
Abstract
Fibromyalgia (FM) results in pain characterized by low selenium (Se) levels, excessive Ca2+ influx, reactive oxygen species (ROS) production, and acidic pH. TRPM2 and TRPV1 are activated by ROS and acid; nevertheless, their roles have not been elucidated in FM. Therefore, we investigated the contribution of TRPM2 and TRPV1 to pain, oxidative stress, and apoptosis in a rat model of FM and the therapeutic potential of Se. Thirty-six rats were divided into four groups: control, Se, FM, and FM + Se. The Se treatment reduced the FM-induced increase in TRPM2 and TRPV1 currents, pain intensity, intracellular free Ca2+, ROS, and mitochondrial membrane depolarization in the sciatic (SciN) and dorsal root ganglion (DRGN) neurons. Furthermore, Se treatment attenuated the FM-induced decrease in cell viability in the DRGN and SciN, glutathione peroxidase, and reduced glutathione and α-tocopherol values in the DRGN, SciN, brain, muscle, and plasma; however, lipid peroxidation levels were decreased. Se also attenuated PARP1, caspase 3, and 9 expressions in the SciN, DRGN, and muscle. In conclusion, Se treatment decreased the FM-induced increase in hyperalgesia, ROS, apoptosis, and Ca2+ entry through TRPM2 and TRPV1 in the SciN and DRGN. Our findings may be relevant to the elucidation and treatment of FM.
Similar content being viewed by others

Introduction
Fibromyalgia (FM) is a common chronic pain syndrome affecting up to 4 million adults in the United States, about 2% of the adult population1. Various factors such as oxidative stress and calcium ion (Ca2+) influx overload play major roles in the etiology of FM. Several pharmaceutical drugs such as antidepressants and voltage-gated calcium channel blockers are recommended for the treatment of FM; however, they fail to produce a satisfactory response in patients with FM because of the unclear etiology of the disease2,3. Therefore, it is highly important to elucidate the etiology of FM and develop more effective therapeutic strategies. The injection of acidic solutions into the gastrocnemius muscle of rats can produce a bilateral long-lasting hyperalgesia similar to FM in humans4.
The activation of acid-sensitive cation channels may contribute to pain induction in FM5,6. Indeed, acidosis from lactate accumulation and oxidative stress from the ischemia of muscles and tender points are two common triggers of pain in patients with FM6,7. Therefore, FM is strongly associated with acid-sensing and oxidative stress-sensing cation channels. Transient receptor potential (TRP) melastatin 2 (TRPM2) and vanilloid 1 (TRPV1) are two members of the Ca2+-permeable TRP superfamily. The TRPV1 channel is activated by different stimuli, including a pungent hot chili pepper component (capsaicin); on the other hand, the TRPM2 channel is activated by ADP-ribose (ADPR)8,9,10. In addition, the channel activities of TRPM2 and TRPV1 are affected by oxidative stress (independent of ADPR)11,12 and low pH (<5.9)12,13. The dorsal root ganglion neuron (DRGN) and sciatic nerve neuron (SciN) are important neurons for the induction of peripheral pain and FM6,14. The TRPM2 and TRPV1 channels are mainly expressed in the DRGN and SciN8,10. Hence, they are also involved in pain signaling and may be altered in chronic pain development. The results of our recent studies indicated that the TRPM2 and TRPV1 channels are activated in the DRGN and SciN by oxidative stress15,16. Therefore, oxidative stress and the acid-dependent activation of the TRPM2 and TRPV1 channels may be involved in the etiology of FM in the FM rat model.
It has been suggested that the mitochondria and oxidative stress may be important factors in the pathogenesis of FM2,17,18,19. However, there are also conflicting studies on mitochondrial oxidative stress in patients20. The essential trace element selenium (Se) acts as a regulator of the physiological functions of the nervous system, such as signal transduction and development21. Se also acts as a cofactor of the glutathione peroxidase (GSH-Px) enzyme and is incorporated into selenoproteins involved in antioxidant defenses22. Furthermore, Se is implicated as a neuroprotective agent in peripheral pain through the inhibition of apoptosis and regulation of the TRPM2 and TRPV1 channels15,23,24. Studies have reported a decreased blood Se level in patients with FM25,26. Furthermore, GSH-Px activity has been found to be drastically reduced in patients with FM17,18,19,27. Se may modulate Ca2+ influx via the TRPM2 and TRPV1 channels; thus, it may affect oxidative stress and apoptosis in the DRGN and SciN of rats with FM. This effect should be clarified in the DRGN and SciN of FM-induced rats.
The mechanism underlying DRGN and SciN injury through FM induction is still poorly understood. To our knowledge, there is no report of the effect of Se on apoptosis, oxidative stress, and Ca2+ entry in the DRGN and SciN of FM-induced rats. The aim of the current study is to determine the molecular mechanism of the effect of Se on apoptosis, oxidative stress, and Ca2+ entry through TRPV1 and TRPM2 regulation in the DRGN and SciN after FM induction.
Results
Effects of Se on mechanical hyperalgesia in FM-induced rats
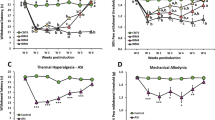
Five days after the second injection of acidic saline into the left gastrocnemius muscle, there was a significant decrease in the mechanical withdrawal threshold of the paw bilaterally, which continued to gradually decrease until the 19th day in all periods analyzed, as demonstrated by hot plate (Fig. 1a) and von Frey (Fig. 1b) hyperalgesia assessments. On the other hand, compared with the control group, acid injections produced mechanical hyper-responsiveness on day 5th and 19th (p ≤ 0.001). However, when only Se was administered, a potent antihyperalgesic effect was observed in the injected paw (p ≤ 0.001). A similar antihyperalgesic effect was also observed in the contralateral paw when Se was administered to rats with FM. However, pretreatment with Se caused a marked increase in the sensitivity threshold to mechanical stimuli (p ≤ 0.001).

Effect of selenium (Se) treatment on paw withdraw threshold of Hot Plate Test (a) and paw withdrawal force of Von Frey test (b) in rats with FM. (n = 8–10 and mean ± SD). (a p ≤ 0.001 versus basal groups. b p ≤ 0.001 5th day groups. c p ≤ 0.001 versus control group of 19th day group. d p ≤ 0.001 versus Se group of 19th day group. e p ≤ 0.001 versus FM group of 19th day group).
