
Abstract
The objective was to investigate the effects of the maternal consumption of the green tea extract during pregnancy and lactation on mothers and offspring metabolism. The female Wistar rats, on the first day of pregnancy until the end of lactation, was divided into groups: MC– received water and ME– received green tea extract (400 mg/kg body weight/day), both ingested control diet. After lactation, at day 28th post-partum, the mothers and pups from each mother were euthanized and composed the groups: FC– pup from mother received water and FE– pup from mother received green tea extract. The ME group increased IL-10/TNF-α ratio and IL-1β content in the mesenteric and IL-1β content in retroperitoneal adipose tissues, and decreased catalase activity. The FE group decreased the retroperitoneal adipose tissue relative weight and SOD activity, but increased adiponectin, LPS, IL-10 and IL-6 content and IL-10/TNF-α ratio in retroperitoneal, IL-10 and TNF-α content in gonadal, and IL-6 content in mesenteric adipose tissues. In summary, the maternal consumption of green tea extract associated with control diet ingestion during pregnancy and lactation altered the inflammatory status of mothers and 28d-old offspring. These data elucidate the effects of green tea during pregnancy and lactation on maternal and offspring metabolism.
Similar content being viewed by others

Introduction
Green tea, made from the dried leaves of Camellia sinensis, is one of the most popular beverages around the world. The green tea is rich in polyphenols, among them the catechins: epigallocatechin, epicatechin, epicatechin gallate and most abundant, epigallocatechin-3-gallate (EGCG)1,2,3. Epidemiological studies suggest that polyphenolic compounds in the tea reduce the risk of a variety of diseases and present antioxidant properties4,5,6.
The metabolic programming can be defined as a response of the organism to a challenge in developmental period that alters the phenotype with potential persistent effect on health of offspring throughout their lives7,8.
However, few studies assess the effects of tea consumption during pregnancy and lactation. One study reported, through in vitro rat embryos exposure to the EGCG during early organogenesis, that most of the tea catechins are very safe during early embryonic development9. Other studies demonstrated that EGCG was absorbed thoroughly, passed through the placenta, and reached the fetus, where it was distributed to major fetal organs in significant quantities10,11.
One study, in rats, investigated potential adverse effects of standardized heat-sterilized green tea catechins (GTC-H) on embryo-fetal development. For this, GTC-H was administered once daily from 6th through 17th day pregnancy and the dosage administered were 0 (control), 200, 600, and 2000 mg/kg/day. The doses used did not affect intrauterine growth and survival fetal malformations or developmental and only the 2000 mg/kg/day dose promoted hair loss in different areas12.
The objective of this study was to investigate the effects of the maternal consumption of the green tea extract during pregnancy and lactation on mothers and offspring metabolism.
Results
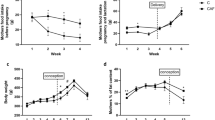
Body weight and delta weight
The mean body weights (BWs) and delta weight of the mothers during pregnancy and lactation, and pups with 28 days old were similar among groups during all treatment (Fig. 1).

Body weight and delta weight in the different groups during all treatment. Mother – C is MC group – mother received water; Mother – E is ME group – mother received green tea extract; Pups 28d – C is FC group – pup from mother received water; Pups 28d – E is FE group – pup from mother received green tea extract. Data are mean ± SEM (n = 8–13).
Tissues relative weight
There was no difference in tissues relative weight in the mothers. RET relative weight in the FE group was significantly lower than in the FC (p = 0.04) group, but there were no differences in others relative weight among groups (Table 1).
Serum concentrations
Serum concentrations of total cholesterol, triacylglycerol, glucose, insulin, adiponectin/SAT and HOMA-IR of the mothers and pups at 28d-old were similar among the groups. However, the serum concentration of adiponectin and LPS were significantly higher in the FE (p = 0.01 and p = 0.04, respectively) group compared with the FC group and were not different in mothers (Table 2).
Tissues cytokine content
Figure 2A shows that the IL-10 content in RET and GON was greater in the FE group than in the FC (p < 0.01 and p = 0.01, respectively) group. In other tissues, the IL-10 content was not significantly different among the MC versus ME groups.

Tissues cytokine content. (A) IL-10 content in RET, GON, MES and Liver; (B) TNF-α content in RET, GON, MES and Liver; (C) IL-10/TNF- α content in RET, GON, MES and Liver; (D) IL-6 content in RET, GON, MES and Liver and (E) IL-1β content in RET, GON, MES and Liver. Mother – C is MC group – mother received water; Mother – E is ME group – mother received green tea extract; Pups 28d – C is FC group – pup from mother received water; Pups 28d – E is FE group – pup from mother received green tea extract. Data are means ± SEM (n = 8–13). *p < 0.05, versus control group of the respective treatment.
