
Abstract
The present work was designed to investigate the characterization, as well as the antioxidation and renoprotection in streptozocin (STZ)-induced diabetic mice, of exopolysaccharides (EPS) and the enzymatic-EPS (EEPS) and acidic-EPS (AEPS) hydrolysates, which were separated from the fermentation liquor of Hypsizigus marmoreus. Animal results demonstrated that EPS, EEPS and AEPS had potential antioxidant and renoprotective effects, especially EEPS. Additionally, they were the most effective, reflecting increases in superoxide dismutase (SOD), glutathione peroxidase (GSH-Px), catalase (CAT), total antioxidant capacity (T-AOC), and albumin (ALB) of 168.33%, 124.8%, 268.17% 179.49%, and 68.71%, respectively, and decreases in the contents of malondialdehyde (MDA), lipid peroxide (LPO) and levels of serum urea nitrogen (BUN) and creatinine (CRE) by 70.58%, 58.43%, 23.97% and 29.60%, respectively, at a dose of 800 mg/kg compared to those of model mice. Three polysaccharides ameliorated the histopathological alterations which were observed in the kidney of diabetic mice. Furthermore, the characterization of polysaccharides had been expressed. These findings indicated that the EEPS from H. marmoreus possesses more effective renoprotection and antioxidation effects and provided insight into its potential clinical values on preventing diabetes.
Similar content being viewed by others
Introduction
Currently, chemical-synthetic medicaments, which are widely used clinically in curing many diseases, can cause serious cellular damage in the intracorporeal organs, including the heart, liver, kidney, and pancreas, and previous literature has indicated that these chemicals can be metabolically activated into highly reactive free radical compounds, which can induce oxidative stress, resulting the aggravation of disease progress1. Internationally, to keep away from these negative effects, many researchers have committed themselves to exploring natural and non-toxic substrates as conventional and effective clinical medicine2. The macrofungi of mushrooms, the most popular natural food, owing to its special mouthfeel and abundant nutrition, have gained increasing academic attention. They were widely used in identifying innovative drugs due to their vast bioactive compounds, such as proteins, polysaccharides, helvolic acid, and p-terphenyls. Additionally, these mushrooms have potential effects as antibacterial, anti-tumor3, antioxidant compounds, in protection against DNA damage4, and as neuroprotective compounds5. Numerous literature has demonstrated that polysaccharides, the most varied nutrient dense and abundant substances, have potential biological properties, such as the antioxidants of Tricholoma mongolicum Imai6, the antitumor compounds of Lentinus edodes7, and the anti-hyperlipidemic compounds of Termitomyces albuminosus8. In this regard, the polysaccharides from H. marmoreus, one of the most widely used traditional Chinese medicines and nutritional mushrooms, have attracted much attention because of its multiple pharmacological effects, including as antioxidant, anti-tumor, antiviral compounds and for their immunomodulating properties9. However, previous studies have focused their attentions on mycelia/intracellular and fruiting-body polysaccharides. However, there is scarce literature about exopolysaccharides/extracellular polysaccharides (EPS). Interestingly, Chen et al.10 indicated that EPS, a class of high-value biopolymers extracted from fermentation broth, have superior industrial application owing to their higher yields and related procedures that are less time-consuming and have less chance of contamination.
Diabetes mellitus (DM), the most common cause of death worldwide that is characterized by hyperglycemia, can lead to impaired metabolic functions of carbohydrates, lipids and proteins, inducing DM complications such as organ damage11. Streptozotocin (STZ), which is commonly employed in establishing diabetic animal models, can cause organ damage which is analogous to human symptoms12. Although the exact mechanism of STZ-induced toxicity is not well understood, several studies have shown the inevitable relationship between STZ-induced oxidative stress and kidney damage13. Experimentally, STZ can generate high amounts of reactive oxygen species (ROS), including superoxide anion, hydroxyl, alkoxyl and peroxyl radicals, and stimulate organic lipid peroxidation14, leading to serious oxidative stress. Notably, our previous studies have demonstrated organic dysfunction associated with STZ-induced toxicity that can be attenuated by the treatment with polysaccharides from mushrooms11.
The aim of the present work was designed to evaluate the renoprotective and antioxidant effects of EPS and its two hydrolysates, enzymatic-EPS (EEPS) and acidic-EPS (AEPS), from the fermentation broth of H. marmoreus in STZ-induced diabetic mice to better understand possible anti-diabetes mechanism and their health benefits in food and the pharmaceutical industry, indicating that the polysaccharides could be developed as valuable functional foods for clinical diabetes treatments. Furthermore, the exploration of fermentation liquor and the utilization of fermentation liquor-derived and value-added products, seem to be significant.
Results
Body weights and glucose (GLU) levels
As can be noted in Table 1, in the pretreatment condition, no significance was shown in both the body weights and GLU levels between the mice of the model control group (MC) groups and dosage groups (P > 0.05). Additionally, the significant increase in the GLU levels in MC mice compared with the normal control group (NC) mice indicated the successful hyperglycemia model. After two weeks, the body weight of the NC mice was significantly higher than those of the MC mice (diabetic mice treated with distilled water) (P < 0.05), and the GLU level of the NC mice was also lower than those of the MC mice (P < 0.05). Interestingly, for mice in the high-dose (800 mg/kg) EEPS group, the body weight was close to the normal group and increased by 41.80% compared with the MC mice. Meanwhile, the GLU level in the experiment group treated with EEPS at dose of 800 mg/kg decreased by 50.38% compared to those of the MC mice.