Effects of Se on [Ca2+]i concentration through modulation of TRPV1 and TRPM2 activation in the DRGN and SciN of FM-induced rats
To clarify the effect of Se on the TRPV1 and TRPM2 channels in the neurons, the neurons of Se-administrated rats were further gated by capsaicin (CAPS; 10 μM) and CumHPx (1 mM). Stimulations with CAPS and CumHPx caused a significant increase in [Ca2+]i influx in the DRGN and SciN of FM-induced rats, which was attributed to the activation of Ca2+-permeable TRPV1 (Fig. 2a and b) and TRPM2 (Fig. 3a and b). As shown in Figs 2c and 3c, despite the higher concentration of [Ca2+]i in the FM groups than in the control group, the TRPV1 antagonist (CPZ) and TRPM2 antagonist (ACA) could effectively decrease the concentration of [Ca2+]I, which was increased by experimental FM induction (p ≤ 0.001). The [Ca2+]i concentration in the neurons was significantly (p ≤ 0.001) lower in the control + CPZ, control + ACA, FM + CPZ, and FM + ACA groups (p ≤ 0.001) than in the FM group. In addition, we found that the concentration of [Ca2+]i was low in the neurons of the Se-treated group. The [Ca2+]i concentration in the neurons was significantly lower in the FM + Se, FM + CPZ, and FM + ACA groups compared with the FM group (p ≤ 0.001). Therefore, Se could modulate the FM-induced [Ca2+]i concentration by regulating TRPV1 and TRPM2 in the neurons.

Effect of selenium (Se) treatment on [Ca2+]i concentration through block of TRPV1 gate in dorsal root ganglion neuron (DRGN) (a) and sciatic nerve (SciN) (b) of control and FM-induced rats. (n = 8–10 and mean ± SD). The animals received intraperitoneal Se for 2 weeks after FM induction. Then, these dissected neurons of control and FM groups were further in vitro treated with CAPS (10 μM) and CPZ (0.1 mM) before loading Fura-2 for 120 seconds. (a p ≤ 0.05 and b p ≤ 0.001 versus control. c p ≤ 0.001 and d p ≤ 0.05 versus control + CPZ and Se groups. e p ≤ 0.001 versus FM group (2c).

Effect of selenium (Se) treatment on [Ca2+]i concentration through block of TRPM2 gate in dorsal root ganglion neuron (DRGN) (a) and sciatic nerve (SciN) (b) of control and FM-induced rats. (n = 8 and mean ± SD). The animals received intraperitoneal Se for 2 weeks after FM induction. Then, these dissected neurons of control and FM groups were further in vitro treated with CumHPx (1 mM) and ACA (0.025 mM) before loading Fura-2 for 120 seconds. (a p ≤ 0.05 and b p ≤ 0.001 versus control. (c p ≤ 0.001 and d p ≤ 0.05 versus control + ACA and Se groups. e p ≤ 0.001 versus FM group (3c).
Effects of Se on CAPS-induced TRPV1 and ADPR-induced TRPM2 currents in the DRGN of control and FM-induced rats
The murine DRGNs in TRPV1 (Fig. 4b and c) and TRPM2 (Fig. 5b and c) experiments were gated by CAPS. CAPS- and ADPR-induced currents were reversibly blocked by CPZ, ACA, and NMDG+ (replacement of Na+), respectively. There were no currents in the absence of the agonists (CAPS and ADPR) (Figs 4a and 5a). The current densities in the neurons were significantly higher in the FM + CAPS and FM + ADPR groups compared with the control, control + CAPS, and control + ADPR groups (p ≤ 0.001); however, the current densities were significantly (p ≤ 0.001) lower in the control + CAPS + CPZ, control + ADPR + ACA, FM + CAPS + CPZ, and FM + ADPR + ACA groups than in the FM group (Figs 4f and 5f). The current densities in the neurons were decreased by Se treatments, and they were low in the Se + CAPS, Se + ADPR, FM + Se + CAPS, and FM + Se + ADPR groups (p ≤ 0.001). These results clearly indicated that CAPS and ADPR induced Ca2+ entry overload through the TRPV1 and TRPM2 channels. However, the FM-induced TRPV1 and TRPM2 currents through oxidative stress modulation were decreased by treatment with the antioxidant Se.

Effects of selenium (Se) on TRPV1 channel activation in dorsal root ganglion neuron (DRGN) of control and FM-induced rat. The TRPV1 currents in DRGN were stimulated by extracellular CAPS (10 μM in patch chamber) but they were blocked by extracellular CPZ (0.1 mM) in the patch-chamber. W.C. is whole cell. (a) Control: Original recordings from control neuron. (b) Control + CAPS group (without FM induction). (c) FM group (with FM induction). (d) FM + Se group: Se was administrated to the rats after FM induction. (e) Se group: Se was administrated to the rats without FM induction. (f) TRPV1 channel current densities in the DRGN. The numbers in parentheses indicated n numbers of groups. (a p ≤ 0.001 versus control. b p ≤ 0.001 versus control + CAPS group. c p ≤ 0.001 versus control + CAPS + CPZ group. d p ≤ 0.001 versus FM + CAPS group. e p ≤ 0.001 versus FM + CAPS + CPZ group).

Effects of selenium (Se) on TRPM2 channel activation in dorsal root ganglion neuron (DRGN) of fibromyalgia (FM)-induced rat. The TRPM2 currents in DRGN were stimulated by intracellular ADPR (1 mM in patch pipette) but they were blocked by extracellular TRPM2 antagonist (ACA and 0.025 mM) in the patch-chamber. W.C. is whole cell. Control (without FM induction and stimulation): Original recordings from control neuron. (b) Control + ADPR group (without FM induction). (c) FM group (with FM induction). (d) FM + Se group: The rats received Se after FM induction. (e) Se group: The rats received Se without FM induction. (f) TRPM2 channel current densities in the DRGN. The numbers in parentheses indicated n numbers of groups were indicated by numbers in parentheses. (a p ≤ 0.001 versus control. b p ≤ 0.001 versus control + ADPR group. c p ≤ 0.001 versus control + ADPR + ACA group. d p ≤ 0.001 versus FM + ADPR group. e p ≤ 0.001 versus FM + ADPR + ACA group).
