
Abstract
Antimicrobial resistance (AMR) is a global health issue. In an effort to minimize this threat to astronauts, who may be immunocompromised and thus at a greater risk of infection from antimicrobial resistant pathogens, a comprehensive study of the ISS “resistome’ was conducted. Using whole genome sequencing (WGS) and disc diffusion antibiotic resistance assays, 9 biosafety level 2 organisms isolated from the ISS were assessed for their antibiotic resistance. Molecular analysis of AMR genes from 24 surface samples collected from the ISS during 3 different sampling events over a span of a year were analyzed with Ion AmpliSeq™ and metagenomics. Disc diffusion assays showed that Enterobacter bugandensis strains were resistant to all 9 antibiotics tested and Staphylococcus haemolyticus being resistant to none. Ion AmpliSeq™ revealed that 123 AMR genes were found, with those responsible for beta-lactam and trimethoprim resistance being the most abundant and widespread. Using a variety of methods, the genes involved in antimicrobial resistance have been examined for the first time from the ISS. This information could lead to mitigation strategies to maintain astronaut health during long duration space missions when return to Earth for treatment is not possible.
Similar content being viewed by others
Introduction
The International Space Station (ISS) is not a sterile environment. Microbial monitoring of air and surfaces have shown that many different microbes exist in this closed, harsh environment, some of which are opportunistic pathogens of human origin1,2,3,4. The presence of such microorganisms within this environment poses a serious risk to crew health, as opportunistic pathogens have shown increased virulence while grown in space and prolonged spaceflight weakens an astronaut’s immune system, rendering him/her more susceptible to infection5. Some human associated opportunistic pathogens that have been isolated from the ISS include Staphylococcus aureus, S. haemolyticus, S. hominis, Acinetobacter pitti, Pantoea conspicua and members of the family Enterobacteriaceae, while others such as Propionibacterium and Corynebacterium have been detected with culture-independent, molecular based methods2,5. This study however, does not discuss virulence – a key factor that determines whether or not a microbe could be harmful to humans. Thus, the AMR gene carrying bacteria reported in this study do not necessarily pose a threat to astronauts on the ISS.
Antibiotics are the primary treatment strategy to combat infections, but this approach may not be as easy to implement on the ISS or on future deep space missions. Studies have shown that some organisms become less susceptible to antibiotic action when exposed to spaceflight. For example, the minimum inhibitory concentration (MIC) of colistin and kanamycin for E. coli, isolated from an astronaut aboard Salyut 7, increased from 4 μg/ml to >16 μg/ml, compared to the ground control, while that of S. aureus increased two fold for oxacillin, chloramphenicol, and erythromycin6. Increased antibiotic resistance has also been noted on Discovery shuttle and Mir space station flight missions7. The mode of action for this resistance is unclear, but it could be due to increased mutations in bacterial genes. A study comparing S. epidermidis grown on the ISS vs Earth, for 122 hours, showed a 24-fold higher frequency in mutations in the rpoB gene (beta subunit of RNA polymerase) when grown on the ISS, which rendered the isolates resistant to rifampicin8.
In a study comparing anti-microbial resistance in bacteria isolated from the ISS and the Antarctic Research Station Concordia, Schiwon et al. found that 75.8% of the ISS isolates were resistant to one or more antibiotics, whereas only 43.6% of the Antarctic isolates were resistant9. From the ISS, 86.2% of the isolates had relaxase and/or transfer genes encoded on plasmids which can promote the dissemination of AMR genes from one microorganism to another9. Importantly, the same genera and even most of the same species were examined between the two locations. The two major differences between these two confined, hostile and extreme environments are microgravity and radiation, which may contribute to the higher prevalence of antibiotic resistant microorganisms on the ISS. However, unlike the Schiwon et al.9 study that measured AMR profiles from isolated strains, this study has examined AMR gene profiles directly from DNA, without culturing.
It is well established that only a small proportion of microorganisms in any given environment can be cultured with standard laboratory techniques and the ISS is no different2. A recent microbial tracking study performed by our lab showed that less than 30% of intact, potentially viable microorganisms (i.e. samples treated with propidium monoazide (PMA) which prevent amplification of dead cells in molecular analyses) could be cultured, leaving 70% of organisms and their resistomes (i.e. the collection of AMR genes within a genome) unaccounted for (Checinska Sielaff et al., manuscript in preparation). In a recent study, propidium iodide ions were shown to move across intact cell membranes for Dinoroseobacter shibae and Bacillus subtilis using BacLight fuorescence staining and hypothesized that high membrane potential facilitated breakthrough of the double-charged propidium iodide and marked viable cells as dead10. In contrast, using polymerase chain reaction (PCR) we have shown that PMA was able to intercalate DNA in compromised cells which eventually blocked the amplification11.
In this study, we performed a thorough analysis of anti-microbial resistance genes within the ISS environment, using traditional and state-of-the art molecular techniques. Samples from eight ISS locations were obtained from three separate sampling events. The bacteria isolated from the samples were tested for resistance to several antibiotics. Concurrently, DNA from PMA treated samples was subjected to a target amplification of 518 antimicrobial genes, designed to match the content of the Antimicrobial Resistance Determinant Microarray (ARDM)12. Samples were treated with PMA before DNA extraction so that our focus would be on intact, potentially viable cells and not dead ones11 as live cells pose a greater risk to astronaut health. In addition, metagenome data generated from the DNA of the same PMA treated samples were mined for the presence of AMR and compared. This is the first report that presents the identification of the most common AMR genes on the ISS without culturing and correlates the results with phenotypic susceptibility and WGS data of cultured isolates. The study of the ISS resistome is crucial for the development of better surveillance and mitigation procedures on future long-duration spaceflight.
Results
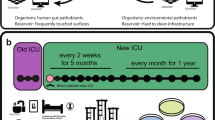
Twenty-four samples were collected during three sampling events, over a span of a year, from eight locations onboard the ISS (Table 1). Samples were both (i) cultured, to obtain isolates for phenotypic antibiotic susceptibility testing and WGS and (ii) subjected to DNA extraction, after propidium monoazide (PMA) treatment, to allow for Ion AmpliSeq™ and metagenomics analyses. PMA is a dye that intercalates with nucleic acid from dead cells and free DNA, allowing one to assess only intact bacteria within a sample11. A summary of the analysis work flow is presented in Fig. 1.

Schematic of analyses performed on 24 samples collected from various locations across the ISS over 3 flight missions.
