
Abstract
Glioblastoma Multiforme (GBM) is the most frequent and lethal primary brain cancer. Due to its therapeutic resistance and aggressiveness, its clinical management is challenging. Platelet-derived Growth Factor (PDGF) genes have been enrolled as drivers of this tumour progression as well as potential therapeutic targets. As detailed understanding of the expression pattern of PDGF system in the context of GBM intra- and intertumoral heterogeneity is lacking in the literature, this study aims at characterising PDGF expression in different histologically-defined GBM regions as well as investigating correlation of these genes expression with parameters related to poor prognosis. Z-score normalised expression values of PDGF subunits from multiple slices of 36 GBMs, alongside with clinical and genomic data on those GBMs patients, were compiled from Ivy Glioblastoma Atlas Project – Allen Institute for Brain Science data sets. PDGF subunits show differential expression over distinct regions of GBM and PDGF family is heterogeneously expressed among different brain lobes affected by GBM. Further, PDGF family expression correlates with bad prognosis factors: age at GBM diagnosis, Phosphatase and Tensin Homolog deletion and Isocitrate Dehydrogenase 1 mutation. These findings may aid on clinical management of GBM and development of targeted curative therapies against this devastating tumour.
Similar content being viewed by others

Introduction
Glioblastoma multiforme (GBM) is the most frequent and the worst type of glioma, in terms of therapeutic resistance, aggressiveness and associated short life expectancy. At present, management of GBMs is based on surgical resection followed by chemotherapy and/or radiotherapy, which is invasive, comes with unwanted side effects, and does not prevent the tumour from being significantly recurrent1,2. Therefore, efforts have been directed to the characterisation of the molecular and genetic profile of this high grade glioma and the discovery of therapeutic markers that may be the targets of a more specific and effective clinical approach3.
In this context, Platelet-derived Growth Factors (PDGF) constitute a family of six subunits assembled into heterodimer and homodimer ligands and tyrosine kinase receptors, which are enrolled in physiological embryogenesis, haematopoiesis, neuroprotection and glial cell development besides of being identified as part of the GBM molecular panel. These genes have been described as drivers of GBM growth and metastasis, for their role in the dedifferentiation of glial cells into stem cells, the epithelial to mesenchymal transition, the activation of cancer-associated fibroblasts as well as the intratumoral stimulation of angiogenesis, lymphangiogenesis and immunosuppression4,5. Of clinical importance, PDGF system expression and genetic profiles have been reported as prognostic factors and valid therapeutic response biomarkers for a number of cancers, such as sarcomas and breast cancer, and these properties are also been investigated on brain tumors6.
For their prominent influence in GBM development, and in keeping with the perspective of molecular-based curative treatments of this tumour, PDGF genes have been suggested as GBM therapeutic targets and clinical trials have been performed with inhibitors of tyrosine kinase receptors such as Imatinib, which showed only limited beneficial effects. Treatment failure may be attributed to the lack of specificity of these inhibitors as to PDGF receptors5,7. Further, heterogeneity is a hallmark of GBM8 and is thought not to be appreciated by current diagnostic and assessment method of single biopsy and whole tissue analysis of a GBM9. This is likely to give a poor perspective of how is the tumour molecular panel configured and how its already well-described biomarkers are distributed in each single tumour block, which has a direct negative impact on the development of successful targeted therapies.
To motivate further research on GBM, the Ivy Glioblastoma Atlas Project [© 2015 Allen Institute for Brain Science. Ivy Glioblastoma Atlas Project. Available from: http://glioblastoma.alleninstitute.org/] is a scientific initiative that investigates the expression of selected genes on different histological regions of GBM blocks, gathers genomic profile and clinical data of the correspondent patients/donors and publishes this valuable resource in the free-access online platforms of Allen Brain Atlas and Ivy GAP Clinical and Genomic Database. In the present study, PDGF system expression data is compiled from those databases with the aim to characterise these genes distribution in different histological regions of a GBM, so that more specific curative strategies against this tumour may be devised. Moreover, PDGF expression is studied regarding clinical parameters that influence life expectancy of GBM patients, to evaluate the potential role of those genes as prognostic factors in GBM.
