
Abstract
Growth differentiation factors 1 (GDF1) and 3 (GDF3) are members of the transforming growth factor superfamily (TGF-β) that is involved in fundamental early-developmental processes that are conserved across vertebrates. The evolutionary history of these genes is still under debate due to ambiguous definitions of homologous relationships among vertebrates. Thus, the goal of this study was to unravel the evolution of the GDF1 and GDF3 genes of vertebrates, emphasizing the understanding of homologous relationships and their evolutionary origin. Our results revealed that the GDF1 and GDF3 genes found in anurans and mammals are the products of independent duplication events of an ancestral gene in the ancestor of each of these lineages. The main implication of this result is that the GDF1 and GDF3 genes of anurans and mammals are not 1:1 orthologs. In other words, genes that participate in fundamental processes during early development have been reinvented two independent times during the evolutionary history of tetrapods.
Similar content being viewed by others


Introduction
Growth differentiation factors 1 (GDF1) and 3 (GDF3) are members of the transforming growth factor superfamily (TGF-β) that were originally isolated from mouse embryonic libraries1,2. They perform fundamental roles during early development, GDF1 has been mainly associated with the regulation of the left-right patterning, whereas GDF3 is mainly involved in the formation of the anterior visceral endoderm, mesoderm and the establishment of anterior-posterior identity of the body3,4,5,6,7,8,9,10,11,12,13,14,15,16,17,18,19,20. Deficiencies in GDF1 and/or GDF3 give rise to a broad spectrum of defects including right pulmonary isomerism, visceral situs inversus, transposition of the great arteries, and cardiac anomalies among others9,21,22,23,24,25. In addition to their developmental roles, GDF1 has been described as a tumor suppressor gene in gastric cells, counteracting tumorogenesis by stimulating the SMAD signaling pathway26, and GDF3 has been associated with the regulation of adipose tissue homeostasis and energy balance during nutrient overload27,28,29.
The evolutionary history of the GDF1 and GDF3 genes is still a matter of debate due to the unclear definition of homologous relationships. Understanding homology is a fundamental aspect of biology as it allows us to comprehend the degree of relatedness between genes that are associated with a given phenotype in a group of organisms. This is particularly important for GDF1 and GDF3 as these genes perform biological functions during early stages of development that define key aspects of the body plan of all vertebrates3,4,5,6,7,8,9,10,11,12,13,14,15,16,17,18,19,20. Until now, most of the inferences of the homology relationships of these genes have been based on functional information. For example, based on the developmental processes that these genes regulate, it has been suggested that the mammalian GDF1 gene is the true ortholog of the Vg1 (GDF1) gene found in amphibians7,9,21. Furthermore, phylogenetic analyses performed by Andersson et al.30 were not able to define orthologous relationships between the GDF1 and GDF3 genes among vertebrates. However after performing genomic comparisons, these authors did propose that the GDF1 gene present in mammals is the true ortholog of the Vg1 (GDF1) gene present in amphibians30. It was also suggested that the GDF3 gene could be an evolutionary innovation of mammals, as Andersson et al.30 did not find GDF3 sequences in the amphibian and bird genomes30,31. Given this scenario, the single copy gene found in amphibians and birds would be a co-ortholog of the mammalian genes30. More recently, a second copy of a GDF gene (derrière) has been annotated at the 3′ side of Vg1 (GDF1) in the genome of the western clawed frog (Xenopus tropicalis), opening new avenues to define homologous relationships and the origin of genes that accomplish fundamental roles during early development in vertebrates.
With emphasis on understanding homologous relationships and their evolutionary origin, the goal of this study was to unravel the evolution of the GDF1 and GDF3 genes of vertebrates. Our phylogenetic analyses revealed that the GDF1 and GDF3 genes of anurans and mammals are the products of independent duplication events of an ancestral gene in the ancestor of each of these groups. We also found the signature of two chromosomal translocations, the first occurred in the ancestor of tetrapods whereas the second was found in the ancestor of mammals. Thus, our results support the hypothesis that in anurans and mammals, descendent copies of the same ancestral gene (GDF1/3) have independently subfunctionalized to perform key developmental functions in vertebrates.
