocyte-myelin glycoprotein (OMG). Patients in the early stages of CAPN5-NIV disease display altered synaptic signaling characterized by an early decrease in the b-wave amplitude on scotopic flash electroretinogram (rod photoreceptors)4,15. NRXN2 is a neuronal cell adhesion protein that is involved in synapse formation and synaptic regulation in the retina16. Loss of neurexin has been shown to significantly impair visual function by decreasing rhodopsin levels16. GluR4 is a metabotropic glutamate receptor expressed in the rod bipolar cells of the mammalian retina17,18. The loss of these synaptic proteins may contribute to the synaptic signaling defect seen in early CAPN5-NIV disease. However, our proteomic analysis could not differentiate which retinal cells are affected by loss of these proteins in CAPN5-NIV.

Gene ontology and pathway analysis reveals differences among early and late CAPN5-NIV: (A) Differentially-expressed proteins from early and late CAPN5-NIV compared to controls. Gene ontology analysis categorized each protein group by biological process, molecular function, and cellular compartment. (B,C) Differentially-expressed proteins from early CAPN5-NIV compared to controls. Results are represented as a volcano plot. The horizontal axis (x-axis) displays the log2 fold-change value (early CAPN5-NIV vs. controls) and the vertical axis (y-axis) displays the noise-adjusted signal as the -log10 (p-value) from the 1-way ANOVA analysis. Proteins with log2 fold-changes greater than 15 (upregulated) are colored cyan (early) and orange (late) while proteins with log2 fold-changes lower than −15 are represented in green (downregulated). (D) Comparative analysis of upregulated proteins (compared to controls) using Venn diagrams. A total of 74 upregulated proteins are shared among the two stages compared to controls (p < 0.05). (E) Comparative analysis of downregulated proteins using Venn diagrams. A total of 49 downregulated proteins are shared among the two stages compared to controls (p < 0.05). (F) Top ten pathways represented in CAPN5-NIV. Pathways are ranked by their −log (p-value) obtained from the right-tailed Fisher’s Exact Test.
Stage-specific protein expression
Each CAPN5-NIV stage is characterized by different clinical phenotypes and pathological processes, suggesting that they may contain distinct proteins. We therefore sought to identify stage-specific protein signatures that may correlate to these clinical features. First, we compared early CAPN5-NIV (Stages II and III) vitreous to controls (Fig. 3B). A total of 144 proteins were differentially-expressed among control and early CAPN5-NIV samples (86 upregulated proteins in CAPN5-NIV samples and 58 downregulated proteins; p < 0.05; Fig. 3A). Among the significantly upregulated proteins were scavenger receptor cysteine-rich type 1 protein M130 (CD163) and plastin-2 (LCP1). CD163 is a macrophage-specific receptor that indicates the presence M2 macrophages. M2 “repair” macrophages are required for vascular stability and produce VEGF. Previous studies have shown that CD163 production by human macrophages drive cytokine secretion in favor of an anti-inflammatory phenotype19. LCP1 plays a role in activating T-cells by facilitating transport of CD69 and CD2520. Among the significantly-downregulated proteins were several proteins involved in retinal function: retinoschisin (RS1), and opticin (OPTC). RS1 is involved in retinal cell adhesion and is required structure and function in the retina. Loss of RS1 function is most-notably associated with retinoschisis, an inherited vitreoretinal dystrophy associated with vitreous hemorrhage, retinal detachment, and neovascular glaucoma21. Opticin is an extracellular matrix glycoprotein associated with the collagen-integrin matrix in the vitreous. It has been previously shown to display anti-angiogenic activity22. Angiogenesis and vitreous hemorrhage are key features of CAPN5-NIV disease and downregulation of opticin may contribute to the increased retinal neovascularization seen in these patients1,2. There were proteins with known antioxidant activity present in controls that were absent in CAPN5-NIV vitreous: superoxide dismutase (SOD1 and SOD3), peroxiredoxin (PRDX2 and PRDX6), catalase (CAT), clusterin, and glutathione peroxidase 3 (GPX3). Clusterin is an antioxidant protein involved in the removal of cellular debris and has been implicated in reducing the breakdown of the blood-retinal barrier23. Past proteomic studies have shown clusterin levels to be reduced in the vitreous of diabetic retinopathy patients23. CAPN5-NIV patients similarly display breakdown of the blood retina barrier leading to vascular leakage of protein into the vitreous2. Taken together, these results indicate that early CAPN5-NIV stages are characterized by reduced defense against oxidative stress.