Results
Clinical and genomic data on GBM patients herein studied
Information gathered from the online Ivy GAP Clinical and Genomic Database and summarised in Table 1 reveal that the patients who donated the GBM blocks analysed by Allen Institute organisation constituted a young population at the time of diagnosis, with no substantially differential distribution between sexes. Most subjects presented high Karnofsky Performance Status score, which indicates mildly compromised functionality/quality of life and favourable prognosis10,11,12. Nevertheless, average overall survival period illustrates the dramatically shortened life expectancy associated with GBM diagnosis. All patients were treated by the standard combination of surgical tumour resection plus radiotherapy and/or chemotherapy. Rates of Phosphatase and Tensin Homolog (PTEN) loss and Isocitrate Dehydrogenase 1 (IDH1) mutation at R132 were consistent with frequencies reported in the literature: PTEN deletion is considered to be a driver alteration very commonly associated with GBM13, whereas IDH1 mutation is described as being much more prevalent in secondary GBM, rather than in primary tumours14, which are being studied here. More extensive details on each donor’s clinical profile and disease progression have been tabulated and are shown in the Supplementary Table 1.
PDGF family shows varied and heterogenic expression patterns among the GBM regions
We analysed gene expression of the PDGF system (PDGFA, PDGFB, PDGFC, PDGFD, PDGFRA, PDGFRB) in 36 GBMs studied on Ivy Glioblastoma Atlas Project. Histological slices from tumour blocks (Fig. 1a–c) were annotated with molecular markers of seven laser-microdissected GBM regions (Fig. 1d): leading edge, infiltrating tumour, cellular tumour, perinecrotic zone, pseudopalisading cells around necrosis, hyperplastic blood vessels and microvascular proliferation (Fig. 1e). In hyperplastic blood vessels (Fig. 2a), PDGFRB shows high expression levels and PDGFC is less expressed than most of the other subunits, whereas the remaining subunits bears similar expression levels. Interestingly, in microvascular proliferation region (Fig. 2b), PDGFRB and PDGFC are also respectively more and less expressed in comparison to the other members of the family. In addition, PDGFRA bears lower expression values than all other subunits but PDGFC. The pseudopalisading cells (Fig. 2c), on the other hand, presents an inverse pattern: PDGFC is the most expressed PDGF subunit, PDGFRB has low expression values, and the remaining subunits are similarly expressed. PDGFB and PDGFRB are less expressed relatively to PDGFC in perinecrotic zone (Fig. 2d), and also in comparison to PDGFRA in cellular tumour block (Fig. 2e). As to the leading edge (Fig. 2f), PDGFB stands out as the most expressed PDGF subunit in the region. Contrastingly, in the area of infiltrating tumour (Fig. 2g), all subunits are expressed uniformly. Hence, GBM tumour is composed by heterogeneous regions, each one bearing a different PDGF expression pattern.

Workflow of Ivy Glioblastoma Atlas Project data production. Each surgically resected tumour is divided into blocks (a), which are sectioned into histological slices (b). Each section is submitted to histopathological evaluation (c), with annotation (d) of the distinct GBM histological regions (e), which are then processed by laser-microdissection and analysed separately by RNA-sequencing technique. As defined in the Technical White Paper: Overview – 2015 [accessible in glioblastoma.alleninstitute.org], Leading Edge Region: border of the tumour, tumour/normal cells ratio is approximately 1–3/100; Infiltrating Tumour: region in between Leading Edge and Cellular Tumour bulk, tumour/normal cells ratio is approximately 10–20/100; Cellular Tumour: tumour core, tumour/normal cells ratio is approximately 100–500/1; Perinecrotic Zone: boundary of tumour cells closely around necrotic areas in tumour core; Pseudopalisading Cells around Necrosis: characteristic rows of lined-up, aggregated cells surrounding necrotic areas in tumour core; Hyperplastic Blood Vessels in Cellular Tumour: aggregated blood vessels with thickened walls, in tumour core; Microvascular Proliferation: glomerulus-like conformation of a couple of blood vessels that share vessel wall, inside the tumour core. All images are credited to Allen Institute. (a), (b) and (e) are available on the Technical White Paper: Overview – 2015, accessible in glioblastoma.alleninstitute.org; (c) and (d) are available at http://glioblastoma.alleninstitute.org/ish/specimen/show/706783?gene=5127. Image credit: Allen Institute.

