
Abstract
The northern corn rootworm (NCR), Diabrotica barberi Smith & Lawrence, is a major pest of maize (Zea mays L.). This pest has developed resistance to insecticides and adapted to crop rotation and may already be in the early stages of adaptation to toxins produced by Bacillus thuringiensis (Bt). Toxicity bioassays using artificial diet have proven to be valuable for monitoring resistance in many species, but no artificial diet has been developed specifically for NCR larvae. Toward this end, we first evaluated known Diabrotica diets to identify a starting media. We then developed a specialized diet for NCR using an iterative approach. Screening designs including 8 diet components were performed to identify the principal nutritional components contributing to multiple developmental parameters (survival, weight, and molting). We then applied mixture designs coupled with response surface modeling to optimize a blend of those components. Finally, we validated an improved NCR diet formulation that supports approximately 97% survival and molting, and a 150% increase in larval weight after 10 days of feeding compared with the best previously published artificial diet. This formulation appears suitable for use in diet bioassays as a tool for evaluating the resistance of NCR populations to insecticides.
Similar content being viewed by others
Introduction
The northern corn rootworm (NCR), Diabrotica barberi Smith & Lawrence, is a major insect pest of maize (Zea mays L.) in the North American Corn Belt. NCR and the western corn rootworm (WCR), Diabrotica virgifera virgifera LeConte, together, are responsible for more than $1 billion annually due to yield reduction and control costs in the United States1. Rootworm larvae feeding on corn roots cause yield loss from disruption to nutrient and water flow2 and can facilitate infection by soil pathogens3,4. Yield loss also occurs from unharvested grain due to the difficulty of mechanically harvesting lodged plants5,6,7. NCR is highly adaptive and has developed resistance and adaptation to several management tactics, including soil insecticides8 and crop rotation9. The latest management tools, transgenic maize hybrids expressing insecticidal proteins from Bacillus thuringiensis (Bt) Berliner, were recently documented to falter due to resistance of WCR10,11,12 to the toxins. Since the targeted pests of Bt maize are Diabrotica species including both WCR and NCR, the recent increase in NCR populations in some areas causes concern that a similar adaptation may occur with NCR to Bt maize and/or other management practices. In fact, NCR may already be in the early stages of adaptation to Bt13.
The United States Environmental Protection Agency (EPA) requires monitoring programs to detect the development of resistance in insect populations. Diet toxicity bioassays, whereby insects are exposed to toxins on artificial diet, can be a valuable component of such programs. However, this requires an artificial diet capable of supporting insect growth and development. To date, artificial diets of NCR for use in evaluating resistance to Bt maize in wild-type populations have been lacking. Because no artificial diet has been developed specifically for NCR, an artificial diet for southern corn rootworm (SCR), Diabrotica undecimpunctata howardi Barber, has been used in diet toxicity bioassays of a laboratory strain of NCR14. Development of an NCR diet to support development (e.g., survival and molting) would be helpful in detecting resistance development of NCR populations.
Diet development for Diabrotica species started with a formulation for larvae of SCR, which is a generalist that feeds on over 100 different plants. The first SCR formulation developed by Sutter et al.15 consisted of wheat germ and casein as key ingredients. This formulation supported SCR development from egg to adult, but development was slower and fecundity was lower for larvae reared on the diet compared with those fed corn roots15. SCR diet improvements were reported by Rose and McCabe16 and Marrone et al.17 by altering linseed oil, sucrose, antibiotics, and potassium hydroxide. The newest formulation improved SCR development and approached that of larvae reared on corn roots after 6 generations of selection for larval vigor and production of adults on the diet17. The SCR diet has been used in WCR diet bioassays, although growth rates for WCR were poorer than that of SCR and microbial contamination was a major issue18. Later, several modifications of this diet were made by industry and a high level of antibiotics was added into the SCR diet to avoid contamination19.
An initial formulation for WCR larvae was made by Pleau et al.20 with modifications made to a diet for SCR17 by adjusting amounts of several ingredients (i.e., wheat germ, linseed oil, and potassium hydroxide), removing formalin and adding corn root powder. The Pleau et al. diet doubled the weight of WCR larvae compared with larvae reared on the SCR formulation20. Later, the Pleau et al. diet was improved by Huynh et al.21, which resulted in a formulation that further increased the weight of WCR larvae and increased both survival and molting after 11 days compared with the first WCR formulation. We utilized response surface modeling to identify and then optimize the key ingredients in the Pleau et al. diet to develop the improved diet21. The Pleau et al. diet was improved by altering several components (i.e., sucrose, wheat germ, casein, cellulose, corn root powder, linseed oil, and agar) and adding wheat germ oil. The Huynh et al. diet had nearly zero contamination after 11 days when clean laboratory practices were applied21, but contamination was also eliminated from 5 other WCR diets using the same techniques22. This diet was also compatible with all current Bt proteins targeting WCR larvae23.