Next, we compared late CAPN5-NIV (Stage IV) vitreous to controls (Fig. 3B). A total of 206 proteins were differentially-expressed among control and late CAPN5-NIV samples (79 upregulated proteins in CAPN5-NIV samples and 127 downregulated proteins; p < 0.05; Fig. 3C). Among the significantly-upregulated proteins were those involved in the acute phase response and coagulation cascade: complement components (C1R, C6, C7, C8, and C9), prothrombin, antithrombin 3 (AT3), hemopexin (HPX), and carboxypeptidase B2 (CPB2). Acute phase proteins are a class of proteins whose plasma levels rise or fall dramatically in response to an injury or acute inflammatory event24,25. Many of the acute phase proteins fall into several overlapping functional categories, including complement signaling, coagulation and fibrinolysis, and inflammatory mediators25. Although most acute phase proteins are produced and secreted by hepatocytes, we have previously reported their expression in the normal human vitreous and RPE-choroid complex13,14. Several crystallin proteins were also upregulated (CRYAA, CRYAB, CRYBB1). Crystallins are soluble proteins found in the lens and cornea and were previously shown to have angiogenic properties26. Patients with proliferative diabetic retinopathy display increased vitreous levels of CRYAB, which correlate to increased levels of VEGF27. Consistent with this observation, CAPN5-NIV patients were previously found to have elevated vitreous VEGF levels and intravitreal injection of bevacizumab (anti-VEGF) suppresses neovascularization and clears vitreous hemorrhages2. Among the downregulated proteins was versican (VCAN/CSPG2). Versican is a chondroitin sulfate proteoglycan involved in maintaining the physiologic structure of the vitreous. Interestingly, loss of versican expression and function is implicated in vitreoretinal degeneration (versican vitreoretinopathy; OMIM: 118661)28. A comparative analysis revealed 49 upregulated proteins and 74 downregulated proteins were shared among the three stages, suggesting that different stages of CAPN5-NIV may share similar pathways and classifications of proteins (Fig. 3D,E).
Pathway analysis
To further classify differentially-expressed proteins in CAPN5-NIV vitreous, we performed pathway analysis, which identifies groups of functionally-linked proteins. The top represented pathways in CAPN5-NIV were: complement activation, CD22 mediated B-cell receptor (BCR) regulation, scavenging of heme from plasma, BCR activation by antigens, binding and uptake of ligands by scavenger receptors, cell surface interactions at the vascular wall, FCGR activation, role of phospholipids in phagocytosis, innate immune system, and integrin signaling (Fig. 3F; Supplemental Tables 1–2). The high representation of coagulation cascade members may be due to the presence of blood in the vitreous during collection. Several of the patients underwent surgery for vitreous hemorrhage. The most significantly represented pathways were associated with the acute phase response. Increased levels of acute phase response proteins are a common feature of neuroinflammation, and complement signaling has been implicated in degenerative eye diseases like age-related macular degeneration (AMD)29,30. We identified a total of 25 interaction acute phase proteins that were significantly upregulated in CAPN5-NIV vitreous, suggesting that innate immune elements play a role in the pathogenesis of CAPN5-NIV (Fig. 4).

Acute phase response network: The acute phase response was the most-significantly represented pathway common to all CAPN5-NIV stages. There were 25 acute phase response proteins elevated in CAPN5-NIV. Results are displayed as a protein interaction network with proteins (nodes) represented as circles and connected by lines representing predicted or experimentally-confirmed interactions (edges). Nodes are highlighted by their respective molecular pathway or function: complement cascade, coagulation cascade, and molecular transport.
Discussion
CAPN5-NIV is a poorly-understood inflammatory disease with few therapeutic options. Our data demonstrate significant molecular differences between CAPN5-NIV stages that may provide insight into the molecular mechanisms underlying the disease. Pathway analysis confirmed molecules and pathways previously identified in CAPN5-NIV vitreous (e.g. angiogenesis, IL-6 signaling, and T-cell signaling pathways)2. These findings support the current analytical approach. By using an unbiased proteomic approach rather than targeted proteomics, we identified affected new molecular pathways that were not previously associated with CAPN5-NIV, including acute phase response signaling, the complement cascade, oxidative stress, and synaptic transmission. Furthermore, the CAPN5-NIV vitreous proteome displayed characteristic enrichment of proteins and pathways previously-associated with non-infectious posterior uveitis, rhegmatogenous retinal detachment (RRD), age-related macular degeneration (AMD), proliferative diabetic retinopathy (PDR), and proliferative vitreoretinopathy (PVR; Tables 2 and 3).