TNF-α content in the GON was higher in the FE group than in the FC (p < 0.01) group. In other tissues, TNF-α content did not differ among the groups (Fig. 2B).
IL-10/TNF-α ratio in the MES of the ME group was higher than in the MC (p = 0.02) group. IL-10/TNF-α ratio was higher in RET of the FE group than FC (p = 0.02) group, but not different in any other tissues or groups (Fig. 2C).
IL-6 content in RET and MES was higher in the FE group than the FC (p = 0.01 and p = 0.03, respectively) group. In the others tissues and groups did not different (Fig. 2D).
In Fig. 2E, we observed that IL-1β content in RET and MES was higher in the ME group than in the MC (p = 0.03 and p = 0.02, respectively) group.
Antioxidant enzymes: superoxide dismutase, glutathione peroxidase and catalase
Table 3 showed the ME has lower catalase than the MC (p = 0.04) group. FE decrease the SOD when compared to the FC (p = 0.045) group. The GPx not differ in the groups.
Discussion
In the present study, maternal consumption of green tea extract during pregnancy and lactation did not promote alteration in body weight and delta weight in mothers and pups. However, mothers that consumed green tea extract demonstrated an increase in IL-10/TNF-α ratio and IL-1β in the mesenteric adipose tissue and a decrease in catalase. The 28 days old pups born from the dams which consumed green tea extract had lower relative mass of the retroperitoneal adipose tissue and lower superoxide dismutase, as well as higher adiponectin and LPS concentrations and inflammatory cytokines in adipose tissue. These results indicate that the consumption of the green tea extract by mothers altered the inflammatory status of mothers and offspring.
The animals did not present difference in body weight and delta weight associated with green tea consumption. Previous studies from our group demonstrated similar results when comparing normolipidic diet ingestion with or without green tea extract in other periods of life13,14. However, other studies observed reduction in body weight or delta weight when compared the consumption of green tea extract with control diet and water with control diet15,16. It is important to remember that in this study the offspring did not drink green tea extract.
The maternal intake of green tea extract promoted decreased in retroperitoneal adipose tissue relative weight in 28d-old pups. Cunha et al.15 demonstrated reduction in retroperitoneal adipose tissue relative weight in group that received control diet and green tea extract (400 mg green tea commercial extract/kg body weight/day) during 8 weeks, in mice. Other study with mice that received green tea extract (50 mg/kg body weight/day) and control diet per 8 weeks presented decrease in epididymal adipose tissue relative weight and SAT17. At this point it is interesting to note that the catechins concentration varies in different green tea extract depending on the types and the extraction time18 and the extract used in the current study had lower EGCG concentration, when compared to the others.
A study from our group evaluated the ingestion of 10% of prebiotic during pregnancy and lactation on 21d-old offspring and identified reduction in RET when compared to the pups from mothers that not ingested prebiotic, both with control diet19. Altogether, these data demonstrated the influence of maternal diet during pregnancy and lactation on pups’ metabolism.
The consumption of 400 mg/kg body mass/day of green tea extract during 8 weeks with control diet promoted increase in adiponectin serum of adult mice15. In another study using mice treated with green tea extract (50 mg/kg body mass/day) during 16 weeks, the authors observed an increase in adiponectin serum levels in animals fed with control diet and green tea extract20. In the present study, we demonstrated an increase in adiponectin serum levels in 28d-old pups demonstrating the effect of green tea consumption by mothers on pups’ metabolism.
We showed the decrease in catalase in the ME group and SOD in the FE group. Green teas possess high content of catechin, especially EGC and EGCG, and they display the highest antioxidant activity scavenging reactive oxygen species (ROS) by generating more stable phenolic radicals21,22. The antioxidant action of catechins in green tea is likely involved in their anti-obesity mechanism owing to the fact that ROS stimulate NFκB, which in turn promotes the expression of proinflammatory cytokines, such as TNF-α and IL-1β23,24.