Past approaches to formulate and develop insect diets involved empirical one-factor-at-a-time (OFAT) and multivariate experiment designs based on mixture designs. For OFAT designs, each diet component was varied independently. Because changing the amount of one factor is confounded with changes in the proportions of other ingredients in a mixture, the OFAT design is not capable of identifying an optimal blend of diet components24. The multivariate geometric approach in combination with a mixture design allows simultaneous varying of multiple diet components and reveals important interactive effects of diet components on several measured responses at a same time21,25,26. Henri Scheffé27 developed mathematical models specifically for response surface modeling based on mixture designs and the corresponding computations are simplified by modern software for design of experiments25,28. The application of response surface methodology combined with a mixture design has been of great value for diet improvement and optimization for WCR21 and another coleopteran insect25,29.
For the current paper, we first compared the performance of 5 published Diabrotica artificial diets for rearing NCR and identified key diet ingredients in the best diet in a 8-component experiment. We then screened 8 protein sources to identify best options. Finally, we optimized specific blends to obtain our desired responses. We combined the response surface method with a mixture design to identify the main drivers of larval development by maximizing larval performance (larval survival, development, and weight) while limiting diet contamination compared with the performance of NCR on existing diets developed for related species (WCR and SCR).
Results
Comparison of five rootworm diets
Larval survivorship to 10 days was significantly different (P < 0.0001, F4,24 = 247.4) among the five tested diets (Fig. 1a). There was no significant difference between WCRMO-1 and Frontier 9800B. The Pleau et al. diet had significantly lower survivorship than WCRMO-1 and Frontier 9800B, whereas no larvae survived to 10 days when reared on Frontier 9757B or the Marrone et al. diet. Average larval dry weight after 10 days varied significantly across diets (P < 0.0001, F4,24 = 122.6). The WCRMO-1 diet had significantly higher larval dry weight than other diets, while the Frontier F9800B diet had significantly lower larval dry weight than the Pleau et al. diet (Fig. 1b). Significantly more NCR larvae molted from 1st to 2nd instar when reared on WCRMO-1 than on any other diet (P < 0.0001, F4,24 = 189.5) (Fig. 1c). The Pleau et al. diet had more successful larval molts compared with Frontier F9800B. Overall, WCRMO-1 supported better NCR larval performance than other diets tested.

Percentage survival (a), average dry weight (b), percentage molting to 2nd instar (c), of northern corn rootworm larvae fed different diet formulations for 10 days. Frontier SCR diets (F9800B & F9757B, Frontier Newark, DE), the Marrone et al. diet17, the Pleau et al. diet20, the WCRMO-1 diet21. Bars with different letters are significantly different (P < 0.0001).
8-component screening experiment for basic nutrition profile
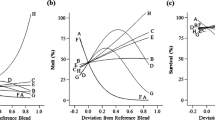
The 8-component mixture design produced significant response surface models for all three measurement criteria; larval weight (P < 0.0001, F9,13 = 37.9), molting (P < 0.0001, F8,14 = 47.3), and survival (P < 0.0001, F8,14 = 19.1). Eight diet components were varied simultaneously: agar, sucrose, wheat germ, casein, cellulose, corn root powder, wheat germ oil and linseed oil. Models for weight and survival had insignificant lack of fit whereas there was a significant lack of fit for molting due to a very small value of pure error. All models were improved by stepwise regression. Values for R2, R2adj and R2pred varied by less than 0.2 (see Supplementary Table S5), indicating good predicted models. The relationships between diet ingredients and larval performance (Fig. 2), depict the effect of increasing the proportion of one component in relation to a reference blend while the relative proportions of all of the other diet components are kept constant30,31 and are called Cox plots. The Cox plot was generated by inverted high and low values, which showed that the slope of the line indicated the inverted direction and magnitude of the influence of the individual factors on the measured response variable, e.g., larval weight. Models for weight and molting indicated that casein, corn root powder and two lipid components had the greatest effects on improving these criteria (Fig. 2a,b) whereas a model for survival revealed that wheat germ, corn root powder and wheat germ oil were the primary drivers (Fig. 2c). Increases in the two lipid components resulted in minor improvements to WCR larval performance21. Casein was the main protein source of the diet formulation, and was an important driver for weight and molting. Based on that observation, we proceeded to test alternative protein sources.

Cox plot of larval responses to deviations from a reference blend diet in the proportions of ingredients in an 8-component mixture experiment. (a) weight, (b) molting, (c) survival. Reference blend proportions: agar = 0.046, sucrose = 0.203, wheat germ = 0.253, casein = 0.203, cellulose = 0.203, corn root: 0.075, wheat germ oil = 0.009, linseed oil = 0.009. A: agar, B: sucrose, C: wheat germ, D: casein, E: cellulose, F: corn root powder, G: wheat germ oil, H: linseed oil. The slope of the line shows the inverted direction and magnitude of the influence of the ingredient on measured responses. The orientation of the graphs is inverted from what is normally expected, so effects that appear negative are positive.