With insights into elevated proteins and pathways, we considered FDA-approved drugs or drugs that are currently in clinical trials that could be repositioned for CAPN5-NIV (Fig. 5A,B). For instance, the decrease in antioxidant proteins in later CAPN5-NIV stages may represent a reduced defense against reactive oxygen species (ROS) in later CAPN5-NIV stages. Oxidative stress is associated with photoreceptor damage in several retinal diseases including PDR, retinitis pigmentosa (RP), and AMD and different retinal regions display varying susceptibility to damage by ROS12. Past proteomic studies have showed enrichment for proteins involved in oxidative stress in AMD vitreous31. In our current study, we observed decreased levels of superoxide dismutase enzymes (SOD1 and SOD3) in CAPN5-NIV vitreous. SOD3 has been shown to regulate oxidative stress at the vitreoretinal interface and loss of Sod3 in mice display inner retina signaling abnormalities32. Compounds that activate antioxidant proteins may be beneficial in treating or preventing photoreceptor damage that results from chronic oxidative stress. Several SOD-mimetic compounds (e.g. M40403 and tempol), for example, have been demonstrated to be protective in numerous animal models of acute and chronic inflammation, chemotoxicity, reperfusion injury, and shock33,34,35,36. These compounds may be administered early in CAPN5-NIV disease to prevent accumulation of damaging ROS.

A CAPN5 vitreoretinopathy disease model for therapeutic testing: (A) Illustrations highlighting the clinical phenotype at each CAPN5-NIV stage. Graphical illustrations by Alton Szeto and Vinit Mahajan. Permission to publish granted by original artist. (B) A constructed disease model highlighting the molecular phenotype with associated proteins and correlation to clinical phenotype of CAPN5-NIV. Potential therapeutics which have already been approved or are in trial for use in other diseases are represented based on differentially-expressed proteins and pathways.
The high levels and number of acute phase proteins suggests a potential important role for the innate immune system in CAPN5-NIV pathogenesis. Our previous proteomic studies linked several cytokine-signaling proteins and pathways involved in the adaptive immune response to CAPN5-NIV (e.g. mTOR and PI3K signaling pathways), although innate immune pathways were not detected due to the targeted nature of our analysis2. Innate immune elements have been previously detected in non-diseased eye tissues (e.g. anterior chamber, vitreous, RPE-choroid) and are implicated in a number of neurodegenerative diseases, such as AMD and RP25,37,38. Notably, there was higher representation of acute phase signaling in late CAPN5-NIV (Figs 3F; 4). This increased acute phase response representation may result from non-specific innate immune activation in response to progressive photoreceptor degeneration. Alternatively, hyperactive CAPN5 activity may cause tissue injury that triggers non-specific activation of these innate immune elements. There is some precedent for this as elevated calpain activity is associated with a wide range of disease, including retinal degeneration and neuronal injury39,40,41,42. Aberrant proteolysis of retinal CAPN5 substrates by a hyperactive protease may similarly lead to exposure of peptide epitopes that trigger the autoinflammatory response seen in CAPN5-NIV. Further research, however, is required to elucidate the interplay between the innate and adaptive immune system in CAPN5-NIV pathogenesis.
Our current study has several limitations. Most notably is the limited sample size, especially for Stage II CAPN5-NIV (Table 1). It is rare to obtain Stage II CAPN5-NIV vitreous samples as the indication for vitrectomy in these patients is exceptional (e.g. in the case of epiretinal membrane formation, recalcitrant macular edema, or non-clearing vitreous opacities) as most symptoms in this stage can be managed without surgical intervention. Nevertheless, these data and analyses are valuable as they provide insight into underlying disease mechanisms in a rare patient population with few therapeutic options. Further prospective validation of these candidate biomarkers is required. Another limitation is the discovery-oriented nature of our proteomic analysis. Due to the rarity of surgical samples from CAPN5-NIV patients, we opted for an unbiased data-independent acquisition (DIA) approach which allows for the relative quantification of proteins in our samples. For more reliable and sensitive quantification of candidate biomarkers, a selective reaction monitoring (SRM) approach or multiplex ELISA would be more appropriate in future studies11.
