
Abstract
The mangrove ecosystem harbors a complex microbial community that plays crucial role in biogeochemical cycles. In this study, we analyzed mangrove sediments from India using de novo whole metagenome next generation sequencing (NGS) and compared their taxonomic and functional community structures to mangrove metagenomics samples from Brazil and Saudi Arabia. The most abundant phyla in the mangroves of all three countries was Proteobacteria, followed by Firmicutes and Bacteroidetes. A total of 1,942 genes were found to be common across all the mangrove sediments from each of the three countries. The mangrove resistome consistently showed high resistance to fluoroquinolone and acriflavine. A comparative study of the mangrove resistome with other ecosystems shows a higher frequency of heavy metal resistance in mangrove and terrestrial samples. Ocean samples had a higher abundance of drug resistance genes with fluoroquinolone and methicillin resistance genes being as high as 28.178% ± 3.619 and 10.776% ± 1.823. Genes involved in cobalt-zinc-cadmium resistance were higher in the mangrove (23.495% ± 4.701) and terrestrial (27.479% ± 4.605) ecosystems. Our comparative analysis of samples collected from a variety of habitats shows that genes involved in resistance to both heavy metals and antibiotics are ubiquitous, irrespective of the ecosystem examined.
Similar content being viewed by others


Introduction
Mangroves are estuarine ecosystems composed of saline tolerant plants and are found in 60–70% of the coastal areas, exclusively in tropical and subtropical regions1. They are exposed to fresh and oceanic water, experiencing a wide variation of salinity throughout the tidal cycles2. Mangroves are important as they are a rich reservoir of microbial diversity and act as a buffer zone between land and sea. Furthermore, mangroves are also a source of novel enzymes and small biomolecules such as LipA-like lipase3, aspergilumamide-A peptide4, pyrrolizidine alkaloid penibruguieramine-A5, GH44 family endoglucanase6, pullularins E, F peptides7 and salt-tolerant endo-β-1, 4-glucanase Cel5A8. They also serve as a potential phytostabilizer to absorb heavy metal pollutants in industrial areas9. In addition, recent studies have shown that mangroves can enhance fish abundance10 and provide an optimal environment for microbial communities, which, in turn, help in nutrient recycling, by sulphate-reducing bacteria (SRB), methanogenic archaea11. Unfortunately, mangroves are under the threat of extinction, having experienced 35% in habitat loss in the last quarter century due to human activities12,13. In spite of the need for extensive studies on mangroves microbial community, they have largely been neglected14,15.
As most microorganisms are unculturable, traditional culture-dependent and fingerprinting methods have been inadequate in accessing the taxonomic and functional diversity of these ecosystems1,16. There are few metagenomic studies about the microbial communities in mangrove from the Brazil17, India18 and the Red Sea region of Saudi Arabia of the grey mangrove Avicennia marina19. Our recent report on mangrove ecosystem has focused on microbial community structure and an overview on functional capabilities18.
The primary objective of our work is to compare the structure and function of the biotic communities of mangroves in India, Brazil, and Saudi Arabia. The following topics are addressed by our study: (i) the comparison of the taxa composition, richness and relative abundance among the study areas; and (ii) functional diversity analyses for the gene composition, richness and relative abundance of genes among the study areas. A robust analysis was performed for preferential metabolic process, drug and heavy metal resistomes, that were further compared among distinct ecosystems.
Materials and Methods
Sampling
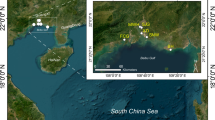
The samples were collected in 2015 during the month of mid-December in the following four sites Kumbla (KMA) (N12°35′39.101″, E74°56′47.842″), Valpadananam (VPM) (N9° 59′ 47.636″, E76° 14′ 49.882″), Kavayi (KAY) (N12° 5′ 17.83″, E75° 10′ 33.706″) and Bangramanjeshwar (BHN) (N12° 42′ 29.998″, E74° 54′ 2.716″), all of which are located within Kerala, India. All the samples had indirect exposure to anthropogenic activities resulting from household drainage. Three soil subsamples (~250 g) of the mangrove rhizosphere from the upper 20 cm depth were collected from each site using both hand gloves and sterilized polythene bags. All the samples were transported to the laboratory and processed within 48 hours.
Metagenomic DNA extraction
Metagenomic DNA from the samples was extracted using soil extraction kits (MoBio PowerSoil). There was deviation from the manufacturer’s instructions, with the elution time extended to 30 minutes at 37 °C. The subsamples in triplicates from the same collection site were pooled and were sequenced using Illumina HiSeq platform 2500 at SciGenome Labs Pvt Ltd, Cochin (India).
Quality control and annotation pipeline of the indian samples
The raw fastQ reads of Kumbla (KMA), Valpadananam (VPM), Kavayi (KAY) and Bangramanjeshwar (BHN) samples were uploaded to the Metagenome Rapid Annotation using Subsystem Technology (MG-RAST) server (http://metagenomics.anl.gov/)20 for analysis. The pipeline, in brief, joins the mate pairs and trims off low quality regions using SolexaQA21 followed by dereplication and artificially duplicated reads (ADRs) analysis22 using DRISEE (Duplicate Read Inferred Sequencing Error Estimation)23. Sequences showing similarity to fly, mouse, cow and humans were removed using Bowtie24. Annotation was done against the RefSeq25 and Subsystems database26,27 for diversity and functional analysis, respectively.
Comparative analysis of metagenomes across distinct mangrove areas
The metagenomic data from this study was compared to samples from Brazil17 and Saudi Arabia19, both of which are available at MG-RAST server (Table 1). Brazilian mangrove data17 consisted of four different samples that had different anthropogenic impacts. The BrMgv01 and BrMgv02 samples were obtained from two different sites that had experienced an oil spill in 1983. The first sample did not show any strong effects from the oil spill but the second sample still show oil effects. BrMgv3 was collected in a site near an urban area while the last sample, BrMgv04, was isolated from what was determined to be pristine conditions. A second dataset collected in Saudi Arabia also had a total of four samples (RSMgr01, RSMgr02, RSMgr03 and RSMgr04) collected from the rhizosphere of Avicennia marina, commonly known as the grey mangrove, in the Red Sea19. All samples are available at MG-RAST.
Comparative analysis of resistance to antibiotics and heavy metals in various ecosystems
For resistome analysis, the twelve mangrove datasets as described above and four mangrove datasets from our previous study18 were included. In addition, datasets collected from soil samples in four agricultural and adjacent grassland samples from Sweden28 were added. Six different forest soil samples from Puerto Rico29 and USA30 were included, as were sixteen oceanic soil samples from the Global Ocean Sampling Expedition31,32 (Table 1).