Results and Discussion
Phylogenetic analyses suggest an independent origin of the GDF1 and GDF3 genes in mammals and anurans
From an evolutionary perspective, the definition of homologous relationships among GDF1 and GDF3 genes is still a matter of debate30,31. The resolution of this homology is important as these genes are involved in fundamental developmental processes which are conserved all across vertebrates16,32,33,34. Thus, if extant species inherited these genes from the vertebrate ancestor, the developmental processes in which GDF1 and GDF3 are involved have a single evolutionary origin and are comparable among species.
Based on evolutionary analyses and the developmental processes in which GDF1 has been shown to participate, it has been suggested that GDF1 in mammals, birds, and amphibians are 1:1 orthologs7,9,21,30,35. Given that it has not been possible to identify copies of the GDF3 gene in amphibians and birds, it has been proposed that this gene is an evolutionary innovation of mammals, and the single gene copy found in amphibians and birds is co-ortholog to the mammalian duplicates30. However, a second copy of a GDF gene has been annotated in the genome of the western clawed frog (Xenopus tropicalis)36. This newly described gene is located at the 3´ side of the GDF1 gene and indicates that amphibians, like mammals, also have a repertoire of two GDF genes (GDF1 (Vg1) and GDF3 (derrière)). This finding opens new ways to define homologous relationships and the origin of genes involved in the early development of all vertebrates16,32,33,34.
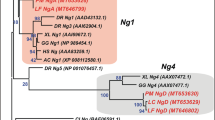
Our maximum likelihood and Bayesian reconstructions strongly suggest that the GDF1 and GDF3 genes of anurans and mammals originated independently in each of these lineages (Fig. 1). In the case of anurans, our phylogenetic tree suggests that the duplication event occurred after the divergence between caudata and anura but before the radiation of anurans. In mammals, the origin of the duplicated genes would have occurred after the divergence between mammals and sauropsids but before mammalian radiation. In agreement with the literature, we found only one gene in sauropsids (e.g. birds, turtles, lizards and allies), suggesting that they retained the ancestral condition of a single gene copy as found caudata, coelacanths, bony fish, chondrichthyes and cyclostomes (Fig. 1). Dot-plot comparisons in representative species of all main groups of sauropsids provided further support for the presence of a single gene copy in sauropsids as no traces of an extra GDF gene were present in the syntenic region of representative species of the group (Fig. 2). However, we cannot rule out an alternative scenario of duplication and subsequent gene loss in the ancestor of sauropsids. Thus, our evolutionary analyses suggest that the GDF1 and GDF3 genes present in anurans and mammals diversified independently in the ancestor of each of these lineages. In other words, genes that participate in fundamental processes during early development have been reinvented two independent times during the evolutionary history of tetrapods. As a consequence of the independent origin of these genes in anurans and mammals, they are not 1:1 orthologs.

Maximum likelihood tree depicting evolutionary relationships among the GDF1 and GDF3 genes of chordates. Numbers on the nodes represent maximum likelihood ultrafast bootstrap and Bayesian posterior probability support values. The arrows indicate the support value in nodes in which space is limited to write it. BMP2 sequences from urochordates (Ciona intestinalis) and cephalochordates (Branchiostoma floridae) were used as outgroups.

Dot-plots of pairwise sequence similarity between the GDF1 and GDF3 genes of the western clawed frog (Xenopus tropicalis) and the corresponding syntenic region in representative species of all main groups of sauropsids, the American alligator (Alligator mississippiensis), Burmese python (Python bivittatus), chicken (Gallus gallus) and green turtle (Chelonia mydas).
