
Abstract
The main objective of this study was to evaluate the functional traits regarding bioturbation activity and its influence in the nutrient cycling of the native clam species Ruditapes decussatus and the invasive species Ruditapes philippinarum in Ria de Aveiro lagoon. Presently, these species live in sympatry and the impact of the invasive species was evaluated under controlled microcosmos setting, through combined/manipulated ratios of both species, including monospecific scenarios and a control without bivalves. Bioturbation intensity was measured by maximum, median and mean mix depth of particle redistribution, as well as by Surface Boundary Roughness (SBR), using time-lapse fluorescent sediment profile imaging (f-SPI) analysis, through the use of luminophores. Water nutrient concentrations (NH4-N, NOx-N and PO4-P) were also evaluated. This study showed that there were no significant differences in the maximum, median and mean mix depth of particle redistribution, SBR and water nutrient concentrations between the different ratios of clam species tested. Significant differences were only recorded between the control treatment (no bivalves) and those with bivalves. Thus, according to the present work, in a scenario of potential replacement of the native species by the invasive species, no significant differences are anticipated in short- and long-term regarding the tested functional traits.
Similar content being viewed by others

Introduction
Coastal ecosystems, like coastal lagoons are complex ecological and socio-economic systems rich in biodiversity and home to a diverse array of habitats and species. Thus, providing numerous economic and societal benefits. Many of these valuable ecosystems are at risk of being irreversibly damaged by invasive species1,2. This pressure threatens the ecological functions of these ecosystems and therefore their provision of ecosystem services. Ria de Aveiro is a shallow coastal lagoon, located in the western Atlantic coast of Portugal, characterized by a wide range of habitats with high ecological importance, natural resources of high economic interest and an important ecological diversity, able to provide a wide range of ecosystem services3. Bivalves are among the most important commercial resources exploited in this ecosystem, namely the native species Grooved Carpet Shell clam Ruditapes decussatus (Linnaeus, 1758) and, since 2011, the invasive species Manila clam Ruditapes philippinarum (Adams & Reeve, 1850). Full description of each species is presented as supplementary material (Supplementary Table S14,5,6,7,8,9,10).
Ruditapes decussatus plays a key role on shell fishing activities and aquaculture due to its high commercial value, about three times that of R. philippinarum11. In 2012, the national aquaculture production of R. decussatus was of 2 320 tonnes11, being Ria de Aveiro and Ria Formosa the main production areas for this species12. At an European level, during the last decades, recruitment failures and excessive pressure on the capture of juveniles on natural banks and severe clam mortalities lead to an important decrease in R. decussatus production13,14. This situation resulted in the introduction of the non-indigenous R. philippinarum. This invasive species originates from the Indo-Pacific region was introduced in Europe for shell fishing activities and aquaculture purposes at the beginning of the 1970s13. It was initially introduced in France, followed by England, Spain, Italy and Portugal13,15,16. In these countries its habitat overlaps with that of R. decussatus, a species which it successfully out-competes, not only in aquaculture farms but also on natural conditions17. In Arcachon Bay (France), Venice Lagoon (Italy) and the western coast of the Cotentin Peninsula (English Channel), the invasive species has clearly supplanted the native R. decussatus1,18,19,20. In 2016, the European production of R. philippinarum was almost seven times higher than that of R. decussatus, corresponding to 5 389 and 35 436 tonnes, respectively11.
In Ria de Aveiro, according to a national management oriented project on bivalves, named Gepeto21, the abundance and dispersion of R. decussatus have been decreasing from 2006/7 to 2013. In a sampling campaign carried out in 2013, 17.0% of the biomass of all bivalves captured in Ria de Aveiro was R. philippinarum and only 1.0% corresponded to R. decussatus. In less than 2 years the invasive species became the second most important bivalve species in commercial terms in this ecosystem, with populations mostly dominated by adult specimens larger than the minimum catch size (40 mm). Stocks of R. decussatus are currently depleted and when the yields recorded from previous campaigns are compared, there was an overall decrease of 67.0% in its biomass and 76.6% in its abundance21. Although its commercial value is lower when compared with the native clams, R. philippinarum constitutes more than 90.0% of the yields of the two species in the European scenario22.
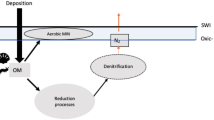
Beyond their socio-economic importance, bivalves are major bioturbators of the sediment in marine and estuarine environments23. One of the consequences of the introduction of non-indigenous species are shifts in the structure and functioning of ecosystems, as similar species may share physiological traits but that does not mean that they will play similar functional roles24. Thus, in spite of the similarities between R. decussatus and R. philippinarum, the replacement of one species by the other can potentially change ecosystem structure and functioning. Sedimentary geochemical processes are influenced by benthic invertebrates through bioturbation, which consists in the mixing of unconsolidated sediments and particulate materials during foraging, feeding and burrowing activities, as well as in the enhancement of pore water and solute advection during burrow ventilation25 which, consequently influences the concentration of oxygen, pH and redox potential1, sediment granulometry26, macrofauna diversity27, bacterial activity and composition28,29 and nutrient cycling30,31. Nutrient (NH4-N, NOx-N and PO4-P) concentrations in the water are commonly used to understand ecosystem functioning and numerous studies have described the influence of bivalve aquaculture on various components of the environment, including nutrient cycling32,33.
The study of bioturbation is therefore crucial to evaluate the importance of key processes in ecosystems34,35. An ecosystem structure and function, may suffer significant changes due to the replacement of its native species by invasive species36. In order to evaluate the potential consequences of an eventual replacement of a native clam species by an invasive species, the present study evaluated the functional traits regarding bioturbation activity and its influence in the nutrient cycling of two clam species that occur in sympatry in Ria de Aveiro: the native species R. decussatus and the invasive species R. philippinarum. The impact of the invasive species was evaluated under controlled microcosmos conditions, through combined/manipulated ratios of R. philippinarum and R. decussatus tested for 21 days. Results will be discussed taking into account sediment luminophores displacement and nutrients concentration in the water column due to bioturbation by R. decussatus and R. philippinarum under the tested ratios versus the control conditions without bivalves.