Analysis of metagenomic data
Venn diagrams were generated using the Venny 2.0 program33. Normality testing using the Shapiro-Wilk test and the Kruskal-Wallis “Nemenyi” tests was performed to evaluate whether the OTU’s and functional genes abundances were different within and between the ecosystems using R34 and PAST35. The dataset was standardized dividing each OTU abundance value by the sum of all abundances in each sample. A Principal Component Analysis (PCA) was used to compare the microbiota of each site using the vegan package36. Multiple linear regressions were conducted with the first two PCs obtained in the PCA analysis and the site as an independent variable. All the analyses were performed using R software34.
Availability of data and materials
The raw fastQ files for the Kerala India samples were uploaded in MG-RAST server and publicly available from the MG-RAST server under the following IDs: mgm4667575.3, mgm4667708.3, mgm4667773.3, and mgm4667861.3. The other publicly available samples used in this study were obtained from the MG-RAST server under the following ids: mgm4671368.3, mgm4671369.3, mgm4671370.3, mgm4671371.3, mgm4451033.3, mgm4451034.3, mgm4451035.3, mgm4451036.3, mgm4523017.3, mgm4523018.3, mgm4523019.3, mgm4523020.3, mgm4441570.3, mgm4441585.3, mgm4441590.3, mgm4441591.3, mgm4441595.3, mgm4441596.4, mgm4441599.3, mgm4441601.3, mgm4441610.3, mgm4441611.3, mgm4441613.3, mgm4441614.3, mgm4441618.3, mgm4441661.3, mgm4441662.3, mgm4442626.3, mgm4446153.3, mgm4477807.3, mgm4477875.3, mgm4477876.3, mgm4477877.3, mgm4477899.3, mgm4528934.3, mgm4529786.3, mgm4529373.3, mgm4527652.3, mgm4528937.3, mgm4529787.3, mgm4529374.3, mgm4527653.3.
Results
The sequencing from the four India datasets resulted in a total of 9 GB, of which 32,080,253 reads were obtained with an average length ranging from 252 ± 9 bp to 409 ± 139 bp. After quality control pipeline, 13 ± 3.08% reads were assigned to ribosomal RNA genes, 38.56 ± 4.41% to predicted proteins with known functions, and 48.62 ± 6.45% to predicted proteins with unknown function (hypothetical proteins) (Table 2).
Analysis at domain level
All the samples had sequences that map to the Bacteria, Archaea, Eukarya and Viruses. A small percentage of the sequences (0.011 to 0.75%) were not assigned to any organism. Bacteria were the most abundant domain recovered from all the mangrove datasets, ranging from 94.8 to 99.2% of the total. Regardless of the low sequence proportion compared to other domains, the number of sequences affiliated with viruses was the highest in Saudi Arabia samples (Fig. 1B).

Doughnut chart representing the distribution of domain, phyla and genus of (A) Brazil (BRMgv-1, BRMgv-2, BrMgv-3 and BrMgv-4) (B) Saudi Arabia (RSMgr01, RSMgr02, RSMgr03 and RSMgr04) and (C) India (MG_KAY, MG_VPM, MG_BNH and MG_KMA) (sample labels from inside to outside in doughnut wheels).
The first two components in the PCA explained more than 98% of variation and there was a clear separation among the samples (Fig. 2A). To determine if the separation among mangrove samples isolated from the different countries were statistically significant, the scores of the first two PC (Principal Component) were used as dependent variables in the multiple linear regression. The clustering effect in the first PC was due to the community abundance at domain level from India. On the other hand, all communities were different when analyzed by the second PC (Table 3). Although the reads mean frequency of Bacteria was not statistically different among the countries, the higher proportion in Indian samples (96.7–99.2%) could explain the separation of this country from the other two (Brazil: 95.1–96.7%; Saudi Arabia: 94.7–96.2%) in the first PC.

PCA plot of domain and bacterial phyla.
Analysis at phylum level
A total of 66 phyla were recovered from all samples. The richness at phylum level was quite similar across the geographic localities, except for the following Eukarya phyla: Annelida, Brachiopoda, Chytridiomycota, Echiura, Entoprocta, Glomeromycota and Xenoturbellida, which were found exclusively in the Indian samples. The Kruskal-Wallis comparison of the reads abundances among the countries indicated that Brazil (Fig. 1A) had 23 and 11 phyla statistically different from Saudi Arabia and India (Fig. 1C), respectively. Only 5 phyla were statistically different between Saudi Arabia and India. Twenty-eight bacterial phyla were retrieved from all mangroves of the three different countries. The most abundant phylum among the samples was Proteobacteria, which accounted for 50.7 to 64.28% of the sequences in Brazil, 62.6 to 64.2% in Saudi Arabia and 56.7 to 90.5% in India. Firmicutes and Bacteroidetes were the second or the third most frequent bacterial phyla recovered. The PCA using only the frequency of the bacteria phyla showed a clear separation of India from Brazil and Saudi Arabia mainly due to Proteobacteria and Bacteroidetes reads abundance (Fig. 2B).
Five archaeal phyla (Crenarchaeota, Korarchaeota, Thaumarchaeota and Nanoarchaeota) were recovered from all samples in all countries (Fig. 3A). The Crenarchaeota abundance was statistically higher in Brazilian samples than in the other countries (BR-SA: P = 0.043; BR- SA: P = 0.021) while Euryarchaeota (P = 0.021) and Korarchaeota (P = 0.043) abundances were statistically lower in India than in Brazil. Euryarchaeota was the most abundant among all archaeal phyla between and within all samples.

Bar chart of archaeal (A) phyla and (B) genera.
Overview of the dominant bacterial and archaeal genera
A total of 593 bacterial and 61 archaeal genera were recovered from all the collection sites (Supplementary Data 1). Most of the bacterial genera were present in all samples (Fig. 4), and all archaeal genera were obtained from all of the geographic locations.

Venn diagram representing the common microbiome diversity within the samples of (A) Brazil (B) Saudi Arabia (C) Kerala and (D) among the entire three sample group.
To get a clear picture of the dominant bacterial community, only those bacterial genera with more than 1% abundance in at least one sample were selected for further analysis. Fifty-four bacterial genera met this criterion (Fig. 1A–C). Most of these genera belonged to Proteobacteria followed by Bacteroidetes and Firmicutes. Other phyla detected in decreasing order of abundance were Actinobacteria, Chloroflexi and Cyanobacteria. Since Archaea were found less frequently compared to bacteria, dominant archaeal genera were examined when they were 0.1% or more of the total sample population. Twenty archaeal genera (Fig. 3B) met this criterion, with Methanosarcina, Nitrosopumilus, Thermococcus, Pyrococcus, Archaeoglobus and Methanocaldococcs being the most abundant. The Indian sample MG_BNH did not have any archaeal genera with abundance higher than 0.1%.
