
Abstract
Despite the effectiveness of mass distribution of long-lasting insecticidal nets (LLINs) in reducing malaria transmission in Africa, in hyperendemic areas such as Burkina Faso the burden of malaria remains high. We here report the results of a 4-month survey on the feeding habits and Plasmodium infection in malaria vectors from a village in Burkina Faso one year following a national LLIN distribution programme. Low values of human blood index (HBI) observed in the major malaria vectors in the area (Anopheles coluzzii: N = 263, 20.1%; An. arabiensis: 5.8%, N = 103) are consistent with the hypothesis that LLINs reduced the availability of human hosts to mosquitoes. A regression meta-analysis of data from a systematic review of published studies reporting HBI and sporozoite rates (SR) for An. gambiae complex revealed that the observed SR values (An. coluzzii: 7.6%, N = 503; An. arabiensis: 5.3%, N = 225) are out of the ranges expected based on the low HBI observed. We hypothesize that a small fraction of inhabitants unprotected by bednets acts as a “core group” repeatedly exposed to mosquito bites, representing the major Plasmodium reservoir for the vectors, able to maintain a high risk of transmission even in a village protected by LLINs.
Similar content being viewed by others

Introduction
Malaria incidence and mortality have globally decreased in the last decade thanks to implementation of integrated control measures, including effective anti-malarial treatments, better health infrastructures, the widespread application of measures reducing human-vector contact (i.e. insecticide treated nets (ITNs), long-lasting insecticide treated nets (LLINs), and indoor residual spraying, IRS)1,2,3,4. It is estimated that 68% of the 663 million malaria cases prevented in Africa since the year 2000 are due to the usage of LLINs4. Between 2008 and 2010, 254 million ITNs were supplied to countries in sub-Saharan Africa, bringing the proportion of African households in possession of a net increased from 3% in 2000 to 50% in 20105. Yet, most of sub-Saharan Africa continues to carry a disproportionately high share of the global malaria burden of malaria deaths6,7,8,9,10. This is certainly the case for Burkina Faso where raising of LLIN coverage from 20% to 70% between 2009 and 2014 led to a significant reduction of malaria prevalence, but did not significantly affect malaria annual incidence1,4,11,12,13,14,15,16,17,18,19 (Fig. 1). Low levels of bednet usage20,21 and/or reduced efficacy in preventing vector/human contact due to physiological and behavioural resistance of mosquito vectors to pyrethroids used to impregnate the nets2,22,23,24,25 may have concurred in producing this heterogeneous efficacy.

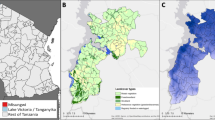
Annual countrywide data on malaria cases for Burkina Faso, estimated per 1,000 inhabitants (2–10 year olds). Left vertical axis: percentage of the population protected by Insecticide Treated Nets (ITN), in red; estimated prevalence of Plasmodium falciparum (Pf) parasite rate in 2–10 year olds, in purple. Right vertical axis: Plasmodium falciparum (Pf) estimated incidence per 1,000 inhabitants, in green (source: Malaria Atlas Project, www.map.ox.ac.uk).
Entomological studies represent an important component for the full understanding of the impact of LLINs on malaria incidence and transmission dynamics. In Burkina Faso, four major vectors are responsible of most malaria burden, i.e. Anopheles coluzzii, Anopheles gambiae sensu stricto, Anopheles arabiensis, and Anopheles funestus. Their ecological features, genetic heterogeneity and plastic behaviours, together with their complementary geographic/seasonal distribution, are major threats to an effective control of malaria in the country23,26,27,28,29,30,31,32,33,34,35,36,37,38. In fact, these anthropophilic and endophilic species have demonstrated the ability to easily switch to more zoophilic and/or exophilic behaviours as a consequence of the implementation of vector control measures39,40,41.
We here present the result of an entomological survey carried out the year after a LLIN mass-distribution campaign in a village of Ziniaré health district in Burkina Faso where high levels of malaria incidence were observed11,12,13,14,19 (Fig. 2). In order to assess whether the ratio between Plasmodium sporozoite rate (SR) and human blood index (HBI) in the study village was in line with those observed in other Afrotropical villages with high LLIN coverage, we compared it with the estimated ratios in Afrotropical villages with different LLIN coverage, as obtained by a systematic review and meta-analysis of published data indexed in PubMed database. Results suggest that, whilst LLIN use led to a strong reduction in human-vector-contact, high Plasmodium infection rates were still present in the mosquito vector population from the study village, suggesting a residual high risk of infection for unprotected inhabitants both indoors and outdoors, despite the high LLIN coverage.

Annual data on malaria cases (blue = total, red = uncomplicated, and green = severe) from 2009 to 2016, calculated per 1,000 inhabitants, in the district of Ziniaré, Burkina Faso. Source: Ministry of Health of Burkina Faso (http://www.cns.bf/spip.php?id_rubrique=17&page=publdetails).