In the literature there are other cases of groups of genes that perform similar biological functions in a diversity of species but that have originated via lineage independent duplication events37,38,39,40,41,42,43. Among these, the independent origin of the β-globin gene cluster in all main groups of tetrapods (e.g. therian mammals, monotremes, birds, crocodiles, turtles, squamates, amphibians) represents a well-documented phenomenon41,42,43. This case is of particular interest as the two β-globin subunits that come from a gene family that has been reinvented several times during the evolutionary history of tetrapods are assembled in a tetramer with two α-globin subunits that belong to a group of genes that possess a single origin37,41,42,43. Besides this example, the independent origin of gene families makes the task of comparison difficult, as the repertoire of genes linked to physiological processes in different lineages do not have the same evolutionary origin. Additionally, the fact that the evolutionary process can give rise to similar phenotypes following different mutational pathways makes the problem of comparing even more difficult44. This is particularly important when extrapolating the results of physiological studies performed in model species to other organisms.
Two translocation events during the evolutionary history of the GDF1 and GDF3 genes of tetrapods
Based on the location of the GDF1 and GDF3 genes in the different chromosomes and the conservation pattern of flanking genes, we propose that during the evolutionary history of tetrapods, these genes underwent two chromosomal translocation events (Fig. 3). According to our analyses the genomic region that harbors the single gene copy of chondrichthyes, bony fish, and coelacanths is conserved, yet this region in these groups differs from the regions in which GDF1 and GDF3 are located in tetrapods. Overall this suggests that the first translocation event occurred in the ancestor of tetrapods (Fig. 3). Interestingly, the genomic region where the single gene copy is located in non-tetrapod vertebrates, which is defined by the presence of upstream (BMP2, HAO1, TMX4 and PLCB1) and downstream genes (FERMT1, LRRN4, CRLS1), is conserved in tetrapods. Given this, it is possible to identify the chromosomal location where the gene was located in the tetrapod ancestor before the first translocation event (Fig. 3). On the other hand, the fact that the mammalian GDF1 and GDF3 genes are located in different chromosomes suggests that the second translocation event occurred in the ancestor of the group (Fig. 3). In humans, the GDF1 gene is located on chromosome 19 while GDF3 is located on chromosome 12. In opossum (Didelphis virginiana) the GDF1 and GDF3 genes are located on chromosomes 3 and 8, respectively.

Structure of the chromosomal region containing the GDF1 and GDF3 genes of vertebrates. Asterisks denote that the orientation of the genomic piece is from 3′ to 5′, gray lines represent intervening genes that do not contribute to conserved synteny.
Evolution of GDF1 and GDF3 genes in vertebrates
In this study we present compelling evidence suggesting that genes involved in fundamental roles during early development3,4,5,6,7,8,9,12,13,14,15,17,18,19,20,30 in anurans and mammals are the product of independent duplications events. As such, these results indicate that the GDF1 and GDF3 genes of anurans and mammals are not 1:1 orthologs.
Thus, according to our results the last common ancestor of vertebrates had a repertoire of one gene (GDF1/3), and this condition is maintained in actual species of cylostomes, chondrichthyes, bony fish, coelacanths and caudata (Fig. 4). In the ancestor of tetrapods, the single gene copy was translocated from a chromosomal region defined by the presence of the BMP2, HAO1, TMX4, PLCB1, FERMT1, LRRN4, CRLS1 genes to a chromosomal region defined by the presence of the CERS1, COPE, DDX49 and HOMER3 genes (Fig. 3). After this translocation, the ancestral gene underwent a duplication event in the anuran ancestor, giving rise to the GDF1 (Vg1) and GDF3 (derrière) genes as they are found in actual species. In the case of the western clawed frog (Xenopus tropicalis) these genes are located in tandem on chromosome 28 (Fig. 3). Sauropsids, the group that includes birds, crocodiles, turtles, lizards and snakes, inherited the ancestral condition of a single gene copy as is seen in non-tetrapod vertebrates (Fig. 4). Finally, in the ancestor of mammals, the ancestral gene also underwent a duplication event, giving rise to the GDF1 and GDF3 genes found in extant species of mammals. After duplication but before the radiation of the group, a second translocation event occurred in the ancestor of the group (Fig. 4). Thus, all mammals inherited a repertoire of two genes (GDF1 and GDF3) that are located on two chromosomes.