Studies with green tea extract showed increased IL-10 and TNF-α levels in retroperitoneal, gonadal and mesenteric adipose tissue in adult mice when associated with normolipidic diet15,17. However, another study showed anti-inflammatory effects of the EGCG and EGC by the inhibition of IL-1β-induced soluble mediators (IL-6 and IL-8), in activated synovial fibroblasts of humans with rheumatoid arthritis25. We observed an increase in IL-10 levels in gonadal and retroperitoneal adipose tissue and TNF-α in gonadal adipose tissue in FE group, and additionally, an increase in IL-10/TNF- α ratio in retroperitoneal adipose tissue in FE group and in mesenteric adipose tissue in ME group. IL-6 in RET, and IL-1β in MES was increased in FE and ME group, respectively. These data demonstrate a proinflammatory effect of maternal consumption of the green tea during pregnancy and lactation in mothers and pup 28d-old.
The increased levels of TNF-α are associated with some inflammatory disorders, and IL-10 is secreted by activated monocytes/macrophages and lymphocytes and is known to possess multifaceted anti-inflammatory properties26. Therefore, suppressing chronic inflammation may be a good strategy to prevent and/or treat obesity that is developed by chronic systemic inflammation. Interestingly, previous studies suggest that the positive impacts of green tea polyphenols could be via their ability to suppress chronic inflammation27,28,29.
The white adipose tissue depots are subdivided into subcutaneous (for example: intramuscular and inguinal) and visceral (for example: retroperitoneal, mesenteric and gonadal) adipose tissue30. The visceral adipose tissue is considered a pro-inflammatory endocrine tissue and is associated with hypertension, dyslipidemia, inflammation, glucose intolerance, insulin resistance and obesity31,32. Previously, it was described that the obese adipose tissue is characterized by production and secretion of inflammatory molecules like TNF-α and IL-6, which may have local and systemic effects33,34,35. Also, the literature shows that LPS induced the activation of the Toll-like receptor 4 (TLR) leading to the production of proinflammatory cytokines, as IL-1β, IL-6 and TNF-α36,37. This study demonstrates that the ingestion of green tea extract during pregnancy and lactation promoted alteration in inflammatory response on maternal and offspring visceral adipose tissue, specific retroperitoneal and mesenteric depots.
Additionally, in the offspring from mothers that received green tea extract, the alterations on the visceral adipose tissue inflammatory milieu were accompanied by increased concentration of LPS on serum, contributing to the proinflammatory state observed in visceral adipose tissue of the pups with 28d-old.
Although it was not evidenced statistically, by analyzing the values of cytokine in the different white adipose tissue depots, we observed that the retroperitoneal adipose tissue of the mothers and offspring of the green tea group presented higher values when compared to the other groups. It seems that different visceral white adipose tissue depots could respond distinctly depending on the nutritional interference.
In summary, the maternal consumption of green tea extract associated with control diet ingestion during pregnancy and lactation promoted proinflammatory status in mothers and 28d-old offspring. These data demonstrated alterations in metabolism development of the offspring by modification in inflammatory milieu in different deposits of visceral adipose tissue. Further studies are needed in order to examine the dose-dependent effects of the green tea extract on metabolic programming.
Methods
Animals and treatments
The Experimental Research Ethic Committee of the Universidade Federal de São Paulo approved all procedures for the care of the animals used in this study (CEUA n°: 718008/2013). The rats were kept under controlled conditions of light (12-h light/12-h dark cycle with lights on at 07:00) and temperature (24 ± 1 °C). Three-month-old female Wistar rats were left overnight to mate, and copulation was verified the following morning by the presence of sperm in vaginal smears.
On the first day of pregnancy, the dams were isolated in individual cages, receiving control diet and were randomly divided into two groups: water (MC group) and green tea extract (ME group). The treatment was maintained throughout pregnancy and lactation.
On the day of delivery, considered day 0 of lactation, litter sizes were adjusted to nine pups each. The animals were weighed weekly.
After lactation, at day 28th post-partum, the mothers and one pup from each mother were euthanized and composed the groups: FC – pup from mother received water and FE – pup from mother received green tea extract.
The green tea extract was offered in amber bottle daily at the concentration of 400 mg/kg body weight/day diluted in the water according to the volume ingested in the previous day. Composition of catechins of the green tea extract used in this study was: 16 μg/mg catechin, 29 μg/mg epicatechin, 24 μg/mg epicatechin gallate, 40 μg/mg epigalocatequinagalate and 58 μg/mg epigallocatechin.
The control diet was modified according to the recommendations of the American Institute of Nutrition (AIN- 93 G)38. The composition of 1 kg of diet is cornstarch (629.5 g), casein (200 g), soybean oil (70 g), fiber (50 g), vitamin mix (10 g), mineral mix (35 g), L-cysteine (3 g), choline bitartrate (2.5 g) and tert-butylhydroquinone (0.014 g). The calories percentage of the macronutrients are 63.8% carbohydrates, 20.3% protein and 16% lipids, and the energy values are 3.9 kcal/g.