Results
Bioturbation
Sediment used in the aquaria was classified as muddy fine sand (ϕ = 2) with 16.3% of fines and an average organic matter content of 5.6 ± 0.4%.
The f-SPILmax (maximum extent of mixing over the long-term), f-SPILmed (typical short-term depth of mixing) and f-SPILmean (time-dependent indication of mixing) mixed depth of particle redistribution recorded for the luminophores following the bioturbation process of bivalves, taking into account different ratios of R. decussatus (Rd) and R. philippinarum (Rp) tested, are shown in Fig. 1A–C, respectively.

Maximum (f-SPILmax) (A), median (f-SPILmed) (B) and mean (f-SPILmean) (C) mixed depth of particle redistribution (mm, mean ± SD) in the control (C) aquaria and in the aquaria with different treatments of Ruditapes decussatus (Rd) and Ruditapes philippinarum (Rp) at D21.
Despite the differences recorded in the f-SPILmax between the different treatments, these were not statistically significant (pseudo-F = 1.755; p = 0.1782). However, significant differences were found in the f-SPILmax between the control and all the treatments (Table 1). The f-SPILmax recorded for luminophores was lower in the control aquaria (7.8 ± 0.7 mm). Despite there were no significant differences between treatments, the treatment 2 Rd + 6Rp presented the highest value of f-SPILmax (24.0 ± 7.6 mm), while the lowest value corresponded to the treatment only using R. decussatus (8 Rd + 0Rp) (17.0 ± 3.0 mm). It was possible to observe an increase of the f-SPILmax when individuals of both species cohabitated the same aquarium (treatments 6 Rd + 2Rp, 4 Rd + 4Rp, 2 Rd + 6Rp) and with the increasing number of R. philippinarum (Fig. 1A).
The f-SPILmed between different treatments did not show significant differences (pseudo-F = 1.059; p = 0.3977) but significant differences were found in the f-SPILmed between the control and all the treatments as is shown in Table 1. The highest and lowest f-SPILmed values (Rp − 10.0 ± 3.2 mm; Rd − 5.9 ± 1.4 mm) were observed in aquaria stocked solely with R. philippinarum and R. decussatus, respectively. The control presented lower f-SPILmed values comparatively with the aquaria stocked with bivalves (0.3 ± 0.4 mm). In general, there was a tendency for increase of f-SPILmed with the increasing number of specimens of R. philippinarum in the treatment (Fig. 1B).
The statistical analysis allowed to test the influence of the ratio of both species on the f-SPILmean reached by the luminophores, representing the depth in which the organisms were found during most of the study period, and no significant differences were found between treatments (pseudo-F = 1.323; p = 0.2891). In turn, there were significant differences in the luminophores f-SPILmean between the control and the aquaria with bivalves (Table 1). Considering the aquaria in which only one species was present (8 Rd + 0Rp; 0 Rd + 8Rp), there was a tendency for the f-SPILmean values to be lower comparatively with the aquaria in which both species were present (Fig. 1C). Thus, the lowest values of f-SPILmean were recorded in the monospecific treatment 8 Rd + 0Rp (5.9 ± 1.4 mm) while the highest values were registered in the treatment 2 Rd + 6Rp (10.0 ± 3.2 mm). Considering the total period of the study, aquaria with both species and with highest number of R. philippinarum tended to reach higher average depths. Control aquaria displayed lower f-SPILmean values than those with bivalves.
The SBR (surface boundary roughness) calculated for the aquaria with bivalves at the end of the experiment is shown in (Fig. 2). The results obtained in the statistical analyses showed no significant differences in the SBR values between the different treatments of R. decussatus and R. philippinarum (pseudo-F = 0.830; p = 0.5237). The average SBR was lower in the aquaria with 6 Rd + 2Rp (7.0 ± 4.0 mm) and higher in the aquaria with the opposite treatment 2 Rd + 6Rp (13.0 ± 4.0 mm). In the treatments with both species, the SBR values tended to increase with the increasing number of R. philippinarum (Fig. 2).

Surface boundary roughness (SBR; mm, mean ± SD) considering different treatments of Ruditapes decussatus (Rd) and Ruditapes philippinarum (Rp) at D21.
Water physicochemical characterization and nutrients concentration
Water temperature, pH, salinity and oxygen concentration of all the aquaria at D−1 (day before bivalves’ introduction (D0)) and D21 (end of the experiment) are shown in Table 2. Temperature ranged from 16.9 ± 0.2 to 17.4 ± 0.2 °C, pH from 7.8 ± 0.3 to 8.2 ± 0.1 and oxygen concentration from 8.5 ± 0.1 to 9.5 ± 0.1 mg L−1. Salinity stayed stable at 33 ± 0.0 through D−1 to D21 in all the aquaria.
Figure 3A displays the principal components ordination analysis of the average concentration of ammonium (NH4-N), oxidised form of dissolved inorganic nitrogen (NOx-N) and phosphate (PO4-P) in the control aquaria and those stocked with different treatments of R. decussatus and R. philippinarum at days 0, 2, 5, 8, 12, 16 and 21. Axis 1 explained 91.0% of the total variance and showed an increasing concentration gradient of NH4-N and PO4-P from D0 to D21 in the aquaria with bivalves. Control was mainly characterized by comparatively higher concentrations of NOx-N.