Results and Discussion
Plasmodium infection rates
Overall, Plasmodium infection rates obtained by nested-PCR on DNA-templates extracted from the entire carcasses of 1,115 An. gambiae s.l. female mosquitoes (hereafter, whole mosquito infection, WMI) were 11.2% for An. coluzzii, 9.4% for An. arabiensis and 12.5% for An. gambiae s.s. (Table 1). PCR-bands amplified from 10 Plasmodium-positive specimens were sequenced and identified as Plasmodium falciparum-specific.
No significant differences in Plasmodium infection rates were observed among species, sampling methods, indoor vs outdoor collections, and months of sampling (GLMM-1, Table (a) in Supplementary File 1). The proportion of observed WMI was within the 95% confidence intervals computed by simulating model results (Figure (a) in Supplementary File 1).
Sporozoite Rates (SRs) and Oocyst Rates (ORs) obtained using DNA extracted either from heads + thoraxes or abdomens of 728 females collected by BP and PIT, were 7.6% and 5.2% for An. coluzzii, and 5.3% and 4.9% for An. arabiensis, respectively (Table 2; rates are not reported for An. gambiae due to the small sample size; N = 31). No statistically significant differences were observed between species, sampling methods, indoor vs outdoor collections, and months of sampling (GLMM). Plasmodium infection was detected in heads + thoraxes and abdomens of the same mosquito in 8 specimens (1.1%) only.
Blood-meal origin identification
Blood meal origin identification was successful for 63% of the 602 An. gambiae s.l. females collected with blood-traces in the abdomens (Table 3). GLM-a analysis showed differences in rates of successful identification between methods of collections (73.3% and 58.7% for the 116 and 303 females collected by SRB and BP-PIT, respectively; likelihood ratio test, df = 3, deviance = 10.43, p-value = 0.015, Supplementary File 2), confirming that that presence of glue in SRB-collected samples does not represent a constraint to molecular analyses32. GLM-a also highlighted that the sensitivity of the PCR technique was affected by the amount of blood in the abdomens and/or by its partial digestion, being 74.5% in freshly/fully-fed females (N = 102), and 62.9% and 58.5% in half gravid (N = 35) and partially-fed mosquitoes (N = 282), respectively (Table (b) and Figure (a) and (B) in Supplementary File 2). This is most likely due to a more abundant and/or less degraded DNA template in freshly/fully-fed females, as already reported by Kent & Norris42. This is also consistent with the higher blood-meal identification rate in SRB-collected specimens, which included a lower proportion of half-gravid and partially-fed females. Overall, ~25% of good-quality blood-meals (corresponding to the proportion of non-identified blood-meals in fully-fed females, in which an optimal DNA-template can be assumed to have been extracted) were not identified. This not-negligible proportion is in line with literature data on An. gambiae complex feeding habits40,43,44,45,46 and is likely due to blood-meals carried out on non-tested hosts, such as chickens (which were not tested, as An. coluzzii/An. gambiae are not known to be ornithophilic)46, donkeys (which were rare in the village), or wild birds/mammals.
Overall, the Human Blood Index (HBI) calculated only specimens with successful blood-meal identification was significantly higher for An. coluzzii (20.1%, including 3 specimens showing a mixed cow-human blood meal) than for An. arabiensis (5.8%) and An. gambiae s.s. (4.3%), (Table 3; Fig. 3; Table (c) in Supplementary File 2). The higher HBI observed for An. coluzzii compared to An. arabiensis is expected based on the known higher anthropophily of the former species38,43,46, while the low HBI observed in highly anthropophilic An. gambiae s.s. may be an artefact of the small sample size.

Results of the Binomial GLMM-2 investigating the Human Blood Index in An. coluzzii and An. arabiensis fed mosquitoes. The dots represent fitted values: vertical bands are the 95% confidence intervals. On the y-axis the probability of a human blood meal (HBI), AR = An. arabiensis, CO = An. coluzzii. IN = indoor collection, OUT = outdoor collection.
Due to the expected presence of unidentified blood-meals in mosquitoes feeding on animals different from the tested ones, the reported HBI values likely represent an overestimate of the real HBI in the village. In fact, when calculated on the overall fully-fed female sample (including specimens with non-identified blood-meals), HBIs were 12.6% and 3.8% in An. coluzzii and An. arabiensis, respectively. HBI values <15% are exceptionally low for the three species, especially for An. coluzzii and An. gambiae s.s.,the most anthropophilic species of the An. gambiae complex. Notably, a bias due to higher numbers of non-identified human blood-meals in low quality blood-meal samples can be ruled out, as the PCR-fragment amplified for human-host is shorter than those of most other hosts42, and thus easier to be amplified. However, the low HBI values are parsimonious with the supposed reduced availability of human hosts to Anopheles vectors due to implementation of LLIN with coverage rates estimated at 90% in the study site the year prior to sampling. In fact, similar studies highlighted a plastic strategy of feeding behaviour of these usually highly anthropophilic species as a consequence of the reduced accessibility of human hosts and of the greater accessibility of readily available, although less-preferred, hosts40,47,48. This is consistent with the high (>70%) reported proportion of blood-meals carried out on cattle by all the 3 taxa, which is possibly associated with the presence of a settlement of pastoral Fulani families and of their cattle herds at ~400 meters from the sampled compounds.
