
Abstract
Animal morphology and behavior often appear to evolve cooperatively. However, it is difficult to assess how strictly these two traits depend on each other. The genitalia morphologies and courtship behaviors in insects, which vary widely, may be a good model for addressing this issue. In Diptera, phylogenetic analyses of mating positions suggested that the male-above position evolved from an end-to-end one. However, with this change in mating position, the dorsoventral direction of the male genitalia became upside down with respect to that of the female genitalia. It was proposed that to compensate for this incompatibility, the male genitalia rotated an additional 180° during evolution, implying evolutionary cooperativity between the mating position and genitalia direction. According to this scenario, the proper direction of male genitalia is critical for successful mating. Here, we tested this hypothesis using a Drosophila Myosin31DF (Myo31DF) mutant, in which the rotation of the male genitalia terminates prematurely, resulting in various deviations in genitalia direction. We found that the proper dorsoventral direction of the male genitalia was a prerequisite for successful copulation, but it did not affect the other courtship behaviors. Therefore, our results suggested that the male genitalia rotation and mating position evolved cooperatively in Drosophila.
Similar content being viewed by others


Introduction
The evolution of animal behaviors often appears to be accompanied by morphological changes, suggesting that the behavior and morphology evolved cooperatively1,2,3,4,5,6. One well-known example of such evolutionarily cooperativity is the relationship between the beak shape of Darwin’s finches and their various eating habits1,3. These birds show variations in food sources, including the nectar and pollen of flowers, fruits, seeds, insects, snails, and even the blood of seabirds1,3, and their beak shapes vary depending on their eating habits1,3. Furthermore, the genes responsible for the evolutionary diversification of the beak morphology have been identified1,2. Therefore, morphological changes that appear to show cooperative evolution with behaviors can be attributed to genomic modification. If the beak shapes, but not the behaviors associated with eating habits, changed, such changes would not make sense and vice versa. This logic leads to the idea that morphology and behavior need to evolve cooperatively. However, it has been difficult to assess to what extent organ morphology is critical for and couples with a particular animal behavior, mostly because the experimental modification of organ morphology is difficult to achieve without generating other side effects.
Insect species exhibit a great variety of mating behaviors4. Given that insects also have large variations in copulatory organ morphologies, insects could be particularly useful for studying the cooperative evolution between mating behavior and genitalia morphology. For example, insects have at least five typical mating positions: end-to-end, male-above, female-above, belly-to-belly, and false-male above4. In the end-to-end position mating, a male and a female copulate end-to-end in opposing directions through copulatory organs located at their posterior ends (Fig. 1a, upper and 1b, left). In the male-above position mating, a male mounts a female, and they copulate via their copulatory organs at the posterior end (Fig. 1a, lower and 1b, right).

Evolutionary relationship between mating position and male genitalia rotation in dipterans. (a) Phylogenetic tree showing the evolution of mating positions in Diptera. Illustrations show the mating of mosquitos (upper, end-to-end position) and flies (bottom, male-above position). (b) Hypothesis for the cooperative evolution between mating position and male genitalia rotation. Red-blue triangles show male genitalia, and black-grey triangles represent female genitalia. If the male genitalia rotate 180°, their dorsoventral direction becomes upside down. Under this condition, the dorsoventral directions of the male and female genitalia are in accordance in the end-to-end position of mosquitoes (left). However, in the male-above position, this relative dorsoventral direction of the male genitalia with respect to those of the female would be upside down. It was previously proposed that this inconsistency in the dorsoventral direction of the female and male genitalia could be overcome by an additional 180° rotation of the male genitalia in flies (right). This hypothesis implies that cooperative evolution occurs between the mating position and the rotation of male genitalia.
The order Diptera encompasses species with one pair of wings and halters, including flies and mosquitos (Fig. 1a)6. Primitive dipteran species like mosquitos tend to mate in the end-to-end position, while the male-above position is common in higher dipterans, such as flies, suggesting that the end-to-end position is ancestral to the male-above position (Fig. 1a)4,5,6. Upon the presumptive evolution from the end-to-end to the male-above position, the dorsoventral direction of the male genitalia became reversed with respect to that of the female genitalia (Fig. 1b). Therefore, the genitalia direction needed to be modified so that the male and female copulatory organs would physically couple. It was proposed that one way to solve the incompatibility in the dorsoventral direction of the genitalia was to rotate the male copulatory organ (Fig. 1b)4,5,6. Interestingly, in many dipteran species, the male genitalia rotate to a certain angle during the pupal or adult stage, temporarily or permanently4,5,6. In species that show temporary rotation, the genitalia usually have some torsional flexibility4,7, while the rotation angle is fixed in species in which the male genitalia rotate permanently7,8,9.
In Diptera, the rotation angle of the genitalia is well correlated with the mating positions. A 180° rotation is found in species that mate in the end-to-end position or its derivatives. For example, the common house mosquito Culex pipiens mates end-to-end10 (Fig. 1b, left), and the yellow fever mosquito, Aedes aegypti, uses a belly-to-belly position, which is a variant of the end-to-end position and keeps the same dorsoventral direction of the male genitalia4,5,11. On the other hand, in the fruit fly Drosophila melanogaster, the male genitalia rotate 360° clockwise as viewed from the posterior end, and the male-above position is used (Fig. 1b, right). Thus, the additional 180° rotation of the male genitalia in Drosophila (D.) melanogaster appears to compensate for the discordance of the male genitalia direction in flies in the evolution from the end-to-end to the male-above position (Fig. 1b). These observations led to the proposal that incompatibility in the dorsoventral direction of the genitalia, which is associated with the evolution of the mating position, may be overcome by rotating of the male genitalia6. According to this scenario, the evolution of morphology (genitalia rotation) and behavior (mating position) could have occurred cooperatively (Fig. 1b).