Histopathological analysis
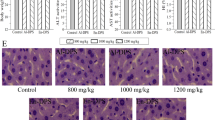
Obviously, the kidney damage in diabetic mice could also be confirmed by histopathological observations (Fig. 1), indicating that kidney damage was successfully established by STZ injection. In the MC group, diabetic mice exhibited serious glomerular degeneration and renal lesions reflected by inflammatory infiltration, extracellular matrix deposition, and glomerular basement membrane distortion, while the normal mice showed an integrated glomerulus and intact glomerular basement membranes. This was especially the case for the histopathological damage, which can be relieved in EEPS treatment mice at a dose of 800 mg/kg, and similar remission has been shown in the positive control (PC) group mice (Fig. 1).

Effects of EPS, EEPS, and AEPS on kidney tissue damage in NC, MC, PC, and treated diabetic mice with EPS, EEPS, and AEPS at doses of 800 mg/kg, 400 mg/kg and 200 mg/kg (HE staining, magnification ×600, *cellular degeneration;  glomerulus destruction).
glomerulus destruction).
Effects of EPS, EEPS, and AEPS on kidney
As seen in Fig. 2A, significant increase in the kidney index observed in the MC group in comparison with the NC group. Presently, after the treatment with EEPS, AEPS and EPS, the kidney index increase observed in the MC group was alleviated (Fig. 2A), especially the treatment of EEPS at the dose of 800 mg/kg.

Effects of EPS, EEPS and AEPS on (A) kidney index, (B–D) serum analysis, (E–H) hepatic enzymatic analysis and (I,J) lipid peroxidation. The values are reported as means ± SD (n = 10). Bars with the same letter are not significantly different (P < 0.05).
As displayed in Fig. 2B,C, the serum urea nitrogen (BUN) and creatinine (CRE) were 8.47 ± 0.33 mM and 85.84 ± 3.00 µM, respectively, in the diabetic mice (MC group), which were markedly increased in comparison with the NC mice (6.35 ± 0.30 mM and 52.95 ± 4.51 µM), while the albumin (ALB) declined in MC (10.93 ± 1.01 g/L) compared with NC mice (19.76 ± 1.20 g/L). Interestingly, three polysaccharides at different doses could ameliorate the biochemical enzyme activities of the STZ-induced mentioned above in comparison with the MC mice, especially the EEPS at a dose of 800 mg/kg, which decreased the BUN and CRE by 23.97% and 29.60%, respectively, compared to those of the MC group. Meanwhile, BUN and CRE decreased by 21.72% and 25.59%, respectively after treatment with AEPS, and BUN and CRE also decreased by 19.95% and 23.25%, respectively, by treatment with EPS at the same dose (800 mg/kg), while the ALB levels were increased by 68.71%, 64.23% and 55.72% in EEPS, AEPS and EPS treatment, respectively, in mice at a dose of 800 mg/kg. The enzyme activities contained superoxide dismutase (SOD), glutathione peroxidase (GSH-Px), catalase (CAT), total antioxidant capacity (T-AOC), as well as the contents of lipid peroxide (LPO) and malondialdehyde (MDA), which were observed in Fig. 2E–J. Compared with the mice in the NC group, the significant decrease in the SOD, GSH-Px, CAT and T-AOC activities and significant increase in the LPO and MDA contents in the MC group indicated that the kidney had been seriously damaged (P < 0.05). Obviously, the treatment of EEPS, AEPS and EPS expressed concentration-dependent patterns at the tested concentrations with increasing activities of SOD, GSH-Px, CAT and T-AOC, as well as decreasing contents of LPO and MDA (P < 0.05). Especially, the activities of SOD, GSH-Px, CAT and T-AOC were increased by 168.33%, 124.8%, 268.17% and 179.49%, respectively, and the contents of LPO and MDA were decreased by 70.58% and 58.43%, respectively, compared to those of the MC mice with EEPS at a dosage of 800 mg/kg. The PC group mice treated with glibenclamide also expressed remarkable increase in the GSH-Px, CAT, SOD and T-AOC activities and significant decline in the MDA and LPO contents compared with the MC mice.
Effects of EPS, EEPS, and AEPS on serum lipid levels
Figure 3 shows that the serum total cholesterol (TC) levels, triglyceride (TG) levels, high density lipoprotein cholesterol (HDL-C) levels, low-density lipoprotein cholesterol (LDL-C) levels, and very low-density lipoprotein cholesterol (VLDL-C) levels in diabetic mice were significantly higher (P < 0.05), while the HDL-C level was significantly lower than those in the MC groups (P < 0.05), indicating the disordered circulatory lipoproteins. After two weeks of gavage administration, the TC, TG, LDL-C, and VLDL-C levels were decreased significantly (P < 0.05), and the HDL-C levels were increased significantly in the dosage groups compared to those in the MC groups (P < 0.05, Fig. 3), indicating that the three polysaccharides had potential protective effects. In EEPS treated mice at 800 mg/kg, the TC, TG, LDL-C, and VLDL-C levels were decreased by 20%, 52.94%, 58.88% and 49.29%, respectively, and the HDL-C level was increased by 68.81% compared to those in the MC group. Similar conclusions could be drawn from the positive control mice treated with glibenclamide, suggesting the protective effects of positive medicine.