Principal component analysis (A) and graphic bars (B) based on the average concentrations (mg/L) of ammonium (NH4-N), oxidised form of dissolved inorganic nitrogen (NOx-N) and phosphate (PO4-P) in the control (C) and different treatments of Ruditapes decussatus (Rd) and Ruditapes philippinarum (Rp) since the beginning until the end of the experiment (D0, D2, D5, D8, D12, D16 and D21).
Average concentrations of NH4-N, NOx-N and PO4-P in the control aquaria and those stocked with different treatments of R. decussatus and R. philippinarum at different sampling days are shown in Fig. 3B–D. The statistical analysis showed that NH4-N concentrations were not significantly different between treatments (pseudo-F = 2.221; p = 0.1016). Significant differences were only found between the control and the aquaria with bivalves (Table 3). In general, NH4-N concentrations increased from D0 to D21, tended to be higher in aquaria with 8 Rd + 0Rp (11.18 mg L−1 at D21) and decreasing in aquaria with a lower number of individuals of this species (Fig. 3B). In the control, NH4-N concentrations were lower, comparatively with the other treatments, achieving a maximum concentration of 3.56 mg L−1 at D21. In aquaria with 0 Rd + 8Rp the NH4-N concentration was lower at the end of the experiment, comparatively with other treatments, reaching a maximum concentration of 9.24 mg L−1 (Fig. 3B).
For NOx-N, significant differences were only found in their concentrations at D21 between the control and the aquaria with different treatments (Table 3). Between different treatments, concentrations of NOx-N were not significantly different (pseudo-F = 0.358; p = 0.8326). As is shown in Fig. 3C, the average concentration of NOx-N was higher in the control than in the aquaria with bivalves, increasing from 0.17 mg L−1 at D0 to 1.42 mg L−1 at D21. In the aquaria with bivalves, NOx-N concentrations did not show any particular trend for none of the bivalves’ ratios (Fig. 3C).
The statistical analysis of PO4-P concentrations showed that there were no significant differences in PO4-P concentrations between treatments (pseudo-F = 1.615; p = 0.2100) but there were significant differences in PO4-P concentrations between the control and the aquaria with different treatments (Table 3). The average concentrations of PO4-P tended to increase along time, with the control aquaria showing the lowest concentrations that ranged from 0.10 mg L−1 at D0 to 0.32 at D21 (Fig. 3D). At the beginning of the experiment, PO4-P concentrations for each ratio were of 0.16 mg L−1 for 8 Rd + 0Rp, 0.19 mg L−1 for 6 Rd + 2Rp, 0.13 mg L−1 for 4 Rd + 4Rp and 0.12 mg L−1 for treatments 2 Rd + 6Rp and 0 Rd + 8Rp. At the end of the experiment, PO4-P concentrations were higher in the aquaria with higher number of specimens of R. decussatus (8 Rd + 0Rp − 0.72 mg L−1 and 6 Rd + 2Rp − 0.55 mg L−1) decreasing in the aquaria with equal or higher number of R. philippinarum (4 Rd + 4Rp − 0.49 mg L−1; 2 Rd + 6Rp − 0.42 mg L−1; 0 Rd + 8Rp − 0.49 mg L−1).
Discussion
Bivalves are key species in many habitats due to their influence on habitat engineering and nutrient cycling. While the borrow depth of clams differs from species to species, in general, clams could be considered as shallow-burrowers rather than deep-burrowers, as they tend to preferentially move along and bioturbate the uppermost sediment layers23. The depths considered in this study correspond to the depth of the sediment that clams remobilize due to their bioturbation process, being an indicator of their intervention in the ecosystems structure1. Results obtained in this study allowed to understand the remobilization behaviour of both clam species, suggesting that, in the short term (f-SPILmed), Ruditapes philippinarum seems to contribute more to sediments remobilization in the superficial layer. At long-term (f-SPILmax), and when both species occur in sympatry, clams can reach greater depths due to their highest bioturbation intensity. In addition, this trend also occurred in the mean depth (f-SPILmean), since the depth at which clams were found in the sediment, during most part of the study period, was greater in aquaria with both species and with higher number of individuals of the invasive species R. philipinarum. This finding may be explained by the fact that specimens of R. decussatus can bury up to 120 mm deep6 while those of R. philippinarum can only reach depths of 80 mm7, thus indicating that the native species has the ability to bury at a greater depth. In this way, it is possible that, in the presence of higher densities of R. philippinarum the native species tends to bury deeper in order to avoid the invasive species. According to Sobral37, the immediate response of R. decussatus to a physical stimuli of sediment movement is a decline in the clearance rate and even the periodic valve closure, which reduces the ability of individuals to maintain their feeding activity and physiological condition, reinforcing the effect that the invasive species can have in the native clam species. However, according to our results, the remobilization depths reached by both species, resulting from their bioturbation activity, were not statistically different. Nonetheless, both species produce a significant effect on sediment remobilization, evidenced by the existence of significant differences between the control aquaria with no bivalves and those stocked with clams. This trend is in accordance with the studies by Sgro et al.38 and Spencer et al.39, in which the amount of remobilized sediment was higher in areas exposed to clams bioturbation, revealing that areas with higher densities of clams are characterized by more unstable sediments. The biological modifications carried out by bioturbator organisms increase water-sediment interface roughness and decrease the physical stability of the sediment, which may induce stress in other organisms40. In this way, the analysis of the surface boundary roughness (SBR) contributes to the identification of the bioturbation impacts on surface sediment. High values of SBR, resulting from increased fluid transport by the bioturbation activity near the water-sediment interface, reflect a critical zone in which intense physical and mechanical reactions of sediment changes (diagenesis) are promoted41. However, according to the results obtained in this study, there were no significant differences in SBR values between different ratios of R. decussatus and R. philippinarum.