8-protein mixture experiment
The protein screening design yielded significant response surface models for larval weight (P < 0.0001, F12,17 = 35.6), molting (P < 0.0001, F9,20 = 16.4), and survival (P < 0.0001, F8,21 = 33.7) by varying 8 different protein sources: corn gluten meal, cottonseed meal, casein, plant protein, whey protein, perfect amino, yeast extract, and egg powder (see Supplementary Table S6). Models for weight had insignificant lack of fit whereas models for molting and survival had significant lack of fit due to very small values of pure error. R2, R2adj and R2pred of all models were clustered with differences <0.3. The relationships between protein sources and larval performance (Fig. 3) indicated animal protein sources (casein, egg powder, and whey protein) had the greatest effects on improving all three criteria of larval performance (Fig. 3a–c). In addition, plant protein had positive effects on larval performance and cottonseed meal had positive effect on survival. In contrast, corn gluten meal, perfect amino, and yeast extract had negative effects on all developmental traits. In Fig. 3, the slope shows the direction and magnitude of the impact of each ingredient on all measured responses.

Cox plot of larval responses to deviations from a reference blend diet in the proportions of ingredients in an 8-protein screening experiment. (a) weight, (b) molting, (c) survival. Reference blend proportions: corn gluten meal = cottonseed meal = casein = plant protein = whey protein = perfect amino = yeast extract = egg powder = 0.375. A: corn gluten meal, B: cottonseed meal, C: casein, D: plant protein, E: whey protein, F: perfect amino, G: yeast extract, H: egg powder. The slope of the line indicates the direction and magnitude of the influence of the ingredient on measured responses.
3-protein mixture-amount experiment
The 3-protein mixture-amount was used to identify an optimal 3-component blend for larval weight (P < 0.0001, F5,25 = 10.2), molting (P < 0.0001, F8,22 = 14.1), and survival (P < 0.0001, F6,23 = 8.8) using: casein, egg powder, and whey protein (see Supplementary Table S7). R2, R2adj and R2pred of all models were clustered with differences <0.3. Lack of fit of models for weight was significant due to very small values of pure error, while models for molting and survival had insignificant lack of fit. The relationships between protein sources and larval performance were shown in ternary plots (Fig. 4). Possible combinations of proportions of the mixtures of three components were revealed in the ternary plots. The magnitude of the response variables is coded in color and can be envisioned as perpendicular to the page as indicated by labelled isobars (Fig. 4a–c).

Contour plots of larval responses to a mixture-amount design of casein: egg powder: whey protein at 10 days post infestation. (a) weight, (b) survival, and (c) molting. (d–f) interaction of egg powder, casein and concentration on (d) weight, (e) survival, and (f) molting; proportions of whey protein = 0.34. Color bars display the magnitude of the measured response. (g) Overlaid optimization plot of larval responses; yellow area represents the optimum region for weight, molting, and survival that are obtained when three ingredients (i.e., egg powder, casein, whey protein) are used.
Models for weight and molting indicated that a blend of casein and egg powder was optimal for maximum weight and molting (Fig. 4a,b). High proportions of casein resulted in a decrease in all developmental traits. Whey protein had positive effect on survival (Fig. 4c), but a negative effect on weight and molting when added as a component of the protein mixture. Results also showed very strong blending effects of egg powder and casein on larval weight and molting, but an antagonistic blending effect of these components on survival (see Supplementary Fig. 1). Such a trade-off between survival and weight gain has been shown to occur in other beetles reared on artificial diets32.
The contour plots (Fig. 4d–f) represent the interactive effects of casein and egg powder and the amount of mixture while proportions of whey protein to diet mixture were constant at 0.34:1. The labelled isobars and color indicate the magnitude of the measured response. All contour plots indicated that a higher total amount of the mixture could yield a better formulation. Ratios of egg powder: casein at 4:1, 1:1, 3:1 are predicted to produce the best formulation for weight, molting and survival, respectively (Fig. 4d–f).
Contamination
Since all experiments had minor contamination (<1%), there was no evidence for a relationship between contamination and diet ingredients. Similarly low contamination rates were seen in Huynh et al.21, Ludwick et al.23, and Meihls et al.22.
Diet optimization
The diet development for NCR larvae produced a formulation, referred to hereafter as NCRMO-1 (Table 1). This formulation was calculated with Design-ExpertTM by optimizing key ingredients based on the integrated evaluation of all developmental traits for WCR. An overlay graph comprised of the contour plots from each response laid on top of each other was generated to identify a region where all measured responses were simultaneously maximized (Fig. 4g).