PDGF family expression patterns on each GBM histological region reveal intratumoral heterogeneity. PDGF system expression was evaluated in GBM regions of hyperplastic blood vessels in cellular tumour (a); microvascular proliferation (b); pseudopalisading cells around necrosis (c); perinecrotic zone (d); cellular tumour (e); leading edge (f), and infiltrating tumour (g).*p < 0.05; **p < 0.01; ***p < 0.001; ****p < 0.0001.
PDGF subunits show differential expression along the GBM tumour
Next, we analysed the distribution of each PDGF subunit along the GBM areas, by means of expression values. Both PDGFA (Fig. 3a) and PDGFB (Fig. 3b) subunits appear to be most expressed on microvascular proliferation, and expression levels in hyperplastic blood vessels are higher than on most of the remaining GBM regions. Differently, PDGFC (Fig. 3c) is less expressed in microvascular proliferation in comparison to all other GBM regions aside from hyperplastic blood vessel and leading edge, and similarly to PDGFA (Fig. 3a) and PDGFRA (Fig. 3e), it bears a preferential expression in cellular tumour bulk over leading edge. As to PDGFD (Fig. 3d) and PDGFRB (Fig. 3f), expression is the highest in areas of angiogenic alterations. Contrastingly, PDGFRA (Fig. 3e) is not uniformly distributed among vascular regions, being more expressed in hyperplastic blood vessels than on microvascular proliferation. Additionally, Allen Brain Institute has made available a set of in situ hybridization images of PDGF subunits, for eight GBMs. Although, at visual analysis, each subunit presents a particular expression pattern, comparison to the correspondents annotated histological sections does not reveal a clear correlation between gene expression and GBM region (see Supplementary Fig. S1). Altogether, these results characterise the heterogenic distribution of the PDGF family over GBM blocks, with the preferential expression of most of the subunits on areas of hyperplastic blood vessels and microvascular proliferation. However, such differential expression may not be appreciated in limited samples, with subjective methods of analysis.

PDGF genes are differentially expressed over different histological regions of a GBM. Expression pattern along distinct areas of a GBM was assessed for each PDGF system component: PDGFA (a); PDGFB (b); PDGFC (c); PDGFD (d); PDGFRA (e); PDGFRB (f). CT-HBV: Hyperplastic Blood Vessels in Cellular Tumour; CT-MVP: Microvascular Proliferation; CT-PAN: Pseudopalisading Cells around Necrosis; IT: Infiltrating Tumour; LE: Leading Edge; CT: Cellular Tumour. *p < 0.05; **p < 0.01; ***p < 0.001; ****p < 0.0001.
PDGF family is also heterogeneously expressed over the different locations of GBM in the brain
Following the analysis of PDGF expression in histological GBM regions, we studied PDGF system expression with regard to the GBM-affected part of the brain. Although PDGF family seems to have overall similar expression levels between GBMs located in the right hemisphere and in the left one (Fig. 4a), analysis of PDGF expression per lobes of the brain reveals a more heterogeneous panel. PDGFRA is more expressed in GBMs of the left temporal lobe than in those of the right one (Fig. 4b), whereas PDGFA expression is higher when the tumour is set on right frontal (Fig. 4c) and parietal (Fig. 4d) lobes as opposed to the left correspondents. Therefore, GBM appears to present intertumoral heterogeneity as to PDGF family, of which the expression pattern changes depending on the location of the tumour in the brain.