Sporozoite rates in Anopheles coluzzii sub-samples
Sporozoite rates of An. coluzzii (the only taxon with an adequate sample size for more detailed studies) females were higher for the human-fed sub-sample (22.9%; N = 35) than for animal-fed (8.4%; N = 143) or unfed-gravid subsamples (5.5%; N = 200), as well as for the sub-sample of fed-females with unidentified blood-meals (5.6%; N = 125) (GLMM-3; Supplementary File 3). This is in agreement with previous studies suggesting that females that have already taken a blood-meal on a (infected) human host and/or that are ready to transmit the malaria parasite, have a higher tendency to bite human again49,50,51. It is believed that Plasmodium can potentially affect the complex mosquito host-seeking behaviour (in the experimental model P. yoelii-An. stephensi) due to neurophysiological mechanism, based on changes in the responsiveness of mosquito odorant receptors52. Thus, the higher SR observed in human-fed A. coluzzii could be due to a significant increase in infected mosquito attraction to human odours triggered by the parasite itself in order to enhance its chances of transmission to the appropriate host49,50,51.
Systematic review of Human Blood Index and Sporozoite Rate
The systematic review performed on published field studies indexed in PubMed reporting both SR and HBI in An. gambiae complex species retrieved 133 articles assessed for eligibility, of which 49 were retained according to eligibility criteria (see Materials and Methods), corresponding to 88 records that were considered in the analysis (Supplementary File 4). The proportion meta-analysis of the published data show a high inconsistency (I2) among studies/records in all subsamples (Table 4). Random effects estimates of HBI and SR show that the two estimates are reduced in presence of bednets when all members of the An. gambiae complex are combined. This pattern is confirmed also for An. arabiensis, while in the case of An. coluzzii and An. gambiae s.s. data were not sorted for bednet presence because of the low numbers of records, due to their recent recognition as separate species53. HBI values obtained from meta-analysis are far higher than the HBI recorded in the present study, even if only villages with bednets are included in the meta-analysis (An. gambiae s.l. 95% c.i. = 55.5–75.3%).
A positive association was shown between SR and HBI (Fig. 4, Table (a) in Supplementary File 5) also when the analysis was restricted to the 46 records including data on mosquito net use (untreated, ITN or LLIN, Table (b) and Figure (a) in Supplementary File 5). Significantly higher SRs were reported in the 10/46 records stating lack of mosquito net usage; however, this result should be interpreted with care because of the small sample size and the presence of a single record with HBI < 80% that affects the reliability of dataset for the whole range of HBI.

Results of Beta GLM-b investigating the relationship between Human Blood Indexes (HBI) and Sporozoite Rates (SR) calculated from data retrieved by systematic review. Solid line = fitted values; Dashed lines = 95% confidence intervals.
Beta GLM-b investigating the relationship between SR and HBI showed that the observed SRs obtained by nested-PCR in the current study largely exceeded the 95% confidence intervals for both An. coluzzii and An. arabiensis (red dots, Fig. 5; Figure (b) Supplementary File 5). This is still the case if An. coluzzii SRs were reduced to 5.4%, based on results from Calzetta et al.54 reporting a 1.4-fold higher sensibility of nested-PCR compared to ELISA, i.e. the most commonly used method exploited in the quoted publications) (Fig. 5; Figure (b) Supplementary File 5). However the underrepresentation of published studies reporting very low HBI may affect the reliability of the intervals for the calculated range of the corresponding SR.

Results of the Beta GLM-b investigating the relationship between Sporozoite rates (SR) and Human-Blood Index (HBI). Upper histograms represent the distribution of SR for HBI values observed in this study; x axis represents the SR expressed in percent values; y axis represent the frequency of events occurred in the simulation (see Materials & Methods); black lines are the 95% confidence intervals (bold for meta-analysis estimate, narrow for our study). Lower plot represent the observed values of HBI and SR (black dots). Vertical bands are the 95% confidence intervals of the expected SR value from the Beta GLM. Red dots represent SR obtained by nested-PCR. Blue dots represent SR obtained by nested-PCR halved according to the higher sensitivity of nested-PCR compared to widely used ELISA method.