Effects of EPS, EEPS and AEPS on lipid metabolism. (A) TC levels, (B) TG levels, (C) HDL-C levels, (D) LDL-C levels, and (E) VLDL-C levels. The values are reported as means ± SD (n = 10). Bars with the same letter are not significantly different (P < 0.05).
Acute toxicity study
During the whole period of treatment with EEPS, AEPS and EPS, even at a dose of 4000 mg/kg, all the mice showed normal activities and did not show any clinical signs of toxicity. Furthermore, no death was observed either originally or finally, indicating that these three polysaccharides were all practically non-toxic substances15.
The primer analysis of characteristics
High-performance liquid chromatography (HPLC) chromatograms indicated that the Mw (weight-average molecular weight), Mn (number-average molecular weight) and Mz (z-average molecular weight) of EEPS were 2.14 × 103, 1.12 × 103 and 1.39 × 103 Da, while the Mw, Mn and Mz of EPS were 1.07 × 103, 1.15 × 103 and 1.14 × 103 Da, and the Mw, Mn and Mz of AEPS were 1.20 × 103, 1.95 × 103, 1.41 × 103 Da, respectively.
Scanning electron microscope (SEM) images of EPS, EEPS and AEPS are shown in Fig. 4A–C. Under 25000 × magnification, EPS showed an integrated surface with several regular shapes (Fig. 4A). Different from EPS, the microstructure of EEPS and AEPS presented an irregular stratified structure, and many holes with a non-uniform size were bestrewed on its surface (Fig. 4B,C).

The SEM analysis of (A) EPS, (B) EEPS and (C) AEPS, and GC spectra of (D) EPS, (E) EEPS and (F) AEPS.
The monosaccharide compositions of EPS, EEPS and AEPS were identified by comparing the retention time with standard monosaccharides (Fig. 4D–F) by gas chromatography (GC) analysis. Obviously, EEPS contained rhamnose, mannose, galactose and glucose with a molar ratio of 1.1:2.3:1.0:6.4 (Fig. 4E), while EPS and AEPS were both composed of mannose, galactose and glucose with different molar ratios of 1.1:1.0:2.3 (EPS, Fig. 4D) and 2.5:1.0:6.9 (AEPS, Fig. 4F), respectively.
Fourier transform infrared (FT-IR) spectroscopy revealed a typical major broad peak of hydroxyl stretching vibration around at 3332.37 cm−1 (EPS), 3422.01 cm−1 (EEPS), and 3407.76 cm−1 (AEPS), respectively. The three polysaccharides showed bands around 2900–3000 cm−1 for C-H stretching vibration. The bands in the region of 1631.69 cm−1 (EPS), 1631.46 cm−1 (EEPS) and 1631.40 cm−1 (AEPS) indicated the presence of C=O groups of amide stretching vibration (-CO-NH2). This demonstrated that the small amounts of protein in the samples may be sugar-binding proteins. The bands at 1450.59, 1449.98 and 1452.96 cm−1 corresponded to the characteristic C-O (-COOH) stretching vibration. The bands at 1375.17, 1375.26 and 1375.64 cm−1 corresponded to the C-H variable angle vibration in EPS, EEPS and AEPS (Fig. 5A–C). The band at 1239.69 cm−1 indicated the O-H (-COOH) variable angle vibration in EEPS. Additionally, the fingerprint region between 650 and 1350 cm−1 was usually associated with the stretching vibrations of C-C and C-O, as well as the bending mode of the C-H band. It had previously been suggested as a promising area for the analysis of structural conversions, especially the region of 980–1170 cm−1, which was sensitive to OH groups and could significantly affect the main band positions. As shown in Fig. 5, the bands in the 1010.96 cm−1 (AEPS) region were due to C-O and O-H stretching vibrations, suggesting that both pyranose rings existed in AEPS16,17,18.

The FT-IR spectra of (A) EPS, (B) EEPS and (C) AEPS.
The 1H nuclear magnetic resonance spectroscopy (NMR) spectrum of EPS showed four signals at 4.91, 4.90, 4.27 and 4.26 ppm corresponding to anomeric protons (Fig. 6A). In this spectrum, the 4.1 and 4.6 ppm signals corresponded to signals obtained for β glucan (Fig. 6A). Similarly, the 13C NMR spectrum contained four anomeric carbon signals from 92.69 to 104.61 ppm (Fig. 6D). The results suggested that EPS contained both α- and β-anomeric configurations. The 1H spectra of EEPS and AEPS showed a chemical shift in the anomeric region at 4–6 ppm (Fig. 6B,C). Similarly, the 1H spectrum of the polysaccharides exhibited a set of wide and intense signals (3.0–4.0 ppm) due to the CH2-O and CH-O groups of the sugars, while the chemical shifts from 3.2 to 4.1 ppm were assigned to the H-2 to H-6 protons. Furthermore, the regions between 1.4 and 2.5 were also observed to relate to the glucan-protein structure. Figure 6E,F showed the 13C NMR spectra of EEPS and AEPS from H. marmoreus. The presence of glucose could be observed by signals at 102.97 and 103.15 ppm19,20.