Analysis of PDGF genes expression regarding GBM location in the brain: Comparison was made on PDGF family expression between tumours located at left and at right hemispheres (a); PDGF subunits expression was studied on GBMs at temporal lobe (b), frontal lobe (c) and parietal lobe (d). *p < 0.05; **p < 0.01; ***p < 0.001; ****p < 0.0001.
PDGF expression is a potential bad prognosis marker
Thanks to the extensive clinical and genomic data on the donors of the GBM blocks herein studied, available on Ivy GAP Clinical and Genomic Database, we were able to analyse PDGF genes expression on the GBM as to the presence of molecular or clinical markers that determine a bad prognosis. No significant correlation was found between PDGF expression and Epidermal Growth Factor Receptor genetic and mutational status, Karnofsky Performance Status score and methylation status of O6-methylguanine-DNA-methyltransferase promoter (see Supplementary Fig. S2). On the other hand, GBMs that present PTEN deletion, considered to be a bad prognosis marker13,15,16,17, have higher PDGF family expression levels than the tumours with PTEN gain, which correlates with a better prognosis (Fig. 5a); analysis of each PDGF subunit separately reveals that PDGFA is differentially expressed between these two types of GBM (Fig. 5b). Likewise, GBMs with wild-type IDH1, a bad prognosis determinant10,11,14, show significantly (p value 0.0004) higher overall PDGF family expression over the ones bearing the mutated version of this gene (Fig. 5c), and subunit-by-subunit analysis shows that PDGFA, in particular, follows this pattern (Fig. 5d). As to the age at the time of GBM diagnosis, those with more than 65 years-old are reported to have poorer prognosis12, and among the subjects herein investigated, tumours from patients in this age group are characterised by greater PDGF family expression levels (Fig. 5e) and increased PDGFA expression over PDGFRA (Fig. 5f). Contrastingly, GBMs of patients diagnosed before or at 65 years-old present lower PDGF family expression (Fig. 5e) and uniform expression of PDGF subunits (Fig. 5g). Thus, the expression levels of PDGF family, especially the subunit PDGFA, correlate with the presence of poor prognostic factors, which suggests these genes may be viewed as prognostic markers themselves.

PDGF expression correlates with prognostic factors of GBM. PDGF system expression was analysed according to prognostic factors of Phosphatase and Tensin Homolog (PTEN) deletion (a,b); Isocitrate Dehydrogenase 1 (IDH1) mutation (c,d), and age at diagnosis of GBM (e,f,g). *p < 0.05; **p < 0.01; ***p < 0.001; ****p < 0.0001.
Discussion
This study presents a broad perspective on the inter- and intratumoral heterogeneity of the PDGF family expression in GBMs, along with the potential prognostic significance of these genes expression, from analysis of the comprehensive database of the Ivy Glioblastoma Atlas Project – Allen Institute for Brain Science.
The “multiforme” designation of this high-grade tumour is much accurate on making explicit its wide heterogeneous milieu, which has been studied by different groups at cytogenic, transcriptional, mutational, epigenetic and proteomic levels, being regarded as key point to understanding differential responses to treatment as well as therapeutic resistance so common to GBM patients18,19,20,21,22,23,24,25. The level of heterogeneity on a GBM is linked to poor survival26 and seems to represent a spatially and temporally dynamic multistep-process that involves genetic instability and clonal proliferation, equipping the tumour cells with aggressiveness and resourcefulness to survival and growth27,28.
GBM has been reported to bear copy number aberrations and overexpression of receptor tyrosine kinases, especially PDGFRA, EGFR (Epidermal Growth Factor Receptor) and MET proto-oncogene, in a significantly non-homogeneous presentation, with subpopulations of the same tumour presenting each genetic alteration in a mutually exclusive, mosaic-like way26,29,30,31,32. Interestingly, PDGF genes have been shown to present differential expression between glial cells of tumour mass and endothelial cells of angiogenic alterations in the tumour, with description of autocrine and paracrine loops involving specific PDGF receptors and ligands, which are thought to perpetuate cell proliferation and tumour growth in the different areas33,34,35. Moreover, PDGFRβ has been described to be the type of PDGF receptor preferentially expressed in GBM stem-cells and to promote their self-renewal and invasion, which is likely correlated with tumour recurrence and resistance to therapeutics36.