Phenotypic susceptibility tests of cultured organisms
Fifty-seven Biosafety Level-2 (BSL-2) microorganisms, isolated from the ISS, were tested for their resistance against the following 9 antibiotics using the disc diffusion method: cefazolin, cefoxitin, ciprofloxacin, erythromycin, gentamycin, oxacillin, penicillin, rifampin and tobramycin. Table S1 shows the measured zone of inhibition along with the percentage of resistant, intermediate resistant and susceptible strains to each antibiotic. Out of the total 57 strains tested, 92% were resistant to penicillin while only 14% were resistant to gentamicin and ciprofloxacin. For the remaining 6 antibiotics the resistance percentages were as follows: oxacillin (68%), rifampin (66%), erythromycin (64%), cefoxitin (49%), cefazolin (29%) and tobramycin (19%). Enterobacter bugandensis, a novel species isolated from the ISS (manuscript in preparation), was resistant to the most antibiotics: all 6 E. bugandensis isolates tested were resistant against cefazolin, cefoxitin, erythromycin, oxacillin, penicillin and rifampin, while for ciprofloxacin and gentamycin, strains were either resistant or intermediate resistant. For tobramycin some strains were resistant, some intermediate resistant and some susceptible. S. haemolyticus on the other hand, was not resistant to any of the antibiotics tested. Due to the serious health concerns of methicillin resistant S. aureus (MRSA), which are resistant to a variety of antibiotics, all 12 S. aureus isolates obtained on the ISS were further tested with the Vitek system, which included a more comprehensive panel of antibiotics. Antibiotics that were tested with both the disc diffusion assay and Vitek showed the same results (with the exception of three isolates which were susceptible with Vitek but resistant with the disc assay) and the additional antibiotics tested with Vitek showed all strains to be susceptible, except four that were resistant to clindamycin (Table S1). In addition, assessment of methicillin resistance (i.e. MRSA) was performed using the HardyCHROM MRSA biplates and all 12 S. aureus strains tested were shown to not be MRSA.
Examining bacterial genomes for antibiotic resistance genes
WGS was performed on 15 of the 57 BSL-2 organisms that were isolated from the ISS. In addition, six S. aureus mutant colonies that were present within the clear zone of inhibition after exposure to rifampin, also had their genomes sequenced. The presence or absence of AMR genes in each isolate is displayed in Fig. 2A. It is evident that S. aureus genomes show a distinct AMR gene profile compared to other staphylococcal species and other genera. Furthermore, amongst S. aureus isolates, the two that were resistant to erythromycin (IF4SW_P1_Sau and IF7SW_P3_Sau), as determined by the disc diffusion assay and Vitek testing, display a distinct AMR gene profile compared to all the others, indicating a possible genetic basis for resistance.

Whole genome sequencing analysis of 21 BSL-2 strains isolated from the ISS. (A) The heatmap shows the gene presence (blue) and absence (red) profile of anti-microbial resistance (AMR) genes in whole genome sequenced isolates. Rows represent isolates and columns report the AMR genes found in that specific isolate. Genes with at least 90% nucleotide identity and 90% of the query gene aligned were considered to be positive matches. Resistance genes include the gene name and function class. Where multiple distinct genes of the same type were identified a numeric integer (e.g. 1, 2) is used to denote the difference. Sau: Staphylococcus aureus, Sha: Staphylococcus haemolyticus, Sho: Staphylococcus hominis, Pco: Pantoea conspicua, Epi: Enterobacter bugandensis, Api: Acinetobacter pitti, and Kqu: Klebsiella quasipneumoniae. Naming nomenclature: IF and IIF refers to isolates cultured from Flight 1 and Flight II samples respectively. The number after IF or IIF refers to the location sampled. For example IF6SW.P2.RA_Sau refers to S.aureus cultured from Flight 1, location 6 sample. (B) Antibiotic resistance genes in Enterobacter bugandensis IF2SW-P2, isolated from location 2 (waste and hygiene compartment), during Flight 1. Six specific antibiotic resistance gene clusters were shown to be present in four pathogens but absent from Enterobacter species isolated on Earth. Annotations were done using the NCBI Prokaryotic Genome Annotation Pipeline and visualized with SEED viewer50. Chromosomal region window (16000 bp) is centered on the focus gene, which is red and numbered one. Set of gene with high similarity in sequence are grouped with same color and number. Grey represent genes with similar relative position and functionally coupled.
Mutations in the rpoB gene have been implicated in rifampin resistance8, therefore the rpoB gene sequence from the spontaneous rifampin resistant mutants and the sensitive parent strains were compared. With the exception of a base pair change from C to T at position 144, there were no other differences between parent and mutant strains.
Seven antibiotic resistance gene clusters were analyzed in the novel species E. bugandensis, isolated from the waste and hygiene compartment on the ISS (manuscript in preparation). These AMR gene clusters were associated with known pathogens and were not found in Enterobacter species isolated on Earth (Fig. 2B). This suggests that horizontal gene transfer (HGT) might have occurred but more research is needed to determine if HGT happened on the ISS.
Anti-microbial resistance gene profiles on the ISS using Ion AmpliSeq™
To survey the AMR gene content of a large collection of samples and independent of culture restrictions, DNA isolated from the 24 ISS samples was analyzed using Ion AmpliSeq™. Ion AmpliSeq™ technology consists of a pool of primer pairs, with each pair designed to amplify a specific gene. In our study, this panel consisted of primer pairs designed against 518 known antibiotic resistance genes from a variety of bacteria12. Of the 518 genes that were tested for, a total of 123 were detected, but only those that were over 100 reads from the Ion Torrent sequencer were kept for further analysis. This was a threshold setting to remove potential “false positives” which can arise from non-specific primer binding, mutations during amplification and Ion Torrent sequencing error13,14. A comprehensive summary of the 63 detected genes (those with read counts over 100), their annotation numbers in NCBI (GI), the microorganisms they are related to, their function and the resistance they confer are found in Table S2.
Figure 3 shows the relative proportion of each detected AMR gene within a sample, with each colored bar representing a different AMR gene. The different colors and color patterns for each location suggest differences in spatial distribution of AMR genes at each location. This variability was confirmed with biplots, which show the distribution of the samples in relation to each other, and the contribution of each variable (i.e. gene) to the overall composition of that sample (Fig. S1). In addition to spatial differences, the dendrogram in Fig. 4A, shows temporal differences as well, with a clear separation of Flight 3 (F3) samples from Flight 1 (F1) and Flight 2 (F2). While there was no overall separation between F1 and F2, most paired samples (i.e. F1_8 vs F2_8) do cluster apart between the two sampling events (i.e. locations 1, 2, 4, 6, 7, 8). The separation between F1/F2 and F3 may be attributed to the low relative abundances of AMR genes present on the ISS during F3. As the heatmap in Fig. 4B shows, F3 samples had a higher number of genes shaded yellow, an indicator of abundances lower than the geometric mean abundance of all the 24 samples combined.

Relative abundance of anti-microbial resistance genes (AMR) aboard the ISS. Bar plot showing the relative abundances of AMR genes in samples collected from different locations aboard the ISS, during three sampling events. F refers to the flight number, followed by the location (i.e. F1_1 is Flight 1 location 1 (see Table 1 for more details)). Each color represents a gene and the height of the colored box represents the proportion of that gene within that sample. The remaining fraction, colored grey, groups genes that were less than 1% abundance in that sample. The legend shows the gene name (i.e. mecA) followed by the antibiotic that gene confers resistance to (i.e. methicillin). The number in front of the gene name is the personal identifier that can be matched to the metadata listed in Table S2. The legend is read from bottom to top, as indicated by the red arrow, with the bottom AMR gene on the legend corresponding to the bottom colored box on the bar plot.