The 1H NMR spectra of (A) EPS, (B) EEPS and (C) AEPS, and 13C NMR spectra of (D) EPS, (E) EEPS and (F) AEPS.
Discussion
Pharmacologically, toxicity studies have always been considered as vital evidence during drug development. Different drugs have been synthesized for the treatment of DM; however, many synthetic drugs have a number of serious side effects1. With comparatively low side effects, polysaccharides from mushroom were used as potential medicines to open new avenues for the treatment of various diseases due to their biological activities6,7,8. Many investigators focused their academic attention on the study of antioxidation from either the fruiting body or mycelia of various medicinal fungi. However, there still been poor publication of reports focusing on the toxicity and biological activities of polysaccharides with the necessary modifications. It was necessary determine whether the biological activities will remain when EEPS and AEPS are orally administered. Hence, our present work was designed to investigate the renoprotective effects of EPS and their hydrolysates (EEPS and AEPS) from H. marmoreus in mice against oxidative stress induced by STZ injection.
In recent years, mushroom polysaccharides have been confirmed to possess higher antioxidant activities in protecting against lots of diseases induced by ROS13. DM, a group of metabolic diseases accompanied by organ damage, had been reported to be a complicated disease related to oxidative stress and have become currently the most common danger, greatly threatening human health21. Development of therapies to prevent the generation of free radicals may influence the progression of oxidative organ damage induced by STZ. Although the exact mechanisms of STZ-induced toxicity remain poorly understand, previous studies suggested that lipid peroxidation and free radical formation had devastating roles in the development and progress of diabetes11. The possible mechanism may be that ROS could interact with many biological macromolecules, such as lipids, proteins and DNA, causing structural changes, functional abnormalities, and diabetic complications4,5. Clinically, the complications were associated with organ damage, mainly focused on liver, kidney, pancreas and heart21. Therefore, antioxidants may play important roles in preventing DM and its complications by directly interfering with the generation of ROS. Currently, the protective effects on the kidney in STZ-treated mice, which are accompanied by the antioxidant activities of EPS and its two hydrolysates (EEPS and AEPS) from H. marmoreus, were investigated using serum and tissue parameters.
The kidney is an organ that plays a vital role in the glucose metabolism of diabetic animals. Three serum parameters, including the BUN, CRE and ALB levels were always used for clinical kidney examination. BUN, as the first endogenous substance generated by the decomposition of liver proteins, was excreted by the filtration of the glomerulus. CER, as the byproduct of creatine and phosphocreatine catabolism, was endogenously produced and released into the body fluids22. In addition, the ALB levels could also be an indication of nephropathy, reflecting increments of the glomerular filtration rate because the damaged kidney could expedite excessive plasma protein filtering into the urine11. Previous literature has indicated that STZ could directly induce hyperglycemia, owing to its selective toxicity on pancreatic β-cells, resulting in the disordered secretion of insulin23. Huang et al.24 indicated the pancreatic protection of polysaccharides may be attributed to its anti-diabetic activity.
As a metabolic disorder, DM was characterized not only by increased glucose levels but also by dysregulation of lipid profiles. In the cholesterol families, the HDL-C could carry the cholesterol ester from the peripheral tissues to the liver by the “reverse cholesterol transport” pathway during blood circulation, inhibiting the risk of atherosclerotic cardiovascular diseases. However, the other two lipoproteins of LDL-C and VLDL-C could accelerate the risk factors for coronary heart disease8. In addition, higher TC and TG levels could be a risk for atherosclerotic plaque lesions, attributing to their pathological accumulation in the blood vessel walls11. In this work, the administration of three polysaccharides (EPS, EEPS and AEPS) from H. marmoreus led to the improvement of TC, LDL-C and VLDL-C and the decline of HDL-C, providing clear evidence that the polysaccharides had potential protective effects on organs.
Furthermore, the activities of the antioxidant enzymes (SOD, GSH-Px, CAT and T-AOC), and lipid contents (LPO and MDA) were observed to investigate the protective effects of STZ on organ damage against oxidative stress. Sabir et al.25 reported that SOD and GSH-Px, the important free radical scavenging enzymes in the first line of defense against oxidative injury, were responsible for the detoxification of deleterious oxygen radicals and played important roles in protecting the cell from oxidative damage. Meanwhile, the decrease in the SOD activity could lead to an excess of superoxide anion and hydrogen peroxide (which in turn generated hydroxyl radicals) in biological systems, resulting in the initiation and propagation of lipid peroxidation. The decomposition of hydrogen peroxide could be catalyzed into less-reactive gaseous oxygen and water molecules by CAT26. In addition, the GSH-Px, which played a vital role mainly in the detoxication and metabolism as a cofactor or a substrate for some enzymes, could catalyze the reduction of H2O2 into H2O and O225. Thus, GSH-Px may be measured as a common marker of free radical damage in protecting tissues from oxidative stress.