Experimental procedures
The mothers and pups were decapitated on postnatal 28 days, where 8–13 animals per group were used in the post coming analyzes. The mothers were fasted but not the 28d-old pups to avoid the weaning stress. Trunk blood was collected and immediately centrifuged. The serum was separated and stored at −80 °C for later determinations. The retroperitoneal (RET), mesenteric (MES) and gonadal (GON) white adipose tissue and liver were isolated, weighed, immediately frozen in liquid nitrogen and stored at −80 °C.
Biochemical and hormonal serum analyses
Serum glucose, total cholesterol and triacylglycerol concentrations were measured using a commercial enzymatic colorimetric kit (Labtest®, Brazil). Insulin (Millipore®, USA), adiponectin (AdipoGen Life Sciences®), and lipopolysaccharideo (LPS) (Lonza®) concentrations were quantified using specific commercial kits. HOMA-IR and adiponectin/SAT ratio were calculated. SAT is the index of adiposity and it was calculated by the sum of MES, RET, and GON relative weight.
IL-10, TNF-α, IL-6 and IL-1β protein concentration determined by ELISA
Following decapitation, portions of the adipose tissue (0.3 g) and liver (0.1 g) were homogenized in 800 µL of chilled extraction buffer (100 mM Trizma Base pH 7.5; 10 mM EDTA; 100 mM NaF; 10 mM Na4P2O7; 10 mM Na3VO4; 2 mM PMSF; 0.1 mg/ml aprotinin). After homogenization, 80 μl of 10% Triton X-100 was added to each sample. These samples were held on ice for 30 minutes and then centrifuged (20817 g, 40 minutes, 4 °C). The supernatant was collected, and protein concentrations were determined using the Bradford assay (Bio-Rad®, Hercules, California) with bovine serum albumin as a reference. Quantitative assessment of IL-10, TNF-α, IL-6 and IL-1β proteins was carried out using ELISA (DuoSet ELISA, R&D Systems, Minneapolis, MN, USA) following the recommendations of the manufacturer.
Antioxidant enzymes activity
The liver was weighted and homogenized in specific buffer. Superoxide dismutase (SOD) and glutathione peroxidase (GPx) enzyme activities in the serum were determined using RANSOD and RANSEL Kits (Randox Laboratories, Crumlin, UK), respectively. For the catalase activity the hydrogen peroxide consumption method was used39. The protein concentration was measured by the Bradford method.
Statistical analysis
All results were presented as the means ± standard error of the mean (SEM). The statistical significance of the differences between the means of the samples of the groups (MC versus ME; and FC versus FE) was assessed using independent test-t. Differences were considered to be significant when p < 0.05.
References
Chen, Z., Zhu, Q. Y., Tsang, D. & Huang, Y. Degradation of green tea catechins in tea drinks. Journal of agricultural and food chemistry 49, 477–482 (2001).
Chen, N. et al. Green tea, black tea, and epigallocatechin modify body composition, improve glucose tolerance, and differentially alter metabolic gene expression in rats fed a high-fat diet. Nutrition research 29, 784–793, https://doi.org/10.1016/j.nutres.2009.10.003 (2009).
Graham, H. N. Green tea composition, consumption, and polyphenol chemistry. Preventive medicine 21, 334–350 (1992).
Dulloo, A. G. et al. Efficacy of a green tea extract rich in catechin polyphenols and caffeine in increasing 24-h energy expenditure and fat oxidation in humans. The American journal of clinical nutrition 70, 1040–1045 (1999).
Klaus, S., Pultz, S., Thone-Reineke, C. & Wolfram, S. Epigallocatechin gallate attenuates diet-induced obesity in mice by decreasing energy absorption and increasing fat oxidation. International journal of obesity 29, 615–623, https://doi.org/10.1038/sj.ijo.0802926 (2005).
Yoshino, K., Hara, Y., Sano, M. & Tomita, I. Antioxidative effects of black tea theaflavins and thearubigin on lipid peroxidation of rat liver homogenates induced by tert-butyl hydroperoxide. Biological & pharmaceutical bulletin 17, 146–149 (1994).
Loche, E. & Ozanne, S. E. Early nutrition, epigenetics, and cardiovascular disease. Current opinion in lipidology 27, 449–458, https://doi.org/10.1097/MOL.0000000000000338 (2016).