Temporal comparison of AMR gene profiles. (A) Hierarchical clustering of Euclidian distances of centered log ration transformed (clr) data, using the Ward clustering method. Red values are AU (Approximately Unbiased) p-values, and green values are BP (Bootstrap probability) values. Clusters with AU p-values larger than 90% are highlighted by rectangles (significance level 0.10), meaning that there is a 90% certainty of these clusters being a distinct group. F”x” refers to the flight number, followed by the location (i.e. F1_1: Flight 1, location 1). (B) Heat map of centered log ratio transformed data. Red and orange boxes indicate samples that have genes with relative abundances higher than the geometric mean abundance (represented as 0), which has been calculated from all samples. Simply, it means that these samples have a high abundance overall. Boxes in yellow indicate samples having genes with relative abundances that are lower than the geometric mean abundance (i.e. low abundance overall). The columns show the samples and the rows show the gene, followed by the antibiotic that the gene confers resistance to. The number in front of the gene name is the personal identifier that can be matched to the metadata listed in Table S2 and to the genes shown in Fig. 3.
At the functional level, the genes for beta-lactam and trimethoprim resistance were abundant and widespread across the ISS (Fig. 5A). However, when a metagenomics approach was used to examine the AMR gene profiles, genes for beta-lactam resistance, while found in every sample, were not as abundant compared to other functional groups and genes for trimethoprim resistance was not detected (Fig. 5B). The metagenomics approach also showed that genes conferring resistance to metals (i.e. zinc, copper etc) and those involved in multi-drug efflux pumps were the most abundant across the ISS, both of which were not detected with Ion AmpliSeq™. Fluoroquinolone resistance was also highly represented across the ISS, while Ion AmpliSeq™ only detected this gene family in only 3 samples. Ion AmpliSeq™ detected 23 antibiotic groups and metagenomics 13, with 6 shared between the two methods: aminoglycosides, beta-lactams, quinols (i.e. fluoroquinolones), fosfomycin, methicillin and streptomycin. At the gene level AmpliSeq™ and metagenomics both detected tetR, macB, fosA, and femB. As the two methods use a different set of reference genes and the raw data is processed differently, the fact that there are differences is not surprising, but it does highlight the usefulness of using a variety of analytical approaches.

Distribution of antibiotic resistance across samples at the function level from both Ion AmpliSeq™ and metagenomics analysis. (A) Heat map showing read counts of AMR genes grouped together based on the class of antibiotic they are resistant to, as detected by AmpliSeq™. Samples were rarefied to 1830 reads, with those lower than 1830 read counts removed during rarefaction. (B) Read counts of AMR genes grouped together based on class of antibiotic they are resistant to, as detected by metagenomics. Grey boxes indicate genes that were absent in a particular sample. Red indicates the highest read count and yellow, the lowest read count. Columns represent samples and rows, antibiotic resistance.
Multiple AMR genes can be transferred on a single plasmid, giving rise to multi-drug resistance in the recipient. We were therefore interested in assessing whether any genes were associated with each other and thus possibly found together. A correlation analysis showed statistically significant negative and positive correlations between the genetic determinants of AMR (see Table 2 for Spearman rho and P values).
Discussion
We have provided a comprehensive summary of the AMR genes present across the ISS over a span of a year using phenotypic assays, WGS, Ion AmpliSeq™ and metagenomics. The WGS of BSL-2 isolates showed the collective presence of 51 genes conferring resistance to 18 antibiotics. In most cases, the genotype of the sequenced strains could predict the phenotype, such as the presence of ermA and blaZ in S. aureus, which protects against erythromycin and penicillin respectively15 and the presence of oqxA and oqxB in K. quasipneumoniae, which protects against ciprofloxacin16, all of which were resistant to their respective antibiotics in the disc diffusion assay. In other cases, the AMR profiles were not as straightforward to interpret. For example, one S. aureus isolate, (IF4SW.P1_Sau) lacking the blaZ gene was still resistant to penicillin. In another case, the presence of norA in multiple S. aureus isolates did not protect against ciprofloxacin, even though transfer of this gene to susceptible strains has been proven to be enough to confer resistance17,18. The lack of consensus between genotype and phenotype in this case could be due to inadequate gene expression, which has been shown in numerous studies to be a barrier against resistance16. Moreover, this study was focused on the detection of genomic sequences of known AMR genes so any mutations and variations in gene expression of AMR genes were not analyzed in this study.
E. bugandensis was the most antibiotic resistant species tested with our phenotypic assay, with all six isolates being resistant to all antibiotics tested (with the exception of two that were susceptible to tobramycin). Enterobacter species, a member of the coliform group of bacteria, have been reported to be pathogenic and cause opportunistic infections in immunocompromised individuals and are commonly associated with urinary and respiratory tract infections19. Of note, approximately 6% of the isolates cultured from the ISS were identified as Enterobacter. Carbapenems, a class of beta-lactams, are the most effective treatment against Enterobacter infections as they remain stable against extended spectrum beta lactamases, unlike penicillin and oxacillin. Ertapanem is the only carbapenem included in the medical kit onboard the ISS, however this may need to be re-evaluated, as numerous clinical isolates of Enterobacter species have shown resistance to ertapanem, though not other carbapenems, such as imipenem and meropenem20,21. There is the possibility that our ISS Enterobacter isolates, like the clinical isolates, are resistant to ertapanem, and potentially other carbapenems, as the IMP-2 gene, a metallo beta-lactamase that is able to inhibit the action of this class of antibiotic, was detected in our samples22. Further evaluation of this resistance profile and any other multi-drug resistant microorganisms, onboard the ISS, should be made a priority, in order to assess risk of infection and to ensure that proper strategies are in place to combat such infections.
Ion AmpliSeq™ revealed a collective of 65 AMR genes onboard the ISS, representing resistance to 23 different antibiotics. Nine of those detected genes (qacB, blaZ, bla- OKP, oqxA, oqxB, fos, tet38, norA and bla1) were also identified in the WGS analysis of our selected cultured isolates. One of the detected genes was mecA, a biomarker for methicillin resistant strains of S. aureus, commonly referred to as MRSA. MRSA are strains of Staphylococcus that have become resistant to methicillin and other beta-lactams (i.e. oxacillin, penicillin) used to treat ordinary Staphylococcus infections can cause serious complications (skin infections, sepsis and pneumoniae) in immune-compromised individuals. Even though mecA was detected in numerous samples across the ISS via AmpliSeqTM, none of the 12 S. aureus isolates tested with Vitek were MRSA positive nor was mecA detected in their genomes. While mecA is commonly associated with S. aureus, it has also been detected in other Staphylococcus species23,24 which may have been the source of the detected mecA during this study. Despite the fact that none of the S. aureus isolates were MRSA at the time of collection, there is the possibility that they acquire mecA from other staphylococci. Indeed, the results of a hospital in the Netherlands, showed that mecA gene transfer occurred in a patient between mecA+ S. epidermidis and a mecA- strain of S. aureus25. This is not surprising as there is mounting evidence in favor of the “resistance gene reservoir” where commensal organisms in the body act as reservoirs for AMR genes that can be transferred to resident or transient bacteria, including pathogens26,27. Further studies assessing the proportion and types of mobile genetic elements within the ISS metagenome could shed some light on how readily AMR (and possibly virulence genes) may be acquired and transferred by bacteria on the ISS and if space conditions promote the transfer of AMR genes between different species and even different genera more readily then when these organisms are grown together on Earth.