Conclusion
Proteomic analysis is a powerful tool for studying the molecular basis of inflammatory diseases, like CAPN5-NIV9,11,43,44. Although several CAPN5-NIV-causing mutations have been identified, it is unclear how a hyperactive protease leads to retinal disease. Thus, in the near-term the genotype of our CAPN5-NIV patient does little to change treatment outcomes. On the other hand, our proteomics dataset, points to differential expression of multiple disease-associated molecular pathways, a finding that has implications for the immediate treatment of CAPN5-NIV. Further validation of these novel biomarkers and drug targets will improve our understanding of molecular mechanisms underlying CAPN5-NIV and potentially point to therapies for more common causes of blindness.
Methods
Study approval
The study was approved by Stanford University and the University of Iowa’s Institutional Review Board and adhered to the tenets set forth in the Declaration of Helsinki (IRB: 201803853). Patients provided written informed consent for the use of their tissue samples. Patients underwent eye exams that included slit-lamp examination, dilated retinal bio-microscopy and indirect ophthalmoscopy. Data was collected from August 2009 to July 2018. Written informed consent was provided for pictures appearing in the manuscript.
Sample collection
Pars plana vitrectomy was performed using a single-step transconjunctival 23-gauge trocar cannular system (Alcon Laboratories Inc, Fort Worth, TX), and an undiluted 0.5-cc sample of the vitreous was manually aspirated into a 3-cc syringe. Vitreous samples were immediately centrifuged in the operating room at 15,000 g for 5 minutes at room temperature to remove impurities and then finally stored at −80 °C, as previously described9.
Proteomic analysis
Shotgun proteomic mass spectrometry-based measurements were performed in duplicate for control and CAPN5-NIV vitreous samples (Table 1). A liquid chromatography-tandem mass spectrometry (LC-MS/MS) approach was used for the relative quantitation and simultaneous identification of proteins from all samples. Briefly, vitreous protein concentration was determined using the Qubit Protein Quantification assay (Thermo Fisher). Proteins were then extracted from vitreous (20 µg protein per sample), precipitated in chloroform-methanol, dissolved in 0.1% Rapigest detergent in 50 mM ammonium bicarbonate. Trypsin was added to each sample at a ratio 1:40 enzyme/protein and digested overnight at 37 °C. The reaction was quenched by adding 90% formic acid to a final concentration of 2%. A total of 5 µl (1 µg of trypsin-digested peptides) of each sample was then injected into a ChromXP C18 reverse phase analytical column (10 cm in length). High-performance liquid chromatography (HPLC) was performed on a NanoLC Eskigent HPLC pump at a flow rate of 0.2 µl/min using a linear gradient of Buffer B (0.1% formic acid, 98% acetonitrile). Buffer A consisted of 5% acetonitrile, 0.1% formic acid. Mass spectrometry was performed on a Q ExactiveTM HF Hybrid Quadrupole-Orbitrap (Thermo Fisher) mass spectrometer equipped with a nano-LC electrospray ionization source (ThermoFinnigan). Full MS data were recorded on peptides over a 400 to 100 m/z range (positive ion mode). Data-independent acquisition (DIA) was used to generate MS data within a 25 Da fixed window (at 26% normalized collision energy). Biognoysis SpectronautTM Pulsar was used to search the DIA data. The human Uniprot database was used in the database search. Positive identification was set at 1% peptide FDR. K-Nearest Neighbor (KNN) imputation was for missing values. The mass spectrometry proteomics data have been deposited to the ProteomeXchange Consortium via the PRIDE partner repository with the dataset identifier PXD01198745,46,47.
Statistical and bioinformatics analysis
Results were also saved in Excel as.txt format and were uploaded into the Partek Genomics Suite 6.5 software package. The data was normalized to log base 2 and compared using principal component and 1-way ANOVA analysis. For better interpretation, all proteins with non or less significant (p > 0.05) changes were eliminated from the table. The significant values were mapped using the ‘cluster based on significant genes’ visualization function with the standardization option chosen. Gene ontology (GO) analysis was performed in PANTHER48,49. Pie charts were created for the visualization of GO distributions within the list of proteins under the Batch ID search menu. Pie charts were created for each GO term category including biological process, molecular function, and cellular component. Reactome Pathway Analysis software was utilized to determine the most significant pathways50.