Model validation and diet improvement
The NCRMO-1 diet yielded better larval performance compared with the best previously published diet for WCR, WCRMO-1. The formulation for NCR larvae was modified from WCRMO-1, with several modifications including the elimination of corn root powder, linseed oil and wheat germ oil, substitution of sucrose with glucose, and addition of egg powder (Table 1). Other ingredients were at the same level as in WCRMO-121. After 10 days of feeding on NCRMO-1, larval dry weight, survival and molting rate were 0.44 mg, 97%, and 97% compared to 0.28 mg, 98% and 96%, respectively when reared on WCRMO-1 (Fig. 5).

Larval dry weight, percent successful completion of molt, and survival for northern corn rootworm larvae reared on NCRMO-1 and WCRMO-1 diets for 10 days. Means within bars followed by different letters are significantly different (P < 0.05). Mean ± SEM.
Discussion
We applied a multivariate geometric approach in combination with an n-dimensional mixture design to develop the first artificial diet formulated specifically for rearing NCR. The algorithm used consisted of five steps including evaluation of existing diets, identification of basic nutritional profiles, exploration of key nutrients, maximizing key ingredients, and evaluation of formulations, as presented in Fig. 6. The application of geometric and mathematical approaches including response surface modeling33, response surface modeling combined with n-dimensional mixture design21,25,29, and orthogonal experimental designs34 have been used for improvement of existing diets for other insect species. By applying the response surface methodology based on a multivariate mixture design, we identified key ingredients and then optimized the key ingredients to create the best formulation that maximized life history criteria (survival, molting, and weight) for NCR.

Flow chart for diet development algorithm.
Our results indicated that protein ingredients were key components that had the greatest effect on larval performance, i.e., changing the concentration of these ingredients strongly affected the life history parameters. In contrast, other ingredients (wheat germ, cellulose, sucrose, linseed oil, wheat germ oil, and agar) had little or negative contributions on the overall response parameters (Fig. 2). We found that NCR larvae performed better with animal proteins compared with 8 other protein sources tested, including plant and yeast proteins. The addition of animal protein sources (casein, egg powder, and whey protein) and plant protein improved all measured responses (weight, molting, and survival) whereas the inclusion of other protein ingredients (corn gluten meal, cottonseed meal, perfect amino, and yeast extract) resulted in a decrease in all three measured responses, except for a positive effect of cottonseed meal on survival (Fig. 3a–c). The nonlinear blending effects indicated that a blend of casein and egg powder contributed the most to improved insect performance compared with all combinations of the three animal protein sources (Fig. 4d–f). Casein at high proportions (Fig. 4c) caused a decrease in survival when changed in combination with egg powder and whey protein. Our results revealed that wheat germ had a complex relationship with the life history parameters. Wheat germ at high proportions improved survival, but caused a decrease in molting (Fig. 2b,c). Similar effects of wheat germ on insect performance have been reported previously for WCR larvae20,21. Wheat germ is documented as a key ingredient for nutritional value, digestibility, bioavailability, and palatability, but at high proportions can cause detrimental effects on insect development35.
We found that corn root powder was a key ingredient that had a strong positive influence on life history parameters. The addition of corn root powder increased all larval performance parameters such as weight, molting and survival (Fig. 2a–c). The important role of corn root powder was also documented in WCR diets, whereas an increase in the proportion of root powder improved molting and survival but also caused a decrease in weight when added at high concentrations20,21. Corn roots contain host recognition cues and feeding stimulants, but also contain factors which have a repellent effect on WCR larvae36,37,38. Corn root powder is not currently available for purchase, making it a limiting factor for users. In this study, we identified, characterized and optimized key protein sources (Figs 3 and 4) that allowed us to eliminate corn root powder, making our formulation more widely applicable.
Our results revealed significant differences in NCR larval development among rootworm diets (Figs 1 and 5). WCRMO-1, a superior diet for WCR21, supported NCR larval performance better than SCR diets and another WCR diet. This is possibly because both WCR and NCR are nearly monophagous on corn roots, although they can also survive on some grass species39,40. There was zero NCR survival to day 10 when feeding on diets containing approximately 1% formalin, e.g., Frontier F9757B and Marrone et al. diets. A detrimental effect of formalin on weight was reported previously for WCR. Larval weight reduced to half when feeding on diets with an addition of 1% formalin20. In contrast, at 10 days post-infestation, survival and molting rate of NCR larvae reared on NCRMO-1 (without formalin) were approximately 97%. Larval dry weight increased 150% with NCRMO-1 compared with WCRMO-1 (Fig. 5).
Although the ultimate goal of insect artificial diet research might be to serve as a comparable substitute for natural foods25, only a few diet formulations can be considered to have achieved that goal over the past 60 years35. However, artificial diets are also desired to enable the study of insect species of interest41,42,43, to reduce labor or space41, to reduce time for host plant rearing44, and to provide a standardized diet for insect bioassays21,23. This is the first report of a diet formulation developed specifically for NCR larvae. NCRMO-1 supported high survival, weight, and percentage of larvae molted to second instar after 10 days. Additionally, this formulation had basically zero contamination. Consequently, this formulation is suitable for use in diet bioassays for monitoring resistance programs of NCR, and for the study of basic NCR biology and physiology.