However, at the moment of this publication, the authors had not found any report regarding analysis of the entire PDGF family expression among different histologically-defined GBM regions, as performed here. Likewise, intertumoral heterogeneity as to PDGF genes expression had not been studied by means of comparison of tumours located in different brain lobes. Thus, the present work reaffirms the emblematic feature of heterogeneity on GBM, characterising the distribution and pattern of expression of PDGF, which is associated not only to normal neurogenesis but also to glial tumour initiation and progression37,38,39.
Overall and progression-free survival periods are usually very short after GBM clinical presentation, as illustrated by the subjects studied here, the majority of which died less than one year after GBM diagnosis. Patients with post-diagnosis survival period of more than 2.5 years are classified as long-term survivors40. Because of this tragic prognosis, multiple markers have been investigated with the aim of stratifying patients accordingly to disease severity and therapeutic options. In this context, age at GBM diagnosis, IDH1 mutations and PTEN deletions have been described as independent prognostic factors10,11,12,13,14,15,17,41. The herein shown correlation between PDGF genes expression and the clinical and genomic prognostic factors aforementioned suggests that those genes should be further considered as additional prognosis markers that may aid clinical management of GBM patients.
Growing evidence as to the significance of understanding inter- and intratumoral heterogeneity, as well as prognosis biomarkers, for the stratification of GBM patients and decisions over therapeutic strategies for them has been taken into consideration in the clinical setting. The characterisation of tumour subtypes with specific transcriptional profiles described by groups such as Verhaak et al., 201042 has been applied in the interpretation of GBM heterogeneity and prognosis in recent studies and has motivated the newest revision of World Health Organisation classification of central nervous system tumours towards a molecular-based analysis of each tumour together with the traditional histopathological appraisal43,44. A more precise classification of GBM, taking into account its heterogeneity, will certainly have a positive effect on implementation of targeted therapy tailored for each patient, which greatly increases the chances of changing the current poor prognosis paradigm and reaching curative treatments in the near future.
In brief, the present study contributes to the characterisation of GBM heterogeneity as it reveals that PDGF genes show specific expression patterns through different regions of a GBM as well as differential expression accordingly to the location of the tumour in the brain. Of note, the PDGF family can also be linked to prognostic factors of the GBM. Taken together, these results should contribute to the realization of personalised medicine towards the development of successful therapeutics against this so common and so devastating tumour.
Methods
Compilation from Allen Institute resources
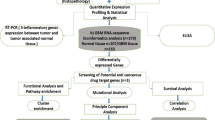
Gene expression, clinical and genomic data on 36 primary GBMs and their donors were compiled from two different platforms of Ivy Glioblastoma Atlas Project [© 2015 Allen Institute for Brain Science. Ivy Glioblastoma Atlas Project. Available from: glioblastoma.alleninstitute.org]. Z-score normalised expression values of PDGF subunits were downloaded from the Anatomic Structures RNA-Sequencing data set of the online Allen Brain Atlas [Available in: glioblastoma.alleninstitute.org/rnaseq/search/index.html, last access in May 2016]. Gene expression data available in this open access atlas was obtained by RNA sequencing technique, applied to the seven GBM histological structures that were isolated by laser micro-dissection in each histological section of tumour blocks: Leading Edge, Infiltrating Tumour, Cellular Tumour, Perinecrotic Zone, Pseudopalisading Cells around Necrosis, Hyperplastic Blood Vessels in Cellular Tumour and Microvascular Proliferation. All values are described to be processed through post-hoc data normalisation followed by TbT normalisation. More detailed description on the methodology of gene expression data production is available on the Technical White Paper: Overview – 2015, accessible in glioblastoma.alleninstitute.org. Additionally, extensive clinical and genomic data of the patients recruited for the study are available in the online Ivy GAP Clinical and Genomic Database [Available in: ivygap.swedish.org/home, last access in May 2016] and were gathered and tabulated.