It should be noted that HBI values utilized in the present study were obtained from specimens with successful blood meal origin identification. This implies that these values, as discussed above, likely represent an overestimation of actual number of blood-meals taken on humans. It is therefore overall safe to conclude that, particularly in the case of An. coluzzii, the SR in the study site was significantly higher than expected based on the low HBI observed and on the consequent lower probability for the mosquito to come in contact with Plasmodium parasite. Although less striking, a similar trend is also observed for An. arabiensis.
Conclusions
LLINs provide protection from malaria to net users by both the physical barrier of the net, which reduces contact with the mosquito, and by the insecticide within the net fibres, which reduces the likelihood that a mosquito will feed through the net. In addition, they provide protection to non-users by reducing the total mosquito population and shortening their life span. This community effect is observed when net usage rates exceed 50% in the population55. In the Burkinabe village of Goden, LLINs had been distributed the year prior to the study and reported net usage was highly above this value. Thus, one would have expected human vector-contact and sporozoite rates to be reduced in people both protected and unprotected by bednets, also considering the presence of alternative hosts (cattle) in the village55. Instead, the main malaria vector species in Goden showed low HBI values and rates of Plasmodium infection in the range of those normally observed in the presence of much higher rates of feeding on humans typical of these species in absence of extensive LLIN coverage. It can be hypothesized that, even if HBI is reduced due to mosquitoes feeding on alternative hosts, as humans are largely protected by LLINs, the infectious reservoir could be being maintained by non-bednet users if the local vector population was resilient to the toxic effects of the pyrethroids on the nets. Although we did not measure insecticide resistance in Goden and no data are available in literature for this village, pyrethroid resistance is widespread in Burkina Faso25,56,57,58,59 and may have played a role by reducing daily mosquito mortality, lengthening the vector population average age and increasing the risk of malaria transmission to people when unprotected by a bednet37.
In the study site the fraction of the human population not protected by a bednet and thus more readily available to the vectors may thus constitutes a “core group” of people who are repeatedly exposed to mosquito bites. These people would represent the major reservoir of Plasmodium gametocytes capable of maintaining the high levels of infection observed in the vector population. These high levels of mosquito infection, in association with high mosquito densities32,60 may sustain the transmission even to the fraction of the inhabitants commonly protected by a bednet. Although we cannot exclude that some people from Goden have been infected by travelling in other areas, the above hypothesis is supported by modelling studies showing that the effect of uneven distribution of mosquito bites - with a “core group” of people more exposed to infections than the rest of the population - is associated with an increase in human-to-mosquito transmission efficiency as malaria is controlled61. If confirmed by additional studies, this would highlight the need to specifically target the “core group” of people sustaining the transmission in order to achieve a significant reduction in transmission.
Despite the limitations of this study (i.e. lack of SR and HBI data before large-scale LLIN distribution and lack of clinical data on malaria incidence), these results may contribute to explain the low effect of LLIN distribution on malaria incidence in some African regions, such as Burkina Faso. Further studies measuring HBI and sporozoite rates in areas of high levels of pyrethroid resistance, accompanied by information on net usage and the prevalence of malaria in bednet users versus non-bednet users, are needed to establish whether the pattern we report in this study reflects contemporary patterns of malaria transmission in areas with high mosquito densities and high (but not complete) LLIN use.
Methods
Study site
Field collections were carried out from August to November 2011 in the village of Goden (Ziniaré health district; 12°25′N–1°21′W) located in a Sudanese-savanna area 41 km East of Ouagadougou, the capital city of Burkina Faso. Goden is a rural village with approximately 800 inhabitants mainly belonging to the Mossi ethnic group, mostly devoted to agriculture and rearing a few animals (e.g. pigs, dogs, goats and chickens) in their compounds. In the year of sampling, two families belonging to Fulani nomadic pastoral ethnic group settled nearby the village with approximately 200 bovines, representing the second most abundant host for mosquitoes. LLINs were distributed in 2010 to approximately 90% of the population (N’F S and MG, unofficial data).
Mosquito sampling and species identification
Field sampling is described in Pombi et al.32. In brief, mosquitoes were collected from 5 compounds, 2 houses per compound (10 in total), randomly chosen in the village, where LLINs were available. The sampling has been done in the same houses for the whole sampling season. Indoor and outdoor resting Anopheles mosquitoes were collected by back-pack aspirations either inside houses (BP) or in pit-shelters (PIT) and by sticky resting boxes (SRB). Mosquitoes glued on sticky sheets were removed by cutting out a small sheet fragment around them and washing it with acetone for 2 minutes. All collected mosquitoes were morphologically identified under a stereomicroscope62, separated by species, gender and gonotrophic stage (i.e. unfed, freshly/fully fed, half-gravid, gravid) and stored in individual tubes containing silica-gel. DNA was extracted from entire specimens in the case of SRB collections, and from heads + thoraxes and abdomens separately in the case of BP and PIT. Anopheles gambiae species complex were identified by PCR32.