Comparative functional analysis of mangrove sediments
A total of 7410 protein coding genes were annotated, with 1942 found in all samples and 1023 in only one sample (Supplementary Data 2). The comparison of the top 25 most abundant functional genes from all the samples consisted of 65 genes (Fig. 5). Protein metabolism was the most diversified function with 11 different sub-functions, which includes ATP-dependent protease La (EC 3.4.21.53) (0.282% ± 0.077) and diverse tRNA synthetase, such as valyl (EC 6.1.1.9) (0.266% ± 0.037), glycyl (EC 6.1.1.14) (0.089% ± 0.08), leucyl (EC 6.1.1.4) (0.246% ± 0.049) and lysyl (class II) (EC 6.1.1.6) (0.154% ± 0.068). Within the DNA replication and transcription functional category, DNA primase (EC 2.7.7.-) (0.13% ± 0.08) and DNA-directed RNA polymerase beta subunit (EC 2.7.7.6) (0.471% ± 0.13) were found most frequently. The most common gene in the Cell Division and Cell Cycle functional category was the carbamoyl-phosphate synthase large chain (EC 6.3.5.5) (0.369% ± 0.09). The metabolism of aromatic compounds through Long-chain-fatty-acid-CoA ligase (EC 6.2.1.3) exhibited a similar pattern of low frequency between MG_VPM (0.165%) and the MG_BNH (0.206%) while the remaining samples showed higher level of abundance (0.45% ± 0.05). Kruskal-Wallis comparison of the most abundant functional genes also indicated statistically significant (p < 0.05) differences between AS, BR and ID (Supplementary Data 3).

The top 25 most abundant functions from each sample annotated against subsystems.
There were also three genes related to antibiotics resistance and toxic compounds: Cation efflux system protein CusA (0.311% ± 0.101), Acriflavine resistance protein (0.428% ± 0.0843) and Topoisomerase IV subunit A (EC 5.99.1.3) (0.136% ± 0.073). Cation efflux system protein CusA is involved in resistance to copper and silver while Acriflavine resistance protein and Topoisomerase IV subunit A are involved in resistance to antiseptic Acriflavine and fluoroquinolones antibiotics respectively, which are both clinically relevant37,38,39.
Overall, the DNA-directed RNA polymerase beta subunit (EC 2.7.7.6) (0.471% ± 0.131) was the most abundant gene followed by Acriflavine resistance protein (0.42% ± 0.08). Within the resistome, Acriflavine resistance protein was the most abundant followed by Cation efflux system protein CusA and Topoisomerase IV subunit A as the 2nd and 3rd most abundant features. All the samples between and within the group of India, Brazil and Saudi Arabia showed high abundance (0.428% ± 0.08) of Acriflavine resistance protein with no significant difference within the samples (p 0.4433).
Comparative functional analysis across different ecosystems
The high abundance of acriflavine resistance protein and the widespread presence of genes related to fluoroquinolone resistance in all the mangroves samples was intriguing and resulted in a deeper examination of the resistance to antibiotics and toxic compounds in the mangroves and other ecosystems. Publicly available metagenomes of marine and terrestrial (forest, grassland and agricultural soil samples) ecosystems were compared to the mangroves sediments (Table 1). Remarkable patterns were observed across the ecosystems showing sharp distinction between the terrestrial and aquatic sites. Enrichment of Multidrug Resistance Efflux Pumps were similar in oceans (23.274% ± 2.931) and mangroves (25.406% ± 2.922) although statistically different (p = 0.028). Interestingly, terrestrial samples had a much lower abundance (14.897% ± 4.116) (p < 0.005) of Multidrug Resistance Efflux Pumps (Fig. 6A). However, a deeper look into the functional level of Multidrug Resistance Efflux Pumps shows acriflavine resistance protein to be highly enriched in all the samples irrespective of ecosystem and anthropogenic activities. The relative abundance of the acriflavine resistance was similar in mangroves and terrestrial which were in turn significantly different from Ocean (p < 0.005) (Fig. 6B). Similarly, other clinically relevant antibiotic resistance genes (ARGs) such as those related to resistance to fluoroquinolones and beta-lactamase were significantly higher (p < 0.005) in ocean (28.178% ± 3.619, 9.913% ± 2.208) compared to mangroves (9.82% ± 3.776, 5.489% ± 0.742) and terrestrial (11.18% ± 8.327, 10.247% ± 5.826). Methicillin resistance was found to be statistically different in all the ecosystems although the relative percentage was more similar between mangrove (3.034% ± 0.808) and terrestrial (2.159% ± 0.682) as compared to Ocean (10.776% ± 1.823) respectively. In addition, resistance genes related to heavy metals such as cobalt, zinc and cadmium were significantly (p < 0.015) different in Ocean, Mangroves, and Terrestrial ecosystems (Supplementary data 4).

(A) Bar chart of resistance to antibiotics and toxic compounds and (B) Multidrug Resistance Efflux Pumps in Ocean, Mangroves and Land (Forest, Agriculture and Grassland).
Discussion
In this study, whole-metagenome of Indian mangrove samples were sequenced to examine the community structure and functional content using the Illumina technology. These were compared to samples isolated from mangroves in Brazil17 and Saudi Arabia19, which were sequenced by a different platform (Roche 454). Although the three datasets were generated by two different NGS platforms, our analysis showed that they had similar taxonomic diversity in their microbial communites40.
Bacterial and archaeal diversity in the mangrove sediments
The bacterial phyla, Proteobacteria (61.2% ± 10.82), was the most abundant in the samples examined from each of the geographic locations (Fig. 1A–C). Proteobacteria had previously been noted as being highly abundant in mangrove samples41. This phylum has a high metabolic diversity, with a wide distribution in marine environments, playing an important role in nutrient cycling42. The second most frequently found phyla within the mangrove metagenomics samples we examined was Archaea. Members belonging to this phylum inhabit extreme environments, playing important roles in the biogeochemical cycles. However, our knowledge as to the niche they occupy, and the role they play in the mangrove microbial community is still limited43. Within the archaeal kingdom, Euryarchaeota (81.29% ± 7.993) were found most frequently within and between all the samples when compared to other Archaeal phyla (Fig. 3), and were also found to be highly abundant in mangrove sediments of Sundarbans (India)43 and other ecosystems such as marine sediment (North Sea of Atlantic Ocean)44 and German bight45 (a shallow region of the North Sea that borders Germany). The archaeal community we found in the mangroves had many methanogen genera, including Methanothermobacter, Methanocaldococcus, Methanococcus, Methanosarcina, Methanococcoides, Methanospirillum, Methanoculleus, Methanosaeta, and Methanoregula. The diversity and presence of these specific genera is an indication that methane metabolism is important in mangrove ecosystems. The archaeal ammonia oxidizer Nitrosopumilus (0.15% ± 0.2) and Cenarchaeum (0.03% ± 0.04) of the phylum Thaumarchaeota was also observed in all samples.