Effect of Se on apoptosis, cell viability (MTT), caspase activity, intracellular ROS production, and JC-1 level in the SciN and DRGN of control and FM groups
Apoptosis levels (6a and b), caspase 3 and caspase 9 activities (6c and d), and JC-1 and ROS levels (6e and f) were markedly increased (p ≤ 0.001) following TRPV1 and TRPM2 stimulation in the DRGN (Fig. 6) and SciN (data are not shown) by FM induction; however, MTT levels were markedly decreased (p ≤ 0.001) in the neurons. The pre-treatment of cells with CPZ (without Se) followed by CAPS stimulation reversed CAPS- and CumHPx-induced oxidative cytotoxicity (apoptosis, MTT, JC1-1, ROS, caspase 3, and caspase 9 values), and the CPZ and ACA treatment resulted in significant additive effects with a 20–50% increase in cell viability (Fig. 6a and b). In addition, apoptosis, JC-1, and ROS levels and caspase 3 and caspase 9 activities were markedly decreased (p ≤ 0.05 and p ≤ 0.001, respectively) in the neurons of the FM groups following the blockage of the TRPV1 and TRPM2 channels by Se with/without CPZ and ACA treatments; however, MTT levels were markedly increased (p ≤ 0.05 and p ≤ 0.001, respectively) in the FM groups by Se with/without CPZ and ACA treatments. These results implied that TRPV1 and TRPV1 cytotoxicity may be attributed to TRPV1 and TRPM2 activation.

Effects of selenium on the apoptosis, cell viability (MTT), mitochondrial membrane depolarization (JC-1), intracellular ROS production and caspase 3 and 9 values through TRPV1 (a,c and e) and TRPM2 (b,d and f) channel activations in the DRGN of FM-induced rats (mean ± SD and n = 3). Values expressed as fold increase (experimental/control). These neurons were dissected from control, FM and treated animals. The TRPV1 and TRPM2 gates in the neurons were opened with capsaicin (10 μM) and cumene hydroperoxide (1 mM) although they were blocked by CPZ (0.1 mM) and ACA (0.025 mM), respectively. (a p ≤ 0.05 and b p ≤ 0.001 versus control and control + CPZ groups. c p ≤ 0.05 versus Se group. d p ≤ 0.001 versus Se and Se + CPZ groups. e p ≤ 0.001 versus FM and FM + CPZ groups. f p ≤ 0.05 versus FM + Se groups).
Effect of Se on caspase 3, caspase 9, and PARP1 expression level in the DRGN, SciN, and muscle of FM-induced rats
PARP1 activity plays an essential role in DNA damage, and the TRPM2 channel agonist ADPR is produced in the nucleus by stimulation of oxidative stress. In apoptotic pathways, caspase 8 activates a proteolytic caspase cascade that transmits and amplifies death signals by the activation of apoptotic caspases such as executioner caspase 3 and initiator 9. These initiator and executioner caspases cleave several substrate proteins including PARP1, resulting in the self-destruction of the cells28. Caspase 3 and caspase 9 expression levels are associated with the progress of apoptosis in neuronal injuries15,24. In the current study, the expression levels of caspase 3, caspase 9, and PARP1 in the DRGN (Fig. 7a and d), SciN (Fig. 7b and d), and muscle (Fig. 7c and d) were markedly higher (p ≤ 0.001) in FM group than in the control group. However, the expression levels of caspase and PARP1 in the three samples were decreased by FM + Se treatments, and their expression levels in the SciN (p ≤ 0.05), DRGN (p ≤ 0.05), and muscle (p ≤ 0.001) were significantly lower in the FM + Se group than in the FM group. In addition, the expression levels of caspase and PARP1 in the three samples were further decreased by Se treatments. These results demonstrated of the roles of caspase 3 and 9 in the apoptotic pathway activated by FM; these caspases could trigger a proteolytic cascade that amplifies death signals by inducing DNA damage, executioner caspase 3, and initiator 9.

Effects of selenium on the caspase 3, caspase 9 and PARP1 expression levels in DRGN (a and d), SciN (b and d) and gastrocnemius muscle (c and d) of rats with FM (mean ± SD and n = 3). (a p ≤ 0.05 and bp ≤ 0.001 versus control. c p ≤ 0.05 and d p ≤ 0.001 versus FM group. e p ≤ 0.001 versus FM + Se group).
Effects of Se on MDA, GSH, GSH-Px, and antioxidant vitamin concentration
Decreased blood GSH, GSH-Px, and antioxidant vitamin levels were reported in patients with FM17,18,19. On the other hand, an increase in their levels17 and a decrease in muscle pain intensity29 were reported in patients with FM following α-tocopherol supplementation. Furthermore, GSH-Px activity has been found to be drastically reduced in patients with FM17. Therefore, we measured MDA concentration, GSH concentration, and GSH-Px activity in the muscle, brain homogenate, and hemolyzed erythrocytes of rats with FM to clarify the role of oxidative stress in the etiology of FM (Table 1).
GSH level and GSH-Px activity in the DRGN, brain, muscle, erythrocyte, and SciN and α-tocopherol concentration in the brain, muscle, and plasma were significantly decreased (p ≤ 0.05 and p ≤ 0.001, respectively) by the induction of FM; however, MDA levels in the samples were markedly decreased (p ≤ 0.05 and p ≤ 0.001, respectively) (Table 1). On the other hand, GSH-Px activity and GSH level (in the DRGN, brain, muscle and SciN) and α-tocopherol concentration (in the brain, muscle, and plasma) were increased in the FM group following Se treatment (p ≤ 0.05 and p ≤ 0.001, respectively). There was no change in retinol and β-carotene concentration in the four groups (Table 1).
Discussion
The results of the current study indicated that Se treatment might decrease FM-induced pain intensity, [Ca2+]i accumulation, mitochondrial ROS, and apoptosis levels in the DRGN and SciN by blocking the TRPM2 and TRPV1 channels. To our knowledge, this is the first evidence demonstrating the FM pathophysiological process and implicating the DRGN and SciN in central and peripheral pain diseases.