A positive correlation was observed between oliquadox (oqxA and oqxB) and fosfomycin (fosA) resistance on the ISS, which conforms with ground studies showing the co-transfer of oqxA/oqxB and fosA genes from single plasmids28,29. A positive correlation was also observed between mupirocin and lincosamide/streptograminA resistance genes on the ISS. While studies have documented the co-transfer of β-lactam, quinolone, aminoglycoside, amphenicol and fosfomycin resistance genes to transconjugants28, this is the first study, to our knowledge, that has observed an association between muprirocin and lincosamide/streptogtanimA resistance genes.
It has been documented that metal exposure and the resulting resistance towards those metals leads to co-selection of unrelated antibiotic resistance30,31, most likely a consequence of metal resistance genes and AMR genes present on the same plasmid and/or transposon32. Indeed, bacterioplankton isolated from metal contaminated Ash basins were significantly more tolerant to a variety of antibiotics, which were not found in these basins, compared to bacterioplankton collected from basins void of metals33,34. Similar phenomena have been reported from metal contaminated freshwater34 and soil samples35. Our metagenomics data showed that the highest resistance genes across all time points and all locations were in fact metal resistance genes. It is unknown whether the material of the ISS surfaces that these organisms were collected from promotes metal resistance dissemination amongst the microbiota but if that is the case, this metal resistance may play a role in the spread and abundance of AMR genes on the ISS.
To summarize, this is the first study to utilize an array of molecular methods to characterize the resistome on the ISS unveiling the presence of numerous genes conferring resistance to 28 different antimicrobial agents. The results from this study highlight the importance of using a variety of analytical methods to get a comprehensive picture of the resistome on the ISS and to utilize this information to develop concrete antibiotic resistance mitigation strategies. Some of these strategies could include the development of bioinspired alternative drugs, cleaning procedures for the eradication of specific organisms, bacterial specific microbial monitoring and evaluation of actual doses of antibiotics to be given in space. This will help maintain crew health, especially on long duration flight missions, when return to Earth for treatment is not an option.
Methods
Sample kit preparation and sample collection
Sampling wipes were assembled at the Jet Propulsion Laboratory (JPL) following the procedure outlined by36. Briefly, each polyester wipe (9″ × 9″) (ITW Texwipe, Mahwah, NJ) was folded two times and soaked in 15 mL of sterile molecular grade water (Sigma-Aldrich, St. Louis, MO) for 30 minutes followed by transfer to a sterile zip lock bag. At the Ames Research Center (ARC, Moffett Field, CA), the wipes were packed along with other items, such as sterile gloves, as part of a kit sent to the ISS. Each sampling kit was then sent to the ISS on the Space-X cargo, and returned to the Earth on Soyuz TM-14 or Dragon capsule (Table 1). The kits were delivered to JPL immediately upon return to Earth.
During each sampling session on the ISS, only one astronaut collected samples from eight different locations. The description of each location is depicted in Table 1. Each wipe was used to collect a sample of one square meter. The control wipe (environmental control) was only taken out from the Zip lock bag, unfolded and packed back to the zip lock. The samples were stored at 4 °C until the return trip to Earth and subsequent processing.
Sample processing
The sample processing took place in a 10 K class cleanroom at JPL immediately upon return to Earth. Each wipe was aseptically taken out from the zip lock bag and transferred to a 500-mL bottle containing 200 mL of sterile PBS. The bottle with the wipe was shaken for two minutes followed by concentration with a Concentrating Pipette (Innova Prep, Drexel, MO) using 0.45 µm Hollow Fiber Polysulfone tips and PBS elution fluid. The environmental control and each sample were concentrated to 5 mL.
PMA treatment and DNA extraction
A 1.5-mL aliquot was treated with PMA to allow for amplification of DNA from only intact cells11. 18.25 µL of 25 µM PMA in water was added to each sample followed by 5-minute incubation at room temperature in the dark, followed by a 15-minute exposure to the activation system (PMA LED device, Biotium, Hayward, CA). 750 µl was then transferred to bead beating tubes containing Lysing Matrix E (MP Biomedicals, Santa Ana, CA) and subjected to bead beating for 60 s using the vortex sample holder (MO Bio, Carlsbad, CA). The bead-beaten half and the unprocessed half were combined, followed by DNA extraction with the Maxwell 16 automated system using the Maxwell 16 Tissue LEV Total RNA purification kit in accordance with the manufacturer’s instructions (Promega, Madison, WI). Resulting DNA samples in water (50 μL each) were stored at −20 °C until downstream analysis.
Isolation and identification of bacteria from samples
The concentrated samples were diluted in PBS (up to 10−6 of each original sample) and 100 µL (in duplicate), were plated on R2A agar for environmental bacteria (incubated at room temperature for 7 days), and blood agar (BA) for human commensals (incubated at 35 °C for two days). On average, five isolates of distinct morphologies were picked up for each location from each type of media. The isolates were archived in semi solid R2A slants (dilution 1:10) and stored at room temperature. For identification, each bacterial isolate was revived on R2A media. Once a culture was confirmed to be pure, DNA extraction was performed by colony PCR, UltraClean DNA kit (MO Bio, Carlsbad, CA) or Maxwell Automated System (Promega, Madison, WI). The 1.5 kb 16S rRNA gene was used to identify the bacterial isolates using the following primers: 27 F (5′-AGAGTTTGATCCTGGCTCAG-3′) and 1492 R (5′-GGTTACCTTGTTACGACTT-3′). The PCR conditions were as follows: denaturation at 95 °C for 5 min, followed by 35 cycles consisting of denaturation at 95 °C for 50 s, annealing at 55 °C for 50 s, and extension at 72 °C for 1 min 30 s and finalized by extension at 72 °C for 10 min. When 16S rRNA identification was ambiguous (B. cereus group isolates), the primers for gyrB (UP-1: 5′-GAA GTC ATC ATG ACC GTT CTG CAY GCN GGN GGN AAR TTY GA-3′ and UP-2R: 5′-AGC AGG ATA CGG ATG TGC GAG CCR TCN ACR TCN GCR TCN GTC AT-3′) were used and a 1.2-kb fragment of gyrase B was amplified as described previously37,38. The sequences were assembled using SeqMan Pro from DNAStar Lasergene Package (DNASTAR Inc., Madison, WI). The bacterial sequences were searched against EzTaxon-e database39 and identified based on the closest similarity to 16S rRNA sequences of bacterial type strains.