In an ecosystem, the existence of a species is conditioned by all the environmental processes that occur there, and its existence or disappearance also influences the ecosystem itself, which reveals the network of complex interactions between organisms and their habitat42. High densities of clams affect the environment due to the excretion of nitrogenous compounds and the enrichment of sediment with organic matter (faeces and pseudofaeces)32,43. High bivalves’ densities are synonymous of high nutrient cycling rates due to bioturbation, since this process intervenes in the nutrient cycle and carbon flux in sediments34. The performance of bioturbation processes by a species is favourable to its survival and to the ecosystem itself, but may become unfavourable and of concern if its intensity inhibits the same process in other species and ultimately threatens their survival through a severe sediment destabilization and resuspension increasing38. Nevertheless, in the particular case of this study, considering the ratios of clams tested (8 Rd + 0Rp, 6 Rd + 2Rp, 4 Rd + 4Rp, 2 Rd + 6Rp, 0 Rd + 8Rp), there were no significant differences in water nutrient concentrations.
Water concentrations of NH4-N and PO4-P registered in this study, carried out in microcosms under controlled conditions, were much higher than those observed in natural conditions in Ria de Aveiro. According to Lopes et al.44, in a study carried out in 2001 and 2012 in Ria de Aveiro, the highest concentrations of NH4-N registered were of 0.15 mg L−1 in winter 2001 and 0.05 mg L−1 in summer 2012, and the highest PO4-P concentrations were registered in winter 2001 (0.09 mg L−1) and in summer 2012 (0.04 mg L−1). Differences in NH4-N and PO4-P concentrations between microcosms and natural conditions could be explained by the fact that, the sediment was rich in organic matter (5.6%) and that during the experimental period, water in the aquariums was not replaced, thus contributing for a potential build-up of nutrients due to mineralisation processes30. In the case of natural conditions, water is daily renewed. The main forcing agent driving water circulation in the lagoon is the tide, which is mesotidal and characterized by semidiurnal tides, presenting an average tidal amplitude at the inlet of 2 m, and amplitudes of 0.6 m in neap tides and 3.2 m in spring tides45. Despite the highest concentrations of NH4-N and PO4-P, no mortality was registered, revealing the capacity of tolerance of both species to high concentrations of these nutrients.
However, the same was not observed in Ria Formosa (south Portugal). In this case, a high mortality of R. decussatus was registered especially in the intertidal mudflats, which could be related with high summer temperatures at low tide46,47, resulting in the accumulation of ammonia in the sediment surface due to the enhancement of mineralization. In fact, Falcão & Vale48 found a sharp release rate of ammonium from sediment in the presence of R. decussatus in Ria Formosa. In this scenario, when tide rises and R. decussatus starts to feed, this species is exposed to high concentrations of ammonium (>1.40 mg L−1) which will decrease with the mixing in the sediment-water interface. The fact that this species exhibits high mortality rates in Ria Formosa when exposed to much lower ammonium concentrations than those observed in the microcosm assay, where no mortality was registered, could be related with the stress to which organisms are exposed during low tide (higher temperature and, consequently, higher salinity). In our study, specimens were always under the same salinity and temperature conditions and always experiencing a “high tide scenario”, as tide was not simulated in the microcosm. Other study, developed by Bartoli et al.49 in Sacca di Goro shallow coastal lagoon in Italy, showed that R. philippinarum excrete large quantities of ammonium, which may represent about 90.0% of the total DIN released from sediments with clams, as compared to only 30.0% of the total DIN released from sediments without clams. These results are in accordance with the results obtained in this study in which NH4-N concentrations were not significantly different between treatments but were significantly different between the control and the aquaria with bivalves.
In what concerns to PO4-P, it is known that sediments act as a sink or source of phosphate, being sediment physicochemical properties (e.g. granulometry, organic matter content, sediments exchangeable P content, salinity), water temperature and hydrodynamics the main environmental variables influencing the fluxes between sediment and water column50,51. Thus, the highest concentrations of PO4-P in the aquaria with higher densities of R. decussatus could be explained by its lower sediment remobilization. Lower hydrodynamics, promotes a higher settlement of organic rich fine particles which adsorb phosphate and decrease the vertical exchanges with the water column (e.g. Mortimer et al.52, Lillebø et al.53).
Contrarily to other nutrients, NOx-N concentrations in the microcosm (ranged from 0.34 mg L−1 to 1.42 mg L−1) were in the same range to that observed in Ria de Aveiro in 2001 and 2012 (ranged from 0.01 mg L−1 to 2.33 mg L−1) (personal info.). Bivalves’ bioturbation minimizes the potential negative effects of excessive biodeposition, as particle and solute transport by bioturbation significantly influences rates and pathways of organic matter mineralization, particularly by increasing oxygen penetration depth in sediments. Thus, contrarily to what was observed, it was expected that NOx-N concentrations in treatments with bivalves were higher than those in the control52. In the presence of bivalves, sediment was more aerated due to a higher remobilization, thus enabling the bacterial activity in the sediment, mediating nitrification by oxidising NH4 to NOx, using O2.
Overall, this study allowed to conclude that, based on the ratios tested, will not occur significant changes in sediment bioturbation, at short- and also at long-term, and nutrient cycling in a scenario of potential replacement of a native and an invasive clam of the genus Ruditapes occurring in sympatry in a coastal lagoon.
Material and Methods
Natural ecosystem under study
Ria de Aveiro is located on the northwestern coast of Portugal, between 40°38′N and 40o 57′N. This temperate system is a shallow coastal lagoon characterized by intertidal mud and sand flats, salt marshes and islands, forming four main channels, Mira, Ílhavo, Espinheiro and S. Jacinto-Ovar. The only connection with the Atlantic Ocean is made through an artificial inlet constructed in 1808 (1.3 km length; 350 m wide; 20 m depth). Ria de Aveiro is mesotidal, characterized by semidiurnal tides. It exhibits a longitudinal gradient of salinity from about 0 at upstream areas, to about 36 at the Ocean boundary.