Analysis of compiled data
Gene expression values of PDGF subunits (PDGFA, PDGFB, PDGFC, PDGFD, PDGFRA, PDGFRB) were analysed with regard to variables such as GBM histological structure, clinical parameters and genomic data. GraphPad Prism version 7.00 for Windows, GraphPad Software, La Jolla California USA (www.graphpad.com) was used for graphical representation and statistical analysis. Differences between two sample groups were assessed by Mann-Whitney test, whereas multiple comparisons were evaluated by Kruskal-Wallis test followed by Dunn’s Multiple Comparisons test. Values were assessed as medians and correlations were considered statistically significant if p value <0.05.
Data availability statement
The datasets analysed during the current study are from the © 2015 Allen Institute for Brain Science - Ivy Glioblastoma Atlas Project, available in: glioblastoma.alleninstitute.org and ivygap.swedish.org/home.
References
Urbanska, K., Sokolowska, J., Szmidt, M. & Sysa, P. Glioblastoma multiforme - An overview. Wspolczesna Onkol. 18, 307–312 (2014).
Omuro, A. & LM, D. Glioblastoma and other malignant gliomas: A clinical review. JAMA 310, 1842–1850 (2013).
Vitucci, M., Hayes, D. N. & Miller, C. R. Gene expression profiling of gliomas: merging genomic and histopathological classification for personalised therapy. Br. J. Cancer 104, 545–553 (2011).
Cao, Y. Multifarious functions of PDGFs and PDGFRs in tumor growth and metastasis. Trends Mol. Med. 19, 460–473 (2013).
Nazarenko, I. et al. PDGF and PDGF receptors in glioma. Ups. J. Med. Sci. 117, 99–112 (2012).
Paulsson, J., Ehnman, M. & Ostman, A. PDGF receptors in tumor biology: prognostic and predictive potential. Future Oncology. 10, 1695–1708 (2014).
Heldin, C.-H. Targeting the PDGF signaling pathway in tumor treatment. Cell Commun. Signal. 11, 97 (2013).
Inda, M.-D.-M., Bonavia, R. & Seoane, J. Glioblastoma multiforme: a look inside its heterogeneous nature. Cancers (Basel). 6, (226–39 (2014).
Parker, N. R., Khong, P., Parkinson, J. F., Howell, V. M. & Wheeler, H. R. Molecular heterogeneity in glioblastoma: potential clinical implications. Front. Oncol. 5, 55 (2015).
Molenaar, R. J. et al. The combination of IDH1 mutations and MGMT methylation status predicts survival in glioblastoma better than either IDH1 or MGMT alone. Neuro. Oncol. 16, 1263–1273 (2014).
Ohno, M. et al. Glioblastomas with IDH1/2 mutations have a short clinical history and have a favorable clinical outcome. Jpn. J. Clin. Oncol. 46, 31–39 (2016).
Ening, G., Osterheld, F., Capper, D., Schmieder, K. & Brenke, C. Charlson comorbidity index: an additional prognostic parameter for preoperative glioblastoma patient stratification. J. Cancer Res. Clin. Oncol. 141, 1131–1137 (2015).
Xu, J., Li, Z., Wang, J., Chen, H. & Fang, J. Y. Combined PTEN mutation and protein expression associate with overall and disease-free survival of glioblastoma patients. Transl. Oncol. 7, 196–205 (2014).
Chen, J.-R., Yao, Y., Xu, H.-Z. & Qin, Z.-Y. Isocitrate Dehydrogenase (IDH)1/2 Mutations as Prognostic Markers in Patients With Glioblastomas. Medicine (Baltimore). 95, e2583 (2016).
Sano, T. et al. Differential Expression of MMAC / PTEN in Glioblastoma Multiforme: Relationship to Localization and Prognosis Advances in Brief Differential Expression of MMAC / PTEN in Glioblastoma Multiforme: Relationship to Localization and Prognosis 1. Cancer Res. 1820–1824 (1999).