Plasmodium detection and blood meal identification
DNA extracted from either heads + thoraxes or abdomens in the case of BP- and PIT-collected specimens (in order to distinguish between infective and infected mosquitoes), and from entire specimens in the case of SRB collections (which could not be easily dissected due to the glue presence) were processed by nested-PCR protocol targeting mtDNA to detect Plasmodium sp.54. A subsample of PCR products from Plasmodium positive specimens was sequenced (BMR genomics sequencing service, Padua, Italy) for species identification by BLAST (National Centre for Biotechnology Information, Bethesda, MD, USA).
DNA from either abdomens (BP and PIT) or entire specimens (SRB) was used as template for multiplex-PCR protocol targeting mtDNA to detect blood meal origin42; primers for human, cow, pig, dog and goat were used.
Statistical analyses
Generalized Linear Mixed Models (GLMM) and Generalized Linear Model (GLM) were performed assuming a binomial distribution with logit link. To incorporate a dependency structure among observations from the same house or in the same day the variable house was considered as a random effect nested into the variable compound, while the variable collection days/weeks was considered as a crossed random effect. Assessment of model assumption was carried out by graphically inspect Pearson residuals versus fitted values and each covariate.
GLMM-1 was carried out to investigate whether the probability of a mosquito being infected by Plasmodium sp. is different in Anopheles gambiae complex species (An. arabiensis, An. coluzzii, An. gambiae s.s.) accounting for collection periods (i.e. month) and sampling positions (indoors or outdoors). As part of the model validation process, 10,000 data sets have been simulated from the model to investigate if the predicted probability of infected mosquito was in accordance with the observed value (see Supplementary File 1).
The following analyses were carried out on Anopheles coluzzii and An. arabiensis (i.e. the two most represented taxa) from BP and PIT collections. Plasmodium infection in abdomen and in head-thorax was regressed against the taxa by means of Binomial GLMM.
GLM-a was carried assuming a binomial distribution with logit link out to assess differences in blood meal detection between sticky (SRB) and non-sticky methods (BP + PIT), as well as in fed mosquitoes with different degrees of digestion of blood for An. coluzzii fed mosquitoes. Tukey post-hoc test was carried out to assess the significance of all pairwise comparison. A simulation study was carried out to understand how the different sample size in each group affected the results (see Supplementary File 2).
GLMM-2 was carried out to investigate whether the probability of mosquito having fed on human was different in Anopheles gambiae complex species (An. arabiensis, An. coluzzii) accounting for indoors or outdoors collections. The model was fitted only on the subsample of identified blood meal (see Table (b) on Supplementary File 2).
GLMM-3 was carried out to investigate whether the source of blood meal (blood from human, animal or unidentified hosts plus unfed/gravid mosquitoes) affected the probability of positive infection to sporozoite in specimens of An. coluzzii.
Systematic review and meta-analysis
A systematic search and review of field studies indexed in the publication repository PubMed (www.ncbi.nlm.nih.gov/pubmed) was carried out for studies published in English, French or Italian between 1964 and August 2016. As no review protocol are already available for this systematic review, articles were searched using the keywords “gambiae”, “coluzzii”, and “arabiensis” in association with any one of the following keywords: “human blood index” (or “HBI”), “blood meal sporozoite”, “blood index Plasmodium”, “human blood sporozoite index”. From the initial set of publications the following information were retrieved and recorded in an Excel database: country of collection, village/area of collection, sampling season (dry-rainy), year of collection, presence of bednets (untreated, ITN, LLIN), mosquito species collected, human blood index, HBI sample size, protocol for HBI detection (ELISA, PCR, other), sporozoite rate (SR), SR sample size, protocol for SR detection (ELISA, PCR, other) whenever available. Articles not reporting both HBI and SR with their relative sample size were excluded from the analysis. A meta-analysis was performed to obtain weighted estimates of mean values of SR and HBI. Homogeneity across studies was assessed by the Cochran Q and I2 statistics63,64. Whenever the P-value of Cochran Q was greater than 5% and it was associated with a value of I2 ≤ 25%, the studies were considered homogeneous and the SR and HBI were calculated considering a fixed-effects combined value. Conversely, if the studies were significantly heterogeneous, a random-effects combined value was calculated. Under the fixed-effect model it is assumed that the true effect is the same in all studies, while under the random-effects model allowance is made for the true effect to vary across studies65.
The relationship between HBI and SR values obtained from the systematic review of the literature was investigated by means of GLM-b assuming a beta distribution with logit link. Since SR values included zeroes, which are excluded by the beta distribution, the transformation (Y*(N-1) + 0.5)/N was employed66. Y is the SR values and N the sample size. Transformed SR values were regressed against HBI values accounting for sampling season, region and species studied. Information on the presence of simple Mosquito Nets and Insecticide Treated Nets (including LLINs) was not available for all the studies considered. Therefore, to investigate the effects of mosquito nets on SR, Beta GLM-c was carried out using only the subsample with available data. HBI and Presence/Absence of Mosquito Nets were considered as independent variables. Finally, the expected SR values for the data observed in this study was computed using both Beta GLMs and compared to the actual value observed.