Thus, assuming that the evolution of the genitalia rotation and mating positions occurred cooperatively, it is important to know how strictly these two traits depend on each other. If these two traits are very tightly interdependent, it is possible that an intermediate step occurred between the cooperative evolution of the genitalia rotation and mating position. However, although the angle of the genitalia rotation is precisely fixed in flies and mosquitoes, it is possible that mating behavior is flexible enough to overcome discordance in the dorsoventral direction of the genitalia. Here we addressed these issues using D. melanogaster as an experimental model.
The mechanisms underlying the male genital rotation have been studied extensively in D. melanogaster7,9. In particular, the directional rotation of the male genitalia is an excellent system for studying the mechanisms of left-right asymmetric development7,9. In the pupal stage, the 360° clockwise rotation is achieved by a combination of 180° rotations in two adjacent abdominal segments, abdominal segment 8 anterior and posterior7,9. During studies of left-right asymmetric development in this species, Myosin31DF (Myo31DF) mutants were identified, in which the left-right asymmetric development is reversed in various organs, including the male genitalia rotation7,12,13,14,15,16. Myo31DF encodes a Drosophila ortholog of Myosin ID12,13,15,16. The null mutation of Myo31DF is homozygous viable and fertile14,15. In most adult males homozygous for null or loss-of-function alleles of Myo31DF, the male genitalia rotate 360° counterclockwise7,12,13,14. Therefore, the roles of Myo31DF are specific to the left-right asymmetric development to some extent, although recent studies revealed that Myo31DF also plays essential roles in other cellular events17,18. Importantly, in addition to the inversion of the rotation direction, the rotation of the male genitalia is sometimes prematurely terminated in Myo31DF mutant flies14. Therefore, we can obtain male adult flies with genitalia that have rotated to various angles, from 0° (no rotation) to 360° (full rotation; circumversion), in the same genetic background.
In this study, using these Myo31DF mutant flies, we analyzed whether the normal dorsoventral direction of the male genitalia is a prerequisite for successful mating in the male-above position in D. melanogaster. We found that successful copulation required a certain degree of accuracy in the dorsoventral direction of the male genitalia. Our results suggest that the male–above position of this species evolved cooperatively with the additional 180° rotation of the male genitalia from the ancestral end-to-end position, as previously proposed based on the phylogenic relationship between mating behavior and male genitalia rotation6,7.
Results
Myo31DF mutant male flies have genitalia with angle deviations
In Myo31DF homozygotes, the rotation of the male genitalia in some individuals stopped at various angles, resulting in male genitalia with “angle deviation” (Fig. 2). In this study, we obtained male flies with genitalia showing angle deviation in homozygotes of Myo31DFK2, a null mutant allele of Myo31DF that is homozygous viable and fertile13,14.

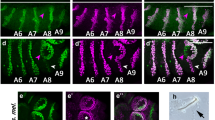
Classification of male genitalia with angle deviation. (a and b) Definition of the angle deviation in male genitalia. We measured the angle (shown in red) between the midline of the abdomen (solid black line) and the line connecting the anus and penis (broken magenta line). A photograph of male genitalia with the angle deviation (a) and its schematic representation (b) are shown. cl, clasper; ap, anal plate. (c) Eight classes of angle deviation in male genitalia: 0° (Left 22°-Right 22°), Right 45° (Right 22°-Right 67°), Right 90° (Right 67°-Right 112°), Right 135° (Right 112°-Right 157°), 180° (Right 157°- Left 157°), Left 135° (Left 157°-Left 112°), Left 90° (Left 112°-Left 67°), Left 45° (Left 67°-Left 22°). (d–g) Male genitalia of wild-type (d) and of Myo31DFK2 mutant (Myo31DF) flies with 0° (e), Right 45° (f), and 180° (g) angle deviation. The head of the magenta arrow shows the penis, and the tail indicates the anus in (d)–(g).
To evaluate how much the angle deviation of the male genitalia influenced reproduction efficiency, we classified the male genitalia with various degrees of angle deviation into eight categories (Fig. 2a–c). As schematically shown in Fig. 2b, we first defined the dorsoventral line of the male genitalia, which passed through the anus and penis (middle of claspers) (broken arrow in Fig. 2a,b). We then measured the angle formed by the midline of the abdomen (solid line in Fig. 2a,b) and the dorsoventral line of the male genitalia (broken arrow in Fig. 2a,b). Based on these angles, we defined 8 classes of angle deviation (classes: 0°, Right 45°, Right 90°, Right 135°, 180°, Left 135°, Left 90°, Left 45°) (Fig. 2c). Figure 2d–g shows typical examples of male genitalia with normal and deviated angles.
Proper dorsoventral direction of the male genitalia is a prerequisite for successful reproduction
We next analyzed the success rate of reproduction involving the male flies with genitalia belonging to each of the 8 classes of angle deviation obtained from Myo31DFK2. A single male belonging to each class was mated with three virgin females. Following the removal of each male, the females were kept in the medium to allow egg-laying for another five days. The females were then discarded, and the medium was incubated for another five days. If offspring were found in the medium, the mating was judged as successful.
First, we validated that our experimental conditions were sufficient for reproduction between wild-type females and males with genitalia of the normal direction (Fig. 3a,b). Single wild-type males or Myo31DFK2 homozygous males with genitalia in the normal direction (the genitalia rotated counterclockwise, but their final dorsoventral direction was normal) were mated with wild-type females for 1, 2, 3, and 4 days, followed by the recovery of offspring as described above. In all cases, the success rate of reproduction was more than 80% (Fig. 3a,b, Supplementary Table S1). We decided to use a 4-day mating period, because the success rate of reproduction was 100% in these two types of males (Fig. 3a,b and Supplementary Table S1).