In the free radical hypothesis, these reactive radicals could react with polyunsaturated fatty acids in cell membranes, leading to lipid peroxidation and dysfunction of membranes and subsequently resulting in tissue injury. Simultaneously, excess free radical could also be generated during abnormal lipid peroxidation11. Can et al.27 indicated that the in vivo organ damage caused by diabetes was probably due to free radicals produced by lipid peroxidation. Reed28 had also reported that the content of MDA, an end product of lipid peroxidation, may provide a convenient index of lipid peroxidation.
The biological functions, including antioxidant properties, were mainly associated with their monosaccharide compositions, molecular weights, bond types, and so on26. In the present work, all the results indicated that EEPS exhibited potentially superior protective effects on kidney damage compared to EPS and AEPS. Based on the monosaccharide analysis, only EEPS contained L-rhamnose, indicating that the L-rhamnose may play important role in maintaining protective effects against kidney damage. Furthermore, the NMR analysis of EEPS agreed with preliminary test results. According to previous studies29, chemical shifts correspond to C2 and C6. These observations were in accordance with other studies19. The analysis of 13C NMR spectra showed that the polysaccharides from H. marmoreus contained a high level of glucose, and the possible mechanism may be due to the type of homopolysaccharides (glucans). The results indicate that the superior effects on STZ-induced kidney damage were responsible for these special characteristics. Meanwhile, similar results of Grifola frondosa19 and Macrocybe gigantea30 could confirm our conclusions. The characteristic changes coincided with the shape changes in the polysaccharides. The SEM of EPS, EEPS and AEPS as well as their derivatives at magnifications of 25000 demonstrated these results. The results showed that the different extraction methods induced different physical changes in size and shape. The SEM analysis demonstrated that EPS was relatively regular and homogeneous in shape, while the surface of EEPS and AEPS was rough and had an irregular shape, with the possible mechanism potentially owing to the branches and network structures of the polysaccharides. Previous literature has indicated that the surface topography of a polysaccharide may be influenced by different methods of extraction, purification, and preparation31. In the present work, the difference in the SEM of the three polysaccharides may be that the polysaccharide properties had been well changed by enzymatic hydrolysis, showing superior physicochemical properties with good water solubility, high stability, safety and a lack of toxicity32. Furthermore, the main advantages of enzymatic hydrolysis were the high selectivity and substrate specificity, enabling products with well-defined and stereospecific structure33. In agreement with these results, similar conclusions were also reported by Zhao et al.8 for Termitomyces albuminosus, Jia et al.34 for Cordyceps militaris fruit bodies, and Yang et al.35 for Phoma herbarum. Previous and present conclusions demonstrated that the extraction methods played important role in maintaining the biological properties due to their unique effects on influencing the shape and structure of the substances.
Conclusions
In the present work, EPS and its two EEPS and AEPS hydrolysates were isolated and characterized from H. marmoreus, and the protective effects on the kidney in diabetic mice induced by STZ were investigated. The results provided evidence that EPS, EEPS, and AEPS could effectively protect organs against STZ toxicity and could be used as potentially natural and functional ingredients in the prevention and alleviation of DM and its complications.
Materials and Methods
Chemicals and strain
The diagnostic kits for assaying SOD activity, GSH-Px activity, CAT activity, T-AOC activity, and LPO and MDA content were purchased from Nanjing Jiancheng Bioengineering Institute (Nanjing, China). Snailase and STZ were purchased from Sigma Chemicals Company (St. Louis, USA). Blood glucose test strips were purchased from Sinocare Biosensing Corporation Limited (Changsha, China). All other chemicals were of analytical grade and purchased from local chemical suppliers.
Culture media and conditions
The H. marmoreus strain used in this experiment was provided by the Shandong Agricultural Academy of Sciences (Shandong, China) and maintained on a potato dextrose agar (PDA) slant at 4 °C. The liquid culture (1 L) with natural pH was composed of 20 g of glucose, 3 g of peptone, 4 g of yeast extract, 1 g of KH2PO4 and 1 g of MgSO4. A 0.5-cm2 portion of the agar plate culture was sliced from the slant for inoculating the seed culture medium. Seeding cultivation in liquid media was cultured in a 1 L filter flask containing 600 mL of potato dextrose broth at 25 °C for 24 h without shaking and then shaken on a rotary shaker (160 rpm, Anting, Shanghai, China) for 7 days. The submerged fermentation was carried out in a 100-L fermentation tank (Xianmin, Luoyang, China) for 14 days (temperature 25 °C and pH 7) with the previous liquid culture media.
Preparation of EPS
The fermentation broth of H. marmoreus was centrifuged at 3000 rpm for 15 min, and supernatant liquid was collected. After concentrating fivefold under vacuum, the supernatant was mixed with 3 volumes of 95% ethanol (v/v), standing at 4 °C for 24 h. After centrifugation at 3000 rpm for 15 min, the precipitate was collected and purified by the Sevag method36. Then, the purified polysaccharides were considered as EPS by lyophilization, and vacuum freeze drying (Labconco, USA) with the contents determined by phenol-sulfuric acid, using glucose as the standard37 with the EPS yields of 1.44 ± 0.57 g/L.