An evolutionary hypothesis of the evolution of the GDF1 and GDF3 genes in vertebrates. According to this model the last common ancestor of vertebrates had a repertoire of one gene (GDF1/3), a condition that has been maintained in actual species of cylostomes, chondrichthyes, bony fish, coelacanths, caudata and sauropsids. In the ancestor of tetrapods, the single gene copy was translocated to a different chromosomal location. After that, in the anuran ancestor, the single gene copy underwent a duplication event giving rise to the anuran GDF1 (Vg1) and GDF3 (derrière) genes. In the ancestor of mammals, the single gene copy also underwent a duplication event, giving rise to the mammalian GDF1 and GDF3 genes. After the duplication, but before the radiation of the group, a second translocation event occurred in the ancestor of the group. Thus, all mammals inherited a repertoire of two genes (GDF1 and GDF3) that were located on two chromosomes.
Concluding remarks
This study provides a comprehensive evolutionary analysis of the GDF1 and GDF3 genes in representative species of all main groups of vertebrates. The main focus of this study was to unravel the duplicative history of the GDF1 and GDF3 genes and to understand homologous relationships among vertebrates. Understanding homology in this case is particularly important as these genes perform fundamental roles during early development that are conserved across vertebrates16,32,33,34. Our results revealed that the GDF1 and GDF3 genes present in anurans and mammals are the product of independent duplication events in the ancestor of each of these groups. Subsequently, the GDF1 and GDF3 genes of anurans and mammals are not 1:1 orthologs. Our results also show that all other vertebrate groups - i.e non-tetrapods, caudata and sauropsids – maintained the ancestral condition of a single gene copy (GDF1/3).
From an evolutionary perspective the independent duplication events that occurred in the ancestors of mammals and anurans could have resulted in the division of labor, with some degree of redundancy of the function performed by the ancestral gene. In support of this idea, it has been shown that the GDF1 and GDF3 genes of mammals have partially redundant functions during development where GDF1 can to some degree compensate for the lack of GDF330. Additionally, it has also been shown that double knockout animals (GDF1−/− and GDF3−/−) present more severe phenotypes than those of either single knockout (GDF1−/− or GDF3−/−)30. Detailed comparisons of the developmental roles of the GDF1 and GDF3 genes between mammals and anurans will shed light regarding the reinvention of genes that possess fundamental roles during early development. On the other hand, comparing mammals and anurans with sauropsids will provide useful information regarding the evolutionary fate of duplicated genes. Finally, it would be interesting to study the evolutionary history of genes that cooperate with GDF1 and GDF3 during early development (e.g. nodal)16,33,45 in order to understand the evolutionary nature of the entire developmental network.
Material and Methods
DNA sequences and phylogenetic analyses
We annotated GDF1 and GDF3 genes in representative species of chordates. Our study included representative species from mammals, birds, reptiles, amphibians, coelacanths, holostean fish, teleost fish, cartilaginous fish, cyclostomes, urochordates and cephalochordates (Supplementary dataset 1). We identified genomic pieces containing GDF 1 and GDF3 genes in the Ensembl database using BLASTN with default settings or NCBI database (refseq_genomes, htgs, and wgs) using tbalstn (Altschul et al., 1990) with default settings. Conserved synteny was also used as a criterion to define the genomic region containing GDF1 and GDF3 genes. Once identified, genomic pieces were extracted including the 5′and 3′ flanking genes. After extraction, we curated the existing annotation by comparing known exon sequences to genomic pieces using the program Blast2seq with default parameters (Tatusova and Madden 1999). Putatively functional genes were characterized by an open intact reading frame with the canonical exon/intron structure typical of vertebrate GDF1 and GDF3 genes. Sequences derived from shorter records based on genomic DNA or cDNA were also included in order to attain a broad and balanced taxonomic coverage. Amino acid sequences were aligned using the L-INS-i strategy from MAFFT v.746 (Supplementary Dataset 2). Phylogenetic relationships were estimated using maximum likelihood and Bayesian approaches. We used the proposed model tool from IQ-Tree47 to select the best-fitting model (JTT + F + R5). Maximum likelihood analysis was also performed in IQ-Tree47 to obtain the best tree. Node support was assessed with 1000 bootstrap pseudoreplicates using the ultrafast routine (Supplementary dataset 3). Bayesian searches were conducted in MrBayes v.3.1.248. Two independent runs of six simultaneous chains for 10 × 106 generations were set, and every 2,500 generations were sampled using default priors. The run was considered to have reached convergence once the likelihood scores formed an asymptote and the average standard deviation of the split frequencies remained <0.01. We discarded all trees that were sampled before convergence, and we evaluated support for the nodes and parameter estimates from a majority rule consensus of the last 2,000 trees (Supplementary dataset 4). Sea squirt (Ciona intestinalis) and Florida lancelet (Branchiostoma floridae) BMP2 sequences were used as outgroups.