Males with genitalia exhibiting less angle deviation had an advantage in reproduction. (a,b) Reproduction success rates (%) after mating for 1, 2, 3, and 4 days. A single wild-type male (a) or Myo31DFK2 male with genitalia in the normal dorsoventral direction (b) was mated with three wild-type virgin females. (c and d) Reproduction success rates (%) for males with 8 classes of genitalia angle deviation, after mating for 4 days. Virgin females used for the mating were wild type (c) or Myo31DFK2 homozygotes (d).
Using the above experimental conditions, we next examined the reproduction rates using males with genitalia belonging to each of the eight classes of angle deviation (Fig. 3c,d). As shown in Fig. 3c, matings between wild-type females and males with genitalia of the 0°, Right 45°, Right 90°, Right 135°, Left 90°, and Left 45° angle deviations led to offspring at rates of 66.2, 33.9, 1.96, 3.18, 3.03, and 46.2%, respectively, whereas the males with genitalia of the 180° and Left 135° angle deviation did not have any offspring (Fig. 3c and Supplementary Table S2). Matings between Myo31DFK2 homozygous females and males with genitalia of the 0°, Right 45°, Right 90°, and Left 45° angle deviations led to offspring at rates of 75.0, 60.0, 9.80, and 65.0%, respectively, whereas the males with genitalia of the Right 135°, 180°, Left 135°, and Left 90° angle deviation did not have any offspring (Fig. 3d and Supplementary Table S2). Therefore, mating success strongly favored males with no or small angles of deviation (classes of the 0°, Left 45°, and Right 45° angle deviations). The same tendencies were observed with wild-type and Myo31DFK2 homozygous females, indicating that the strong reproductive superiority of the normal dorsoventral genitalia direction did not depend on a specific genetic background. Although males with genitalia in the normal direction yielded offspring at 100% frequency under these experimental conditions (Fig. 3a,b), males with the class 0° angle deviation had offspring only at rates of 66.2 and 75.0% with wild-type and Myo31DFK2 homozygote females, respectively (Fig. 3c,d and Supplementary Table S2). These results suggested that even a small angle deviation that was less than 22° decreased the reproduction success rate with wild-type and Myo31DFK2 homozygous females, with statistical significance in the case of wild-type females (Fisher’s exact test, P = 0.02879 for wild type and P = 0.1026 for the Myo31DFK2 homozygote).
Although males that had genitalia with the normal or almost normal direction had a great advantage for successful reproduction, we also found that males with an angle deviation of 45° or more still had offspring. In particular, males with genitalia with an angle deviation more than 67° (see Fig. 2c) still had offspring, although their success rate was very low (Supplementary Table S2). These results suggested that the males may have some ability to adjust the angle of their genitalia for copulation, although we cannot exclude the possibility that females contribute to this adjustment.
We also noted that males with the Left 45° versus Right 45° angle deviations did not show a marked difference in their reproduction success rate (Fig. 3c,d). In Myo31DFK2 homozygous males, the genitalia rotate counterclockwise12,14. Therefore, the genitalia only slightly rotated in males with the Left 45° angle deviation, but they rotated until slightly short of completion in males with the Right 45° angle deviation; that is, the Right 45° and Left 45° angle deviation was achieved by a 315° and 45° counterclockwise rotation, respectively. However, despite the 270° difference in rotation angle, the males with genitalia belonging to these two classes of angle deviation showed similar reproduction success rates with wild-type or Myo31DFK2 homozygous females (Fisher’s exact test, P = 0.3349 for wild type, P = 0.7921 for Myo31DFK2) (Fig. 3c,d). Thus, we speculated that the final direction of the genitalia, but not the genitalia rotation itself, had a significant physiological role in the success of reproduction.
Angle deviation of the male genitalia reduces the success rate of copulation
The mating behavior of Drosophila consists of multiple steps, including orientation, tapping, following, singing (wing vibration), licking, and attempted copulation (mounting trial), before copulation19,20. Thus, to understand how the proper dorsoventral direction of the male genitalia affects reproduction success, we next investigated which step of the mating behavior was obstructed by the angle deviation of the male genitalia. Among them, copulation appeared the most likely to be sensitive to an abnormality in the dorsoventral direction of the genitalia. Thus, we first examined the success rate of copulation between wild-type females and males with genitalia with angle deviations classified as 0°, Right 45°, and 180°, obtained from Myo31DFK2 stock, and a control (w1118 otherwise wild type). A single virgin female was placed in a standard mating chamber, followed by the addition of a single male with genitalia at a normal or deviated angle (Fig. 4a). To analyze the mating behavior quantitatively, we video-recorded them for one hour after the addition of the male. To facilitate the video analysis, we developed a program to quickly identify the period of copulation. In this program, the image of flies (Fig. 4b) was binarized (Fig. 4c), and the shape of the flies was automatically selected and arranged in 3D space, where the binarized images were stacked along a time line (Fig. 4d and Supplementary videos 1–3).

Analysis of courtship behavior using a novel image-processing program. (a) A snapshot of the video data of mating behavior between a virgin wild-type female and a male with genitalia with angle deviation or a control male with genitalia in the normal dorsoventral direction. (b and c) Snapshot of the original video (b) and its corresponding binarized picture (c) from the video recording of mating behavior in a single chamber. (d) Processed image in which the binarized images were automatically arranged in 3D space along a time line. A moving bar (yellow line) that was synchronized with the 2D image in b, c was inserted into the 3D image. Black triangles show time points.