Enzymatic and acidic degradation
The EEPS and AEPS were separately prepared according to the methods reported by Yang, et al.35 with slight modifications.
The enzymatic hydrolysis of EPS was carried out in snailase solution (1%, dissolved in sodium acetate buffer) depending the following conditions with a ratio of 1:4 (materials to snailase solution, w/v), extraction temperature of 40 °C, pH of 6, and extraction time of 5 h35. The final EEPS was obtained by concentration, deproteinzation36, and lyophilization.
The EPS in the test tube (18 mm × 180 mm) was hermetically hydrolyzed with H2SO4 (1 M, 1:20, w/v) in boiled water for 8 h. After centrifugation at 3000 rpm for 15 min, the supernatant was neutralized with NaOH solution (2 M). The AEPS was consecutively extracted by concentration, deproteinzation36 and lyophilization.
Animal experiments
The experiments were performed and approved by the institutional animal care and use committee of Shandong Agricultural University and in accordance with the Animals (Scientific Procedures) Act of 1986 (amended 2013). The Kunming strain of mice, weighing 20 ± 0.2 g, were purchased from the Taibang Company (Taian, China) and housed in stainless steel cages under controlled conditions (temperature 23 ± 2 °C, lights on 12 h every day). After three days to allow the mice to get acclimated, diabetes was induced by an intraperitoneal injection with STZ (80 mg/kg, freshly prepared in citrate buffer solution, 0.1 M, pH 4.5) for three successive days (once every day). The mice were considered diabetic when the tail-vein GLU levels were higher than 13.3 mM14. Successful diabetic mice were randomly divided into eleven groups of ten mice each, including one MC treated with distilled water; one PC group that received glibenclamide (20 mg/kg); and nine dosage groups treated with EPS, EEPS and AEPS at three different doses (200, 400 and 800 mg/kg), while another ten mice in the NC groups (normal mice without STZ-intervention) were treated with distilled water. The glibenclamide, polysaccharide and distilled water gavages were processed with a syringe daily, and the experiments lasted for 14 days. The body weights of the mice were measured every day. At the end of the experiment, the GLU levels from the tail vein were monitored by blood glucose test strips after fasting overnight, and the mice were sacrificed under ether anesthesia.
The serum in all the mice was collected by centrifugation (14000 rpm, 4 °C) for serum analysis. The serum levels of BUN, CRE, ALB, TC, TG, HDL-C, VLDL-C and LDL-C were measured using an automatic biochemical analyzer (ACE, USA).
The right kidney was randomly isolated from one mouse in each group and freshly immersed in PBS buffer (10% formalin, pH 7.4) for over 24 h and embedded in paraffin. Thin sections (4–5 µm thickness) were prepared using a microtome and stained with hematoxylin-eosin. Each section was photographed under a microscope to show the histopathological changes (×600 magnifications).
The other kidneys were rapidly weighed and homogenized (1:9, w/v) in 0.2 M phosphate buffer (4 °C, pH 7.4). The organic supernatants were collected by centrifugation (14000 rpm, 10 min), and the SOD, GSH-Px, CAT, and T-AOC activities, as well as the MDA and LPO contents, were determined by commercial reagent kits according to their instructions. The tissue index was calculated using the following equation: (tissue weight/body weigh) (g/100 g body weight).
Acute toxicity assay
An acute toxicity study was performed by the method of Chao et al.38 Twenty Kunming strain mice were randomly divided into four groups (five in each group). In the control group, mice were given free access to diet and water, while in the experimental groups, mice were orally given three polysaccharides at the ultimate dosage of 4000 mg/kg. The mice were observed continuously for the first 24 h for any gross behavioral changes and toxic symptoms, as well as for mortality in the first 48 h.
Characterization of EPS, AEPS and EEPS
The molecular weights and homogeneities were determined by HPLC on an HPLC system (Shimadzu LC-2010AT, Japan) equipped with an Atlantis C18 column (250 mm × 4.6 mm × 5 µm) and a refractive index detector. Deionized water was used as the mobile phase with a flow rate of 1 mL/min, and the column temperature was maintained at 30 °C. A series of standard dextrans were used to make the calibration curve. The molecular weights were analyzed by Agilent GPC software.
The morphological features of EPS, EEPS and AEPS were analyzed by SEM analysis (S-4800, FE-SEM, Hitachi High-Technologies, Japan). The dried powder of the polysaccharides was affixed on a glass slide and coated with gold powder to make the samples conductive. The images with a magnification of 25000 × were taken with an accelerating voltage of 10 kV.
Monosaccharide composition was determined by GC (GC-2010, Shimadzu, Japan) equipped with a capillary column of Rtx-1 (30 mm × 0.25 mm × 0.25 µm) using a previously published method39. Composition identification was processed by comparison with standard monosaccharides (mannose, rhamnose, glucose, galactose, arabinose, ribose, and xylose). The relative molar ratios were calculated by the area normalization method according to the chromatogram.