Statistical analysis were carried out using Stats Direct statistical software67, R statistical software64 version 3.3.2 and the lme468, betareg69, plyr70 packages.
References
WHO. World Malaria Report 2015. (World Health Organization, 2015).
Trape, J.-F. et al. The rise and fall of malaria in a west African rural community, Dielmo, Senegal, from 1990 to 2012: a 22 year longitudinal study. Lancet Infect. Dis. 14, 476–488 (2014).
O’Meara, W. P. et al. The impact of primary health care on malaria morbidity–defining access by disease burden. Trop. Med. Int. Health 14, 29–35 (2009).
Bhatt, S. et al. The effect of malaria control on Plasmodium falciparum in Africa between 2000 and 2015. Nature 526, 207–211 (2015).
WHO. World Malaria Report 2011. (World Health Organization, 2011).
Jagannathan, P. et al. Increasing incidence of malaria in children despite insecticide-treated bed nets and prompt anti-malarial therapy in Tororo, Uganda. Malar. J. 11, 435 (2012).
Roca-Feltrer, A. et al. Lack of decline in childhood Malaria, Malawi, 2001–2010. Emerg. Infect. Dis. 18, 272–278 (2012).
Coulibaly, D. et al. Stable malaria incidence despite scaling up control strategies in a malaria vaccine-testing site in Mali. Malar. J. 13, 374 (2014).
Mwesigwa, J. et al. On-going malaria transmission in The Gambia despite high coverage of control interventions: a nationwide cross-sectional survey. Malar. J. 14, 314 (2015).
CPS/SSDSPF, INSTAT/MPATP & ICF. Enquête Démographique et de Santé au Mali 2012–2013. (2014).
Ministry of Health of Burkina Faso. Annuaire statistique 2011 (2012).
Ministry of Health of Burkina Faso. Annuaire statistique 2014 (2015).
Ministry of Health of Burkina Faso. Annuaire statistique 2012 (2013).
Ministry of Health of Burkina Faso. Annuaire statistique 2010 (2011).
Ministry of Health of Burkina Faso. Annuaire statistique 2013 (2014).
Ministry of Health of Burkina Faso. Annuaire statistique 2006 (2007).
Ministry of Health of Burkina Faso. Annuaire statistique 2007 (2008).
Ministry of Health of Burkina Faso. Annuaire statistique 2008 (2009).
Ministry of Health of Burkina Faso. Annuaire serie longue de la santé, 1960–2009 (2011).
Sangaré, L. R. et al. Determinants of use of insecticide treated nets for the prevention of malaria in pregnancy: Jinja, Uganda. PLoS One 7 (2012).
Teklemariam, Z., Awoke, A., Dessie, Y. & Weldegebreal, F. Ownership and utilization of insecticide-treated nets (ITNs) for malaria control in Harari National Regional State, Eastern Ethiopia. Pan Afr. Med. J. 21, 1–9 (2015).
Snow, R. W. et al. Changing Malaria Prevalence on the Kenyan Coast since 1974: Climate, Drugs and Vector Control. PLoS One 10, e0128792 (2015).
Gatton, M. L. et al. The importance of mosquito behavioural adaptations to malaria control in Africa. Evolution (N. Y). 67, 1218–1230 (2013).
Russell, T. L., Beebe, N. W., Cooper, R. D., Lobo, N. F. & Burkot, T. R. Successful malaria elimination strategies require interventions that target changing vector behaviours. Malar. J. 12, 56 (2013).
Strode, C., Donegan, S., Garner, P., Enayati, A. A. & Hemingway, J. The Impact of Pyrethroid Resistance on the Efficacy of Insecticide-Treated Bed Nets against African Anopheline Mosquitoes: Systematic Review and Meta-Analysis. PLoS Med. 11 (2014).
Guelbeogo, W. M. et al. Seasonal distribution of anopheles funestus chromosomal forms from Burkina Faso. Malar. J. 8, 1–7 (2009).
Costantini, C. et al. Living at the edge: biogeographic patterns of habitat segregation conform to speciation by niche expansion in Anopheles gambiae. BMC Ecol. 9, 16 (2009).
Pombi, M. et al. Chromosomal plasticity and evolutionary potential in the malaria vector Anopheles gambiae sensu stricto: insights from three decades of rare paracentric inversions. BMC Evol. Biol. 8, 309 (2008).
Gimonneau, G. et al. Larval habitat segregation between the molecular forms of the mosquito Anopheles gambiae in a rice field area of Burkina Faso, West Africa. Med. Vet. Entomol. 26, 9–17 (2012).