In this analysis, a mating was defined as successful copulation if the male stayed on the back of a female for more than one minute during the one-hour experimental period. In the images processed by our program, the shapes of female and male flies partially overlap each other during copulation (Fig. 4d). We can then find the pictures from the video that correspond to “slices” in the 3D space; the yellow line in Fig. 4d corresponds to the pictures shown in Fig. 4b and c. We confirmed that using this program, we could accurately detect copulation events and determine the period of copulation using the processed image data from the video. If the male did not start wing vibration within 10 minutes, we did not include these data in the subsequent analysis, according to previous reports21,22. In our experiments, however, most of the males started the wing vibration (Supplementary Fig. S1 and Supplementary Table S3). Using the video data and image-analysis program, we found that males with the 0° angle deviation successfully copulated at 88.9% (Fig. 5a and Supplementary Table S4). We also found that the copulation rate of the control males, otherwise wild-type males whose genetic background was normalized to Myo31DFK2 by backcrosses, was 56.3% (Fig. 5a and Supplementary Table S4). However, the rate of successful copulation was severely reduced if the male had genitalia classified as Right 45° (7.14%) and 180° (0%) angle deviation (Fig. 5a and Supplementary Table S4). Therefore, the normal dorsoventral direction of the male genitalia was a great advantage for successful copulation. To confirm that our definition of successful copulation (a male mounting a female for more than one minute) was correlated with fecundity, we recovered each female from the chamber and examined whether she left offspring in the standard medium (Fig. 5b and Supplementary Table S4). In these experiments, all of the pairs of single females and males were found to copulate only once during one-hour period (Supplementary Fig. S2 and Supplementary videos 1 and 2). We found that males with genitalia classified as the 0° and Right 45° angle deviation and the control male had offspring at 83.3, 7.14, and 56.3%, respectively (Fig. 5b and Supplementary Table S4). These results showed a very similar tendency to the results of the copulation success rates (compare Fig. 5b with Fig. 5a). We also found that males with genitalia classified as 0° and Right 45° angle deviation and the control males that successfully copulated in the courtship assay had offspring at 93.8, 100, and 100% frequency, respectively (Supplementary Table S5). On the other hand, males with genitalia classified as 0°, Right 45°, and 180° angle deviation and the control males that failed to copulate in the courtship assay, did not have any offspring in any case (Supplementary Table S6). These results demonstrated that the copulation success rate showed a very good correlation with fecundity. Therefore, we concluded that the superiority of the males that had genitalia in the normal dorsoventral direction in having offspring (Fig. 3), was mostly attributed to a high success rate of copulation.

Males with genitalia exhibiting less angle deviation had more successful copulation. (a) Copulation success rates (%) of males with genitalia with angle deviation classified as 0° (0), Right 45° (45), and 180° (180) and of control males (control). (b) Reproduction success rates (%) of males with genitalia with angle deviation classified as 0° (0), Right 45° (45), and 180° (180) and of control males (control). (c) Courtship latency (seconds) of males with genitalia with angle deviation classified as 0° (0), Right 45° (45), and 180° (180) and of control males (control), shown as boxplots. In each boxplot, the middle band is the median (50th percentile), and the length of the box shows the 1st and 3rd quartile (25th and 75th percentile). (d) CI of males with genitalia with angle deviation classified as 0° (0), Right 45° (45), and 180° (180) and of control males (control). The mean and standard error (error bars) are shown. In a-d, all the females were wild type, and the number of examined males is shown in parentheses as n. ***P-value < 0.01. N.S., not significant statistically.
Deviation in the genitalia direction does not affect courtship behaviors
Although we ascertained that the reduction in successful copulation was the major cause of the decrease in fecundity associated with the angle deviation of the male genitalia, it was still possible that other courtship behaviors were also influenced by the angle deviation of the genitalia. Using the video images, we measured the start time of courtship (first wing vibration) for the first 10 minutes after adding a single male that had genitalia with 0°, Right 45°, or 180° angle deviation or a single control male (backcrossed w1118) to the standard mating chamber containing a virgin female (Fig. 5c). This measurement was designated as the “courtship latency,” as described previously21,22,23. We found that the majority of these males initiated wing vibration within the first 10 minutes (Supplementary Fig. S1 and Supplementary Table S3). However, if a male did not start the wing vibration within 10 minutes, we did not include it in the rest of the analysis, as previously reported21,22. Our results revealed that the courtship latency did not show a significant difference among males with the 0°, Right 45°, and 180° angle deviation and control males (Kruskal-Wallis test, P = 0.3559) (Fig. 5c, Supplementary Table S7, and Supplementary Table S8).
The “courtship index” (CI) is generally used to quantify how enthusiastically a single male courts a single female for the first 10 minutes21,22,24. The CI is calculated as the percentage of time spent in all of the components of courtship, including tapping, following, wing extension and vibration, licking, and attempted copulation19,20,22,24,25. Using the same video data, we obtained the CI and compared the values among males with the 0°, Right 45°, and 180° angle deviation and control males (backcrossed w1118). We did not find a significant difference in the CI among the males with genitalia with angle deviation (Fig. 5d, Supplementary Table S7 and Supplementary Table S9). However, the CI of control males was lower than that of males with genitalia classified as 0°, Right 45°, and 180° angle deviation, which carried the Myo31DFK2 mutation (Fig. 5d, Supplementary Table S7 and Supplementary Table S9). Although we do not know the reason for the difference in CI at this point, it is possible that Myo31DF has a weak phenotype in courtship behavior. Alternatively, our backcross may not have been sufficient to uniformalize the genetic background. Nevertheless, our results demonstrated that the deviation in genital rotation did not affect the courtship latency or CI. Therefore, we concluded that the decrease in successful copulation was the main cause of the reduction in fecundity associated with angle deviation of the male genitalia.