At the bacterial genus level, Pseudomonas (2.61% ± 1.7) was found to be most abundant in MG_BNH (8.06%) (India). Pseudomonas spp. thrives in many diverse environments that range from individual humans to the rhizosphere46. Pseudomonas has been found to influence plant growth19,46,47 by releasing siderophores, antibiotics, biosurfactants and solubilization of potassium into forms that are accessible for plants, and has also been noted for its critical nitrogen fixation role in the mangrove ecosystem48,49. In addition, they are also tolerant to aromatic hydrocarbons, organic and heavy metal contaminants50,51. Another aromatic hydrocarbons degrader, Geobacter, was found at a similar frequency in the samples from Brazil (2.65% ± 0.08) and Saudi Arabia (2.05% ± 0.6). Its presence in the Indian samples (1.03% ± 0.53) differed statistically (p 0.02) from the samples of the other two countries. Geobacter has also been found in petroleum contaminated environments and pristine deep aquifers capable of Fe (III) reduction52 and are also strong candidates for immobilization uranium53 (U(VI)) and bioremediation of aromatic hydrocarbon contaminants. Neptuniibacter, a copiotrophic microorganism which can degrade aromatic hydrocarbon such as carbazole54, was the 3rd most abundant genus in all the samples (1.61% ± 4.61). The high standard deviation was due to the overwhelmingly high frequency in MG_BNH (16.91%) (Indian sample) that could be due to this region having higher anthropogenic activity when compared to the other samples. In addition, Marinobacter, a ubiquitous marine aromatic hydrocarbon degradation genus55 of the Proteobacteria phylum, was significantly (p 0.0124) abundant in the Indian (3.15% ± 2.23) samples compared to Brazil (0.44% ± 0.04) and Saudi Arabia (0.82% ± 0.35). Our previous study also showed a dominance of Marinobacter in mangrove samples18. Genome analysis of Marinobacter has revealed its potentiality to survive in oil-polluted water55 and has suggested that it could be used for bio-monitoring of oil spills in mangroves56. Two important genera from the phylum Actinobacteria, Streptomyces and Mycobacterium, were found. Mycobacterium were found at more significant frequency (p 0.0209) in Brazil (1.17% ± 0.34) than in Saudi Arabia (0.35% ± 0.03) or India (0.35% ± 0.16). Interestingly, a pristine Brazilian sample (BRMgv-1: 1.72%) and one that was highly impacted by human activity (BRMgv-2: 0.98%) had a higher frequency of Mycobacterium compared to the samples associated with anthropogenic activity (BRMgv-3: 1.19%), or to the sample from pristine (BRMgv-4: 0.79%) environment. It is interesting that Mycobacterium spp., such as M. chlorophenolicum, M. farcinogenes and M. austroafricanum, were observed in samples from mangrove sediments that were contaminated with PAH (Polycyclic aromatic hydrocarbon)57. Seven genera belonging to order Desulfobacterales were found in all samples and showed similar frequency across all samples. Desulfotalea is a psychrophilic genus58. Desulfovibrio is known to be aerotolerant59. Geobacter is recognized as an aromatic hydrocarbons degrader60 and Pelobacter is an iron and sulfur-reducing mesophilic anaerobe61. These were significantly less abundant (p 0.0209, 0.034, 0.026 and 0.037 respectively) in Indian samples when compared to Brazil and Saudi Arabia. An ammonia oxidizing bacteria, Nitrosococcus (0.97% ± 0.28), was present in all the samples with no significant difference within or between the groups. Samples collected from mangroves in each of these countries shared a total of 97.9% of the OTUs, which accounted for 99.97% of the total reads abundance (Fig. 4). Similar results were obtained when compared to forest and vineyard soils62. The high number of shared OTUs between the mangroves corroborates the functional genes statistical analysis (Supplementary data 3) between the samples.
Resistance to antibiotics and heavy metals in various ecosystems
High abundance of fluoroquinolones and acriflavine resistance proteins were found in the mangrove samples of India, Brazil and Saudi Arabia irrespective of the collection site. In order to examine the consistency of these resistance genes in other ecosystems, whole metagenomic datasets from others studies, including18, ocean31,32, forest29,30 agricultural and grassland soil samples28 were compared, specifically targeting marine and terrestrial environments with and without anthropogenic activity (Table 1).
Heavy metal resistome in diverse ecosystems
The presence of genes involved in heavy metal resistance in rivers, activated sludges, aquaculture farm sediments, etc.63,64 have been previously described. In our study, genes that play a role in the resistance to antibiotics and toxic compounds were found across all of the ecosystems. Cobalt-zinc-cadmium resistance were found in all the samples, but the percentage of reads that mapped to these genes was found to be significantly lower (p < 0.01) in the samples collected from the ocean (5.713% ± 2.589) compared to those collected from mangroves (23.495% ± 4.701) or terrestrial (27.479% ± 4.605) ecosystems (Fig. 6). Among the genes that determine cobalt-zinc-cadmium resistance, the cation efflux system protein CusA was the most abundant gene. CusA and the cation efflux system provide bacteria with resistance to copper and silver. Although copper is an essential element, it can be lethal to plants even at low concentrations65 and can lead to several ill effects such as chlorosis, yellow coloration, and retardation of growth66. This copper resistance symbiotic bacterium is associated with plants found in mine tailings67. Metal ion solubility generally increases with decreasing pH68, and the presence of CusA has been found to be associated with soil types with low pH68. Marine samples showed significantly lesser enrichment of CusA (5.37% ± 3.55), which could be due to the higher pH of marine water69 and the lack of plants in this ecosystem. Another annotated function that was seen involved copper homeostasis, but all the ecosystems exhibited comparable level of this particular functionality (7.068% ± 1.154, 6.058% ± 1.343 and 7.238% ± 2.116 for marine, mangroves and terrestrial, respectively). Arsenic resistance genes were also found consistently across all the samples (Fig. 6) having significant difference in ocean vs. mangrove (p < 0.001) and terrestrial vs. mangrove (p < 0.002) ecosystem (Supplementary data 4). A recent study by Xiao et al.70 demonstrated a similar presence of genes involved in arsenic metabolism in paddy soil, with the authors concluding that these genes play an important role in avoiding arsenic risk through biotransformation.
Antibiotic resistance genes (ARGs) patterns across ecosystems
Genes involved in antibiotic resistance have been observed in distinct patterns across different ecosystems71. In our study, the Multidrug Resistance Efflux Pumps functional category was the most abundant drug resistance function across all the ecosystems (Fig. 6A). Interestingly, among the subtypes within this functional category, Acriflavine resistance proteins were significantly abundant in every sample (Fig. 6B). Acriflavine has antibacterial properties and is used as an antibiotic72. It has been shown to have antiviral and antitumor activities through its topoisomerase inhibition properties73,74. The widespread prevalence of acriflavine resistance was also observed in the past from clinical samples in 11 Asian countries75. In the recent years, metagenomic analyses showed that acriflavine resistance genes were highly abundant in South China paddy soil76 and aerobic activated sludge and anaerobically digested sludge77. In our analysis, we found that the acriflavine resistance genes were widespread in aquatic and terrestrial ecosystems that had significant human activity or were from pristine environments (Fig. 6A and B). Fluoroquinolone drugs, which target DNA gyrase and topoisomerase IV, are widely used as the first line for nosocomial infections78,79. Fluoroquinolone as well as methicillin resistance gene were found to be significantly higher in marine (28.178% ± 3.619 and 10.776% ± 1.823 respectively, p < 0.001) as compared to mangroves (9.82% ± 3.776 and 3.034% ± 0.808, respectively) and terrestrial (11.18% ± 8.327 and 2.159% ± 0.682, respectively) ecosystems.