The dried polysaccharides samples (1 mg) were mixed with KBr powder (100–200 mg), and then the mixture was ground in a mortar under an infrared lamp to prevent the air-slake of KBr. After tableting, spectra were recorded on a 6700 Nicolet Fourier transform-infrared spectrophotometer (Thermo Co., Madison, WI, USA) within the range from 4000 to 400 cm−1.40
13C and 1H NMR spectroscopy experiments were conducted using a 700-MHz Varian Mercury 2010 Magneto Oxford spectrometer at 60 °C, and the samples were dissolved in dimethyl sulfoxide (DMSO).
Statistical analysis
All data were processed and analyzed using SAS and expressed the means ± standard deviation (SD). Two-way analysis of variance (ANOVA) followed by post-hoc Tukey’s tests were performed to statistically test the differences, and P < 0.05 was considered significant.
References
Gao, J. et al. Protective effect of polysaccharides from Opuntia dillenii Haw. fruits on streptozotocin-induced diabetic rats. Carbohyd. Polym. 124, 25–34 (2015).
Liu, Y. et al. Structural characterization and antidiabetic activity of a glucopyranose-rich heteropolysaccharide from Catathelasma ventricosum. Carbohyd. Polym. 149, 399–407 (2016).
Xiao, J. H. et al. Synergistic antitumor efficacy of antibacterial helvolic acid from Cordyceps taii and cyclophosphamide in a tumor mouse model. Exp. Biol. Med. 242, 214–222 (2016).
Jin, Q. L. et al. Antioxidant and DNA damage protecting potentials of polysaccharide extracted from Phellinus baumii using a delignification method. Carbohyd. Polym. 152, 575–582 (2016).
Chon, S. H., Yang, E. J., Lee, T. & Song, K. S. β-Secretase (BACE1) inhibitory and neuroprotective effects of p-terphenyls from Polyozellus multiplex. Food Funct. 7(9), 3834–3842 (2016).
Zhao, Y. M. et al. Optimization of cellulase-assisted extraction process and antioxidant activities of polysaccharides from Tricholoma mongolicum Imai. J. Sci. Food. Agr. 96(13), 4484–4491 (2016).
Yu, Z. et al. Structure, chain conformation and antitumor activity of a novel polysaccharide from Lentinus edodes. Fitoterapia. 81(8), 1163–1170 (2010).
Zhao, H. J. et al. The antihyperlipidemic activities of enzymatic and acidic intracellular polysaccharides by Termitomyces albuminosus. Carbohyd. Polym. 151, 1227–1234 (2016).
Chang, J. S. et al. Cancer preventive potential of methanol extracts of Hypsizigus marmoreus. J. Med. Food. 12, 493–500 (2009).
Chen, W., Zhao, Z., Chen, S. F. & Li, Y. Q. Optimization for the production of exopolysaccharide from Fomes fomentarius in submerged culture and its antitumor effect in vitro. Bioresour. Technol. 99, 3187–3194 (2008).
Lin, L. et al. Antioxidative and renoprotective effects of residue polysaccharides from Flammulina velutipes. Carbohyd. Polym. 146, 388–395 (2016).
Tesch, G. H. & Allen, T. J. Rodent models of streptozotocin-induced diabetic nephropathy. Nephrology. 12(3), 261–266 (2007).
Mollazadeh, H., Sadeghnia, H. R., Hoseini, A., Farzadnia, M. & Boroushaki, M. T. Effects of pomegranate seed oil on oxidative stress markers, serum biochemical parameters and pathological findings in kidney and heart of streptozotocin-induced diabetic rats. Renal Failure. 38(8), 1256–1266 (2016).
Szkudelski, T. The mechanism of alloxan and streptozotocin action in B cells of the rat pancreas. Physiol. Res. 50(6), 537–546 (2001).
Zhang, Y., Wu, X., Ren, Y., Fu, J. & Zhang, Y. Safety evaluation of a triterpenoid-rich extract from bamboo shavings. Food. Chem. Toxicol. 42(11), 1867–1875 (2004).
Yang, L. Q. & Zhang, L. M. Chemical structural and chain conformational characterization of some bioactive polysaccharides isolated from natural sources. Carbohyd. Polym. 3, 349–361 (2009).
Liu, H. H. et al. Polysaccharides from Lycium barbarum leaves: Isolation, characterization and splenocyte proliferation activity. Int. J. Med. Mushrooms. 51(4), 417–422 (2012).
Li, J. H. et al. Purification, characterization and bioactivities of polysaccharides from Pleurotus ferulate. Food Funct. 8(5), 1905–1914 (2017).
Meng, M., Cheng, D., Han, L. R., Chen, Y. Y. & Wang, C. L. Isolation, purification, structural analysis and immunostimulatory activity of water-soluble polysaccharides from Grifola Frondosa fruiting body. Carbohyd. Polym. 157, 1134–1143 (2017).
Kozarski, M. et al. Antioxidative and immunomodulating activities of polysaccharide extracts of the medicinal mushrooms Agaricus bisporus, Agaricus brasiliensis, Ganoderma lucidum and Phelllinus liteus. Food Chem. 129, 1667–1675 (2011).
Mohamed, A. K. et al. The role of oxidative stress and NF-κB activation in late diabetic complications. Biofactors. 10(2-3), 157–167 (1999).