Microcosm setting
The microcosm set-up was designed to simulate different degrees of sympatry occurrence in Ria de Aveiro of a native and an invasive clam of the genus Ruditapes, and run with specimens of R. decussatus (Rd) and R. philippinarum (Rp) from the lagoon. Taking into account the large number of specimens necessary, the convenience of being of the same size and the effort associated with their capture, post-depuration clams were acquired in the local depuration centre that receives the specimens collected by local fishermen. Shell length of the individuals used in this experiment was measured with a Vernier calliper to the nearest 0.05 mm. The average length was of 44.8 ± 1.8 mm for Rd and of 41.8 ± 2.3 mm for Rp. Specimens were transported to the laboratory and placed in containers with synthetic saltwater (prepared by mixing Red Sea Coral Pro Salt (Germany) and freshwater purified by a reverse osmosis unit) without sediment, with a salinity of 33 during 72 hours. This salinity was stablished based on the distribution areas of both species in Ria the Aveiro21,54,55. No food was added during acclimation and continuous aeration was kept by air-bubbling the water with air-stones. The sediment used in the experiment was collected in Mira channel, homogenized and resident macrofauna was visually detected and removed. Five sediment sub-samples were analysed to determine grain-size and organic matter content. Sediment was distributed along 30 glass aquaria (150 × 150 × 500 mm), creating a 150 mm column about 1/3 of the height of the aquarium and filled with saltwater (about 2/3 of the height of the aquarium) and left to settle for 48 h. Then, all saltwater was replaced and left to stabilize for 24 h before the introduction of bivalves. Aquaria were gently aerated by air-bubbling using an air pump (Aqua Medic Mistral 4000, 4000 l/h) connected to a silicon tube (ø = 4.00 mm) in which were connected silicon capillary tubes (ø = 0.84 mm) that aerated each aquarium. Aquaria were randomly distributed by the bench and exposed to natural light conditions with a 10 h day: 14 h dark photoperiod. Specimens of Rd and Rp were placed in the aquaria at different ratios (8 Rd + 0Rp, 6 Rd + 2Rp, 4 Rd + 4Rp, 2 Rd + 6Rp, 0 Rd + 8Rp), with 5 replicates per each one of them, totalling 25 aquaria. Five aquaria were kept without bivalves (0 Rd + 0Rp) and used as control. All aquaria were covered with parafilm to minimize evaporation and hence pronounced salinity shifts. In order to avoid the initial activity associated with bivalves establishment and borrowing, 40 g of fluorescent-dyed sediment particles (luminophores: 125–250 µm diameter, pink colour; Brian Clegg Ltd., UK) were distributed in the front face of each aquaria 24 h after the introduction of bivalves (D0). The same quantity of luminophores was added to all the aquaria during the experiment at days 0, 4, 11 and 17 to allow the monitoring of bioturbation. The water of each container was not renewed and neither sediment nor food were added. The temperature in the room ranged from 16.4 °C at D−1 to 17.9 °C at D21.
Bioturbation
Particle reworking was measured non-invasively using fluorescent sediment profile imaging (f-SPI56) and luminophores. To photograph the aquaria, a black box was used, illuminated with four actinic fluorescent lamps (22.000 K, 80 W (λ = 400–450 nm), Red-Sea) (Fig. 4). Images were captured once a day, using a SONY Cyber-shot G (14MP; aperture f = 5.6) camera. Considering the main objective of this study, solely the images collected at D21 were analysed. Images obtained were previously cut according to the internal width of the aquarium (1968 pixels; effective resolution = 0.07 mm per pixel) and then converted to a red-blue-green (RGB) stack and saved with JPEG compression (Joint Photographic Experts Group (Fig. 5)). Images were then analysed using a custom-made plugin that runs within ImageJ (Version 1.48c), a java-based public domain program developed at the US National Institutes of Health (available at http://rsb.info.nih.gov/ij/index.html). In the particular case of this study, as we added luminophores several times along the experiment, in the control aquaria, luminophores created a superficial layer which was not remobilized. For this reason, in the control aquaria the interface considered was between the sediment and the layer of luminophores. As all luminophores were remobilized in the aquaria stocked with bivalves, the sediment-water interface was considered for analysis. At D21 the following parameters were determined: mean (f-SPILmean, time-dependent indication of mixing), median (f-SPILmed, typical short-term depth of mixing), and maximum (f-SPILmax, maximum extent of mixing over the long-term) mixed depth of particle redistribution, following Hale et al.57. In addition, the maximum vertical deviation of the sediment-water interface (upper-lower limit = surface boundary roughness, SBR), which provides an indication of surficial faunal activity, was also determined. SBR was only calculated for aquaria stocked with bivalves, as these were the only ones in which there was a sediment-water interface remobilization.

Particle reworking was measured inside a black box, photographing the pink luminophores exposed to four actinic lights 22000 K Rede-sea 80 W (λ = 400–450 nm).

Image analysis for the quantification of luminophores showing (A) the original image with the sediment appearing in black and luminophore particles in pink, and (B) the processed image, showing the flattened sediment-water interface.
Sediment grain-size was analysed by dry sieving following Dias58. Median and percentage of fines were used to classify the sediment, according to the Wentworth scale59,60. Total organic matter concentration was obtained as described by Kristensen & Anderson61.