Assessment of conserved synteny
We examined genes found up- and downstream of GDF1 and GDF3 in species representative of vertebrates. Synteny assessment were conducted for human (Homo sapiens), chicken (Gallus gallus), western-clawed frog (Xenopus tropicalis), axolotl (Ambystoma mexicanum), coelacanth (Latimeria chalumnae), spotted gar (Lepisosteus oculatus), elephant shark (Callorhinchus milii) and sea lamprey (Petromyzon marinus). Initial ortholog predictions were derived from the EnsemblCompara database49 and were visualized using the program Genomicus v91.0150. In other cases, the genome data viewer platform from the National Center for Biotechnology information was used.
References
Lee, S.-J. Identification of a Novel Member (GDF-1) of the Transforming Growth Factor-beta Superfamily. Mol. Endocrinol. 4, 1034–1040 (1990).
Jones, C. M., Simon-Chazottes, D., Guenet, J.-L., Hogan, B. L. M. & Pasteur, L. Isolation of Vgr-2, a Novel Member of the Transforming Growth Factor-P- Related Gene Family. Mol. Endocrinol. 6, 1961–1968 (1992).
Lee, S.-J. Expression of growth/differentiation factor 1 in the nervous system: Conservation of a bicistronic structure. Biochemistry 88, 4250–4254 (1991).
Dale’, L., Matthews, G. & Colman, A. Secretion and mesoderm-inducing activity of the TGF-j-related domain of Xenopus Vgl. EMBO J. 12, 4471–4480 (1993).
Thomsen, G. H. & Melton, D. A. Processed Vg1 protein is an axial mesoderm inducer in xenopus. Cell 74, 433–441 (1993).
Hyatt, B. A., Lohr, J. L. & Yost, H. J. Initiation of vertebrate left–right axis formation by maternal Vg1. Nature 384, 62–65 (1996).
Hyatt, B. A. & Yost, H. J. The left-right coordinator: The role of Vg1 in organizing left-right axis formation. Cell 93, 37–46 (1998).
Söderström, S. & Ebendal, T. Localized expression of BMP and GDF mRNA in the rodent brain. J. Neurosci. Res. 56, 482–492 (1999).
Rankin, C. T., Bunton, T., Lawler, A. M. & Lee, S. J. Regulation of left-right patterning in mice by growth/differentiation factor-1. Nat. Genet. 24, 262–265 (2000).
Wright, C. V. E. Mechanisms of Left-Right Asymmetry: What’ s Right and What’ s Left? A recent meeting at the Juan March Foundation in. Cell 1, 179–186 (2001).
Hamada, H., Meno, C., Watanabe, D. & Saijoh, Y. Establishment of vertebrate left-right asymmetry. Nat. Rev. Genet. 3, 103–113 (2002).
Somi, S., Houweling, A. C., Buffing, A. A. M., Moorman, A. F. M. & Van Den Hoff, M. J. B. Expression of cVg1 mRNA during chicken embryonic development. Anat. Rec. 273A, 603–608 (2003).
Chen, C. The Vg1-related protein Gdf3 acts in a Nodal signaling pathway in the pre-gastrulation mouse embryo. Development 133, 319–329 (2006).