Whole Genome Sequencing of Opportunistic Strains
Genomic DNA (gDNA) extracted from 21 BSL-2 bacterial strains were sent to Lawrence Livermore National Laboratory for WGS using the Illumina NextSeq. 500 sequencing system (San Diego, CA). The concentration of the gDNA was determined using a Qubit fluorimeter (ThermoFisher Scientific, Waltham, MA). The sequencing library preparation was performed on the gDNA samples using the TruSeq DNA PCR-Free LT kit (Illumina, San Diego, CA) according to the manufacturer’s protocol. Briefly, the concentration of each gDNA was normalized to 40 ng/µl and 55 µl of each sample was fragmented on a Covaris M220 ultrasonicator (Covaris, Woburn, MA) using settings to generate 550 base pair inserts. The Illumina TruSeq protocol was further followed to repair the fragmented DNA ends, adenylate the 3’ ends of the fragments, and finally ligate Illumina sequencing adaptors to the fragmented DNAs.
The libraries were validated using the Library Quantification Kit for Illumina platforms (KAPA, Wilmington, MA) following manufacturer’s protocols. Following the quantification, each library was normalized, pooled, and denatured according to Illumina protocols. Briefly, each library was normalized to 0.5 nM and 5 µl of each normalized sample was pooled together. The 40-µL pooled sample was denatured by combining with 40 µL of 0.2N NaOH. The denatured, pooled sample was further diluted to a concentration of 1.8 pM where it was mixed with denatured PhiX control DNA (Illumina, San Diego, CA) at a concentration of 1%. This sample was loaded on to the NextSeq Sequencer using the NextSeq High Output v2 300 cycle sequencing kit (Illumina, San Diego, CA). The NextSeq was set up to run a paired end, 150 nucleotide (2 × 150) run with single indexing.
Data assembly and gene annotation
Draft assemblies of the bacterial isolates were generated as described in40. Starting with the draft assemblies as input, genes were predicted for each genome using Glimmer v. 3.0241 using the g3-from-scratch.csh script with default settings to generate genome specific gene models. Each gene was searched against a collection of curated genes taken from the comprehensive antibiotic resistance database (CARD)42, ResFinder43 and ARG-ANNOT44. Each gene was searched against the curated gene database using the top hit from a blastn search (NCBI BLAST v2.2.27+) with word_size 8, num_threads 20, outfmt 7, max_target_seqs. 1, and all other options set to default. A table was generated recording individual genes that matched to an annotated resistance gene for each organism with at least 90% identity between the query and reference gene over at least 90% of the length of reference gene. Gene labels and gene function were manually assigned based on annotation assignments given in the included curated gene databases.
Phenotypic antibiotic resistance testing
The isolates were streaked from glycerol stocks onto R2A plates. A single colony was inoculated into 5-mL Tryptic Soy Broth (TSB) and grown overnight at 30 °C. Aliquots of 100 µL were plated on TSA. Agar diffusion discs (BD BBLTM Sensi-DiscTM, Franklin Lakes, NJ) were placed aseptically on a plate and the strains were incubated at 37 °C for 24 hours. The tested antibiotics included: 30-µg cefazolin (CZ-30); 30-µg cefoxitin (FOX-30), 5-µg ciprofloxacin (CIP-5), 15 µg erythromycin (E-15), 10-µg gentamicin (GM-10), 1-µg oxacillin (OX), 10-µg penicillin (P-10), 5-µg rifampin (RA-5), and 10-µg tobramycin (NN-10). The diameter of inhibition zones was measured for each antibiotic disk and recorded in millimeters. The resistance results were compared with the zone diameter interpretive charts provided by the manufacturer. When the spontaneous mutants were present in response to some antibiotics, they were isolated, subcultured and tested for the specific antibiotic resistance.
Vitek®2 Compact AST-GP67 cards (BioMerieux, Inc., Hazelwood, MO) were selected to test the S. aureus isolates. The AST-GP67 cards are designed for testing gram-positive cocci. The tested antibiotics were ampicillin, benzylpenicillin, beta-lactamase, cefoxitin, ciprofloxacin, clindamycin, erythromycin, gentamicin, levofloxacin, linezolid, moxifloxacin, nitrofurantoin oxacillin, quinupristin/dalfopristin, rifampicin, streptomycin, tetracycline, tigecycline, trimethoprim/sulfamethoxazole and vancomycin. The cards were initially analyzed for expected outcomes by running the manufacturers recommended quality control organisms S. aureus ATCC 29213, ATCC BAA-1026, ATCC BAA-976, and ATCC BAA-977, and the results were as predicted (data not shown). Liquid aliquots of the organisms were shipped overnight from JPL to AlloSource® (Centennial, CO) and refrigerated upon arrival. The aliquot of 0.1 mL subcultures of each microorganism was spread onto Trypticase Soy Agar (TSA) plates (BD BBLTM) and incubated overnight at 30–35°C. Using a sterile cotton swab, a pure colony of each organism was suspended in 3.0 mL sterile 0.9% saline at 0.55 McFarland using a DensiCheck® densitometer (BioMerieux, Inc.). A second dilution was prepared by aliquoting 0.28 mL of the initial inoculant into a fresh 3.0 mL sterile 0.9% saline solution. The swab was inoculated onto a TSA plate to check for inoculant purity. Both concentrations of each organism were loaded into separate AST-GP67 cards and analyzed by the Vitek®2 Compact (BioMerieux, Inc.) applying the CLSI+ Natural Resistance susceptibility standards per the manufacturers recommendations. Reports were verified by AlloSource® technicians. Confirmation of methicillin sensitivity (MRSA) was performed using HardyCHROM MRSA biplates (Hardy Diagnostics, Santa Maria, CA). Each isolate was streaked on both sides of the bi-plate and incubated at 30–35 °C overnight. Isolate sensitivity was interpreted using the manufacturers recommendation
Ion AmpliSeq™
Library preparation and targeted Sequencing
Ion AmpliSeq™ libraries (Life Technologies) were constructed with the Ion AmpliSeq™ library protocol as previously described45. The PCR conditions were as follows: 2X primer mix, 6 µL of sample and 16 PCR cycles consisting of a 99 °C for 15 sec; 60 °C for 4 min. Both primer pools were utilized to create two unique Ion AmpliSeq™ libraries for each metagenome sample. All libraries were quality checked using the Agilent BioAnalyzer and quantitated using the Ion Library Quantitation Kit. Template was diluted to target 10–30% enriched beads and clonally amplified using Ion PGM OT2 400 kit. Enriched beads were sequenced using the Ion PGM 400 Sequencing kit on an Ion 318™ chip at default instrument parameters. Libraries were sequenced in multiplex.