TRPV1 is a pronociceptive polymodal receptor that senses CAPS, oxidative stress, high temperature, and acidic pH. Oxidative stress and acidic pH play significant roles in the activation of TRPM2 and TRPV1,12,13. The activation of TRPM2 and TRPV1 through oxidative stress and acidic pH could enhance [Ca2+]i accumulation, and they are involved in several physiological and pathological processes such as neuronal apoptosis and neuronal recovering signaling5,15,30. Therefore, TRPM2 and TRPV1 could act as the final oxidative stress mediators that function via distinct intracellular signaling pathways such as NADPH oxidase31. The blockage of TRPM2 and TRPV1 generally results in antinociceptive effects in animal models; however, the roles of TRPM2 and TRPV1 in maintaining acid- and oxidative stress-induced chronic hyperalgesia in animal models have not been elucidated. Recent studies have revealed that TRPV1 may play a role in the development of hyperalgesia after the induction of acid-induced FM5. Se as a cofactor of the cytosolic antioxidant enzyme GSH-Px is a strong antioxidant21,22, and it has strong superoxide radical scavenger and NADPH oxidase blocker effects in several types of cells5,32. In recent studies, we observed that Se treatment decreased the levels of Ca2+ influx by blocking TRPM2 and TRPV1 in the DRGN and SciN of rats15,30. To our knowledge, there is no report on calcium signaling and pain intensity in relation to the activation of TRPM2 and TRPV1 in the DRGN and SciN of rats with FM. In the current study, we observed that pain intensity and Ca2+ influx through the TRPM2 and TRPV1 channels were increased in the DRGN and SciN of FM-induced rats; however, pain intensity and Ca2+ influx were decreased in the neurons following treatment with the antioxidant Se. Similar to the pain intensity and Ca2+ results of the current study, decreased chronic hyperalgesia has been demonstrated by blocking CaV3.2 T-type Ca2+ channel signaling in mice at 15 min before the second acid injection in another study33. In addition, TRPV1 overexpression in the development of a FM-like pain state has been reported in an acid-injected TRPV1-null mice model of FM34.
The increased activation of cation channels including TRPM2 and TRPV1 leads to a high [Ca2+]i concentration, which disrupts the Ca2+ contents of the intermembrane space through mitochondrial permeability transition activation35. The dysfunction of the mitochondria activates three pathways: (1) generation of endogenous mitochondrial ROS, (2) apoptosis through caspase production, and (3) DNA damage through PARP1 activation34. The role of TRPM2 and TRPV1 in mitochondrial dysfunction and apoptosis in the DRGN and SciN has been reported in recent studies15,34. In the current study, apoptosis, caspase 3, caspase 9, PARP1, JC-1, and intracellular ROS values were increased in the SciN and DRGN following FM induction; however, their values were decreased in the neurons by blocking TRPM2 and TRPV1 with Se.
The antagonist effects of Se on the TRPM2 and TRPV1 channels in the DRGN were revealed in recent studies15,24. In addition, the involvement of NADPH oxidase and protein kinase C molecular pathways in the activation of TRPM2 and TRPV1 in the DRNG were reported in a recent study31. Besides functioning as an antagonist of TRPM2 and TRPV1, Se also inhibits NADPH oxidase in various cell lines32,33. Based on present data on Se, we hypothesize that Se as a potential antagonist of TRPM2 and TRPV1 may also function via this mechanism. Indeed, we found that treatment with Se resulted in decreased FM-induced mitochondrial ROS in the DRGN and SciN, and FM-induced apoptosis was reversed by Se treatments. Therefore, the current results indicated that Se may modulate FM-induced cellular oxidative stress in the DRGN and SciN of rats with FM. It is well known that patients with FM are under oxidative stress because of the ischemia of tender points and abnormalities in mitochondrial functions, resulting in a delicate balance between blood ROS levels and the antioxidant capabilities of the cell17,18,19,36. Therefore, increases in ROS above the basal level could disrupt this fine balance, thereby triggering ROS-induced apoptosis. This may explain why Se was able to inhibit ROS, apoptosis, caspase 3, and caspase 9 in the DRGN and SciN of rats with FM.
The human body is equipped with a complete arsenal of defenses against ROS. These ROS are scavenged by enzymes, such as GSH-Px and catalase, and non-enzymatic species, such as GSH and vitamin E37. α-Tocopherol is the major scavenger of ROS in the lipid phase of neuronal membranes, and its oxidized form is recovered by the antioxidant action of GSH. An imbalance between the oxidant and antioxidant system is a common feature in the clinical dysfunction of the nervous system. It is well known that an increase in mitochondrial ROS and a decrease in GSH-Px activity and Se level are known to be implicated in the etiology of pain and FM17,18,19,25,26,27. In addition to acting as a cofactor of GSH-Px, Se is an important antioxidant enzyme in the brain and neurons for removing lipid hydroperoxides and hydrogen peroxide21. In the current study, we observed that GSH-Px activity, GSH concentration, and α-tocopherol concentration were reduced in the DRGN, SciN, brain, muscle, and plasma; however, they were increased to near basal levels following Se treatments. These results demonstrated that Se administration conferred protection against FM in the rats, possibly by limiting the lifetime of ROS. Retinol and β-carotene concentration in the brain and muscle did not change in the four groups; however, they also have antioxidant roles in neurons. The adaptive antioxidant responses of retinol and β-carotene were accompanied by GSH-Px activity, GSH concentration, and α-tocopherol concentration upregulation.
In summary, our data suggested for the first time that the mechanisms of FM may be mediated via the TRPM2 and TRPV1 channels by inducing mitochondrial ROS, apoptosis, and pain in the DRGN and SciN. However, the pain, oxidant, and apoptotic effects of FM were reversed through the blockage of TRPM2 and TRPV1 in the neurons by Se treatment. Therefore, the use of Se may be an effective novel approach for treating FM-induced pain, mitochondrial oxidative stress, and apoptosis. In addition, the TRPM2 and TRPV1 channels may be important pharmacological targets in the treatment of FM-induced apoptosis and pain.
Methods
Animals
It is well known that incidence rate of FM is high in women1,2,3. Hence, we used 36 female Wistar rats (aged 12 weeks old and 170 ± 10 g body weight) in the current study. The rats were housed in controlled-temperature (21 ± 2 °C) and humidity (65%) rooms, under a 12:12 h light-dark cycle, with ad libitum access to water and food until use. Hot plate and Von Frey tests were performed between 9:00 and 10.00 a.m. at baseline, 5th and 19th days of the experiments.