In addition to the courtship latency and CI, we examined whether the male-above mating position was influenced by angle deviation of the male genitalia. We examined all of the video data of the courtship assay involving the males with genitalia classified as Right 45° and 180° angle deviation. However, we did not observe any atypical mating positions attempted by these males, in any case examined (see Supplementary videos 2 and 3 as typical examples). Therefore, regardless of the dorsoventral direction of the male genitalia, the mating behaviors leading to the male-above position were fixed. Taking all of our results together, we demonstrated that the normal dorsoventral direction of the male genitalia is a prerequisite for successful mating in the male-above mating position in D. melanogaster. Therefore, our findings suggest that cooperative evolution can exist between mating behavior and male genitalia rotation in this species.
Discussion
Cooperative evolution between organ morphology and behavior is a well-established concept1,2,3,4,5,6. Phylogenetic analyses in various animal groups have demonstrated remarkable coincidences between organ morphology and behavior1,2,3,4,5,6. In this study, we sought to obtain evidence supporting the idea that mating behaviors cooperatively evolved with the rotation of male genitalia in dipteran insects.
In these insects, the evolution from the end-to-end to the male-above position should result in an incompatibility in the dorsoventral direction of the male genitalia with respect to that of the female genitalia4,5. It was previously proposed that this incompatibility was overcome by the additional 180° rotation of the male genitalia (in total, a 360° rotation called “circumversion”)4,5,7. In this study, we showed that males with genitalia resulting from complete circumversion had a great advantage for reproduction over those with deviation in the rotation angle of the genitalia. Our results indicated that the proper dorsoventral direction of the male genitalia is a prerequisite for successful copulation in the male-above mating position in Drosophila. Therefore, our results are consistent with the previously proposed idea that the mating position of dipterans evolved cooperatively with changes in the rotation angle of the male genitalia4,5.
In this analysis, we found that males with genitalia classified as the Left 45° and Right 45° angle deviation still copulated and had offspring (Fig. 3c,d). Therefore, males may be able to overcome a small aberration in the dorsoventral direction of their genitalia, although we could not exclude the possibility that females also contribute to this adjustment. In D. melanogaster, the males have genitalia sensilla, including bristles on the genital claspers, which detect the position of the mating partner and contribute to the male’s proprioception26. Acebes et al. showed that ablating the genital sensilla caused a reduction in mating frequency and decreased the male’s ability to detect his position on the female’s back26. Therefore, in our experiments, it is possible that the sensitivity of the male genitalia contributed to the adjustment to a small aberration in the dorsoventral direction of the male genitalia. However, Acebes et al. also showed that ablating the genital sensilla does not affect the male’s attempt to mount the female back, demonstrating that the male’s attempt to copulate on the female’s back is an unalterable component of male-above mating26. This idea is consistent with our observation that the males with genitalia classified as Right 45° and 180° (upside down genitalia) angle deviation still used the male-above mating position, and no other mating positions, in all of the video data examined (n = 14 and n = 10, respectively) (see Supplementary videos 2 and 3 as typical examples). Upon an alteration in body structure, mating behaviors are also found to be inflexible in other insects. For example, the South American four spot-roach, Eublaberus distanti has a highly ritualized mating behavior, which requires male wing rise27. Males in which the wings have been partially or completely surgically removed still perform the full sequence of mating behaviors and complete their courtship ritual27. Therefore, in cases in which a particular body structure and mating behavior are cooperatively required to achieve successful mating, the mating behavior is often inflexible to adjust to a change in body structure, as found here for the dorsoventral direction of the male genitalia.
In this study, we obtained Myo31DF mutant male flies with genitalia exhibiting various deviations in the rotation angle. In our classification of the angle deviation, the Right 45° and Left 45° males had a deviation of the same angle, which was left-right inversed. However, there is a 270° difference in the amount of rotation needed to achieve these deviations; the Right 45° and Left 45° angle deviations were achieved by 315° and 45° counterclockwise rotations, respectively. However, we did not find a noticeable difference between the success rates of reproduction between these two categories (Fig. 3c,d, Supplementary Table S2). These results suggested that the amount of genital rotation itself does not impact reproduction; rather, the final dorsoventral angle of the genitalia resulting from the rotation is critical for successful copulation. In this context, Huber et al. proposed based on phylogenetic analyses that morphological asymmetry itself is not advantageous for reproduction in spiders and insects, but rather the newly adopted mating position is4. Their idea is consistent with our experimental observation that the degree of genitalia rotation did not affect reproduction if the final dorsoventral direction of the male genitalia was the same.
In our experiments, males with genitalia demonstrating all classes of angle deviation were obtained from Myo31DF. We found that the males with genitalia classified as 0°, Right 45°, and 180° angle deviation and control males, whose genetic background was unified with Myo31DF through five backcrosses, did not show significant differences in courtship latency or in the percentage of males initiating the courtship (Fig. 5c and Supplementary Table S8). However, although the CI was not significantly different among males with genitalia classified as 0°, Right 45°, and 180° angle deviation, the CI of the w1118/Y control males was lower than those of the males with angle deviations in their genitalia (Fig. 5d, Supplementary Table S7 and Supplementary Table S9). Therefore, the Myo31DFK2 mutation may affect the CI. In support of this possibility, Myo31DF was recently shown to be involved in the formation of neuromuscular junctions and in TNF signaling17,18. Alternatively, the genetic background of the control males may not have become completely unified with that of Myo31DF. However, all of our conclusions were based on comparisons between males with genitalia showing the classified angle deviations as well as the normal direction, all of which were on the sameMyo31DF genetic background. Therefore, the observed difference in the CI of the control males does not affect our conclusion that the normal dorsoventral direction of the male genitalia was highly preferable for successful copulation.