Ion AmpliSeq™ panel design
An Ion AmpliSeq™ custom panel (AmpliSeq MDR v.1, Thermo Scientific, South San Francisco, CA, USA) was designed to match the content of the Antimicrobial Resistance Determinant Microarray (ARDM)12. Primer design was performed by Life Technologies using the Ion AmpliSeq™ strategy and the Ion AmpliSeq™ software. The Ion AmpliSeq MDR v.1 consists of two primer pools that are capable of generating a total of 1,358 amplicons targeting 518 known, non-SNP mediated resistance determinants. The assay was designed to generate two unique amplicons per target for 404 resistance genes. For the remaining 114 genes corresponding to bla (beta-lactam) and qnr (quinolone) resistance determinants, the assay was designed to generate a sufficient number of overlapping amplicons to span the entire gene. Amplicon sequences are available upon request. Primer information is not supplied by Life Technologies.
Amplicon sequence read quality control
Sequence read files were imported into CLC Genomics Workbench v 8.0 (CLC Inc, Aarhus, Denmark). Reads were trimmed at default parameters based on quality score (≤0.05), ambiguous nucleotides (≤2) and filtered on read length (≥75 bp). Trimmed reads were saved as a separate file and used in all downstream bioinformatics analyses.
Reference mapping
Sequence read mapping was performed using CLC Genomics Workbench v8.0 CLC Reference Mapper was run with default settings (Insertion cost = 3, Deletion cost = 3, Mismatch cost = 2, Length fraction = 0.5 and similarity fraction = 0.8). Mapping of all sequence reads was performed using a multi-fasta file that contained the PCR amplicons that comprise the ARDM array.
The raw sequence reads have been deposited into the NCBI SRA database under accession #SRP082440.
Bioinformatics analysis
Dendrogram, heatmaps, barplot and biplots were all generated in R (http://www.R-project.org/). Relative abundances displayed in the bar plot were generated by taking the read count of a gene in a sample and dividing by the total read count of that sample. Data was centered log ratio (clr) transformed46 using the “compositions” package in R. The dendrogram was created with the “pvclust” package, which assesses the uncertainty in hierarchical cluster analysis. The heatmap was generated with the “gplot” package. Counts rarefied to 1830 reads were used in the heatmap and were generated in QIIME using the single_rarefaction.py script. Correlation analyses between the microbiome and anti-microbial resistance (AMR) genes were performed as per47. Briefly, the Spearman’s rank correlation between the relative abundances of each genus (clr transformed) in 128 inferred technical replicates and clr transformed AMR relative abundances were calculated with the aldex.corr function from the ALDEx2 package in R. Spearman’s rho values were converted to P-values and corrected by the Benjamini-Hochberg method. Correlations between AMR genes were performed as above, with the AMR dataset substituted for the microbiome dataset in the script.
Metagenome analysis
Extracted DNA was sent to Second Genome (South San Francisco, CA) for sequencing. All samples were quantified via the Qubit® Quant-iT dsDNA High Sensitivity Kit (Invitrogen, Life Technologies, Grand Island, NY) to ensure that they met minimum concentration and mass of DNA. Samples were prepared for sequencing with the Illumina Nextera kit and quantified with Quant-iT dsDNA High Sensitivity assays. Libraries were pooled and run with 100 bp paired-end sequencing protocols on the Illumina HiSeq. 2500 platform. The NGS QC Toolkit v 2.3 was used to filter the raw data for high-quality (HQ) vector- and adaptor-free reads for genome assembly (cutoff read length for HW, 80%; cutoff quality score, 20)48. Filtered DNA sequences were mapped against a reference database of all proteins within the KEGG databases using MEGAN649. Reads over 100 for each gene, that were predicted for antimicrobial resistance were clustered together and visualization was done using MEGAN. A total of 8 million reads were generated for each sample. The heatmap used to visualize the data was generated in R using the gplot function.
Availability of sequence data
The data presented in this manuscript are available in the NCBI Sequence Read Archive under accession number: SRP082440. The raw metagenomics sequences can be downloaded from the following GeneLab website (https://genelab-data.ndc.nasa.gov/genelab/accession/GLDS-66/).
Disclaimer
This document was prepared as an account of work sponsored by an agency of the United States government. Neither the United States government nor Lawrence Livermore National Security, LLC, nor any of their employees makes any warranty, expressed or implied, or assumes any legal liability or responsibility for the accuracy, completeness, or usefulness of any information, apparatus, product, or process disclosed, or represents that its use would not infringe privately owned rights. Reference herein to any specific commercial product, process, or service by trade name, trademark, manufacturer, or otherwise does not necessarily constitute or imply its endorsement, recommendation, or favoring by the United States government or Lawrence Livermore National Security, LLC. The views and opinions of authors expressed herein do not necessarily state or reflect those of the United States government or Lawrence Livermore National Security, LLC, and shall not be used for advertising or product endorsement purposes.
References
Ichijo, T., Yamaguchi, N., Tanigaki, F., Shirakawa, M. & Nasu, M. Four-year bacterial monitoring in the International Space Station—Japanese Experiment Module “Kibo” with culture-independent approach. Npj Microgravity 2, 16007, https://doi.org/10.1038/npjmgrav.2016.7, http://www.nature.com/articles/npjmgrav20167-supplementary-information (2016).
Checinska, A. et al. Microbiomes of the dust particles collected from the International Space Station and Spacecraft Assembly Facilities. Microbiome 3, 50, https://doi.org/10.1186/s40168-015-0116-3 (2015).
Venkateswaran, K. et al. International Space Station environmental microbiome - microbial inventories of ISS filter debris. Applied microbiology and biotechnology 98, 6453–6466, https://doi.org/10.1007/s00253-014-5650-6 (2014).
Castro, V. A., Thrasher, A. N., Healy, M., Ott, C. M. & Pierson, D. L. Microbial Characterization during the Early Habitation of the International Space Station. Microbial ecology 47, 119–126, https://doi.org/10.1007/s00248-003-1030-y (2004).
Klaus, D. M. & Howard, H. N. Antibiotic efficacy and microbial virulence during space flight. Trends in Biotechnology 24, 131–136, https://doi.org/10.1016/j.tibtech.2006.01.008 (2006).
Tixador, R. et al. Study of minimal inhibitory concentration of antibiotics on bacteria cultivated in vitro in space (Cytos 2 experiment). Aviat Space Environ Med 56, 748–751 (1985).
Taylor, P. W. Impact of space flight on bacterial virulence and antibiotic susceptibility. Infection and Drug Resistance 8, 249–262, https://doi.org/10.2147/IDR.S67275 (2015).
Fajardo-Cavazos, P. & Nicholson, W. L. Cultivation of Staphylococcus epidermidis in the Human Spaceflight Environment Leads to Alterations in the Frequency and Spectrum of Spontaneous Rifampicin-Resistance Mutations in the rpoB Gene. Front Microbiol 7, 999, https://doi.org/10.3389/fmicb.2016.00999 (2016).