Availability of data and materials
Data and approve of rats were taken from Experimental Animal Research Center of Suleyman Demirel University (SDU) according to protocol number (HADYEK-21438139-320). All methods in the manuscript were performed in accordance with the relevant guidelines and regulations of SDU by including a statement in the methods section to this effect. The dataset and analyses were generated in Neuroscience Research Center of SDU and they are available from the corresponding author on reasonable request. Graphics and tables in the manuscript were prepared by the corresponding author.
Study groups
The rats were divided into four groups as follows: The control group (n = 8) had no FM and was not administrated. They received intraperitoneal (i.p.) 0.9% w/v saline solution for 2 weeks. In the Se group (n = 8), they received i.p. Se 1.5 mg/kg/over day for two weeks (total seven doses)15. In FM group (n = 10), the rats were exposed FM induction procedure4. Then, the rats received i.p. saline solution for 2 weeks. In FM + Se group (n = 10), the rats received Se (same as the Se group) after FM induction (same as the FM group).
Twelve hours after the last saline solution and Se dose administration, all rats were decapitated under propofol inhalation anesthesia in accordance with SDU experimental animal legislation. Selenium (Sodium selenite, Sigma Chemical Co., MO, USA) was diluted to appropriate concentration in sterile 0.9% w/v saline solution. In patch-clamp experiment and [Ca2+]i concentration assays, the DRGN and SciN were further treated with CumHPx (1 mM) or ADPR (1 mM) and capsaicin (10 μM) for activation of TRPM2 and TRPV1 channels, respectively and they were also blocked the TRPM2 channels antagonist, N-(p-amylcinnamoyl) anthranilic acid (ACA and 0.025 mM) and TRPV1 antagonist, capsazepine (CPZ and 0.1 mM). Doses of capsaicin and CPZ on TRPV1 in cells are changing between 100 nM and 0.1 mM. In a recent study31, we tested different doses of capsaicin and CPZ in the DRGN and TRPV1 against and antagonist doses of capsaicin and CPZ in the study was found as 10 μM and 0.1 mM, respectively. Therefore, the doses were used in the current study.
Induction of hyperalgesia
For produce a bilateral long-lasting hyperalgesia similar to fibromyalgia in humans, acidic saline solution (adjusted to pH 4.0) was injected into the gastrocnemius muscle in the rats4. Before the first injection of acidic or normal saline into the gastrocnemius muscle, paw withdrawal threshold values of the rats were recorded in order to record the baseline value. Each animal received two repeated injections of the acidic saline solution (100 μl) in the same unilateral gastrocnemius muscle under propofol inhaled anesthesia, a procedure which is repeated five days later4. After the second injection of the acidic saline, hyperalgesia in the animal is detected increase the paw withdrawal threshold by the von Frey filament and hot plate tests in the muscle.
Hyperalgesia tests
The assessment of hyperalgesia was measured by the using calibrated von Frey filaments (20PC Aesthe Model, NO. 160615, Muromachi Kikai Co., Ltd. Tokyo, Japan), to the plantar aspect of the hind paw of mice that were kept in suspended wire mesh cages. A response was indicated by lifting of the hind paw38.
Heat-controlled plate (Varioma, Thermo Fischer Inc., Langenselbold, Germany) for Hot Plate test was used to assess paw withdrawal latency to thermal nociceptive stimuli. The hot plate test apparatus consisted of an electrically heated surface kept at a constant temperature of 55.0 ± 0.6 °C. Each mouse was placed on hot plate, and the reaction time was measured until the mouse either demonstrated hind paw licking or jumping (‘up and down’ method). A cutoff time of 60 s was used to prevent tissue damage for hot plate test38. Withdrawal threshold was determined by sequentially increasing and decreasing the stimulus strength.
Preparation of brain, blood, primary DRGN and SciN samples
Details of isolations of DRGN and SciN were given in previous studies15,34. Briefly, the DRGN and SciN were minced with iridectomy scissors and incubated with enzymes including trysin (type III, Sigma) and 0.5 mg/ml collagenase (type XI, Sigma) in 5 ml DMEM at 37 °C in a shaking bath for 40 min after removing the attached nerves and surrounding connective tissues. To stop the enzymatic digestion 1.25 mg/ml soybean trypsin inhibitor (type II-S1, Sigma) was added. After dissociation with a sterile syringe, the DRGN and SciN suspensions of mediums were centrifuged at 1,500 g and the medium and high size neurons were removed for the analysis. The isolated neurons were transferred into a 35-mm culture dish and kept still for at least 30 min.
The brain was also taken as follows; the cortex was dissected out after the brain was split in the mid-sagittal plane. The gastrocnemius muscle samples from right legs were also taken. After preparing brain and muscle homogenates in ice-cold Tris-HCl buffer (50 mM, pH 7.4), they were stored at −85 °C for analyses of lipid peroxidation and antioxidants. Half of the muscle sample with DRGN and SciN was frozen at −85 °C for using Western blot analyses. Plasma and erythrocyte samples from anticoagulated blood (sodium EDTA) were obtained as described in a previous studies17,18 and they were stored at −85 °C.
Electrophysiology
Whole-cell voltage clamp recording was taken from the DRGN at 22–24 °C (EPC10 patch-clamp set, HEKA, Lamprecht, Germany). Resistances of the recording electrodes were adjusted to about 3–6 MΩ by a puller (PC-10 Narishige International Limited, London, UK). We used standard extracellular bath and pipette solutions as described in previous studies16,30. Holding potential of the patch-clamp analyses in the DRGNs was −60 mV. Voltage clamp technique was used in the analyses and current-voltage (I–V) relationships were obtained from voltage ramps from −90 to +60 mV applied over 200 milliseconds. All experiments were performed at room temperature (22 ± 1 °C).
In the experiments, TRPM2 is intracellularly (via patch pipette) gated by ADPR (1 mM), although they were extracellularly (via patch chamber) blocked by ACA (0.025 mM). TRPV1 was extracellularly gated by CAPS (10 μM), and the channels were extracellularly blocked by CPZ (0.1 mM). For the analysis, the maximal current amplitudes (pA) in a DRGN were divided by the cell capacitance (pF), a measure of the cell surface. The results in the patch clamp experiments are the current density (pA/pF).