Although our hypothesis is based on the single example of D. malanogaster, it supports the idea that the male–above position evolved cooperatively with the additional 180° rotation of the male genitalia from the ancestral end-to-end position, which was proposed before4,5,6,7. However, we also found that Drosophila males have a very limited ability to adjust to a deviation in the direction of their own genitalia. In contrast, the ancestral species that used the end-to-end position might have had high flexibility in the mating position, which gave it the ability to tolerate the future additional 180° rotation of the male genitalia. That is, such flexibility might have subsequently allowed cooperative evolution between the mating position and the rotation of male genitalia. Alternatively, the evolution from the end-to-end to male-above position might have gone through an intermediate step, such as the false-male-above or male-above with flexed male abdomen position4,5,6,28. In this scenario, the mating position evolved before the additional 180° rotation of the male genitalia. To evaluate these ideas, further phylogenetic analyses of the relationship between mating behavior and male genitalia rotation are required.
Materials and Methods
Fly stocks
Flies were cultured on standard medium at 25°. Canton-S was used as the wild-type strain. Myo31DFK2 is a null mutant of Myo31DF12. In all experiments, the Myo31DFK2 mutant line carrying w1118 was used (w1118; Myo31DFK2). white1118 (w1118) is a previously described loss-of-function allele of w29,30.
Collection and classification of Myo31DF homozygous males with deviations in the rotation angle of their genitalia
In the majority of Myo31DFK2 homozygous males, the genitalia rotate 360° counterclockwise, which is the left-right inversed direction of their wild-type counterpart12,14. However, the rest of the males had genitalia whose rotation stopped before it was completed. The deviation in the rotation angle of the male genitalia (angle deviation) was defined as previously described with slight modifications14. The angle formed by the midline of the abdomen and the dorsoventral line that passes though the anus and penis (middle of the claspers) was measured with Image J (http://rsb.info.nih.gov/ij/) on a photograph of the genitalia of each male (VB-7010, Keyence) (Fig. 2a,b). Based on this angle, the angle deviations were classified into eight groups: 0° (Left 22°-Right 22°), Right 45° (Right 22°-Right 67°), Right 90° (Right 67°-Right 112°), Right 135° (Right 112°-Right 157°), 180° (Right 157°- Left 157°), Left 135° (Left 157°-Left 112°), Left 90° (Left 112°-Left 67°), and Left 45° (Left 67°-Left 22°) (Fig. 2c).
Evaluating the effect of male genitalia angle deviation on fecundity
Wild-type and Myo31DFK2 homozygous females were collected six to eight hours after eclosion and kept in fresh medium for 3–5 days. To confirm that all females were virgin, the medium in which these females were kept was incubated for at least another week at 25 °C to assure that they did not bear progeny.
One male classified according to its genitalia angle deviation and three virgin females were then cultured together in a vial of standard medium for four days. Under these conditions, the wild-type and Myo31DFK2 homozygous males with genitalia in the normal direction successfully mated with wild-type virgin females and yielded progenies at 100% (Fig. 3a,b). After four days, the males were recovered from the culture, their genitalia were photographed, and the angle deviation was measured as described above. To collect the potential progenies, the females were kept in the same medium for another 5 days and then discarded, followed by further incubation of the medium for another 5 days. If progeny were found in the medium, the mating was regarded as successful.
Courtship assays
To analyze the potential influences of the male genitalia angle deviation on mating behaviors, we measured a set of courtship parameters: copulation success rate, courtship latency21,22,23, courtship index (CI)19,20,22,24,25, and reproduction success rate (fecundity). All females were wild type. The males were w1118/Y; Myo31DFK2 with genitalia with classified angle deviations and the control w1118 line that was backcrossed with w1118/Y; Myo31DFK2 (see backcross method below). The flies used in these experiments were collected six to eight hours after eclosion as virgins using carbon dioxide anesthesia, and males were collected and roughly classified according to angle deviation (following Fig. 2a–c). Until used for the tests, the female and male flies were maintained individually for another three to five days and five to seven days, respectively, in vials containing standard medium, under a 12-hour light-dark cycle19,31.
For courtship assays, a wild-type virgin female was placed in a small chamber (8-mm in diameter and 3-mm high) followed by the addition of a male with genitalia in the normal direction or classified according to angle deviation, and their courtship behavior was video-recorded for one hour or extended to the end of copulation if they were copulating at the end of the one-hour period, using an HC-V550M video camera (Panasonic). The copulation success rate was the percentage of males that mounted a female continuously for more than one minute during the one-hour courtship assay. To facilitate this analysis, we wrote image analysis software, in which the outlines of flies were obtained by the binarization of video images, and these images were arranged in chronological order (see video analysis program below). Using this analysis, we could efficiently detect when flies were copulating, although it was not sensitive enough to define each step of the courtship. This image analysis software also enabled us to observe the video image that corresponded to every time point of the processed data, which allowed us to confirm successful copulation from snapshots of the video data. After taking the video, the males were recovered, their genitalia were photographed, and the angle deviation was measured (VB-7010, Keyence).
Courtship initiation was defined as the beginning of male wing vibration23. Courtship latency was defined as the period of time from the observation start time to the beginning of the first wing vibration (courtship initiation)23. Males that did not start the wing vibration were excluded from this analysis, although there were very few of them (Supplementary Fig. S1). The courtship index (CI) was calculated as the percentage of time that was spent on the courtship, including orientation, tapping, following, wing extension and vibration, licking, and attempted copulation for 10 minutes, as described previously19,20,22,24,25. We used a stopwatch to record the total time in which a male engaged in courtship activity from video-recordings obtained as described above21,25. Males that did not copulate for 10 minutes were excluded from this analysis, as described previously22. The percentage of males that initiated courtship is shown in Supplementary Fig. S1. The reproduction success rate represents the percentage of males that yielded offspring after the one-hour courtship analysis. After the courtship analysis, each female was recovered, separately maintained in standard medium for five days, and discarded. The vials were incubated for another five days to determine whether offspring were present.