Water characteristics and nutrient analyses
Water aliquots (10 ml) were sampled from each aquarium at days 0, 2, 5, 8, 12, 16 and 21 (the volume of water sampled throughout the experiment represented less than 10.0% of the microcosm total volume). Samples were filtered through Whatman GF/C glass-fibre filters and stored at −20 °C until analysis of dissolved inorganic nutrients (ammonium, NH4-N; oxidised form of dissolved inorganic nitrogen, NOx-N; phosphate, PO4-P). The determination of the concentrations of NH4-N and PO4-P was performed following standard methods described in Limnologisk Metodik62. The concentrations of NOx-N were determined using a flow injection system (FIAstar 5000 Analyzer, Höganäs, Sweden), following the Strickland & Parsons63 method. To ensure analytical quality control, calibration curves, using a standard solution, were run at the beginning of the analysis and in parallel with blanks and samples. Water temperature, pH, concentration of dissolved oxygen and salinity were measured using a WTW – pH 330i/set equipped with SenTix® 41; a WTW – cond 3110/set 1 equipped with TetraCon® 325 and a WTW – Oxi 3210/set 2 equipped with CellOx® 325-3.
Statistical analyses
Bioturbation activity and water nutrient concentrations were analysed using a model with one fixed factor (clam ratios) for D21. Aquaria without bivalves were used as a control for analysis, with the exception of the SBR variable, as there was no vertical variation on the superficial layer due to the absence of organisms. The resemblance matrix between samples was obtained using Euclidean distances following a Log(X + 1) transformation. The f-SPILmax data were analysed under the null hypothesis (H0) of no significant differences in the maximum extent of luminophores mixing in the sediment over the long-term considering different treatments of R. decussatus (Rd) and R. philippinarum (Rp). Data concerning f-SPILmed was analysed under the H0 of no significant differences in the luminophores mixing in the sediment over the short-term, considering different treatments of Rd and Rp. Concerning f-SPILmean data, this was analysed under the H0 of no significant differences in the average depth reached by luminophores in the sediment, considering different treatments of Rd and Rp. The SBR analysis was performed under the H0 of no significant differences recorded in SBR determined considering different treatments of Rd and Rp. Water nutrient concentrations was analysed under the H0 of no significant differences in the water nutrient concentrations (NH4-N, NOx-N and PO4-P, separately) considering different treatments of Rd and Rp. All H0 detailed above were tested between the control aquaria and different treatments of Rd and Rp for all the variables previously described, with the exception of SBR. Statistical differences between treatments were tested based on a matrix only with data from aquaria with bivalves, excluding the control. The comparison between control aquaria and those stocked with different treatments of clams was performed separately for each clam treatment. Hypothesis testing was performed by Permutation Multivariate Analysis of Variance64, using the software PRIMER v665, with the add-on PERMANOVA+66. To run the PERMANOVA tests, we considered 9999 Monte Carlo permutations. The pseudo-F values in the main tests and the t-statistic in the pairwise comparisons were evaluated in terms of the significance among levels of the tested factor. Values of p < 0.05 revealed that the groups differed significantly. Water nutrient concentrations of NH4-N, NOx-N and PO4-P were represented in ordination analyses, using a Principal Component Analysis (PCA)64.
References
de Moura Queirós, A., Hiddink, J. G., Johnson, G., Cabral, H. N. & Kaiser, M. J. Context dependence of marine ecosystem engineer invasion impacts on benthic ecosystem functioning. Biol. Invasions 13, 1059–1075 (2011).
Gallardo, B., Clavero, M., Sánchez, M. I. & Vilà, M. Global ecological impacts of invasive species in aquatic ecosystems. Glob. Chang. Biol. 22, 151–163 (2016).
Sousa, L. P., Sousa, A. I., Alves, F. L. & Lillebø, A. I. Ecosystem services provided by a complex coastal region: Challenges of classification and mapping. Sci. Rep. 6, 1–14 (2016).
FAO Fisheries & Aquaculture - Species Fact Sheets - Ruditapes decussatus (Linnaeus, 1758). Available at: http://www.fao.org/fishery/species/3542/en. (Accessed: 28th May 2018).
FAO Fisheries & Aquaculture - Cultured Aquatic Species Information Programme - Ruditapes philippinarum (Adams & Reeve, 1850). Available at: http://www.fao.org/fishery/culturedspecies/Ruditapes_philippinarum/en. (Accessed: 28th May 2018).
Vilela, H. Vida bentonica de Tapes decussatus (L.). Arq. Museu Bocage 21, 120 (1950).
Lee, S. Y. Distribution pattern and interaction of two infaunal bivalves, Tapes philippinarum (Adams and Reeve) and Anomalocardia squamosa (Linnaeus) (Bivalvia: Veneridae). J. Exp. Mar. Bio. Ecol. 201, 253–273 (1996).
Mistri, M. Effect of Musculista senhousia mats on clam mortality and growth: Much ado about nothing? Aquaculture 241, 207–218 (2004).
Poppe, G. T. & Goto, Y. European Seashells. (Verlag Christa Hemmen, 1993).
Komorita, T. et al. Food Sources for Ruditapes philippinarum in a Coastal Lagoon Determined by Mass Balance and Stable Isotope Approaches. https://doi.org/10.1371/journal.pone.0086732 (2014).
Food and Agriculture Organization of the United Nations (FAO). FishStat Plus -Universal software for fishery statistical time series. Accessed 10 Aug. 2018. http://www.fao.org/fi/statist/FISOFT/FISHPLUS.asp.
DGRM. Plano Estratégico para a Aquicultura Portuguesa 2014-2020. 10.15713/ins.mmj.3 (2016).
Flassch, J. P. & Leborgne, Y. Introduction inEurope, from 1972 to 1980, of the Japanese Manila clam (Tapes philippinarum) and the effects on aquaculture production and natural settlement. ICES mar. Sci. Symp 194, 92–96 (1992).
Matias, D. et al. The reproductive cycle of the European clam Ruditapes decussatus (L., 1758) in two Portuguese populations: Implications for management and aquaculture programs. Aquaculture 406–407, 52–61 (2013).
Humphreys, J. et al. Introduction, dispersal and naturalization of the Manila clam Ruditapes philippinarum in British estuaries, 1980–2010. J. Mar. Biol. Assoc. United Kingdom 95, 1163–1172 (2015).
de Montaudouin, X. et al. Asari clam (Ruditapes philippinarum) in France: history of an exotic species 1972 – 2015. Bull. Japan Fish. Res. Educ. Agency. 35–42 (2016).