Beta-lactamase was highly abundant in the marine (9.913% ± 2.208) and terrestrial (10.247% ± 5.826) ecosystems. Within the terrestrial samples, forest (4.38% ± 1.66) had similar abundance comparable to mangroves (5.489% ± 0.742) while the agricultural and grassland samples were found to be highly enriched (14.64% ± 3.54). Antibiotic-resistant genes have been found to be similarly abundant in soils that contain or lack manure80. Similarly, our result showed a high abundance of β-lactamases genes in agricultural and adjacent grasslands samples with comparable frequency while the forest and mangroves samples had relatively lesser abundance. The high abundance of β-lactamases in agricultural soil have been demonstrated in a recent functional metagenomic study by Lau et al.81 who identified 34 new antibiotic resistance genes that were related to multi-drug efflux systems, indicating a potential high-level resistance towards aminoglycosides, sulfonamides, and a broad range of beta-lactams. As β-lactamases have been hypothesized to play a vital role in the survival of the bacteria in its natural habitat82, the presence of the genes involved in this resistance have been noted in metagenomic samples from a variety of habitats. For instance, a proficient β-lactamase enzyme was isolated from Oceanobacillus iheyensis in the ocean sediments at a depth of 1050 meters83. Recently, a novel β-lactamase gene was discovered from Pelagibacterium halotolerans B2T, which was isolated from the East China Sea84. The high abundance of β-lactamases in oceans also indicates the rich diversity of enzymes and the promising prospects of novel antibiotic discoveries.
Antibiotics and antibiotic resistance genes have been found in diverse environments that include deep terrestrial subsurface, glacier ice core and samples collected from deep sea that have not been in contact with humans83,85,86,87,88, but they are mostly present at non-inhibitory concentrations89,90,91. The antibiotic resistance genes were dominant in the resistome having significant differences among the ecosystems with the ocean having highest relative abundance compared to mangrove and terrestrial ecosystems (Supplementary data 4). It has been hypothesized that the function of such resistance genes in the natural environment could be related to some basic physiological processes such as biosynthesis of the cell wall92,93, trafficking of signaling molecules, detoxification of metabolic intermediates88 or antibiotic detoxification88,94,95,96. Untouched environments can have novel antibiotic resistance genes97 that can give rise to more multidrug resistant strains via horizontal transfer when human activities encroach upon them. For instance, when soil samples from pre-and post-antibiotic areas were compared, plasmids from the earlier era had fewer antibiotic resistance genes97,98 and this was followed by a significant rise in their presence in later sampes99. The notion of clinically relevant pathogens acquiring resistance genes from the environment is a likely possibility97,100,101.
Conclusion
We have analysed the metagenomic profiles of mangrove sediments across India and compared them with publicly available samples from Brazil and Saudi Arabia mangrove. Distinct patterns unique to the Brazilian and Saudi Arabian mangroves were observed which differentiated them from samples collected in India. Although there were differences, a significant number of microbial genera were found to be present across all of the three geographic regions. Proteobacteria and Euryarchaeota were the most abundant phyla within and between all the mangroves for bacteria and archaea, respectively. A functional analysis that compared the mangroves samples with metagenomic sample taken from ocean, forest, agriculture and grassland showed the presence of highly enriched acrylflavine, copper, fluoroquinolone, β-lactamase and methilicin resistant genes distributed consistently in patterns throughout all the examined ecosystems. Further, our study showed that heavy metals and antibiotic resistance genes are founnd in microbial populations from mangroves and the other ecosystems, including both pristine areas and environments that experience significant human activity. The widespread existence of antibiotic resistance genes could be a warning bell, indicating a source of new genes that could further increase the rise in antimicrobial resistance that could have clinical significance.
References
Ghosh, A. et al. Culture independent molecular analysis of bacterial communities in the mangrove sediment of Sundarban, India. Saline systems 6(1), 1 (2010).
Holguin, G., Vazquez, P. & Bashan, Y. The role of sediment microorganisms in the productivity, conservation, and rehabilitation of mangrove ecosystems: an overview. Biology and fertility of soils 33(4), 265–278 (2001).
Couto, G. H. et al. Isolation of a novel lipase from a metagenomic library derived from mangrove sediment from the south Brazilian coast. Genet Mol Res 9(1), 514–523 (2010).
Zheng, C. J. et al. Structure and Absolute Configuration of Aspergilumamide A, a Novel Lumazine Peptide from the Mangrove‐Derived Fungus Aspergillus sp. Helvetica Chimica Acta 98(3), 368–373 (2015).
Zhou, Z. F. et al. Penibruguieramine A, a novel pyrrolizidine alkaloid from the endophytic fungus Penicillium sp. GD6 associated with Chinese mangrove Bruguiera gymnorrhiza. Organic letters 16(5), 1390–1393 (2014).
Mai, Z., Su, H., Yang, J., Huang, S. & Zhang, S. Cloning and characterization of a novel GH44 family endoglucanase from mangrove soil metagenomic library. Biotechnology letters 36(8), 1701–1709 (2014).
Ebrahim, W. et al. Pullularins E and F, two new peptides from the endophytic fungus Bionectria ochroleuca isolated from the mangrove plant Sonneratia caseolaris. Marine drugs 10(5), 1081–1091 (2012).
Gao, Z., Ruan, L., Chen, X., Zhang, Y. & Xu, X. A novel salt-tolerant endo-β-1, 4-glucanase Cel5A in Vibrio sp. G21 isolated from mangrove soil. Applied microbiology and biotechnology 87(4), 1373–1382 (2010).
Almahasheer, H. B., Al-Taisan, W. A. & Mohamed, M. K. Metals accumulation in grey mangrove (avicennia marina (forsk.) vierh.) inhabiting tarut bay, eastern Saudi Arabia. Journal of Agricultural Science 6(1), 137 (2013).
Serafy, J. E., Shideler, G. S., Araújo, R. J. & Nagelkerken, I. Mangroves enhance reef fish abundance at the Caribbean regional scale. PloS one 10(11), e0142022 (2015).
Taketani, R. G., Yoshiura, C. A., Dias, A. C. F., Andreote, F. D. & Tsai, S. M. Diversity and identification of methanogenic archaea and sulphate-reducing bacteria in sediments from a pristine tropical mangrove. Antonie van Leeuwenhoek 97(4), 401–411 (2010).