Materials and Methods
Insects
NCR eggs (a diapausing strain) were obtained from the North Central Agricultural Research Laboratory in Brookings, South Dakota. Eggs were surface-treated using a procedure described by Pleau et al.20 (see Supplementary Methods).
Diet preparation
Frontier SCR diets (F9800B & F9757B, Frontier, Newark, DE) were purchased and poured according to procedures by manufacturers. Other diets including the Marrone et al. diet, the Pleau et al. diet and the WCRMO-1 diet were made using a procedure described in Pleau et al.20, with some modifications by Huynh et al.21 (see Supplementary Methods).
Insect diet bioassays
The diet bioassays were conducted as described in Huynh et al.21. All materials used in the diet assays were surface-treated via exposure to UV light for 10 min in a biological cabinet. Each formulation was tested in a 12-well row of a 96 well plate and values from the 12 sub-samples were averaged to yield a single replicate. Each formulation was replicated at least 4 times. Eight different formulations were randomly assigned on the plate. One larva (<24 h after hatching) was placed in each well of the diet plate using a fine paintbrush. A sealing film (TSS-RTQ-100, Excel Scientific, Inc., Victorville, CA) was used to cover the plate and one vent hole was made per well using an insect pin (size 0). The plates were stored at 25 °C in darkness for 10 days. Collections made at 10 days were used to determine differences in survival, weight and molting to the 2nd instar. Larval survival, time of molt to 2nd instar, and evidence of diet contamination were recorded daily. Live larvae were collected and pooled within a replication into 95% ethanol, dried in an oven (602752, Blue M Therm Dry Bacteriological Incubator) at 50 °C for 2 days, and then were weighed using a micro balance (MSU6.6S-000-DM, Sartorius Lab Instruments GmbH & Co. KG, Goettingen, Germany).
Experimental approach
An iterative approach was used to develop an artificial diet for NCR. An initial experiment was conducted by comparing five current rootworm diets. A screening design was conducted to explore the nutritional profile of NCR by varying 8 components using a mixture design. Polynomial equations were generated to describe the impact of diet ingredients on the measured responses (i.e., larval weight, molting, survival, and contamination). A screening experiment was used to characterize the effects of 8 different protein sources on the measured responses using an 8-component mixture design. A mixture-amount design45 was also used to identify an optimum formulation of 3 protein ingredients that had the greatest effect on the measured responses. The predicted optimal blend was then validated in a comparison experiment.
Comparison of five rootworm diets
Three SCR diets included the two Frontier diets (F9800B & F9757B, Frontier, Newark, DE) plus the Marrone et al. diet17. The two WCR diets included Pleau et al. diet20 and WCRMO-121. An improved formulation was chosen based on overall larval performance (i.e., larval weight, molting, and survival) after 10 d of feeding in diet assays.
8-component mixture design for basic nutrition profile
Since the WCRMO-1 diet was the best of the five public rootworm diets evaluated, improvements started from this diet. It consisted of 18 diet ingredients21. An exploratory experiment to identify the basic nutritional profile of NCR simultaneously varied 8 diet components including casein, cellulose, corn root powder, wheat germ, wheat germ oil, linseed oil, and agar. Other ingredients (vitamin and salt blends, preservatives and antibiotics) were kept constant at the levels in the WCRMO-121. No effort was made to de-convolute the mixtures of Vanderzant vitamin and Wesson salt that are widely used in insect diets. The exploratory design was created with Design-Expert (v.10.0, Stat-Ease, Inc., Minneapolis, MN) resulting in 24 design points including vertex, center, 7-blend, and axial check blend points (see Supplementary Table S1)28. The design included 7 model, 11 lack of fit, and 5 pure error degrees of freedom46.
8-protein screening design
The 8-component mixture experiment indicated casein, corn root powder, lipid ingredients and agar had positive effects on both NCR larval weight and molting. Survival was removed as a criterion due to more than 90% survival across almost all design points. Lipid ingredients (wheat germ oil and linseed oil) were removed due to their minor contributions in WCR diets21. Glucose was substituted for sucrose because glucose is a main component of sugar in corn roots36. In this study, 8 different protein sources were concurrently screened to characterize their contributions to larval performance (weight, molting, and survival) by constructing an I-optimal mixture design47,48. This design consisted of 32 design points with 7 model, 17 lack of fit and 5 pure error degrees of freedom (see Supplementary Table S2). Concentrations of other ingredients were the same as for WCRMO-1 (see Supplementary Table S3).