Data Availability
The mass spectrometry proteomics data have been deposited to the ProteomeXchange Consortium via the PRIDE partner repository with the dataset identifier PXD011987.
References
Mahajan, V. B. et al. Calpain-5 mutations cause autoimmune uveitis, retinal neovascularization, and photoreceptor degeneration. PLoS Genet 8, e1003001, https://doi.org/10.1371/journal.pgen.1003001 (2012).
Velez, G., Bassuk, A. G., Colgan, D., Tsang, S. H. & Mahajan, V. B. Therapeutic drug repositioning using personalized proteomics of liquid biopsies. JCI Insight 2, https://doi.org/10.1172/jci.insight.97818 (2017).
Schaefer, K. et al. Calpain-5 gene expression in the mouse eye and brain. BMC Res Notes 10, 602, https://doi.org/10.1186/s13104-017-2927-8 (2017).
Schaefer, K. A. et al. Calpain-5 Expression in the Retina Localizes to Photoreceptor Synapses. Invest Ophthalmol Vis Sci 57, 2509–2521, https://doi.org/10.1167/iovs.15-18680 (2016).
Wert, K. J. et al. CAPN5 mutation in hereditary uveitis: the R243L mutation increases calpain catalytic activity and triggers intraocular inflammation in a mouse model. Hum Mol Genet 24, 4584–4598, https://doi.org/10.1093/hmg/ddv189 (2015).
Wert, K. J. et al. Functional validation of a human CAPN5 exome variant by lentiviral transduction into mouse retina. Hum Mol Genet 23, 2665–2677, https://doi.org/10.1093/hmg/ddt661 (2014).
Velez, G. et al. A novel de novo CAPN5 mutation in a patient with inflammatory vitreoretinopathy, hearing loss, and developmental delay. Cold Spring Harb Mol Case Stud 4, https://doi.org/10.1101/mcs.a002519 (2018).
Bassuk, A. G. et al. Structural modeling of a novel CAPN5 mutation that causes uveitis and neovascular retinal detachment. PLoS One 10, e0122352, https://doi.org/10.1371/journal.pone.0122352 (2015).
Velez, G. et al. Precision Medicine: Personalized Proteomics for the Diagnosis and Treatment of Idiopathic Inflammatory Disease. JAMA Ophthalmol 134, 444–448, https://doi.org/10.1001/jamaophthalmol.2015.5934 (2016).
Zhang, Y., Fonslow, B. R., Shan, B., Baek, M. C. & Yates, J. R. 3rd. Protein analysis by shotgun/bottom-up proteomics. Chem Rev 113, 2343–2394, https://doi.org/10.1021/cr3003533 (2013).
Velez, G. et al. Personalized Proteomics for Precision Health: Identifying Biomarkers of Vitreoretinal Disease. Transl Vis Sci Technol 7, 12, https://doi.org/10.1167/tvst.7.5.12 (2018).
Velez, G. et al. Proteomic analysis of the human retina reveals region-specific susceptibilities to metabolic- and oxidative stress-related diseases. PLoS One 13, e0193250, https://doi.org/10.1371/journal.pone.0193250 (2018).
Skeie, J. M. & Mahajan, V. B. Proteomic landscape of the human choroid-retinal pigment epithelial complex. JAMA Ophthalmol 132, 1271–1281, https://doi.org/10.1001/jamaophthalmol.2014.2065 (2014).
Skeie, J. M., Roybal, C. N. & Mahajan, V. B. Proteomic insight into the molecular function of the vitreous. PLoS One 10, e0127567, https://doi.org/10.1371/journal.pone.0127567 (2015).
Tang, P. H. et al. Progression of Scotopic Single-Flash Electroretinography in the Stages of Capn5 Vitreoretinopathy. Retin Cases Brief Rep, https://doi.org/10.1097/ICB.0000000000000828 (2018).
Tian, Y. et al. Neurexin regulates visual function via mediating retinoid transport to promote rhodopsin maturation. Neuron 77, 311–322, https://doi.org/10.1016/j.neuron.2012.11.012 (2013).
Brandstatter, J. H., Koulen, P. & Wassle, H. Diversity of glutamate receptors in the mammalian retina. Vision Res 38, 1385–1397 (1998).