Devi, B. A., Kamalakkannan, N. & Prince, P. S. Supplementation of fenugreek leaves to diabetic rats. Effect on carbohydrate metabolic enzymes in diabetic liver and kidney. Phytother. Res. 17(10), 1231–1233 (2003).
Moncada, S. R., Palmer, R. M. J. & Higgs, E. Nitric Oxide: Physiology, pathophysiology, and pharmacology. Pharmacol. Rev. 43(2), 109–142 (1991).
Huang, G. J. et al. Protective effect of antrosterol from Antrodia camphorate submerged whole broth against carbon tetrachloride-induced acute liver injury in mice. Food Chem. 132(2), 709–716 (2012).
Sabir, S. M. et al. Antioxidant and hepatoprotective activity of ethanolic extract of leaves of Solidago microglossa containing polyphenolic compounds. Food Chem. 131(3), 741–747 (2012).
Liu, J. et al. In vitro and in vivo antioxidant activity of exopolysaccharides from endophytic bacterium Paenibacillus polymyxa EJS-3. Carbohyd. Polym. 82(4), 1278–1283 (2010).
Can, A. et al. Effect of Aloe vera leaf gel and pulp extracts on the liver in type-II diabetic rat models. Biol. Pharm. Bull. 27(5), 694–698 (2004).
Reed, T. T. Lipid peroxidation and neurodegenerative disease. Free. Radic. Biol. Med. 51(7), 1302–1319 (2011).
Chauveau, C., Talaga, P., Wieruszeski, J. M., Strecker, G. & Chavant, L. A water-soluble β-D-glucan from Boletus erythropus. Phytochemistry. 43(2), 413–415 (1996).
Khong, T. Y. Acute atherosis in pregnancies complicated by hypertension, small-for-gestational-age infants, and diabetes mellitus. Arch. Pathol. Lab. Med. 115(7), 722–725 (1991).
Nep, E. I. & Conway, B. R. Physicochemical characterization of grewia polysaccharide gum: Effect of drying method. Carbohyd. Polym. 84(1), 446–453 (2011).
Wu, T. et al. Antioxidant and antimicrobial activity of acidolysis and enzymolysis products of Fenugreek polysaccharides. J. Food. Sci. 28(11), 509–543 (2007).
Gübitz, G. M. & Paulo, A. C. New substrates for reliable enzymes: enzymatic modification of polymers. Curr. Opin. Biotech. 14(6), 577–582 (2003).
Jia, J. Q. et al. Enzymatic modification and antioxidant activity of polysaccharides from Cordyceps militaris fruit bodies. Food Sci. 34(01), 114–120 (2013).
Yang, X. B. et al. Purification, characterization and enzymatic degradation of YCP, a polysaccharide from marine filamentous fungus Phoma herbarum YS4108. Biochimie. 87(8), 747–754 (2005).
Staub, A. M. Removal of protein-Sevag method. Methods Carbohydr. Chem. 5, 5–6 (1965).
Chaplin, M. F., & Kennedy, J. F. Carbohydrate Analysis: A Practical Approach. IRL Press Limited. New York (1994).
Chao, J. et al. Analgesic and anti-inflammatory activities of ethanol root extract of Mahonia oiwakensis in mice. J. Ethnopharmacol. 125(2), 297–303 (2009).
Sheng, J. C. et al. Preparation, identification and their antitumor activities in vitro of polysaccharides from Chlorella pyrenoidosa. Food Chem. 105, 533–539 (2007).
Zhang, W. J. Biochemical research technology of glycoconjugate, Zhejiang University Press. HangZhou. 195–200 (1999).
Acknowledgements
This work was supported by grants from Mushroom Technology System of Shandong Province (SDAIT-07–05), Mushroom Technology System of Shandong Province (SDAIT-07-04), and Shandong Agricultural Fine Breed Project (2014lz058).
Author information
Authors and Affiliations
Contributions
All experimental work and statistical analyses were designed by Min Liu and Le Jia. Yongfa Zhu, Hui Liu, Yufei Lan, Chengye Tian and Wenshuai Wang performed the research. Min Liu and Jianjun Zhang analyzed the data. Min Liu, Jianjun Zhang interpreted the results. Min Liu, Yufei Lan, and Weiru Liu repared figures and drafted the manuscript. Min Liu, Weiru Liu, Jianjun Zhang and Le Jia edited the manuscript. All authors were involved in checked the paper and contributed to the preparation of the final manuscript. All authors read and approved the final manuscript.
Corresponding authors
Ethics declarations
Competing Interests
The authors declare that they have no competing interests.
Additional information
Publisher's note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Liu, M., Lan, Y., Tian, C. et al. The characterization, renoprotection and antioxidation of enzymatic and acidic exopolysaccharides from Hypsizigus marmoreus. Sci Rep 8, 2048 (2018). https://doi.org/10.1038/s41598-018-20440-y
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-018-20440-y
This article is cited by
-
Genetic and functional analysis of the Zn(II)2Cys6 transcription factor HADA-1 in Hypsizygus marmoreus
Applied Microbiology and Biotechnology (2021)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