Usero, J., González-Regalado, E. & Gracia, I. Trace metals in the bivalve molluscs Ruditapes decussatus and Ruditapes philippinarum from the Atlantic Coast of Southern Spain. Environ. Int. 23, 291–298 (1997).
Dang, C. et al. The Manila clam population in Arcachon Bay (SW France): Can it be kept sustainable? J. Sea Res. 63, 108–118 (2010).
Pranovi, F. et al. An ecological imbalance induced by a non-native species: The Manila clam in the Venice Lagoon. Biol. Invasions 8, 595–609 (2006).
Basuyaux, O. et al. Evaluation of Ruditapes spp. Clam Stock on the Western Coast of Cotentin (English Channel). J. Mar. Biol. Oceanogr. 07 (2018).
Maia, F. & Gaspar, M. Caso de Estudo: Apanha de bivalves na Ria de Aveiro - Distribuição e abundância dos moluscos bivalves com maior interesse comercial na Ria de Aveiro Francisco (2014).
Donaghy, L., Lambert, C., Choi, K. S. & Soudant, P. Hemocytes of the carpet shell clam (Ruditapes decussatus) and the Manila clam (Ruditapes philippinarum): Current knowledge and future prospects. Aquaculture 297, 10–24 (2009).
Norkko, J. & Shumway, S. E. In Shellfish Aquaculture and the Environment (ed. Shumway, S. E.) 297–317 (John Wiley & Sons, Inc., 2011).
Beauchard, O., Veríssimo, H., Queirós, A. M. & Herman, P. M. J. The use of multiple biological traits in marine community ecology and its potential in ecological indicator development. Ecol. Indic. 76, 81–96 (2017).
Kristensen, E. et al. What is bioturbation? the need for a precise definition for fauna in aquatic sciences. Mar. Ecol. Prog. Ser. 446, 285–302 (2012).
Montserrat, F. et al. Sediment segregation by biodiffusing bivalves. Estuar. Coast. Shelf Sci. 83, 379–391 (2009).
Volkenborn, N., Hedtkamp, S. I. C., van Beusekom, J. E. E. & Reise, K. Effects of bioturbation and bioirrigation by lugworms (Arenicola marina) on physical and chemical sediment properties and implications for intertidal habitat succession. Estuar. Coast. Shelf Sci. 74, 331–343 (2007).
Mermillod-Blondin, F. & Rosenberg, R. Ecosystem engineering: The impact of bioturbation on biogeochemical processes in marine and freshwater benthic habitats. Aquat. Sci. 68, 434–442 (2006).
Gilbertson, W. W., Solan, M. & Prosser, J. I. Differential effects of microorganism-invertebrate interactions on benthic nitrogen cycling. FEMS Microbiol. Ecol. 82, 11–22 (2012).
Lillebø, A. I., Flindt, M. R., Pardal, M. Â. & Marques, J. C. The effect of macrofauna, meiofauna and microfauna on the degradation of Spartina maritima detritus from a salt marsh area. Acta Oecologica 20, 249–258 (1999).
Bertics, V. J. et al. Burrowing deeper into benthic nitrogen cycling: The impact of Bioturbation on nitrogen fixation coupled to sulfate reduction. Mar. Ecol. Prog. Ser. 409, 1–15 (2010).
Doering, P. H., Kelly, R. & Oviatt, C. A. S. T. Effect of the hard clam Mercenaria mercenaria on benthic fluxes of inorganic nutrients and gases. Mar. Biol. 308, 299–308 (1987).
Lavoie, M., McKindsey, C., Pearce, C. & Archambault, P. Influence of intertidal Manila clam Venerupis philippinarum aquaculture on biogeochemical fluxes. Aquac. Environ. Interact. 8, 117–130 (2016).
ICES. Report of the Benthos Ecology Working Group (BEWG) (2017).
Majdi, N., Bardon, L. & Gilbert, F. Quantification of sediment reworking by the Asiatic clam Corbicula fluminea Müller, 1774. Hydrobiologia 732, 85–92 (2014).
Vaugn, C. & Hakenkamp, C. The functional role of burrowing bivalves in freshwater ecosystems - Vaughn − 2008 -Freshwater Biology - Wiley Online Library. Freshw. Biol. 46, 1431–1446 (2001).
Sobral, P. Ecophysiology of Ruditapes decussatus. Fac. Ciências e Tecnol. Univ. Nov. Lisboa (1995).
Sgro, L., Mistri, M. & Widdows, J. Impact of the infaunal Manila clam, Ruditapes philippinarum, on sediment stability. Hydrobiologia 550, 175–182 (2005).
Spencer, B. E., Kaiser, M. J. & Edwards, D. B. Intertidal clam harvesting: Benthic community change and recovery. Aquac. Res. 29, 429–437 (1998).
Rhoads, D. C. & Young, D. K. The influence of deposit-feeding organisms on sediment stability and community trophic structure. J. Mar. Res. 28, 150–178 (1970).
Aller, R. C. In Nitrogen cycling in coastal marine environments. (eds Blackburn, T. H. & Sorensen, J.) 301–341 (John Wiley & SonsLtd, 1988).
Dame, R. Ecology of Marine Bivalves. (CRC Press, https://doi.org/10.1201/b11220, 2011).
Zhu, S., Saucier, B., Durfey, J., Chen, S. & Dewey, B. Waste excretion characteristics of Manila clams (Tapes philippinarum) under different temperature conditions. Aquac. Eng. 20, 231–244 (1999).
Lopes, M. L., Marques, B., Dias, J. M., Soares, A. M. V. M. & Lillebø, A. I. Challenges for the WFD second management cycle after the implementation of a regional multi-municipality sanitation system in a coastal lagoon (Ria de Aveiro, Portugal). Sci. Total Environ. 586, 215–225 (2017).