Measurement of [Ca2+]i concentration in DRGN and sciatic nerve
The DGRN and SciN samples were separately seeded in sterile cell culture flasks. Then they were loaded with 4 μM Fura-2/AM in loading buffer for 45 min at 37 °C in the dark, washed twice with the phosphate buffer, incubated for an additional 30 min at 37 °C to complete probe de-esterification. The groups were exposed to the stimulations in a water-jacketed cuvette (37 °C) with continuous magnetic stirring. Fluorescence was detected by using a Carry Eclipse Spectrofluorometer (Varian Inc, Sydney, Australia). The fluorescence at 505 nm was measured at 1 second intervals after excitation at 340 nm and 380 nm, respectively.
For calibration of [Ca2+]i, maximum and minimum fluorescence values were obtained by adding the detergent Triton X-100 (0.1%) and the Ca2+ chelator EGTA (10 mM) sequentially at the end of each experiment. Calculation of the [Ca2+]i concentrations were described in previous studies39,40, assuming a Kd of 155 nM. The [Ca2+]i concentrations in TRPM2 and TRPV1 experiments were recorded by using the integral of the rise in [Ca2+]i for 120 seconds after addition of CumHPx (1 mM) or capsaicin (10 μM), respectively. The [Ca2+]i concentration is expressed as nanomolar (nM) taking a sample every second as previously described39,41.
Measurement of intracellular ROS production and mitochondrial membrane depolarization
Dihydrorhodamine (DHR)123 is an uncharged and nonfluorescent intracellular ROS production indicator and the level of intracellular ROS was assessed by fluorescence microplate reader (Infinite pro200, Tecan Life, Männedorf, Switzerland) with DHR123, which oxidizes to cationic rhodamine 123 in the presence of ROS, as described previously39. The resulting data were normalized using the control values. 5,5′,6,6′-tetrachloro-1,1′,3,3′-tetraethylbenzimidazolocarbocyanine iodide (JC-1) dye was used for measurement of the mitochondrial membrane potential and it was determined in he a plate reader (Infinite pro200) by using JC-1 as described in previous studies40,42. The ROS and JC-1 values were expressed as fold-increase over the pretreatment level after calculating fluorescence units/mg protein.
Assay for apoptosis, cell viability, and caspase 3 and 9 values
The apoptosis levels were determined in a spectrophotometer (UV-1800, Shimadzu, Kyoto, Japan) by using a commercial kit of Biocolor Ltd. (Northern Ireland) as described in a previous study41. The method is based on loss of asymmetry in membranes of apoptotic neurons. We used to cell viability analyses as 3-(4,5-Dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) in the neurons as described elsewhere41 and absorbance values of MTT were recorded by the spectrophotometer (UV-1800) at 490 nm.
The determinations of caspase 3 and caspase 9 activities in the sciatic nerve and DRGN neurons were performed in the microplate reader (Infinite pro200) by using caspase 3 (N-acetyl-Leu-Glu-His-Asp-7-amino-4-methylcoumarin (AC-LEHD-AMC), Bachem, Bubendorf, Switzerland) and caspase 9 (N-acetyl-Asp-Glu-Val-Asp-7-amino-4-methylcoumarin (ACDEVD-AMC), Sigma) substrates. Details of the assays were indicated in recent studies16,34. The apoptosis, MTT, caspase and 9 values were expressed as fold-increase after calculating fluorescence units/mg protein.
Lipid peroxidation, GSH-Px, reduced glutathione (GSH) and antioxidant vitamin analyses in plasma, erythrocytes, muscle (gastrocnemius) and brain
The GSH concentration and GSH-Px activity in the DRGN, SciN, muscle, brain homogenate and hemolyzed erythrocyte were spectrophotometrically (UV-1800) assayed at 412 nm by using the method of Sedlak and Lindsay43 and Lawrence and Burk22, respectively. Lipid peroxidation levels as malondialdehyde (MDA) in the DRGN, SciN, muscle, brain homogenate, plasma and hemolyzed erythrocyte were measured with the thiobarbituric-acid reaction by using method of Placer et al.44. The GSH-Px activity, GSH and MDA concentrations were expressed as international unit (IU) of GSH oxidized/min/μg protein (IU/μg pr) and μmol/g protein (μmol/μg pr), respectively. Method of Bradford was used in the muscle and brain homogenate, and hemolyzed erythrocytes for the determination of protein contents.
Results of recent studies indicated low vitamin A (retinol), vitamin E (α-tocopherol) and β-carotene concentrations in plasma of patients with FM. For further clarifying role of antioxidant vitamins in FM-induced oxidative stress, retinol, β-carotene and α-tocopherol were spectrophotometrically determined in the brain, gastrocnemius muscle and plasma samples by a modification of the method described by Suzuki and Katoh45 and Desai46. Calibration was performed using standard solutions of all-trans retinol, β-carotene and α-tocopherol in hexane. All antioxidant vitamin values in plasma, muscle and brain were expressed as μmol/l for plasma and μmol/g tissue for brain and muscle, respectively.
Western Blot analyses
Standard procedures are used in the Western Blot analyses of gastrocnemius muscle, SciN and DRGN16,34. Samples were homogenized in ice-cold RIPA buffer. The protein concentration in the supernatant was determined using the Bradford’s method. Membrane blots were incubated overnight at 4 °C with the following primary antibodies: caspase 3 (p17-specific Polyclonal Antibody), caspase 9 (p35/p10 Polyclonal Antibody), β actin (polyclonal antibody), poly-ADPR polymerase 1 (PARP1) (polyclonal antibody). The primary antibodies were purchased from Proteintech (Istanbul, Turkey) although secondary antibodies (Rabbit IgG, HRP-linked from donkey) were purchased from GE Healthcare (Amersham, UK). Relative levels of immunoreactivity in ECL Western HRP Substrate (Millipore Luminate Forte, USA) were quantified using Syngene G:Box Gel Imagination System (UK). Rabbit anti-β-actin (1:2000) was used as an internal control for the concentration of proteins loaded. Obtained values were expressed as relative density over the control level.
Statistical analyses
All data were represented as means ± standard deviation (SD). The data were analyzed by using 17.0 version of SPSS statistical program (Chicago, Illinois, USA). P value as ≤ 0.05 was considered to indicate a statistically significant difference. Presence of significance in the four groups was once detected by LSD test. Withdrawal thresholds of Hot Plate and Von Frey were analyzed by using a Dixon non-parametric test. Remaining p value levels of significances in the data were analyzed by using Mann-Whitney U test.