3-protein mixture design
The 3-proteins responsible for the largest effects on NCR larval weight, molting, and survival were used to construct a D-optimal mixture-amount design45,49 sufficient for a Scheffé quadratic-quadratic polynomial response surface model. This design consisted of 32 design points with 17 model, 6 lack of fit and 8 pure error degrees of freedom (see Supplementary Table S4). Other ingredients were kept constant as shown in see Supplementary Table S3. Optimal blends were predicted using overlay response surface plots combined with a simplex hill-climbing algorithm included in Design-Expert™ software25.
Diet optimization
A formulation for maximizing larval performance (survival, molting, and weight) was calculated with Design-ExpertTM (Stat-Ease, Inc., Minneapolis, MN). This software used overlay response surface and direct search methods50 to maximize the desirability function51.
Model validation
All three measures of larval performance (survival, molting, and weight) identified by the response surface mixture model were compared with those of larvae reared on WCRMO-1 diet21.
Statistical analyses
Survival and molting data were generated by dividing the number of live larvae and successful larval molts from 1st to 2nd instar per replicate, respectively, by the initial number of larvae infested and multiplying by 100 to obtain percentages. Weight data were generated by dividing total dry weight per replicate by the number of live larvae while dead larvae were recorded as 0 in weight.
In the diet comparison experiments, measured parameters of larval performance on the rootworm diets at 10 days after infestation were analyzed as a randomized complete block design using PROC MIXED in SAS52. All percent variables were arcsine square-root transformed prior to the analysis to meet assumptions of normality and homoscedasticity.
In the mixture experiments, all possible models from 1st degree to 4th degree polynomials for each measured response of larval performance (larval survival, proportion of successful larval molts and larval weight) were generated with Design Expert®. Model selection as described in Lapointe et al.25. Briefly, the criteria included a lack of aliased terms, low residual values, low model P-value, nonsignificant lack of fit, low standard deviation, high R2, R2adj and R2pred 25, close agreement between R2adj and R2pred, and a low PRESS value46,53. When two or more models were satisfactory, the selected model was then further evaluated according to adequacy tests as described by Anderson and Whitcomb28,54.
Data Availability
All pertinent data are found in the figures and tables. Requests for data and additional information should be submitted to the corresponding author.
Change history
15 January 2020
An amendment to this paper has been published and can be accessed via a link at the top of the paper.
References
Mitchell, P. D. Costs and benefits of controlling pest Diabrotica in maize in the United States. In, 24th IWGO Conference and 3rd International Conference of Diabrotica Genetics. IWGO, Freiburg, Germany (2011).
Kahler, A. L., Olness, A. E., Sutter, G. R., Dybing, C. D. & Devine, O. J. Root damage by western corn rootworm and nutrient content in maize. Agron. J. 77, 769–774 (1985).
Palmer, L. T. & Kommedahl, T. Root-infecting Fusarium species in relation to rootworm infestations in corn. Phytopathology 59 (1969).
Kurtz, B., Karlovsky, P. & Vidal, S. Interaction between western corn rootworm (Coleoptera: Chrysomelidae) larvae and root-infecting Fusarium verticillioides. Environ. Entomol. 39, 1532–1538 (2010).
Spike, B. P. & Tollefson, J. J. Relationship of plant phenology to corn yield loss resulting from western corn rootworm (Coleoptera: Chrysomelidae) larval injury, nitrogen deficiency, and high plant density. J. Econ. Entomol. 82, 226–231 (1989).
Spike, B. P. & Tollefson, J. J. Yield response of corn subjected to western corn root worm (Coleoptera: Chrysomelidae) infestation and lodging. J. Econ. Entomol. 84, 1585–1590 (1991).
Spike, B. P. & Tollefson, J. J. Western corn rootworm (Coleoptera: Chrysomelidae) larval survival and damage potential to corn subjected to nitrogen and plant density treatments. J. Econ. Entomol. 81, 1450–1455 (1988).
Hamilton, E. W. Aldrin resistance in corn rootworm beetles. J. Econ. Entomol. 58, 296–300 (1965).
Krysan, J. L., Foster, D. E., Branson, T. F., Ostlie, K. R. & Cranshaw, W. S. Two years before the hatch: rootworms adapt to crop rotation. Bull. ESA 32, 250–253 (1986).
Zukoff, S. N. et al. Multiple assays indicate varying levels of cross resistance in Cry3Bb1-selected field populations of the western corn rootworm to mCry3A, eCry3.1Ab, and Cry34/35Ab1. J. Econ. Entomol. 109, 1387–1398 (2016).
Ludwick, D. C. et al. Minnesota field population of western corn rootworm (Coleoptera: Chrysomelidae) shows incomplete resistance to Cry34Ab1/Cry35Ab1 and Cry3Bb1. J. Appl. Entomol. 141, 28–40 (2017).
Gassmann, A. J., Petzold-Maxwell, J. L., Keweshan, R. S. & Dunbar, M. W. Field-evolved resistance to Bt maize by western corn rootworm. PloS one 6, e22629 (2011).