Schiwon, K. et al. Comparison of antibiotic resistance, biofilm formation and conjugative transfer of Staphylococcus and Enterococcus isolates from International Space Station and Antarctic Research Station Concordia. Microbial ecology 65, 638–651, https://doi.org/10.1007/s00248-013-0193-4 (2013).
Kirchhoff, C. & Cypionka, H. Propidium ion enters viable cells with high membrane potential during live-dead staining. J Microbiol Methods 142, 79–82, https://doi.org/10.1016/j.mimet.2017.09.011 (2017).
Vaishampayan, P. et al. New perspectives on viable microbial communities in low-biomass cleanroom environments. ISME J 7, 312–324, http://www.nature.com/ismej/journal/v7/n2/suppinfo/ismej2012114s1.html (2013).
Leski, T. A. et al. Molecular characterization of multidrug resistant hospital isolates using the antimicrobial resistance determinant microarray. PloS one 8, e69507, https://doi.org/10.1371/journal.pone.0069507 (2013).
McCall, C. M. et al. False positives in multiplex PCR-based next-generation sequencing have unique signatures. The Journal of molecular diagnostics: JMD 16, 541–549, https://doi.org/10.1016/j.jmoldx.2014.06.001 (2014).
Quail, M. A. et al. A tale of three next generation sequencing platforms: comparison of Ion Torrent, Pacific Biosciences and Illumina MiSeq sequencers. BMC genomics 13, 341, https://doi.org/10.1186/1471-2164-13-341 (2012).
Argudin, M. A. et al. Genotypes, exotoxin gene content, and antimicrobial resistance of Staphylococcus aureus strains recovered from foods and food handlers. Applied and environmental microbiology 78, 2930–2935, https://doi.org/10.1128/aem.07487-11 (2012).
Andres, P. et al. Differential distribution of plasmid-mediated quinolone resistance genes in clinical enterobacteria with unusual phenotypes of quinolone susceptibility from Argentina. Antimicrobial agents and chemotherapy 57, 2467–2475, https://doi.org/10.1128/aac.01615-12 (2013).
Ubukata, K., Itoh-Yamashita, N. & Konno, M. Cloning and expression of the norA gene for fluoroquinolone resistance in Staphylococcus aureus. Antimicrobial agents and chemotherapy 33, 1535–1539 (1989).
Yoshida, H., Bogaki, M., Nakamura, S., Ubukata, K. & Konno, M. Nucleotide sequence and characterization of the Staphylococcus aureus norA gene, which confers resistance to quinolones. Journal of bacteriology 172, 6942–6949 (1990).
Mezzatesta, M. L., Gona, F. & Stefani, S. Enterobacter cloacae complex: clinical impact and emerging antibiotic resistance. Future Microbiol. 7, 887–902, https://doi.org/10.2217/fmb.12.61 (2012).
Chung, H. S., Yong, D. & Lee, M. Mechanisms of ertapenem resistance in Enterobacteriaceae isolates in a tertiary university hospital. J Investig Med 64, 1042–1049, https://doi.org/10.1136/jim-2016-000117 (2016).
Woodford, N. et al. Ertapenem resistance among Klebsiella and Enterobacter submitted in the UK to a reference laboratory. International journal of antimicrobial agents 29, 456–459, https://doi.org/10.1016/j.ijantimicag.2006.11.020 (2007).
Riccio, M. L. et al. Characterization of the metallo-beta-lactamase determinant of Acinetobacter baumannii AC-54/97 reveals the existence of bla(IMP) allelic variants carried by gene cassettes of different phylogeny. Antimicrobial agents and chemotherapy 44, 1229–1235 (2000).
Osman, K. M. et al. Antimicrobial Resistance, Biofilm Formation and mecA Characterization of Methicillin-Susceptible S. aureus and Non-S. aureus of Beef Meat Origin in Egypt. Front Microbiol 7, 222, https://doi.org/10.3389/fmicb.2016.00222 (2016).
Petinaki, E., Arvaniti, A., Dimitracopoulos, G. & Spiliopoulou, I. Detection of mecA, mecR1 and mecI genes among clinical isolates of methicillin-resistant staphylococci by combined polymerase chain reactions. Journal of Antimicrobial Chemotherapy 47, 297–304, https://doi.org/10.1093/jac/47.3.297 (2001).
Wielders, C. L. C. et al. Evidence for in-vivo transfer of mecA DNA between strains of Staphylococcus aureus. Lancet 357, 1674–1675, https://doi.org/10.1016/S0140-6736(00)04832-7 (2001).
Huddleston, J. R. Horizontal gene transfer in the human gastrointestinal tract: potential spread of antibiotic resistance genes. Infection and Drug Resistance 7, 167–176, https://doi.org/10.2147/IDR.S48820 (2014).
Salyers, A. A., Gupta, A. & Wang, Y. Human intestinal bacteria as reservoirs for antibiotic resistance genes. Trends in microbiology 12, 412–416, https://doi.org/10.1016/j.tim.2004.07.004 (2004).
Fang, L. et al. Co-spread of metal and antibiotic resistance within ST3-IncHI2 plasmids from E. coli isolates of food-producing animals. Scientific reports 6, 25312, https://doi.org/10.1038/srep25312 (2016).
Ho, P. L. et al. Plasmid-Mediated OqxAB Is an Important Mechanism for Nitrofurantoin Resistance in Escherichia coli. Antimicrobial agents and chemotherapy 60, 537–543, https://doi.org/10.1128/aac.02156-15 (2015).
Flach, C. F. et al. Does antifouling paint select for antibiotic resistance? The Science of the total environment 590–591, 461–468, https://doi.org/10.1016/j.scitotenv.2017.01.213 (2017).
Knapp, C. W. et al. Antibiotic resistance gene abundances correlate with metal and geochemical conditions in archived Scottish soils. PloS one 6, e27300, https://doi.org/10.1371/journal.pone.0027300 (2011).
Pal, C., Bengtsson-Palme, J., Kristiansson, E. & Larsson, D. G. Co-occurrence of resistance genes to antibiotics, biocides and metals reveals novel insights into their co-selection potential. BMC genomics 16, 964, https://doi.org/10.1186/s12864-015-2153-5 (2015).
Stepanauskas, R. et al. Elevated Microbial Tolerance to Metals and Antibiotics in Metal-Contaminated Industrial Environments. Environmental Science & Technology 39, 3671–3678, https://doi.org/10.1021/es048468f (2005).
Wright, M. S., Peltier, G. L., Stepanauskas, R. & McArthur, J. V. Bacterial tolerances to metals and antibiotics in metal-contaminated and reference streams. FEMS Microbiology Ecology 58, 293–302, https://doi.org/10.1111/j.1574-6941.2006.00154.x (2006).
Berg, J. et al. Cu exposure under field conditions coselects for antibiotic resistance as determined by a novel cultivation-independent bacterial community tolerance assay. Environ Sci Technol 44, 8724–8728, https://doi.org/10.1021/es101798r (2010).