Muralimanoharan, S. et al. Sexual dimorphism in the fetal cardiac response to maternal nutrient restriction. Journal of molecular and cellular cardiology 108, 181–193, https://doi.org/10.1016/j.yjmcc.2017.06.006 (2017).
Wang, C. C. et al. Tea epigallocatechin-3-gallate increases 8-isoprostane level and induces caudal regression in developing rat embryos. Free radical biology & medicine 43, 519–527, https://doi.org/10.1016/j.freeradbiomed.2007.04.034 (2007).
Chu, K. O. et al. Pharmacokinetic studies of green tea catechins in maternal plasma and fetuses in rats. Journal of pharmaceutical sciences 95, 1372–1381, https://doi.org/10.1002/jps.20594 (2006).
Chu, K. O. et al. Uptake and distribution of catechins in fetal organs following in utero exposure in rats. Human reproduction 22, 280–287, https://doi.org/10.1093/humrep/del353 (2007).
Morita, O. et al. Effects of green tea catechin on embryo/fetal development in rats. Food and chemical toxicology: an international journal published for the British Industrial Biological Research Association 47, 1296–1303, https://doi.org/10.1016/j.fct.2009.03.005 (2009).
Moreno, M. F. et al. Metabolic profile response to administration of epigallocatechin-3-gallate in high-fat-fed mice. Diabetology & metabolic syndrome 6, 84, https://doi.org/10.1186/1758-5996-6-84 (2014).
Okuda, M. H. et al. Green tea extract improves high fat diet-induced hypothalamic inflammation, without affecting the serotoninergic system. The Journal of nutritional biochemistry 25, 1084–1089, https://doi.org/10.1016/j.jnutbio.2014.05.012 (2014).
Cunha, C. A. et al. Green tea extract supplementation induces the lipolytic pathway, attenuates obesity, and reduces low-grade inflammation in mice fed a high-fat diet. Mediators of inflammation 2013, 635470, https://doi.org/10.1155/2013/635470 (2013).
Santamarina, A. B. et al. Decaffeinated green tea extract rich in epigallocatechin-3-gallate prevents fatty liver disease by increased activities of mitochondrial respiratory chain complexes in diet-induced obesity mice. The Journal of nutritional biochemistry 26, 1348–1356, https://doi.org/10.1016/j.jnutbio.2015.07.002 (2015).
Santana, A. A. et al. Decaffeinated green tea extract rich in epigallocatechin-3-gallate improves insulin resistance and metabolic profiles in normolipidic diet- but not high-fat diet-fed mice. The Journal of nutritional biochemistry 26, 893–902, https://doi.org/10.1016/j.jnutbio.2015.03.001 (2015).
Shiraishi, M. et al. Association between the serum folate levels and tea consumption during pregnancy. Bioscience trends 4, 225–230 (2010).
Hachul, A. C. et al. Oligofructose supplementation (10%) during pregnancy and lactation does not change the inflammatory effect of concurrent trans fatty acid ingestion on 21-day-old offspring. Lipids in health and disease 12, 59, https://doi.org/10.1186/1476-511X-12-59 (2013).
Santamarina, A. B. et al. Green Tea Extract Rich in Epigallocatechin-3-Gallate Prevents Fatty Liver by AMPK Activation via LKB1 in Mice Fed a High-Fat Diet. PloS one 10, e0141227, https://doi.org/10.1371/journal.pone.0141227 (2015).
Forester, S. C. & Lambert, J. D. The role of antioxidant versus pro-oxidant effects of green tea polyphenols in cancer prevention. Molecular nutrition & food research 55, 844–854, https://doi.org/10.1002/mnfr.201000641 (2011).
Suzuki, T., Pervin, M., Goto, S., Isemura, M. & Nakamura, Y. Beneficial Effects of Tea and the Green Tea Catechin Epigallocatechin-3-gallate on Obesity. Molecules 21 https://doi.org/10.3390/molecules21101305 (2016).
Hayakawa, S. et al. Anti-Cancer Effects of Green Tea by Either Anti- or Pro- Oxidative Mechanisms. Asian Pacific journal of cancer prevention: APJCP 17, 1649–1654 (2016).
Suzuki, Y., Miyoshi, N. & Isemura, M. Health-promoting effects of green tea. Proceedings of the Japan Academy. Series B, Physical and biological sciences 88, 88–101 (2012).