Ghosh, K. K., Haverkamp, S. & Wassle, H. Glutamate receptors in the rod pathway of the mammalian retina. J Neurosci 21, 8636–8647 (2001).
Alvarado-Vazquez, P. A. et al. Macrophage-specific nanotechnology-driven CD163 overexpression in human macrophages results in an M2 phenotype under inflammatory conditions. Immunobiology 222, 900–912, https://doi.org/10.1016/j.imbio.2017.05.011 (2017).
Wabnitz, G. H. et al. Costimulation induced phosphorylation of L-plastin facilitates surface transport of the T cell activation molecules CD69 and CD25. Eur J Immunol 37, 649–662, https://doi.org/10.1002/eji.200636320 (2007).
Sikkink, S. K., Biswas, S., Parry, N. R., Stanga, P. E. & Trump, D. X-linked retinoschisis: an update. J Med Genet 44, 225–232, https://doi.org/10.1136/jmg.2006.047340 (2007).
Le Goff, M. M. et al. Opticin exerts its anti-angiogenic activity by regulating extracellular matrix adhesiveness. J Biol Chem 287, 28027–28036, https://doi.org/10.1074/jbc.M111.331157 (2012).
Zhang, C. et al. The emerging roles of clusterin on reduction of both blood retina barrier breakdown and neural retina damage in diabetic retinopathy. Discov Med 21, 227–237 (2016).
Cruz-Guilloty, F. & Perez, V. L. Molecular medicine: Defence against oxidative damage. Nature 478, 42–43, https://doi.org/10.1038/478042a (2011).
Perez, V. L. & Caspi, R. R. Immune mechanisms in inflammatory and degenerative eye disease. Trends Immunol 36, 354–363, https://doi.org/10.1016/j.it.2015.04.003 (2015).
Dimberg, A. et al. alphaB-crystallin promotes tumor angiogenesis by increasing vascular survival during tube morphogenesis. Blood 111, 2015–2023, https://doi.org/10.1182/blood-2007-04-087841 (2008).
Chen, W., Lu, Q., Lu, L. & Guan, H. Increased levels of alphaB-crystallin in vitreous fluid of patients with proliferative diabetic retinopathy and correlation with vascular endothelial growth factor. Clin Exp Ophthalmol 45, 379–384, https://doi.org/10.1111/ceo.12891 (2017).
Tang, P. H., Velez, G., Tsang, S. H., Bassuk, A. G. & Mahajan, V. B. VCAN Canonical Splice Site Mutation is Associated With Vitreoretinal Degeneration and Disrupts an MMP Proteolytic Site. Invest Ophthalmol Vis Sci 60, 282–293, https://doi.org/10.1167/iovs.18-25624 (2019).
McGeer, P. L. & McGeer, E. G. Inflammation, autotoxicity and Alzheimer disease. Neurobiol Aging 22, 799–809 (2001).
Geerlings, M. J., de Jong, E. K. & den Hollander, A. I. The complement system in age-related macular degeneration: A review of rare genetic variants and implications for personalized treatment. Mol Immunol 84, 65–76, https://doi.org/10.1016/j.molimm.2016.11.016 (2017).
Schori, C. et al. The Proteomic Landscape in the Vitreous of Patients With Age-Related and Diabetic Retinal Disease. Invest Ophthalmol Vis Sci 59, AMD31–AMD40, https://doi.org/10.1167/iovs.18-24122 (2018).
Wert, K. J. et al. Extracellular superoxide dismutase 3 (SOD3) regulates oxidative stress at the vitreoretinal interface. Free Radic Biol Med, https://doi.org/10.1016/j.freeradbiomed.2018.06.024 (2018).
Rosales, M. A., Silva, K. C., Lopes de Faria, J. B., Lopes de Faria, J. M. & Exogenous, S. O. D. mimetic tempol ameliorates the early retinal changes reestablishing the redox status in diabetic hypertensive rats. Invest Ophthalmol Vis Sci 51, 4327–4336, https://doi.org/10.1167/iovs.09-4690 (2010).
Ahmed, L. A., Shehata, N. I., Abdelkader, N. F. & Khattab, M. M. Tempol, a superoxide dismutase mimetic agent, ameliorates cisplatin-induced nephrotoxicity through alleviation of mitochondrial dysfunction in mice. PLoS One 9, e108889, https://doi.org/10.1371/journal.pone.0108889 (2014).