Venkateswaran, K., La Duc, M. T. & Vaishampayan, P. Vol. 1 and 2 1–117 (Jet Propulsion Laboratory, California Institute of Technology, Pasadena, CA., 2012).
Yamada, S., Ohashi, E., Agata, N. & Venkateswaran, K. Cloning and nucleotide sequence analysis of gyrB of Bacillus cereus, B. thuringiensis, B. mycoides, and B. anthracis and their application to the detection of B. cereus in rice. Appl. Environ. Microbiol. 65, 1483–1490 (1999).
Yamamoto, S. & Harayama, S. PCR amplification and direct sequencing of gyrB genes with universal primers and their application to the detection and taxonomic analysis of Pseudomonas putida strains. Appl. Environ. Microbiol. 61, 1104–1109 (1995).
Kim, O.-S. et al. Introducing EzTaxon-e: a prokaryotic 16S rRNA gene sequence database with phylotypes that represent uncultured species. Int. J. Syst. Evol. Microbiol. 62, 716–721 (2012).
Checinska Sielaff, A. et al. Draft Genome Sequences of Biosafety Level 2 Opportunistic Pathogens Isolated from the Environmental Surfaces of the International Space Station. Genome announcements 4, https://doi.org/10.1128/genomeA.01263-16 (2016).
Delcher, A. L., Bratke, K. A., Powers, E. C. & Salzberg, S. L. Identifying bacterial genes and endosymbiont DNA with Glimmer. Bioinformatics (Oxford, England) 23, 673–679, https://doi.org/10.1093/bioinformatics/btm009 (2007).
McArthur, A. G. et al. The comprehensive antibiotic resistance database. Antimicrobial agents and chemotherapy 57, 3348–3357, https://doi.org/10.1128/aac.00419-13 (2013).
Zankari, E. et al. Identification of acquired antimicrobial resistance genes. The Journal of antimicrobial chemotherapy 67, 2640–2644, https://doi.org/10.1093/jac/dks261 (2012).
Gupta, S. K. et al. ARG-ANNOT, a new bioinformatic tool to discover antibiotic resistance genes in bacterial genomes. Antimicrobial agents and chemotherapy 58, 212–220, https://doi.org/10.1128/aac.01310-13 (2014).
Gardner, S. N. et al. Targeted amplification for enhanced detection of biothreat agents by next-generation sequencing. BMC research notes 8, 682, https://doi.org/10.1186/s13104-015-1530-0 (2015).
Gloor, G. B. & Reid, G. Compositional analysis: a valid approach to analyze microbiome high-throughput sequencing data. Canadian journal of microbiology 62, 692–703, https://doi.org/10.1139/cjm-2015-0821 (2016).
McMillan, A. et al. A multi-platform metabolomics approach identifies highly specific biomarkers of bacterial diversity in the vagina of pregnant and non-pregnant women. Scientific reports 5, 14174, https://doi.org/10.1038/srep14174 (2015).
Patel, R. K. & Jain, M. NGS QC Toolkit: a toolkit for quality control of next generation sequencing data. PloS one 7, e30619, https://doi.org/10.1371/journal.pone.0030619 (2012).
Huson, D. H. et al. MEGAN Community Edition - Interactive Exploration and Analysis of Large-Scale Microbiome Sequencing Data. PLoS Comput Biol 12, e1004957, https://doi.org/10.1371/journal.pcbi.1004957 (2016).
Overbeek, R. et al. The SEED and the Rapid Annotation of microbial genomes using Subsystems Technology (RAST). Nucleic Acids Res 42, D206–214, https://doi.org/10.1093/nar/gkt1226 (2014).
Acknowledgements
Part of the research described in this publication was carried out at the Jet Propulsion Laboratory, California Institute of Technology, under a contract with NASA. We would like to thank astronauts Captain Terry Virts and J. Williams for collecting samples aboard the ISS, and the Implementation Team at NASA Ames Research Center including project scientist Dr. Fathi Karouia for coordinating this effort. The authors wish to thank Gary Vora, PhD for his intellectual contributions to the design of the AMR panel v. 1.0., Adriana Blachowicz for helping in processing the samples at Jet Propulsion Laboratory, Ganesh Malli Mohan for disc diffusion antibiotic testing of 3rd flight isolates and Dr. Douglas Botkin for coordinating the arrival of first sample set. © 2017 California Institute of Technology. Government sponsorship acknowledged. This research was funded by a 2012 Space Biology NNH12ZTT001N grant no. 19-12829-26 under Task Order NNN13D111T award to KV which also funded post-doctoral fellowship for ACS, NS, and NASA Space Biology program sponsored NPP post-doc for CU. This work was performed under the auspices of the U.S. Department of Energy by Lawrence Livermore National Laboratory under Contract DE-AC52-07NA27344. In addition, the research carried out at Lawrence Livermore National Laboratory was funded by a 2014 Space Biology NNH14ZTT002N grant no. NNX15AJ29G award to CJ which also funded JEA.
Author information
Authors and Affiliations
Contributions
C.U. wrote the manuscript, analyzed the Ion AmpliSeq™ dataset and reviewed the results for all analyses performed. A.C. was involved in study design, responsible for sample processing, performed the disc diffusion assay and provided input for all analyses performed. K.F. performed targeted sequencing with AmpliSeq™ and helped analyze the AmpliSeq™ dataset. J.A. analyzed the antibiotic resistance gene content of the Staphylococcus isolates and performed antibiotic resistance gene analysis from the WGS data. N.S. conducted metagenomics analysis. K.W. performed the VITEK analysis. K.V. led the project and was involved in study setup and upkeep and coordinating sample collection and processing. K.V., C.J. and K.F. were involved in early organization, study design and planning of the research project, providing direct feedback to C.U., A.C., N.S. and J.A. throughout the project and during write-up of the manuscript. All authors read and edited the manuscript.
Corresponding author
Ethics declarations
Competing Interests
The authors declare that they have no competing interests.
Additional information
Publisher's note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Urbaniak, C., Sielaff, A.C., Frey, K.G. et al. Detection of antimicrobial resistance genes associated with the International Space Station environmental surfaces. Sci Rep 8, 814 (2018). https://doi.org/10.1038/s41598-017-18506-4
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-017-18506-4
This article is cited by
-
Genomic, functional, and metabolic enhancements in multidrug-resistant Enterobacter bugandensis facilitating its persistence and succession in the International Space Station
Microbiome (2024)
-
Simulated microgravity facilitates stomatal ingression by Salmonella in lettuce and suppresses a biocontrol agent
Scientific Reports (2024)
-
D-Mannose prevents bone loss under weightlessness
Journal of Translational Medicine (2023)
-
Microgravity and evasion of plant innate immunity by human bacterial pathogens
npj Microgravity (2023)
-
Microbial applications for sustainable space exploration beyond low Earth orbit
npj Microgravity (2023)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