Ayala, D. et al. Chromosome inversions and ecological plasticity in the main African malaria mosquitoes. Evolution (N. Y). 71, 686–701 (2017).
Pombi, M. et al. Dissecting functional components of reproductive isolation among closely related sympatric species of the Anopheles gambiae complex. Evol. Appl. 10, 1102–1120 (2017).
Pombi, M. et al. The Sticky Resting Box, a new tool for studying resting behaviour of Afrotropical malaria vectors. Parasit. Vectors 7, 247 (2014).
Lehmann, T. & Diabate, A. The molecular forms of Anopheles gambiae: A phenotypic perspective. Infect. Genet. Evol. 8, 737–746 (2008).
Coluzzi, M., Sabatini, A., della Torre, A., Di Deco, M. A. & Petrarca, V. A Polytene Chromosome Analysis of the Anopheles gambiae Species Complex. Science (80-.). 298, 1415–1418 (2002).
Ranson, H. & Lissenden, N. Insecticide Resistance in African Anopheles Mosquitoes: A Worsening Situation that Needs Urgent Action to Maintain Malaria Control. Trends Parasitol. 32, 187–196 (2016).
Ouedraogo, A. L. et al. Efficacy and safety of the mosquitocidal drug ivermectin to prevent malaria transmission after treatment: A double-blind, randomized, clinical trial. Clin. Infect. Dis. 60, 357–365 (2015).
Churcher, T. S., Lissenden, N., Griffin, J. T., Worrall, E. & Ranson, H. The impact of pyrethroid resistance on the efficacy and effectiveness of bednets for malaria control in Africa. Elife 5, e16090 (2016).
Carnevale, P. & Robert, V. Les anophèles Biologie, transmission du Plasmodium et lutte antivectorielle. IRD éditions (2009).
Guelbeogo, W. M., Sagnon, N., Liu, F., Besansky, N. J. & Costantini, C. Behavioural divergence of sympatric Anopheles funestus populations in Burkina Faso. Malar. J. 13, 65 (2014).
Lefèvre, T. et al. Beyond nature and nurture: Phenotypic plasticity in blood-feeding behavior of Anopheles gambiae s.s. when humans are not readily accessible. Am. J. Trop. Med. Hyg. 81, 1023–1029 (2009).
Dambach, P. et al. Nightly Biting Cycles of Anopheles Species in Rural Northwestern Burkina Faso Nightly Biting Cycles of Anopheles Species in Rural Northwestern Burkina Faso. 1–8 (2018).
Kent, R. J. & Norris, D. E. Identification of mammalian blood meals in mosquitoes by a multiplexed polymerase chain reaction targeting cytochrome B. Am. J. Trop. Med. Hyg. 73, 336–42 (2005).
Gibson, G. Genetics, ecology and behaviour of anophelines. Ciba Found. Symp. 200, 22-37-47 (1996).
Dekker, T., Takken, W. & Braks, M. A. Innate preference for host-odor blends modulates degree of anthropophagy of Anopheles gambiae sensu lato (Diptera: Culicidae). J. Med. Entomol. 38, 868–71 (2001).
Pates, H. V., Takken, W., Stuke, K. & Curtis, C. F. Differential behaviour of Anopheles gambiae sensu stricto (Diptera: Culicidae) to human and cow odours in the laboratory. Bull. Entomol. Res. 91, 289–96 (2001).
Busula, A. O. et al. Mosquito host preferences affect their response to synthetic and natural odour blends. Malar. J. 14, 133 (2015).
Killeen, G. F. Characterizing, controlling and eliminating residual malaria transmission. Malar. J. 13, 330 (2014).
Meyers, J. I. et al. Increasing outdoor host-seeking in Anopheles gambiae over 6 years of vector control on Bioko Island. Malar. J. 15, 239 (2016).
Koella, J. C., Sørensen, F. L. & Anderson, R. A. The malaria parasite, Plasmodium falciparum, increases the frequency of multiple feeding of its mosquito vector, Anopheles gambiae. Proc. Biol. Sci. 265, 763–8 (1998).
Nyasembe, V. O. et al. Plasmodium falciparum infection increases Anopheles gambiae attraction to nectar sources and sugar uptake. Curr. Biol. 24, 217–21 (2014).
Smallegange, R. C. et al. Malaria Infected Mosquitoes Express Enhanced Attraction to Human Odor. PLoS One 8, 8–10 (2013).
Cator, L. J. et al. ‘Manipulation’ without the parasite: altered feeding behaviour of mosquitoes is not dependent on infection with malaria parasites. Proc. Biol. Sci. 280, 20130711 (2013).
Coetzee, M., Wilkerson, R., della Torre, A., Coulibaly, M. B. & Besansky, N. J. Anopheles coluzzii and Anopheles amharicus, new members of the Anopheles gambiae complex. Zootaxa 3619, 246–274 (2013).