Andersson, O., Reissmann, E., Jörnvall, H. & Ibáñez, C. F. Synergistic interaction between Gdf1 and Nodal during anterior axis development. Dev. Biol. 293, 370–381 (2006).
Bengtsson, H. et al. Generation and characterization of a Gdf1 conditional null allele. Genesis 46, 368–372 (2008).
Komatsu, Y. & Mishina, Y. Establishment of left – right asymmetry in vertebrate development: the node in mouse embryos. Cell. Mol. Life Sci. 70, 4659–4666 (2013).
Arias, C. F., Herrero, M. A., Stern, C. D. & Bertocchini, F. A molecular mechanism of symmetry breaking in the early chick embryo. Scietific Reports 7, 1–6 (2017).
Bisgrove, B. W., Su, Y.-C. & Yost, H. J. Maternal Gdf3 is an obligatory cofactor in Nodal signaling for embryonic axis formation in zebrafish. Elife 6, e28534 (2017).
Montague, T. G. & Schier, A. F. Vg1-Nodal heterodimers are the endogenous inducers of mesendoderm. Elife 6, e28183 (2017).
PellicciaJ., L., Jindal, G. A. & Burdine, R. D. Gdf3 is required for robust Nodal signaling during germ layer formation and left-right patterning. Elife 6, e28635 (2017).
Wall, N. A., Craig, E. J., Labosky, P. A. & Kessler, D. S. Mesendoderm induction and reversal of left-right pattern by mouse Gdf1, a Vg1-related gene. Dev. Biol. 227, 495–509 (2000).
Karkera, J. D. et al. Loss-of-Function Mutations in Growth Differentiation Factor-1 (GDF1) Are Associated with Congenital Heart Defects in Humans. Am. J. Hum. Genet. 81, 987–994 (2007).
Kaasinen, E. et al. Recessively inherited right atrial isomerism caused by mutations in growth/differentiation factor 1 (GDF1). Hum. Mol. Genet. 19, 2747–2753 (2010).
Bao, M. W. et al. Cardioprotective role of growth/differentiation factor 1 in post-infarction left ventricular remodelling and dysfunction. J. Pathol. 236, 360–372 (2015).
Zhang, J. et al. Association of GDF1 rs4808863 with fetal congenital heart defects: A case-control study. BMJ Open 5, e009352 (2015).
Yang, W. et al. Epigenetic silencing of GDF1 disrupts SMAD signaling to reinforce gastric cancer development. Oncogene 35, 2133–2144 (2016).
Witthuhn, B. A. & Bernlohr, D. A. Upregulation of bone morphogenetic protein GDF-3/Vgr-2 expression in adipose tissue of FABP4/aP2 null mice. Cytokine 14, 129–135 (2001).
Wang, W., Yang, Y., Meng, Y. & Shi, Y. GDF-3 is an adipogenic cytokine under high fat dietary condition. Biochem. Biophys. Res. Commun. 321, 1024–1031 (2004).
Andersson, O. et al. Growth/differentiation factor 3 signals through ALK7 and regulates accumulation of adipose tissue and diet-induced obesity. Proc. Natl. Acad. Sci. USA 105, 7252–7256 (2008).
Andersson, O., Bertolino, P. & Ibáñez, C. F. Distinct and cooperative roles of mammalian Vg1 homologs GDF1 and GDF3 during early embryonic development. Dev. Biol. 311, 500–511 (2007).
Levine, A. J., Brivanlou, A. H., Suzuki, A. & Hemmati-Brivanlou, A. GDF3, a BMP inhibitor, regulates cell fate in stem cells and early embryos. Development 133, 209–16 (2006).
Mercola, M. & Levin, M. Left-right assymetry determination in vertebrates. Annu. Rev. Cell Dev. Biol. 17, 779–805 (2001).
Levine, A. J. GDF3, a BMP inhibitor, regulates cell fate in stem cells and early embryos. Development 133, 209–216 (2005).
Nakamura, T. & Hamada, H. Left-right patterning: conserved and divergent mechanisms. Development 139, 3262 (2012).