Calles-Torrez, V. et al. Transgenic Bt corn, soil insecticide, and insecticidal seed treatment effects on corn rootworm (Coleoptera: Chrysomelidae) beetle emergence, larval feeding injury, and corn yield in North Dakota. J. Econ. Entomol. 111, 348–360 (2018).
Oyediran, I. O. et al. Susceptibility of northern corn rootworm Diabrotica barberi (Coleoptera: Chrysomelidae) to mCry3A and eCry3.1Ab Bacillus thuringiensis proteins. Insect Sci. 23, 913–917 (2016).
Sutter, G. R., Krysan, J. L. & Guss, P. L. Rearing the southern corn rootworm on artificial diet. J. Econ. Entomol. 64, 65–67 (1971).
Rose, R. I. & McCabe, J. M. Laboratory rearing techniques for the southern corn rootworm. J. Econ. Entomol. 66, 398–400 (1973).
Marrone, P. G., Ferri, F. D., Mosley, T. R. & Meinke, L. J. Improvements in laboratory rearing of the southern corn rootworm, Diabrotica undecimpuncta howardi Barber (Coleoptera: Chrysomelidae), on an artificial diet and corn. J. Econ. Entomol. 78, 290–293 (1985).
Pereira, A. E., Carneiro, N. P. & Siegfried, B. D. Comparative susceptibility of southern and western corn rootworm adults and larvae to vATPase-A and Snf7 dsRNAs. J. RNAi and Gene Silencing 12, 528–535 (2016).
Zhao, J. Z. et al. mCry3A-selected western corn rootworm (Coleoptera: Chrysomelidae) colony exhibits high resistance and has reduced binding of mCry3A to midgut tissue. J. Econ. Entomol. 109, 1369–1377 (2016).
Pleau, M. J., Huesing, J. E., Head, G. P. & Feir, D. J. Development of an artificial diet for the western corn rootworm. Entomol. Exp. Appl. 105, 1–11 (2002).
Huynh, M. P. et al. Diet improvement for western corn rootworm (Coleoptera: Chrysomelidae) larvae. PloS one 12, e0187997 (2017).
Meihls, L. N. et al. Comparison of six artificial diets for support of western corn rootworm bioassays and rearing. J. Econ. Entomol, https://doi.org/10.1093/jee/toy268 (2018).
Ludwick, D. C. et al. A new artificial diet for western corn rootworm larvae is compatible with and detects resistance to all current Bt toxins. Sci. Rep. 8, 5379 (2018).
Niedz, R. P. & Evens, T. J. Design of experiments (DOE)—history, concepts, and relevance to in vitro culture. In Vitro Cell. Dev. Biol. – Plant 52, 547–562 (2016).
Lapointe, S. L., Evens, T. J. & Niedz, R. P. Insect diets as mixtures: Optimization for a polyphagous weevil. J. Ins. Physiol. 54, 1157–1167 (2008).
Raubenheimer, D. Toward a quantitative nutritional ecology: the right-angled mixture triangle. Ecol. Monogr. 81, 407–427 (2011).
Scheffé, H. Experiments with mixtures. Royal Stat. Soc., Series B (Methodological) 20, 344–360 (1958).
Anderson, M. J. & Whitcomb, P. J. RSM simplified: optimizing processes using response surface methods for design of experiments. (CRC press, 2004).
Lapointe, S. L., Evens, T. J., Niedz, R. P. & Hall, D. G. Artificial diet optimized to produce normative adults of Diaprepes abbreviatus (Coleoptera: Curculionidae). Environ. Entomol. 39, 670–677 (2010).
Smith, W. F. Experimental design for formulation (SIAM, 2005).
Lindig, O. H. & Malone, O. L. Oviposition of boll weevils fed diets containing germinated cottonseed puree or cottonseed meats puree. J. Econ. Entomol. 66, 566–567 (1973).
Lapointe, S. L., Niedz, R. P. & Evens, T. J. An artificial diet for Diaprepes abbreviatus (Coleoptera: Curculionidae) optimized for larval survival. Fla. Entomol. 93, 56–62 (2010).
Lee, K. P. et al. Lifespan and reproduction in Drosophila: New insights from nutritional geometry. Proc. Natl. Acad. Sci. 105, 2498–2503 (2008).
Tan, X.-L., Wang, S. & Zhang, F. Optimization an optimal artificial diet for the predatory bug Orius sauteri (Hemiptera: Anthocoridae). PloS one 8, e61129 (2013).
Cohen, A. C. Insect diets: science and technology, 2nd ed., (Taylor & Francis Group, 2015).
Bernklau, E. J. & Bjostad, L. B. Identification of feeding stimulants in corn roots for western corn rootworm (Coleoptera: Chrysomelidae) larvae. J. Econ. Entomol. 101, 341–351 (2008).