Duke, N. C. et al. A world without mangroves? Science 317(5834), 41–42 (2007).
UNFAO, Status and trends in mangrove area extent worldwide In, Forestry (ed): Italy (2003).
Ghizelini, A. M., Mendonça-Hagler, L. C. S. & Macrae, A. Microbial diversity in Brazilian mangrove sediments: a mini review. Brazilian Journal of Microbiology 43(4), 1242–1254 (2012).
Mendes, R., Garbeva, P. & Raaijmakers, J. M. The rhizosphere microbiome: significance of plant beneficial, plant pathogenic, and human pathogenic microorganisms. FEMS microbiology reviews 37(5), 634–663 (2013).
Basak, P. et al. Spatiotemporal analysis of bacterial diversity in sediments of Sundarbans using parallel 16S rRNA gene tag sequencing. Microbial ecology 69(3), 500–511 (2015).
Andreote, F. D. et al. The microbiome of Brazilian mangrove sediments as revealed by metagenomics. PloS one 7(6), e38600 (2012).
Imchen, M. et al. Searching for signatures across microbial communities: Metagenomic analysis of soil samples from mangrove and other ecosystems. Scientific Reports 7(1), 8859 (2017).
Alzubaidy, H. et al. Rhizosphere microbiome metagenomics of gray mangroves (Avicennia marina) in the Red Sea. Gene 576(2), 626–636 (2016).
Meyer, F. et al. The metagenomics RAST server–a public resource for the automatic phylogenetic and functional analysis of metagenomes. BMC bioinformatics 9(1), 386 (2008).
Cox, M. P., Peterson, D. A. & Biggs, P. J. SolexaQA: At-a-glance quality assessment of Illumina second-generation sequencing data. BMC bioinformatics 11(1), 485 (2010).
Gomez-Alvarez, V., Teal, T. K. & Schmidt, T. M. Systematic artifacts in metagenomes from complex microbial communities. The ISME journal 3(11), 1314–1317 (2009).
Keegan, K. P. et al. A platform-independent method for detecting errors in metagenomic sequencing data: DRISEE. PLoS computational biology 8(6), e1002541 (2012).
Langmead, B., Trapnell, C., Pop, M. & Salzberg, S. L. Ultrafast and memory-efficient alignment of short DNA sequences to the human genome. Genome biology 10(3), R25 (2009).
Wilke, A. et al. The M5nr: a novel non-redundant database containing protein sequences and annotations from multiple sources and associated tools. BMC bioinformatics 13(1), 141 (2012).
Overbeek, R. et al. The subsystems approach to genome annotation and its use in the project to annotate 1000 genomes. Nucleic acids research 33(17), 5691–5702 (2005).
Overbeek, R. et al. The SEED and the Rapid Annotation of microbial genomes using Subsystems Technology (RAST). Nucleic acids research 42(D1), D206–D214 (2013).
Manoharan, L., Kushwaha, S. K., Hedlund, K. & Ahrén, D. Captured metagenomics: large-scale targeting of genes based on ‘sequence capture’ reveals functional diversity in soils. DNA Research 22(6), 451–460 (2015).
DeAngelis, K. M. et al. Strategies for enhancing the effectiveness of metagenomic-based enzyme discovery in lignocellulolytic microbial communities. Bioenergy Research 3(2), 146–158 (2010).
Fierer, N. et al. Cross-biome metagenomic analyses of soil microbial communities and their functional attributes. Proceedings of the National Academy of Sciences 109(52), 21390–21395 (2012).
Rusch, D. B. et al. The Sorcerer II global ocean sampling expedition: northwest Atlantic through eastern tropical Pacific. PLoS biology 5(3), e77 (2007).
Williamson, S. J. et al. The Sorcerer II Global Ocean Sampling Expedition: metagenomic characterization of viruses within aquatic microbial samples. PloS one 3(1), e1456 (2008).
Oliveros, J. C. VENNY. An interactive tool for comparing lists with Venn Diagrams. http://bioinfogp.cnb.csic. es/tools/venny/index. html (2007).
Team, R. C. R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. ISBN 3-900051-07-0 (2012).
Hammer, Ø., Harper, D. A. T. & Ryan, P. D. 2001. Paleontological statistics software: Package for education and data analysis. Palaeontologia Electronica (4) (2014).
Oksanen, J. et al. The vegan package. Community ecology package 10, 631–637 (2007).
Ahmad, N., Khan, A. H., Sulaiman, S. A. S. & Javaid, A. Fluoroquinolones resistance in multidrug-resistant tuberculosis in Pakistan and suitability of guidelines recommended standardized regimen. The International Journal of Mycobacteriology 4(3), 258 (2015).
Strahilevitz, J., Jacoby, G. A., Hooper, D. C. & Robicsek, A. Plasmid-mediated quinolone resistance: a multifaceted threat. Clinical microbiology reviews 22(4), 664–689 (2009).
Lepainteur, M. et al. Prevalence of resistance to antiseptics and mupirocin among invasive coagulase-negative staphylococci from very preterm neonates in NICU: the creeping threat? Journal of Hospital Infection 83(4), 333–336 (2013).
Luo, C., Tsementzi, D., Kyrpides, N., Read, T. & Konstantinidis, K. T. Direct comparisons of Illumina vs. Roche 454 sequencing technologies on the same microbial community DNA sample. PloS one 7(2), e30087 (2012).
Mendes, L. W. & Tsai, S. M. Variations of bacterial community structure and composition in mangrove sediment at different depths in Southeastern Brazil. Diversity 6(4), 827–843 (2014).
Kersters, K. et al. Introduction to the Proteobacteria. In The prokaryotes (pp. 3–37). Springer New York (2006).
Bhattacharyya, A. et al. Diversity and distribution of Archaea in the mangrove sediment of Sundarbans. Archaea 2015 (2015).
Sapp, M., Parker, E. R., Teal, L. R. & Schratzberger, M. Advancing the understanding of biogeography–diversity relationships of benthic microorganisms in the North Sea. FEMS microbiology ecology 74(2), 410–429 (2010).
Wemheuer, B., Wemheuer, F. & Daniel, R. RNA-based assessment of diversity and composition of active archaeal communities in the German Bight. Archaea 2012 (2012).
Nikel, P. I., Martínez-García, E. & De Lorenzo, V. Biotechnological domestication of pseudomonads using synthetic biology. Nature Reviews Microbiology 12(5), 368–379 (2014).
Wu, X. et al. Comparative genomics and functional analysis of niche-specific adaptation in Pseudomonas putida. FEMS microbiology reviews 35(2), 299–323 (2011).
Flores-Mireles, A. L., Winans, S. C. & Holguin, G. Molecular characterization of diazotrophic and denitrifying bacteria associated with mangrove roots. Applied and environmental microbiology 73(22), 7308–7321 (2007).