Compliance with Ethical Standards
The study was approved by the Local Experimental Animal Ethical Committee of Suleyman Demirel University (SDU) (Protocol number: HADYEK-21438139-320).
References
Fibromyalgia fact sheet. Centers for Disease Control and Prevention. CDC 24/7. https://www.cdc.gov/arthritis/basics/fibromyalgia.htm (2017).
Ozgocmen, S. et al. Antioxidant status, lipid peroxidation and nitric oxide in fibromyalgia: etiologic and therapeutic concerns. Rheumatol Int. 26, 598–603 (2006).
Di Tommaso Morrison, M. C. et al. Fibromyalgia and bipolar disorder: extent of comorbidity and therapeutic implications. J Biol Regul Homeost Agents. 31, 17–20 (2017).
Sluka, K. A., Kalra, A. & Moore, S. A. Unilateral intramuscular injections of acidic saline produce a bilateral, long-lasting hyperalgesia. Muscle Nerve 24, 37–46 (2001).
Chen, W. N. et al. Roles of ASIC3, TRPV1, and NaV1.8 in the transition from acute to chronic pain in a mouse model of fibromyalgia. Mol Pain. 10, 40 (2014).
Ross, J. L. et al. Muscle IL1β Drives Ischemic Myalgia via ASIC3-Mediated Sensory Neuron Sensitization. J Neurosci. 36, 6857–6871 (2016).
DeSouza, R. A. et al. Mitochondrial encephalomyopathy with lactic acidosis and strokelike episodes (MELAS): a mitochondrial disorder presents as fibromyalgia. South Med J. 97, 528–531 (2004).
Nazıroğlu, M. TRPM2 cation channels, oxidative stress and neurological diseases: where are we now? Neurochem Res. 36, 355–366 (2011).
Pecze, L., Blum, W. & Schwaller, B. Mechanism of capsaicin receptor TRPV1-mediated toxicity in pain-sensing neurons focusing on the effects of Na(+)/Ca(2+) fluxes and the Ca(2+)-binding protein calretinin. Biochim Biophys Acta. 1833, 1680–1691 (2013).
Tominaga, M. & Tominaga, T. Structure and function of TRPV1. Pflugers Arch. 451, 143–150 (2005).
Nazıroğlu, M. & Lückhoff, A. A calcium influx pathway regulated separately by oxidative stress and ADP-Ribose in TRPM2 channels: single channel events. Neurochem Res. 33, 1256–1262 (2008).
Du, J., Xie, J. & Yue, L. Modulation of TRPM2 by acidic pH and the underlying mechanisms for pH sensitivity. J Gen Physiol. 134, 471–488 (2009).
Jordt, S. E., Tominaga, M. & Julius, D. Acid potentiation of the capsaicin receptor determined by a key extracellular site. Proc Natl Acad Sci USA 97, 8134–8139 (2000).
Goffaux, P., de Souza, J. B., Potvin, S. & Marchand, S. Pain relief through expectation supersedes descending inhibitory deficits in fibromyalgia patients. Pain 145, 18–23 (2009).
Kahya, M. C., Nazıroğlu, M. & Övey, İ. S. Modulation of diabetes-induced oxidative stress, apoptosis, and Ca(2+) entry through TRPM2 and TRPV1 channels in dorsal root ganglion and hippocampus of diabetic rats by melatonin and selenium. Mol Neurobiol. 54, 2345–2360 (2017).
Akpınar, H., Nazıroğlu, M., Övey, İ. S., Çiğ, B. & Akpınar, O. The neuroprotective action of dexmedetomidine on apoptosis, calcium entry and oxidative stress in cerebral ischemia-induced rats: Contribution of TRPM2 and TRPV1 channels. Sci Rep. 6, 37196 (2016).
Nazıroğlu, M. et al. Vitamins C and E treatment combined with exercise modulates oxidative stress markers in blood of patients with fibromyalgia: a controlled clinical pilot study. Stress 13, 498–505 (2010).
Akkuş, S. et al. Levels of lipid peroxidation, nitric oxide, and antioxidant vitamins in plasma of patients with fibromyalgia. Cell Biochem Funct. 27, 181–185 (2009).
Sánchez-Domínguez, B. et al. Oxidative stress, mitochondrial dysfunction and, inflammation common events in skin of patients with Fibromyalgia. Mitochondrion 21, 69–75 (2015).
Ranzolin, A. et al. Evaluation of cytokines, oxidative stress markers and brain-derived neurotrophic factor in patients with fibromyalgia - A controlled cross-sectional study. Cytokine 84, 25–28 (2016).
Schweizer, U., Bräuer, A. U., Köhrle, J., Nitsch, R. & Savaskan, N. E. Selenium and brain function: a poorly recognized liaison. Brain Res Brain Res Rev. 45, 164–178 (2004).
Lawrence, R. A. & Burk, R. F. Glutathione peroxidase activity in selenium-deficient rat liver. Biochem Biophys Res Commun 71, 952–958 (1976).
Nazıroğlu, M. et al. Selenium modulates oxidative stress-induced TRPM2 cation channel currents in transfected Chinese hamster ovary cells. Basic Clin Pharmacol Toxicol. 112, 96–102 (2013).
Balaban, H., Nazıroğlu, M., Demirci, K. & Övey, İ. S. The Protective Role of selenium on scopolamine-induced memory impairment, oxidative stress, and apoptosis in aged rats: The involvement of TRPM2 and TRPV1 channels. Mol Neurobiol. 54, 2852–2868 (2017).
Reinhard, P., Schweinsberg, F., Wernet, D. & Kötter, I. Selenium status in fibromyalgia. Toxicol Lett. 96–97, 177–180 (1998).
Sendur, O. F., Tastaban, E., Turan, Y. & Ulman, C. The relationship between serum trace element levels and clinical parameters in patients with fibromyalgia. Rheumatol Int. 28, 1117–1121 (2008).
Yildirim, T. & Alp, R. The role of oxidative stress in the relation between fibromyalgia and obstructive sleep apnea syndrome. Eur Rev Med Pharmacol Sci. 21, 20–29 (2017).