Backcross to generate the control w 1118 line
As a control for the courtship assays, we used w1118 hemizygous males, because the Myo31DFK2 line also carries w1118 12. To make the genetic background as uniform as possible, we backcrossed w1118 with w1118/Y; Myo31DFK2 five times. To distinguish between wild-type Myo31DF and the Myo31DFK2 mutation, we conducted genomic PCR. At every generation, a single pair in which the female could be homozygous or heterozygous for Myo31DFK2 was recovered. Genomic DNA was extracted and used as a template for PCR to check the genotype using two sets of primers.
The wild-type Myo31DF gene was amplified by a set of forward and reverse primers (5′- TGCAATGCCTTCAAGACCC -3′ and 5′- GCCGTAAATATCCAGCACTCC-3′) that yielded a 472-bp product. The PCR reaction was conducted according to the manufacturer’s instructions (Ex Taq, Takara, Japan), with modifications of the cycle number from 30 to 34 and annealing temperature from 55 °C to 52 °C.
The genotype of the final w1118 line, which carried the wild-type Myo31DF gene, was confirmed by the same PCR reaction and subsequent analysis. The wild-type Myo31DF locus was amplified by a set of forward and reverse primers (5′-TGGGAACACGTAATGCGTAA-3′ and 5′-TCTGCACTCGATTTGTCAGG-3′) that yielded a 484-bp product. To exclude the line with the Myo31DFK2 mutation, this gene was amplified by a set of forward and reverse primers (5′-TAATCCGCCGCAAACGAAAC-3′ and 5′-TTCGTTCTTTTCGTCGGCCA-3′) that yielded a 431-bp product. In both experiments, the PCR reactions were conducted according to the manufacturer’s instructions (Ex Taq, Takara, Japan), with modification of the cycle number from 30 to 34. In PCR reactions for the wild-type Myo31DF and Myo31DFK2 alleles, the annealing temperature was 50 and 52 °C, respectively. A w1118 line that carried only the wild-type Myo31DF and not the Myo31DFK2 mutation was established and used as a control in the courtship assays.
Video analysis program
To facilitate the courtship assays, we generated a program for processing the images from the video data. A time code was inserted into each video image (Fig. 4b), and then images were collected every 10 seconds. The collected images were binarized, and the shapes of the flies were automatically selected (Fig. 4c). The selected shapes were stacked and arranged in 3D space along a time line (Fig. 4d). To identify time points on the 3D image, a moving bar was inserted, which was synchronized with the 2D images (yellow lines in Fig. 4d). Using the slide bar indicated by these yellow lines on Maya version 2018 (Autodesk, San Rafael, CA), the 2D (video image) and 3D constructed images could be correspondingly identified at any time point (Fig. 4d and Supplementary Fig. S2). With this system, the start and end points of copulation could be quickly identified. After identifying these time points from the 3D constructed images, we confirmed them using the video images.
Statistical analysis
Statistical analyses and graph constructions were performed in R 3.3.1 (R Foundation for Statistical Computing, Vienna, Austria). To evaluate the statistical significance of the differences in reproduction success rates, Fisher’s exact test was performed. In the courtship latency analysis, a nonparametric Kruskal-Wallis test was performed, and the results were evaluated by a Steel-Dwass test to judge the significance of the difference between each group. The CI was analyzed with One-Way Analysis of Variance (ANOVA) and evaluated by Tukey test to judge the significance of the difference between each group. A P-value < 0.05 was considered to indicate a significant difference.
References
Lamichhaney, S. et al. Evolution of Darwin’s finches and their beaks revealed by genome sequencing. Nature 518, 371–375, https://doi.org/10.1038/nature14181 (2015).
Lamichhaney, S. et al. A beak size locus in Darwin’s finches facilitated character displacement during a drought. Science 352, 470–474, https://doi.org/10.1126/science.aad8786 (2016).
Grant, P. R. & Grant, B. R. Darwin’s finches. Curr. Biol. 15, 614–615, https://doi.org/10.1016/j.cub.2005.08.014 (2005).
Huber, B. A., Sinclair, B. J. & Schmitt, M. The evolution of asymmetric genitalia in spiders and insects. Biol. Rev. 82, 647–698, https://doi.org/10.1111/j.1469-185x.2007.00029.x (2007).
Huber, B. A. Mating positions and the evolution of asymmetric insect genitalia. Genetica 138, 19–25, https://doi.org/10.1007/s10709-008-9339-6 (2010).
Bickel, D. Sex with a twist in the tail. New Scientist 127, 34–37 (1990).
Suzanne, M. et al. Coupling of apoptosis and L/R patterning controls stepwise organ looping. Curr. Biol. 20, 1773–1778, https://doi.org/10.1016/j.cub.2010.08.056 (2010).
Chevone, B. I. & Richards, A. G. Ultrastructure of the atypic muscles associated with terminalial inversion in male Aedes aegypti (L). Biol. Bull. 151, 283–296, https://doi.org/10.2307/1540661 (1976).
Kuranaga, E. et al. Apoptosis controls the speed of looping morphogenesis in Drosophila male terminalia. Development 138, 1493–1499, https://doi.org/10.1242/dev.058958 (2011).
Kim, S., Trocke, S. & Sim, C. Comparative studies of stenogamous behaviour in the mosquito Culex pipiens complex. Med. Vet. Entomol., https://doi.org/10.1111/mve.12309 (2018).