Phillips, H. S. et al. Molecular subclasses of high-grade glioma predict prognosis, delineate a pattern of disease progression, and resemble stages in neurogenesis. Cancer Cell 9, 157–173 (2006).
Gont, A. et al. PTEN loss represses glioblastoma tumor initiating cell differentiation via inactivation of Lgl1. Oncotarget 4, 1266–79 (2013).
Jung, V. et al. Evidence of Focal Genetic Microheterogeneity in Glioblastoma Multiforme by Area-Specific CGH on Microdissected Tumor Cells. J. Neuropathol. Exp. Neurol. 58, 993–999 (1999).
Parker, N. R. et al. Intratumoral heterogeneity identified at the epigenetic, genetic and transcriptional level in glioblastoma. Sci. Rep. 6, 22477 (2016).
Kumar, A. et al. Deep sequencing of multiple regions of glial tumors reveals spatial heterogeneity for mutations in clinically relevant genes. Genome Biol. 15, 530 (2014).
de Aquino, P. F. et al. A Time-Based and Intratumoral Proteomic Assessment of a Recurrent Glioblastoma Multiforme. Front. Oncol. 6, 1–10 (2016).
Hoelzinger, D. B. et al. Gene expression profile of glioblastoma multiforme invasive phenotype points to new therapeutic targets. Neoplasia 7, 7–16 (2005).
Lemée, J.-M., Clavreul, A. & Menei, P. Intratumoral heterogeneity in glioblastoma: Don’t forget the peritumoral brain zone. Neuro. Oncol. 17, 1322–1332 (2015).
Meyer, M. et al. Single cell-derived clonal analysis of human glioblastoma links functional and genomic heterogeneity. Proc. Natl. Acad. Sci. USA 112, 851–856 (2015).
Harada, K. et al. Intratumoral cytogenetic heterogeneity detected by comparative genomic hybridization and laser scanning cytometry in human gliomas. Cancer Res. 58, 4694–4700 (1998).
Patel, A. P. et al. Single-cell RNA-seq highlights intratumoral heterogeneity in primary glioblastoma. Science 344, 1396–1401 (2014).
Nickel, G. C. et al. Characterizing mutational heterogeneity in a glioblastoma patient with double recurrence. PLoS One 7, 1–8 (2012).
Sottoriva, A. et al. Intratumor heterogeneity in human glioblastoma reflects cancer evolutionary dynamics. Proc Natl Acad Sci USA 110, 4009–4014 (2013).
Little, S. E. et al. Receptor tyrosine kinase genes amplified in glioblastoma exhibit a mutual exclusivity in variable proportions reflective of individual tumor heterogeneity. Cancer Res. 72, 1614–1620 (2012).
Snuderl, M. et al. Mosaic amplification of multiple receptor tyrosine kinase genes in glioblastoma. Cancer Cell 20, 810–817 (2011).
Okada, Y. et al. Selection pressures of TP53 mutation and microenvironmental location influence epidermal growth factor receptor gene amplification in human glioblastomas. Cancer Res. 63, 413–416 (2003).
Szerlip, N. J. et al. Intratumoral heterogeneity of receptor tyrosine kinases EGFR and PDGFRA amplification in glioblastoma defines subpopulations with distinct growth factor response. Proc. Natl. Acad. Sci. 109, 3041–3046 (2012).
Hermansson, M. et al. Endothelial cell hyperplasia in human glioblastoma: coexpression of mRNA for platelet-derived growth factor (PDGF) B chain and PDGF receptor suggests autocrine growth stimulation. Proc. Natl. Acad. Sci. USA 85, 7748–52 (1988).
Hermanson, M. et al. Platelet-derived Growth Factor and Its Receptors in Human Glioma Tissue: Expression of Messenger RNA and Protein Suggests the Presence of Autocrine and Paracrine Loops. Cancer Res. 52, 3213–3219 (1992).