Sancho, R. et al. The CB1/VR1 agonist arvanil induces apoptosis through an FADD/caspase-8-dependent pathway. Br J Pharmacol. 140, 1035–1044 (2003).
Batista, E. D. et al. Food intake assessment and quality of life in women with fibromyalgia. Rev Bras ReumatolEngl Ed. 56, 105–110 (2016).
Lin, J. G., Hsieh, C. L. & Lin, Y. W. Analgesic effect of electroacupuncture in a mouse fibromyalgia model: Roles of TRPV1, TRPV4, and pERK. PLoS One. 10, e0128037 (2015).
Nazıroğlu, M. Activation of TRPM2 and TRPV1 Channels in Dorsal Root Ganglion by NADPH Oxidase and Protein Kinase C Molecular Pathways: a Patch Clamp Study. J Mol Neurosci. 61, 425–435 (2017).
Tsukagoshi, H. et al. Superoxide radical-scavenging effects from polymorphonuclear leukocytes and toxicity in human cell lines of newly synthesized organic selenium compounds. FEBS J. 27, 6046–6054 (2007).
Cheng, Q. et al. The selenium-independent inherent pro-oxidant NADPH oxidase activity of mammalian thioredoxin reductase and its selenium-dependent direct peroxidase activities. J Biol Chem. 285, 21708–21723 (2010).
Uslusoy, F., Nazıroğlu, M. & Çiğ, B. Inhibition of the TRPM2 and TRPV1 Channels through Hypericum perforatum in sciatic nerve injury-induced rats demonstrates their key role in apoptosis and mitochondrial oxidative stress of sciatic nerve and dorsal root ganglion. Front Physiol. 8, 335 (2017).
Pecze, L. et al. Activation of endogenous TRPV1 fails to induce overstimulation-based cytotoxicity in breast and prostate cancer cells but not in pain-sensing neurons. Biochim Biophys Acta 1863, 2054–2064 (2016).
Sarıfakıoğlu, B., Güzelant, A. Y., Güzel, E. C., Güzel, S. & Kızıler, A. R. Effects of 12-week combined exercise therapy on oxidative stress in female fibromyalgia patients. Rheumatol Int. 34, 1361–1367 (2014).
Nazıroğlu, M. Molecular role of catalase on oxidative stress-induced Ca(2+) signaling and TRP cation channel activation in nervous system. J Recept Signal Transduct Res. 32, 134–141 (2012).
Dogrul, A., Gul, H., Yesilyurt, O., Ulas, U. H. & Yildiz, O. Systemic and spinal administration of etanercept, a tumor necrosis factor alpha inhibitor, blocks tactile allodynia in diabetic mice. Acta Diabetol. 48, 135–142 (2011).
Bejarano, I. et al. Melatonin induces mitochondrial-mediated apoptosis in human myeloid HL-60 cells. J Pineal Res. 46, 392–400 (2009).
Uğuz, A. C. et al. Melatonin potentiates chemotherapy-induced cytotoxicity and apoptosis in rat pancreatic tumor cells. J Pineal Res. 53, 91–98 (2012).
Uğuz, A. C. & Nazıroğlu, M. Effects of selenium on calcium signaling and apoptosis in rat dorsal root ganglion neurons induced by oxidative stress. Neurochem Res. 37, 1631–1638 (2012).
Espino, J. et al. Melatonin reduces apoptosis induced by calcium signaling in human leukocytes: Evidence for the involvement of mitochondria and Bax activation. J Membr Biol. 233, 105–118 (2010).
Sedlak, J. & Lindsay, R. H. C. Estimation of total, protein bound and non-protein sulfhydryl groups in tissue with Ellmann’ s reagent. Anal Biochem 25, 192–205 (1968).
Placer, Z. A., Cushman, L. & Johnson, B. C. Estimation of products of lipid peroxidation (malonyl dialdehyde) in biological fluids. Anal Biochem 16, 359–364 (1966).
Suzuki, J. & Katoh, N. A simple and cheap method for measuring vitamin A in cattle using only a spectrophotometer. Jpn J Vet Sci 52, 1282–1284 (1990).
Desai, I. D. Vitamin E analysis methods for animal tissues. Methods Enzymol 105, 138–147 (1984).
Acknowledgements
The abstract of the study was partially published in the 6th World Congress of Oxidative Stress, Calcium Signaling and TRP Channels, 24–27 May 2016, Isparta, Turkey (www.cmos.org.tr). The study was supported by the Scientific Project Unit of SDU (Protocol No: BAP-4562-TU2–16 and coordinator of the project was Prof. Dr. Mehmet Şahin). The authors wish to thank the technician Fatih Şahin (BSN Health Analyses ARGE Ltd., Teknonent, Isparta, Turkey) for helping with plate reader, patch-clamp and western blot analyses.
Author information
Authors and Affiliations
Contributions
M.N. and M.Ş. formulated the present hypothesis and were responsible for writing the report. E.Y. was responsible for inducing experimental F.M. and conducting tests of pain intensity. B.Ç. was responsible for isolating the D.R.N.G. and SciN and analyzing intracellular Ca2+ concentration. All authors approved the final manuscript.
Corresponding author
Ethics declarations
Competing Interests
The authors declare that they have no competing interests.
Additional information
Publisher's note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Yüksel, E., Nazıroğlu, M., Şahin, M. et al. Involvement of TRPM2 and TRPV1 channels on hyperalgesia, apoptosis and oxidative stress in rat fibromyalgia model: Protective role of selenium. Sci Rep 7, 17543 (2017). https://doi.org/10.1038/s41598-017-17715-1
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-017-17715-1
This article is cited by
-
Cross-Talk Between Selenium Nanoparticles and Cancer Treatment Through Autophagy
Biological Trace Element Research (2024)
-
Potential of Natural Products in the Treatment of Glioma: Focus on Molecular Mechanisms
Cell Biochemistry and Biophysics (2024)
-
Involvement of TRPM7 Channel on the Induction of Diabetic Neuropathic Pain in Mice: Protective Role of Selenium and Curcumin
Biological Trace Element Research (2023)
-
Involvement of TRPM2 Channel on Doxorubicin-Induced Experimental Cardiotoxicity Model: Protective Role of Selenium
Biological Trace Element Research (2023)
-
The effects of chronic nanoselenium treatment on sciatic nerve injury: behavioral and biochemical responses
Nutrire (2022)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