Zhang, Y., Dong, J., Yang, Z., Zhang, S. & Wang, Y. Phylogenetic diversity of nitrogen-fixing bacteria in mangrove sediments assessed by PCR–denaturing gradient gel electrophoresis. Archives of microbiology 190(1), 19 (2008).
Jiménez, J. I., Miñambres, B., García, J. L. & Díaz, E. Genomic analysis of the aromatic catabolic pathways from Pseudomonas putida KT2440. Environmental microbiology 4(12), 824–841 (2002).
Pitondo‐Silva, A., Gonçalves, G. B. & Stehling, E. G. Heavy metal resistance and virulence profile in Pseudomonas aeruginosa isolated from Brazilian soils. Apmis 124(8), 681–688 (2016).
Coates, J. D., Phillips, E. J., Lonergan, D. J., Jenter, H. & Lovley, D. R. Isolation of Geobacter species from diverse sedimentary environments. Applied and Environmental Microbiology 62(5), 1531–1536 (1996).
Yun, J., Malvankar, N. S., Ueki, T. & Lovley, D. R. Functional environmental proteomics: elucidating the role of a c-type cytochrome abundant during uranium bioremediation. The ISME journal 10(2), 310–320 (2016).
Nagashima, H. et al. Cloning and nucleotide sequences of carbazole degradation genes from marine bacterium Neptuniibacter sp. strain CAR-SF. Current microbiology 61(1), 50–56 (2010).
Singer, E. et al. Genomic potential of Marinobacter aquaeolei, a biogeochemical “opportunitroph”. Applied and environmental microbiology 77(8), 2763–2771 (2011).
Dos Santos, H. F. et al. Mangrove bacterial diversity and the impact of oil contamination revealed by pyrosequencing: bacterial proxies for oil pollution. PloS one 6(3), e16943 (2011).
Guo, C., Ke, L., Dang, Z. & Tam, N. F. Temporal changes in Sphingomonas and Mycobacterium populations in mangrove sediments contaminated with different concentrations of polycyclic aromatic hydrocarbons (PAHs). Marine pollution bulletin 62(1), 133–139 (2011).
Yang, T. et al. Distinct bacterial communities in surficial seafloor sediments following the 2010 Deepwater Horizon blowout. Frontiers in microbiology 7 (2016).
Bryukhanov, A. L. et al. Components of antioxidant systems in the cells of aerotolerant sulfate-reducing bacteria of the genus Desulfovibrio (strains A2 and TomC) isolated from metal mining waste. Microbiology 85(6), 649–657 (2016).
Kronenberg, M., Trably, E., Bernet, N. & Patureau, D. Biodegradation of polycyclic aromatic hydrocarbons: Using microbial bioelectrochemical systems to overcome an impasse. Environmental pollution (Barking, Essex: 1987) 231(Pt 1), 509 (2017).
An, T. T. & Picardal, F. W. Desulfuromonas carbonis sp. nov., an Fe (III)-, S0-and Mn (IV)-reducing bacterium isolated from an active coalbed methane gas well. International journal of systematic and evolutionary microbiology 65(5), 1686–1693 (2015).
Castañeda, L. E. & Barbosa, O. Metagenomic analysis exploring taxonomic and functional diversity of soil microbial communities in Chilean vineyards and surrounding native forests. PeerJ 5, e3098 (2017).
Suenaga, H. et al. Novel organization of aromatic degradation pathway genes in a microbial community as revealed by metagenomic analysis. The ISME journal 3(12), 1335–1348 (2009).
Ma, L., Li, B. & Zhang, T. Abundant rifampin resistance genes and significant correlations of antibiotic resistance genes and plasmids in various environments revealed by metagenomic analysis. Applied microbiology and biotechnology 98(11), 5195–5204 (2014).
Outten, F. W., Huffman, D. L., Hale, J. A. & O’Halloran, T. V. The independent cue and cusSystems confer copper tolerance during aerobic and anaerobic growth in Escherichia coli. Journal of Biological Chemistry 276(33), 30670–30677 (2001).
Lange, B. et al. Copper and cobalt accumulation in plants: a critical assessment of the current state of knowledge. New Phytologist 213(2), 537–551 (2017).
Fan, L. M. et al. Characterization of a copper-resistant symbiotic bacterium isolated from Medicago lupulina growing in mine tailings. Bioresource technology 102(2), 703–709 (2011).
Malik, A. A., Thomson, B. C., Whiteley, A. S., Bailey, M. & Griffiths, R. I. Bacterial physiological adaptations to contrasting edaphic conditions identified using landscape scale metagenomics. bioRxiv, p.117887 (2017).
Ramos, O., Castro, S. & Laskowski, J. S. Copper–molybdenum ores flotation in sea water: Floatability and frothability. Minerals Engineering 53, 108–112 (2013).
Xiao, K. Q. et al. Metagenomic analysis revealed highly diverse microbial arsenic metabolism genes in paddy soils with low-arsenic contents. Environmental Pollution 211, 1–8 (2016).
Forsberg, K. J. et al. Bacterial phylogeny structures soil resistomes across habitats. Nature 509(7502), 612–616 (2014).
Pépin, G. et al. Activation of cGAS-dependent antiviral responses by DNA intercalating agents. Nucleic acids research 45(1), 198–205 (2017).
Hassan, S. et al. Novel activity of acriflavine against colorectal cancer tumor cells. Cancer science 102(12), 2206–2213 (2011).
Dekervel, J. et al. Acriflavine Inhibits Acquired Drug Resistance by Blocking the Epithelial-to-Mesenchymal Transition and the Unfolded Protein Response. Translational oncology 10(1), 59–69 (2017).
Noguchi, N. et al. Susceptibilities to antiseptic agents and distribution of antiseptic-resistance genes qacA/B and smr of methicillin-resistant Staphylococcus aureus isolated in Asia during 1998 and 1999. Journal of medical microbiology 54(6), 557–565 (2005).
Xiao, K. Q. et al. Metagenomic profiles of antibiotic resistance genes in paddy soils from South China. FEMS microbiology ecology 92(3), fiw023 (2016).
Guo, J., Li, J., Chen, H., Bond, P. L. & Yuan, Z. Metagenomic analysis reveals wastewater treatment plants as hotspots of antibiotic resistance genes and mobile genetic elements. Water research 123, 468 (2017).
Ereshefsky, B. J., Majdi, N. A.-H., Gokun, Y. & Martin, C. A. Comparison of ß-lactam plus aminoglycoside versus ß-lactam plus fluoroquinolone empirical therapy in serious nosocomial infections due to Gram-negative bacilli. Journal of Chemotherapy 29(1), 30–37 (2017).
Moon, D. C. et al. Emergence of a new mutation and its accumulation in the topoisomerase IV gene confers high levels of resistance to fluoroquinolones in Escherichia coli isolates. International journal of antimicrobial agents 35(1), 76–79 (2010).