Lokker, N. A., Sullivan, C. M., Hollenbach, S. J., Israel, M. A. & Giese, N. A. Platelet-derived Growth Factor (PDGF) Autocrine Signaling Regulates Survival and Mitogenic Pathways in Glioblastoma Cells. Cancer Res. 62, 3729–3735 (2002).
Kim, Y. et al. Platelet-derived growth factor receptors differentially inform intertumoral and intratumoral heterogeneity. Genes Dev. 26, 1247–1262 (2012).
Uhrbom, L., Hesselager, G., Nister, M. & Westermark, B. Induction of brain tumors in mice using a recombinant platelet-derived growth factor B-chain retrovirus. Cancer Res. 58, 5275–5279 (1998).
Shih, A. H. et al. Dose-Dependent Effects of Platelet-Derived Growth Factor-B on Glial Tumorigenesis. Cancer Res. 64, 4783–4789 (2004).
Funa, K. & Sasahara, M. The roles of PDGF in development and during neurogenesis in the normal and diseased nervous system. J. Neuroimmune Pharmacol. 9, 168–181 (2014).
Smoll, N. R., Schaller, K. & Gautschi, O. P. Long-term survival of patients with glioblastoma multiforme (GBM). J. Clin. Neurosci. 20, 670–675 (2013).
Duan, S. et al. PTEN deficiency reprogrammes human neural stem cells towards a glioblastoma stem cell-like phenotype. Nat. Commun. 6, 10068 (2015).
Verhaak, R. G. W. et al. Integrated Genomic Analysis Identifies Clinically Relevant Subtypes of Glioblastoma Characterized by Abnormalities in PDGFRA, IDH1, EGFR, and NF1. Cancer Cell 17, 98–110 (2010).
Louis, D. N. et al. The 2016 World Health Organization Classification of Tumors of the Central Nervous System: a summary. Acta Neuropathol. 131, 803–820 (2016).
Verhaak, R. G. W. Moving the needle: Optimizing classification for glioma. Sci. Transl. Med. 8, 350fs14 (2016).
Acknowledgements
This work was funded by the Conselho Nacional de Desenvolvimento Científico e Tecnológico - CNPq, CAPES, PROPESQ-UFPE and the authors are grateful for the genererous donation from Nubia Martins. The authors thankfully acknowledge the comprehensive Allen Institute for Brain Science datasets, especially the Ivy Glioblastoma Atlas Project. Valuable inputs from the post-doctorate researchers Roberta Lemos and Eriton Cunha on data analysis are greatly appreciated. Manuscript revision by the cancer biologist Dr Gaëlle Bougeard-Denoyelle provided significant contribution to the refinement of this paper.
Author information
Authors and Affiliations
Contributions
J.R.M.O. devised and supervised the project. I.G.C. performed compilation and analysis, composed the figures/table and elaborated the manuscript. All authors reviewed and revised the manuscript.
Corresponding author
Ethics declarations
Competing Interests
The authors declare that they have no competing interests.
Additional information
Publisher's note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Cantanhede, I.G., de Oliveira, J.R.M. PDGF Family Expression in Glioblastoma Multiforme: Data Compilation from Ivy Glioblastoma Atlas Project Database. Sci Rep 7, 15271 (2017). https://doi.org/10.1038/s41598-017-15045-w
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-017-15045-w
This article is cited by
-
Shaping the brain vasculature in development and disease in the single-cell era
Nature Reviews Neuroscience (2023)
-
PDGFRA, KIT, and KDR Gene Amplification in Glioblastoma: Heterogeneity and Clinical Significance
NeuroMolecular Medicine (2023)
-
A single-cell based precision medicine approach using glioblastoma patient-specific models
npj Precision Oncology (2022)
-
Scavenger receptor class F member 2 (SCARF2) as a novel therapeutic target in glioblastoma
Toxicological Research (2022)
-
The HIF1α-PDGFD-PDGFRα axis controls glioblastoma growth at normoxia/mild-hypoxia and confers sensitivity to targeted therapy by echinomycin
Journal of Experimental & Clinical Cancer Research (2021)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