Roth, L. M. A study of mosquito behavior. An experimental laboratory study of the sexual behavior of Aedes aegypti (Linnaeus). Am. Midl. Nat. 40, 265–352, https://doi.org/10.2307/2421604 (1948).
Speder, P., Adam, G. & Noselli, S. Type ID unconventional myosin controls left-right asymmetry in Drosophila. Nature 440, 803–807, https://doi.org/10.1038/nature04623 (2006).
Hozumi, S. et al. An unconventional myosin in Drosophila reverses the default handedness in visceral organs. Nature 440, 798–802, https://doi.org/10.1038/nature04625 (2006).
Okumura, T. et al. Class I myosins have overlapping and specialized functions in left-right asymmetric development in Drosophila. Genetics 199, 1183–U1493, https://doi.org/10.1534/genetics.115.174698 (2015).
Inaki, M., Sasamura, T. & Matsuno, K. Cell chirality drives left-right asymmetric morphogenesis. Front Cell Dev. Biol. 6, 34, https://doi.org/10.3389/fcell.2018.00034 (2018).
Inaki, M. et al. Chiral cell sliding drives left-right asymmetric organ twisting. Elife 7, https://doi.org/10.7554/eLife.32506 (2018).
Packard, M. et al. Nucleus to synapse Nesprin1 railroad tracks direct synapse maturation through RNA localization. Neuron 86, 1015–1028, https://doi.org/10.1016/j.neuron.2015.04.006 (2015).
Amcheslavsky, A. et al. Plasma membrane localization of apoptotic caspases for non-apoptotic functions. Dev. Cell 45, 450–464.e453, https://doi.org/10.1016/j.devcel.2018.04.020 (2018).
Ito, H. et al. Fruitless recruits two antagonistic chromatin factors to establish single-neuron sexual dimorphism. Cell 149, 1327–1338, https://doi.org/10.1016/j.cell.2012.04.025 (2012).
Yamamoto, D. & Koganezawa, M. Genes and circuits of courtship behaviour in Drosophila males. Nat. Rev. Neurosci. 14, 681–692, https://doi.org/10.1038/nrn3567 (2013).
Krstic, D., Boll, W. & Noll, M. Influence of the white locus on the courtship behavior of Drosophila males. Plos One 8, 8, https://doi.org/10.1371/journal.pone.0077904 (2013).
Krstic, D., Boll, W. & Noll, M. Sensory integration regulating male courtship behavior in Drosophila. Plos One 4, 13, https://doi.org/10.1371/journal.pone.0004457 (2009).
Luu, P., Zaki, S. A., Tran, D. H. & French, R. L. A novel gene controlling the timing of courtship initiation in Drosophila melanogaster. Genetics 202, 1043–1053, https://doi.org/10.1534/genetics.115.183061 (2016).
Siegel, R. W. & Hall, J. C. Conditioned responses in courtship behavior of normal and mutant. Drosophila. Proc. Natl. Acad. Sci. USA 76, 3430–3434 (1979).
McBride, S. M. et al. Mushroom body ablation impairs short-term memory and long-term memory of courtship conditioning in Drosophila melanogaster. Neuron 24, 967–977 (1999).
Acebes, A., Cobb, M. & Ferveur, J. F. Species-specific effects of single sensillum ablation on mating position in. Drosophila. J. Exp. Biol. 206, 3095–3100, https://doi.org/10.1242/jeb.00522 (2003).
Kotyk, M. & Varadínová, Z. Wing reduction influences male mating success but not female fitness in cockroaches. Sci. Rep. 7, 2367, https://doi.org/10.1038/s41598-017-02647-7 (2017).
Preston-Mafham, K. G. Courtship and mating in Empis (Xanthempis) trigramma Meig., E-tessellata F. and E. (Polyblepharis) opaca F. (Diptera: Empididae) and the possible implications of ‘cheating’ behaviour. J. Zool. 247, 239–246, https://doi.org/10.1111/j.1469-7998.1999.tb00987.x (1999).
Hazelrigg, T., Levis, R. & Rubin, G. M. Transformation of white locus DNA in Drosophila: Dosage compensation, zeste interaction, and position effects. Cell 36, 469–481, https://doi.org/10.1016/0092-8674(84)90240-X (1984).
Kurkulos, M., Weinberg, J. M., Pepling, M. E. & Mount, S. M. Polyadenylylation in copia requires unusually distant upstream sequences. Proc. Nat. Acad. Sci. USA 88, 3038–3042 (1991).
Perry, J. C. et al. Experimental evolution under hyper-promiscuity in Drosophila melanogaster. BMC Evol. Biol. 16, 131 (2016).
Acknowledgements
We are grateful to Dr. S. Noselli for providing the Myo31DFK2 line. We also thank Drs D. Yamamoto and T. Higuchi for valuable advice for recording the mating videos. We thank members of the Matsuno laboratory for critical comments and discussions. This work was supported by the Innovative Educational Research Fund and JSPS KAKENHI Grant Number JP16J01904.
Author information
Authors and Affiliations
Contributions
M.I. designed and performed the experiments, analyzed the data, and wrote the manuscript. D.S. developed the software for analyzing the mating videos. Y.L. prepared the figures. K.M. designed the experiments, analyzed and interpreted data, and wrote the manuscript.
Corresponding author
Ethics declarations
Competing Interests
The authors declare no competing interests.
Additional information
Publisher’s note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Inatomi, M., Shin, D., Lai, YT. et al. Proper direction of male genitalia is prerequisite for copulation in Drosophila, implying cooperative evolution between genitalia rotation and mating behavior. Sci Rep 9, 210 (2019). https://doi.org/10.1038/s41598-018-36301-7
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-018-36301-7
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