Marti, R. et al. Impact of manure fertilization on the abundance of antibiotic-resistant bacteria and frequency of detection of antibiotic resistance genes in soil and on vegetables at harvest. Applied and environmental microbiology 79(18), 5701–5709 (2013).
Lau, C. H. F., van Engelen, K., Gordon, S., Renaud, J. & Topp, E. Novel antibiotic resistance determinants from agricultural soil exposed to antibiotics widely used in human medicine and animal farming. Applied and Environmental Microbiology pp.AEM-00989 (2017).
Kristiansen, A., Grgic, M., Altermark, B. & Leiros, I. Properties and distribution of a metallo-β-lactamase (ALI-1) from the fish pathogen Aliivibrio salmonicida LFI1238. Journal of Antimicrobial Chemotherapy 70(3), 766–772 (2014).
Toth, M., Smith, C., Frase, H., Mobashery, S. & Vakulenko, S. An antibiotic-resistance enzyme from a deep-sea bacterium. Journal of the American Chemical Society 132(2), 816–823 (2009).
Zheng, B., Jiang, X., Xu, Z., Fang, Y. & Li, L. Characterization of a novel metallo-β-lactamases fold hydrolase from Pelagibacterium halotolerans, a marine halotolerant bacterium isolated from East China Sea. Extremophiles 20(1), 37–44 (2016).
Brown, M. G. & Balkwill, D. L. Antibiotic resistance in bacteria isolated from the deep terrestrial subsurface. Microbial Ecology 57(3), 484 (2009).
Miteva, V. I., Sheridan, P. P. & Brenchley, J. E. Phylogenetic and physiological diversity of microorganisms isolated from a deep Greenland glacier ice core. Applied and Environmental Microbiology 70(1), 202–213 (2004).
D’Costa, V. M. et al. Antibiotic resistance is ancient. Nature 477(7365), 457–461 (2011).
Martinez, J. L. et al. Functional role of bacterial multidrug efflux pumps in microbial natural ecosystems. FEMS microbiology reviews 33(2), 430–449 (2009).
Linares, J. F., Gustafsson, I., Baquero, F. & Martinez, J. L. Antibiotics as intermicrobial signaling agents instead of weapons. Proceedings of the National Academy of Sciences 103(51), 19484–19489 (2006).
Yim, G. & Wang, H. H. & FRS, J.D. Antibiotics as signalling molecules. Philosophical Transactions of the Royal Society of London B: Biological Sciences 362(1483), 1195–1200 (2007).
Fajardo, A. & Martínez, J. L. Antibiotics as signals that trigger specific bacterial responses. Current opinion in microbiology 11(2), 161–167 (2008).
Severin, A., Wu, S. W., Tabei, K. & Tomasz, A. Penicillin-binding protein 2 is essential for expression of high-level vancomycin resistance and cell wall synthesis in vancomycin-resistant Staphylococcus aureus carrying the enterococcal vanA gene complex. Antimicrobial agents and chemotherapy 48(12), 4566–4573 (2004).
Blanco, P. et al. Bacterial multidrug efflux pumps: much more than antibiotic resistance determinants. Microorganisms 4(1), 14 (2016).
Mesak, L. R., Miao, V. & Davies, J. Effects of subinhibitory concentrations of antibiotics on SOS and DNA repair gene expression in Staphylococcus aureus. Antimicrobial agents and chemotherapy 52(9), 3394–3397 (2008).
Linares, J. F. et al. The global regulator Crc modulates metabolism, susceptibility to antibiotics and virulence in Pseudomonas aeruginosa. Environmental microbiology 12(12), 3196–3212 (2010).
Martínez, J. L. Natural antibiotic resistance and contamination by antibiotic resistance determinants: the two ages in the evolution of resistance to antimicrobials. Frontiers in microbiology 3 (2012).
Bhullar, K. et al. Antibiotic resistance is prevalent in an isolated cave microbiome. PloS one 7(4), e34953 (2012).
Hughes, V. M. & Datta, N. Conjugative plasmids in bacteria of the ‘pre-antibiotic’ era. Nature 302(5910), 725–726 (1983).
Knapp, C. W., Dolfing, J. L., Ehlert, P. A. & Graham, D. W. Evidence of increasing antibiotic resistance gene abundances in archived soils since 1940. Environmental science & technology 44(2), 580–587 (2009).
Donato, J. J. et al. Metagenomic analysis of apple orchard soil reveals antibiotic resistance genes encoding predicted bifunctional proteins. Applied and environmental microbiology 76(13), 4396–4401 (2010).
Allen, H. K. et al. Resident microbiota of the gypsy moth midgut harbors antibiotic resistance determinants. DNA and cell biology 28(3), 109–117 (2009).
Acknowledgements
The authors would like to acknowledge the gracious access to research datasets made publicly available by Andreote et al. 2012, Alzubaidy et al. 2016, Global Ocean Sampling Expedition, Rusch et al. 2007, Williamson et al. 2008, DeAngelis et al. 2010, Fierer et al. 2012, Manoharan et al. 2015 and to the MG-RAST team for the free availability of the pipeline and server resources. Authors thanks to DST-SERB-EMEQ for partial financial assistance, IM thanks to UGC-NFST fellowship Govt. of India and the research facilities supported by Central University of Kerala. DB thanks TWAS-CNPq for granting postdoctoral fellowship at UFMG.
Author information
Authors and Affiliations
Contributions
Conceived and designed the experiments: R.K. and I.M. Analyzed the data: R.K., I.M., A.V., D.B., and A.R.W. Contributed reagents/materials/analysis tools: R.K. Wrote the paper: R.K., I.M., V.A., S.T., D.B., A.V., P.G., A.R.W. and A.G.N.
Corresponding author
Ethics declarations
Competing Interests
The authors declare no competing interests.
Additional information
Publisher's note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Imchen, M., Kumavath, R., Barh, D. et al. Comparative mangrove metagenome reveals global prevalence of heavy metals and antibiotic resistome across different ecosystems. Sci Rep 8, 11187 (2018). https://doi.org/10.1038/s41598-018-29521-4
Received:
Accepted:
Published:
Version of record:
DOI: https://doi.org/10.1038/s41598-018-29521-4
This article is cited by
-
Metabarcoding reveals unique rhizospheric microbiomes of Rhizophora in Indian Mangroves
Folia Microbiologica (2025)
-
Untapped rich microbiota of mangroves of Pakistan: diversity and community compositions
Folia Microbiologica (2024)
-
Microbial Metabolites Beneficial to Plant Hosts Across Ecosystems
Microbial Ecology (2023)
-
Bacterial diversity in the aquatic system in India based on metagenome analysis—a critical review
Environmental Science and Pollution Research (2023)
-
Metagenomic landscape of taxonomy, metabolic potential and resistome of Sardinella longiceps gut microbiome
Archives of Microbiology (2022)