Fechtner, S., Singh, A., Chourasia, M. & Ahmed, S. Molecular insights into the differences in anti-inflammatory activities of green tea catechins on IL-1beta signaling in rheumatoid arthritis synovial fibroblasts. Toxicology and applied pharmacology 329, 112–120, https://doi.org/10.1016/j.taap.2017.05.016 (2017).
Mallat, Z. et al. Expression of interleukin-10 in advanced human atherosclerotic plaques: relation to inducible nitric oxide synthase expression and cell death. Arteriosclerosis, thrombosis, and vascular biology 19, 611–616 (1999).
Rankinen, T. et al. The human obesity gene map: the 2005 update. Obesity 14, 529–644, https://doi.org/10.1038/oby.2006.71 (2006).
Shen, C. L. et al. Green tea polyphenols mitigate bone loss of female rats in a chronic inflammation-induced bone loss model. The Journal of nutritional biochemistry 21, 968–974, https://doi.org/10.1016/j.jnutbio.2009.08.002 (2010).
Shen, C. L. et al. Green tea polyphenols attenuate deterioration of bone microarchitecture in female rats with systemic chronic inflammation. Osteoporosis international: a journal established as result of cooperation between the European Foundation for Osteoporosis and the National Osteoporosis Foundation of the USA 22, 327–337, https://doi.org/10.1007/s00198-010-1209-2 (2011).
Suchacki, K. J., Cawthorn, W. P. & Rosen, C. J. Bone marrow adipose tissue: formation, function and regulation. Current opinion in pharmacology 28, 50–56, https://doi.org/10.1016/j.coph.2016.03.001 (2016).
Shah, R. V. et al. Visceral adiposity and the risk of metabolic syndrome across body mass index: the MESA Study. JACC. Cardiovascular imaging 7, 1221–1235, https://doi.org/10.1016/j.jcmg.2014.07.017 (2014).
Wajchenberg, B. L. Subcutaneous and visceral adipose tissue: their relation to the metabolic syndrome. Endocrine reviews 21, 697–738, https://doi.org/10.1210/edrv.21.6.0415 (2000).
Bastard, J. P. et al. Recent advances in the relationship between obesity, inflammation, and insulin resistance. European cytokine network 17, 4–12 (2006).
Phillips, L. K. & Prins, J. B. The link between abdominal obesity and the metabolic syndrome. Current hypertension reports 10, 156–164 (2008).
Waki, H. & Tontonoz, P. Endocrine functions of adipose tissue. Annual review of pathology 2, 31–56, https://doi.org/10.1146/annurev.pathol.2.010506.091859 (2007).
Nakamura, Y. K. & Omaye, S. T. Metabolic diseases and pro- and prebiotics: Mechanistic insights. Nutrition & metabolism 9, 60, https://doi.org/10.1186/1743-7075-9-60 (2012).
Tsukumo, D. M. et al. Loss-of-function mutation in Toll-like receptor 4 prevents diet-induced obesity and insulin resistance. Diabetes 56, 1986–1998, https://doi.org/10.2337/db06-1595 (2007).
Reeves, P. G. Components of the AIN-93 diets as improvements in the AIN-76A diet. The Journal of nutrition 127, 838S–841S (1997).
Aebi, H. Catalase in vitro. Methods in enzymology 105, 121–126 (1984).
Acknowledgements
This work was supported by Conselho Nacional de Desenvolvimento Científico, Tecnológico (CNPq), Fundação de Amparo à Pesquisa do Estado de São Paulo (FAPESP 2014/19508-7), and Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES). L.M.O., C.M.O.N., E.B.R., P.O.C., A.C.H.F.S. and A.C.L.H. are recipients of CNPq fellowships.
Author information
Authors and Affiliations
Contributions
Designed the study, analyzed the results and wrote the manuscript: A.C.L.H., C.M.O.N. and L.M.O. Analyzed the composition of the green tea: P.O.C., A.C.H.F.S. Performed the experiments: A.C.L.H., N.I.P.N., V.T.B., M.F.M., P.O.C., A.C.H.F.S. Contributed with reagents/materials/analysis tools: L.M.O., E.B.R., C.M.O.N. All authors revised the manuscript.
Corresponding author
Ethics declarations
Competing Interests
The authors declare that they have no competing interests.
Additional information
Publisher's note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Hachul, A.C.L., Boldarine, V.T., Neto, N.I.P. et al. Effect of the consumption of green tea extract during pregnancy and lactation on metabolism of mothers and 28d-old offspring. Sci Rep 8, 1869 (2018). https://doi.org/10.1038/s41598-018-20174-x
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-018-20174-x
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