Bernklau, E. J., Hibbard, B. E., Dick, D. L., Rithner, C. D. & Bjostad, L. B. Monogalactosyldiacylglycerols as host recognition cues for western corn rootworm larvae (Coleoptera: Chrysomelidae). J. Econ. Entomol. 108, 539–548 (2015).
Bernklau, E. J., Hibbard, B. E., Norton, A. P. & Bjostad, L. B. Methyl anthranilate as a repellent for western corn rootworm larvae (Coleoptera: Chrysomelidae). J. Econ. Entomol. 109, 1683–1690 (2016).
Oyediran, I. O., Hibbard, B. E., Clark, T. L. & French, B. W. Selected grassy weeds as alternate hosts of northern corn rootworm (Coleoptera: Chrysomelidae). Environ. Entomol. 33, 1497–1504 (2004).
Oyediran, I. O., French, B. W., Clark, T. L., Dashiell, K. E. & Hibbard, B. E. Prairie grasses as hosts of the northern corn rootworm (Coleoptera: Chrysomelidae). Environ. Entomol. 37, 247–254 (2008).
Beck, S. D. & Stauffer, J. F. An aseptic method for rearing European corn borer larvae. J. Econ. Entomol. 43, 4–6 (1950).
Vanderzant, E. S. & Reiser, R. Aseptic rearing of the pink bollworm on synthetic media. J. Econ. Entomol. 49, 7–10 (1956).
Vanderzant, E. S. & Reiser, R. Studies of the nutrition of the pink bollworm using purified casein media. J. Econ. Entomol. 49, 454–458 (1956).
Vanderzant, E. S., Reiser, R. & Ivy, E. Methods for the mass rearing of the pink bollworm. J. Econ. Entomol. 49, 559–560 (1956).
Cornell, J. A. Experiments with mixtures: designs, models, and the analysis of mixture data, 3rd ed. (John Wiley & Sons 2002).
Myers, R. H. & Montgomery, D. C. Response surface methodology: process and product optimization using designed experiments, 2nd ed. (Willey, 2002).
Laake, P. On the optimal allocation of observations in experiments with mixtures. Scand. J. Stat., 153–157 (1975).
Goos, P., Jones, B. & Syafitri, U. I-optimal design of mixture experiments. J. Am. Stat. Assoc. 111, 899–911, https://doi.org/10.1080/01621459.2015.1136632 (2016).
Czitrom, V. Mixture experiments with process variables: D-optimal orthogonal experimental designs. Commun. Stat. Theory Methods 17, 105–121 (1988).
Derringer, G. Simultaneous optimization of several response variables. J. Qual. Technol. 12, 214–219 (1980).
Myers, R. H., Montgomery, D. C. & Anderson-Cook, C. M. Response surface methodology: process and product optimization using designed experiments, 4th ed., (John Wiley & Sons, 2016).
SAS version 9.4 (SAS Institute, 2013).
Allen, D. M. Mean square error of prediction as a criterion for selecting variables. Technometrics 13, 469–475 (1971).
Anderson, M. J. & Whitcomb, P. J. Using graphical diagnostics to deal with bad data. Qual. Eng. 19, 111–118 (2007).
Acknowledgements
The authors thank Julie Barry, Nelson Chad, Emily Harrington, and Brock Andreasen for technical assistance. We also thank the Monsanto Corn Rootworm Knowledge Program for helping to fund some of the work. This article reports the results of research only. Mention of a proprietary product does not constitute an endorsement or recommendation for its use by the USDA or the University of Missouri.
Author information
Authors and Affiliations
Contributions
M.P.H., B.E.H. and T.A.C. contributed to writing the first draft of the manuscript. M.P.H., S.L.L., R.P.N. and T.A.C. designed the experiments and performed the analyses. M.P.H. performed the experiments. B.E.H., B.W.F., D.L.F., K.S.S. and T.A.C. provided materials required for the experiments. All authors edited and approved the final version of the manuscript.
Corresponding author
Ethics declarations
Competing Interests
The authors declare no competing interests.
Additional information
Publisher’s note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Huynh, M.P., Hibbard, B.E., Lapointe, S.L. et al. Multidimensional approach to formulating a specialized diet for northern corn rootworm larvae. Sci Rep 9, 3709 (2019). https://doi.org/10.1038/s41598-019-39709-x
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-019-39709-x
This article is cited by
-
Chromobacterium Csp_P biopesticide is toxic to larvae of three Diabrotica species including strains resistant to Bacillus thuringiensis
Scientific Reports (2022)
-
Development of a nondiapausing strain of northern corn rootworm with rearing techniques for both diapausing and nondiapausing strains
Scientific Reports (2021)
-
Evaluation of various commodities for the development of the yellow mealworm, Tenebrio molitor
Scientific Reports (2020)
-
Development of an improved and accessible diet for western corn rootworm larvae using response surface modeling
Scientific Reports (2019)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