Seleiro, E. A. P., Connolly, D. J. & Cooke, J. Early developmental expression and experimental axis determination by the chicken Vg1 gene. Curr. Biol. 6, 1476–1486 (1996).
Suzuki, A. et al. Genome organization of the vg1 and nodal3 gene clusters in the allotetraploid frog Xenopus laevis. Dev. Biol., https://doi.org/10.1016/j.ydbio.2016.04.014 (2017).
Goodman, M., Czelusniak, J., Koop, B. F., Tagle, D. A. & Slightom, J. L. Globins: a case study in molecular phylogeny. Cold Spring Harb. Symp. Quant. Biol. 52, 875–90 (1987).
Kriener, K., O’huigin, C. & Klein, J. Independent Origin of Functional MHC Class II Genes in Humans and New World Monkeys. Hum. Immunol. 62, 1–14 (2001).
Wallis, O. C. & Wallis, M. Characterisation of the GH gene cluster in a new-world monkey, the marmoset (Callithrix jacchus). J. Mol. Endocrinol. 29, 89–97 (2002).
Li, Y. et al. Independent origin of the growth hormone gene family in New World monkeys and Old World monkeys/hominoids. J. Mol. Endocrinol. 35, 399–409 (2005).
Opazo, J. C., Hoffmann, F. G. & Storz, J. F. Genomic evidence for independent origins of -like globin genes in monotremes and therian mammals. Proc. Natl. Acad. Sci. USA 105, 1590–1595 (2008).
Hoffmann, F. G., Storz, J. F., Gorr, T. A. & Opazo, J. C. Lineage-specific patterns of functional diversification in the α-and β-globin gene families of tetrapod vertebrates. Mol. Biol. Evol. 27, 1126–1138 (2010).
Hoffmann, F. G., Vandewege, M. W., Storz, J. F. & Opazo, J. C. Gene Turnover and Diversification of the α- and β-Globin Gene Families in Sauropsid Vertebrates. Genome Biol. Evol. 10, 344–358 (2018).
Natarajan, C. et al. Predictable convergence in hemoglobin function has unpredictable molecular underpinnings. Science 354, 336–339 (2016).
Ramsdell, A. F. & Yost, H. J. Molecular mechanisms of vertebrate left–right development. Trends Genet. 14, 459–465 (1998).
Katoh, K. & Standley, D. M. MAFFT Multiple Sequence Alignment Software Version 7: Improvements in Performance and Usability. Mol. Biol. Evol. 30, 772–780 (2013).
Trifinopoulos, J., Nguyen, L.-T., von Haeseler, A. & Minh, B. Q. W-IQ-TREE: a fast online phylogenetic tool for maximum likelihood analysis. Nucleic Acids Res. 44, W232–W235 (2016).
Ronquist, F. & Huelsenbeck, J. P. MrBayes 3: Bayesian phylogenetic inference under mixed models. Bioinformatics 19, 1572–4 (2003).
Herrero, J. et al. Ensembl comparative genomics resources. Database 2016, baw053 (2016).
Louis, A., Nguyen, N. T. T., Muffato, M. & Roest Crollius, H. Genomicus update 2015: KaryoView and MatrixView provide a genome-wide perspective to multispecies comparative genomics. Nucleic Acids Res. 43, D682–D689 (2015).
Acknowledgements
This work was supported by the Fondo Nacional de Desarrollo Científico y Tecnológico (FONDECYT 1160627) grant to JCO.
Author information
Authors and Affiliations
Contributions
Designed the research: J.C.O.; carried out the research: J.C.O. and K.Z.; contributed materials/reagents/analysis tools: J.C.O.; wrote the paper: J.C.O.
Corresponding author
Ethics declarations
Competing Interests
The authors declare no competing interests.
Additional information
Publisher's note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Opazo, J.C., Zavala, K. Phylogenetic evidence for independent origins of GDF1 and GDF3 genes in anurans and mammals. Sci Rep 8, 13595 (2018). https://doi.org/10.1038/s41598-018-31954-w
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-018-31954-w
Keywords
This article is cited by
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