Calzetta, M. et al. A novel nested polymerase chain reaction assay targeting Plasmodium mitochondrial DNA in field-collected Anopheles mosquitoes. Med. Vet. Entomol. https://doi.org/10.1111/mve.12293 (2018).
Killeen, G. F. & Smith, T. A. Exploring the contributions of bed nets, cattle, insecticides and excitorepellency to malaria control: a deterministic model of mosquito host-seeking behaviour and mortality. Trans. R. Soc. Trop. Med. Hyg. 101, 867–880 (2007).
Dabiré, K. R. et al. Distribution of pyrethroid and DDT resistance and the L1014F kdr mutation in Anopheles gambiae s.l. from Burkina Faso (West Africa). Trans. R. Soc. Trop. Med. Hyg. 103, 1113–1120 (2009).
Toé, K. H. et al. Increased pyrethroid resistance in malaria vectors and decreased bed net effectiveness Burkina Faso. Emerg. Infect. Dis. 20, 1691–1696 (2014).
Hien, A. S. et al. Evidence that agricultural use of pesticides selects pyrethroid resistance within Anopheles gambiae s.l. populations from cotton growing areas in Burkina Faso, West Africa. PLoS One 12, 1–15 (2017).
Badolo, A. et al. Three years of insecticide resistance monitoring in Anopheles gambiae in Burkina Faso: resistance on the rise? Malar. J. 11, 232 (2012).
Pombi, M. et al. Evaluation of a protocol for remote identification of mosquito vector species reveals BG-Sentinel trap as an efficient tool for Anopheles gambiae outdoor collection in Burkina Faso. Malar. J. 14 (2015).
Churcher, T. S., Trape, J.-F. & Cohuet, A. Human-to-mosquito transmission efficiency increases as malaria is controlled. Nat. Commun. 6, 6054 (2015).
Gillies, M. & Coetzee, M. A Supplement to the Anophelinae of Africa South of the Sahara. Publ. South African Inst. Med. Res. 55, 63 (1987).
Viechtbauer, W. Conducting Meta-Analyses in R with the metafor Package. J. Stat. Softw. 36, 1–48 (2010).
R Core Team. R: A language and environment for statistical computing. doi:ISBN 3-900051-07-0 (2013).
Higgins, J. P., Thompson, S. G., Deeks, J. J. & Altman, D. G. Measuring inconsistency in meta-analyses. BMJ 327, 557–60 (2003).
Smithson, M. & Verkuilen, J. A better lemon squeezer? Maximum-likelihood regression with beta-distributed dependent variables. Psychol. Methods 11, 54–71 (2006).
StatsDirect Ltd. StatsDirect statistical software (2013).
Bates, D., Maechler, M., Bolker, B. & Walker, S. lme4: linear mixed-effects models using S4 classes. R package version 1.1-6. R, http://CRAN.R-project.org/package=lme4 (2014).
Cribari-Neto, F. & Zeileis, A. Beta regression in R. J. Stat. Softw. 34, 24 (2010).
Wickham, H. The Split-Apply-Combine Strategy for Data. J. Stat. Softw. 40, 1–29 (2011).
Acknowledgements
We thank the inhabitants of Goden village for their collaboration during the whole course of this study. The work was supported by the EC FP7 Project grant no: 265660 “AvecNet”.
Author information
Authors and Affiliations
Contributions
M.P., W.M.G., N.F. H.R. and A.d.T. designed the research. M.P. and W.M.G. collected the samples. M.C., E.P., V.P. and E.M. performed molecular analyses. M.P. and M.M. analysed the data. M.P., H.R. and A.d.T. wrote the manuscript.
Corresponding author
Ethics declarations
Competing Interests
The authors declare no competing interests.
Additional information
Publisher's note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Pombi, M., Calzetta, M., Guelbeogo, W.M. et al. Unexpectedly high Plasmodium sporozoite rate associated with low human blood index in Anopheles coluzzii from a LLIN-protected village in Burkina Faso. Sci Rep 8, 12806 (2018). https://doi.org/10.1038/s41598-018-31117-x
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-018-31117-x
This article is cited by
-
The interplay between malaria vectors and human activity accounts for high residual malaria transmission in a Burkina Faso village with universal ITN coverage
Parasites & Vectors (2023)
-
The resting behavior of malaria vectors in different ecological zones of Ghana and its implications for vector control
Parasites & Vectors (2022)
-
Quantifying individual variability in exposure risk to mosquito bites in the Cascades region, Burkina Faso
Malaria Journal (2021)
-
Laboratory and field evaluation of MAÏA®, an ointment containing N,N-diethyl-3-methylbenzamide (DEET) against mosquitoes in Burkina Faso
Malaria Journal (2021)
-
Behavioural plasticity of Anopheles coluzzii and Anopheles arabiensis undermines LLIN community protective effect in a Sudanese-savannah village in Burkina Faso
Parasites & Vectors (2020)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