Masini, E. et al. Protective effects of M40403, a selective superoxide dismutase mimetic, in myocardial ischaemia and reperfusion injury in vivo. Br J Pharmacol 136, 905–917, https://doi.org/10.1038/sj.bjp.0704774 (2002).
Setiohadji, B., Irfani, I., Rifada, M., Virgana, R. & Kartasasmita, A. S. The Superoxide Dismutase Mimetic TEMPOL and Its Effect on Retinal Ganglion Cells in Experimental Methanol-Intoxicated Rats. Ophthalmol Ther 7, 167–172, https://doi.org/10.1007/s40123-018-0132-z (2018).
Anderson, D. H., Mullins, R. F., Hageman, G. S. & Johnson, L. V. A role for local inflammation in the formation of drusen in the aging eye. Am J Ophthalmol 134, 411–431 (2002).
Cruz-Guilloty, F. et al. T cells and macrophages responding to oxidative damage cooperate in pathogenesis of a mouse model of age-related macular degeneration. PLoS One 9, e88201, https://doi.org/10.1371/journal.pone.0088201 (2014).
Artal-Sanz, M. & Tavernarakis, N. Proteolytic mechanisms in necrotic cell death and neurodegeneration. FEBS Lett 579, 3287–3296, https://doi.org/10.1016/j.febslet.2005.03.052 (2005).
Randriamboavonjy, V. & Fleming, I. The role of calpain in diabetes-associated platelet hyperactivation. Adv Pharmacol 59, 235–257, https://doi.org/10.1016/S1054-3589(10)59008-2 (2010).
Huang, C. J. et al. Calcium-activated calpain-2 is a mediator of beta cell dysfunction and apoptosis in type 2 diabetes. J Biol Chem 285, 339–348, https://doi.org/10.1074/jbc.M109.024190 (2010).
Ono, Y., Saido, T. C. & Sorimachi, H. Calpain research for drug discovery: challenges and potential. Nat Rev Drug Discov 15, 854–876, https://doi.org/10.1038/nrd.2016.212 (2016).
Mahajan, V. B. & Skeie, J. M. Translational vitreous proteomics. Proteomics Clin Appl 8, 204–208, https://doi.org/10.1002/prca.201300062 (2014).
Velez, G. et al. Proteomic Analysis of Elevated Intraocular Pressure with Retinal Detachment. Am J Ophthalmol Case Rep 5, 107–110, https://doi.org/10.1016/j.ajoc.2016.12.023 (2017).
Vizcaino, J. A. et al. 2016 update of the PRIDE database and its related tools. Nucleic Acids Res 44(456), D447, https://doi.org/10.1093/nar/gkv1145 (2016).
Perez-Riverol, Y. et al. PRIDE Inspector Toolsuite: Moving Toward a Universal Visualization Tool for Proteomics Data Standard Formats and Quality Assessment of ProteomeXchange Datasets. Mol Cell Proteomics 15, 305–317, https://doi.org/10.1074/mcp.O115.050229 (2016).
Deutsch, E. W. et al. The ProteomeXchange consortium in 2017: supporting the cultural change in proteomics public data deposition. Nucleic Acids Res 45, D1100–D1106, https://doi.org/10.1093/nar/gkw936 (2017).
Thomas, P. D. et al. PANTHER: a library of protein families and subfamilies indexed by function. Genome Res 13, 2129–2141, https://doi.org/10.1101/gr.772403 (2003).
Mi, H., Muruganujan, A. & Thomas, P. D. PANTHER in 2013: modeling the evolution of gene function, and other gene attributes, in the context of phylogenetic trees. Nucleic Acids Res 41, D377–386, https://doi.org/10.1093/nar/gks1118 (2013).
Fabregat, A. et al. Reactome pathway analysis: a high-performance in-memory approach. BMC Bioinformatics 18, 142, https://doi.org/10.1186/s12859-017-1559-2 (2017).
Roybal, C. N. et al. Personalized Proteomics in Proliferative Vitreoretinopathy Implicate Hematopoietic Cell Recruitment and mTOR as a Therapeutic Target. Am J Ophthalmol 186, 152–163, https://doi.org/10.1016/j.ajo.2017.11.025 (2018).
Perez, V. L. et al. Elevated levels of interleukin 6 in the vitreous fluid of patients with pars planitis and posterior uveitis: the Massachusetts eye & ear experience and review of previous studies. Ocul Immunol Inflamm 12, 193–201, https://doi.org/10.1080/092739490500282 (2004).