Araújo, I. B., Dias, J. M. & Pugh, D. T. Model simulations of tidal changes in a coastal lagoon, the Ria de Aveiro (Portugal). Cont. Shelf Res. 28, 1010–1025 (2008).
Ferreira, A. M. et al. Mortalidade da ameijoa Ruditapes decussatus na Ria Formosa Algarve. (Relat. Tec. Cient., INIP, 1989).
Sobral, P. & Fernandes, S. Physiological responses and scope for growth of Ruditapes decussatus from Ria Formosa, southern Portugal, exposed to increased ambient ammonia. Sci. Mar. 68, 219–225 (2003).
Falcao, M. & Vale, C. Study of the Ria Formosa ecosystem: benthic nutrient remineralization and tidal variability of nutrients in the water. Hydrobiologia 207, 137–146 (1990).
Bartoli, M. et al. Impact of Tapes philippinarum farming on nutrient dynamics and benthic respiration in the Sacca di Goro. Hydrobiologia 455, 203–212 (2001).
Coelho, J. P., Flindt, M. R., Jensen, H. S., Lillebø, A. I. & Pardal, M. A. Phosphorus speciation and availability in intertidal sediments of a temperate estuary: Relation to eutrophication and annual P-fluxes. Estuar. Coast. Shelf Sci. 61, 583–590 (2004).
Otero, M. et al. Kinetics of the PO4-P adsorption onto soils and sediments from the Mondego estuary (Portugal). Mar. Pollut. Bull. 77, 361–366 (2013).
Mortimer, R. J. G. et al. The effect of macrofauna on porewater profiles and nutrient fluxes in the intertidal zone of the humber estuary. Estuar. Coast. Shelf Sci. 48, 683–699 (1999).
Lillebø, A. I., Neto, J. M., Flindt, M. R., Marques, J. C. & Pardal, M. A. Phosphorous dynamics in a temperate intertidal estuary. Estuar. Coast. Shelf Sci. 61, 101–109 (2004).
Rodrigues, A. M., Quintino, V., Sampaio, L., Freitas, R. & Neves, R. Benthic biodiversity patterns in Ria de Aveiro, Western Portugal: Environmental-biological relationships. Estuar. Coast. Shelf Sci. 95, 338–348 (2011).
Venice System. Final resolution of the symposium on the classification of brackish waters. Arch. Oceanogr. Limnol. 11, 243–248 (1959).
Solan, M. et al. In situ quantification of bioturbation using time-lapse fluorescent sediment profile imaging (f-SPI), luminophore tracers and model simulation. Mar. Ecol. Prog. Ser. 271, 1–12 (2004).
Hale, R., Mavrogordato, M. N., Tolhurst, T. J. & Solan, M. Characterizations of how species mediate ecosystem properties require more comprehensive functional effect descriptors. Sci. Reports 4 (2014).
Dias, J. A. A análise sedimentar e o conhecimentos dos sistemas marinhos (Uma Introdução à Oceanografia Geológica). (Universidade do Algarve, 2004).
Doeglas, D. J. Grain size indices, classification and environment. Sedimentology 10, 83–100 (1968).
Larsonneur, C. La cartographie de’s dépots meubles sur le plateau continental français: méthode mise du points et utilisée em Manche. J. Rech. Océanographique 2, 33–39 (1977).
Kristensen, E. & Andersen, F. Ø. Determination of organic carbon in marine sediments: a comparison of two CHN-analyzer methods. J. Exp. Mar. Bio. Ecol. 109, 15–23 (1987).
Metodik, L. FerskvandsbiologiskLaboratorium. (ed Københavns Universitet, 1992).
Strickland, J. D. H. & Parsons, T. R. A Practical Handbook of Seawater Analysis. 167, (Supply and Services, 1972).
Anderson, M. J. A new method for non-parametric multivariate analysis of variance. Austral Ecol. 26, 32–46 (2001).
Clarke, K. R. & Gorley, R. N. PRIMERv6: User Manual/Tutorial. 190 (2006).
Anderson, M. J., Gorley, R. N. & Clarke, K. R. PERMANOVA+for PRIMER: Guide to Software and Statistical Methods. 214 (2008).
Acknowledgements
This research was supported by the project SmartBioR - “Intelligent Enhancement of Endogenous Marine Biological Resources in a Changing Climate”, reference Center-01-0145-FEDER-000018, financed by the Regional Operational Programme of the Center (Centro2020), through the FEDER, under the new agreement partnership PT2020. Thanks are due for the financial support to CESAM (UID/AMB/50017 - POCI-01-0145-FEDER-007638), to FCT/MCTES through national funds (PIDDAC), and the co-funding by the FEDER, within the PT2020 Partnership Agreement and Compete 2020. M.D. was supported by Investigador FCT programme (IF/00919/2015).
Author information
Authors and Affiliations
Contributions
Conceived and designed the experiments: M.L.L., J.P.R., D.C., M.D., R.C. and A.I.L. Performed the experiments: M.L.L. and J.P.R. Analysed the data: M.L.L., J.P.R., D.C., M.D., R.C. and A.I.L. Contributed reagents/materials/analysis tools: D.C., M.D., R.C. and A.I.L. All authors wrote and reviewed the manuscript.
Corresponding authors
Ethics declarations
Competing Interests
The authors declare no competing interests.
Additional information
Publisher’s note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Lopes, M.L., Rodrigues, J.P., Crespo, D. et al. Functional traits of a native and an invasive clam of the genus Ruditapes occurring in sympatry in a coastal lagoon. Sci Rep 8, 16901 (2018). https://doi.org/10.1038/s41598-018-34556-8
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-018-34556-8
Keywords
This article is cited by
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