Davuluri, G. et al. Activated VEGF receptor shed into the vitreous in eyes with wet AMD: a new class of biomarkers in the vitreous with potential for predicting the treatment timing and monitoring response. Arch Ophthalmol 127, 613–621, https://doi.org/10.1001/archophthalmol.2009.88 (2009).
Praidou, A. et al. Vitreous and serum levels of vascular endothelial growth factor and platelet-derived growth factor and their correlation in patients with non-proliferative diabetic retinopathy and clinically significant macula oedema. Acta Ophthalmol 89, 248–254, https://doi.org/10.1111/j.1755-3768.2009.01661.x (2011).
Praidou, A. et al. Vitreous and serum levels of platelet-derived growth factor and their correlation in patients with proliferative diabetic retinopathy. Curr Eye Res 34, 152–161, https://doi.org/10.1080/02713680802585920 (2009).
Santos, F. M. et al. iTRAQ Quantitative Proteomic Analysis of Vitreous from Patients with Retinal Detachment. Int J Mol Sci 19, https://doi.org/10.3390/ijms19041157 (2018).
Loukovaara, S. et al. Quantitative Proteomics Analysis of Vitreous Humor from Diabetic Retinopathy Patients. J Proteome Res 14, 5131–5143, https://doi.org/10.1021/acs.jproteome.5b00900 (2015).
Yu, J. et al. Vitreous proteomic analysis of proliferative vitreoretinopathy. Proteomics 8, 3667–3678, https://doi.org/10.1002/pmic.200700824 (2008).
Acknowledgements
We thank Emily Chen and Rajesh Soni for technical assistance. V.B.M. and A.G.B. are supported by NIH grants [K08EY020530, R01EY024665, R01EY025225, R01EY024698, R21AG050437, P30EY026877], Doris Duke Charitable Foundation Grant #: 2013103, Research to Prevent Blindness (RPB) and the Stanford ChEM-H Testing Molecular Hypotheses in Human Subjects Seed Grant. The Barbara & Donald Jonas Laboratory of Regenerative Medicine and Bernard & Shirlee Brown Glaucoma Laboratory are supported by the CDMRP TSCRP:TS080017, National Institute of Health [5P30EY019007, R01EY018213, R01EY024698, R01EY026682, R21AG050437]; National Cancer Institute Core [5P30CA013696]; the Research to Prevent Blindness (RPB) Physician-Scientist Award; unrestricted funds from RPB, New York, NY, USA; the Tistou and Charlotte Kerstan Foundation; the Crowley Family Fund; the Schneeweiss Stem Cell Fund; New York State [C029572];and the Gebroe Family Foundation. G.V. is supported by NIH grants [F30EYE027986 and T32GM007337]. The funding organizations had no role in design and conduct of the study; collection, management, analysis, and interpretation of the data; preparation, review, or approval of the manuscript; and decision to submit the manuscript for publication.
Author information
Authors and Affiliations
Contributions
Dr. Mahajan had full access to all the data in the study and takes responsibility for the integrity of the data and the accuracy of the data analysis. Study concept and design: G.V., S.H.T., A.G.B., V.B.M. Acquisition of data: G.V., J.Y., V.B.M. Analysis and interpretation of data: G.V., J.Y., V.B.M. Drafting of the manuscript: G.V., J.Y., A.S.L., S.H.T., A.G.B., V.B.M. Critical revision of the manuscript for important intellectual content: S.H.T., A.G.B., V.B.M. Statistical analysis: G.V., J.Y., V.B.M. Obtained funding: V.B.M. Administrative, technical, and material support: A.G.B., S.H.T., V.B.M. Study supervision: V.B.M.
Corresponding author
Ethics declarations
Competing Interests
The authors declare no competing interests.
Additional information
Publisher’s note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Velez, G., Yang, J., Li, A.S. et al. Proteomic insight into the pathogenesis of CAPN5-vitreoretinopathy. Sci Rep 9, 7608 (2019). https://doi.org/10.1038/s41598-019-44031-7
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-019-44031-7
This article is cited by
-
Applications of artificial intelligence and bioinformatics methodologies in the analysis of ocular biofluid markers: a scoping review
Graefe's Archive for Clinical and Experimental Ophthalmology (2024)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
