
Abstract
As a nongenetic engineering technique, adaptive evolution is an effective and easy-to-operate approach to strain improvement. In this work, a commercial Thermoanaerobacterium aotearoense SCUT27/Δldh-G58 was successfully isolated via sequential batch fermentation with step-increased carbon concentrations. Mutants were isolated under selective high osmotic pressures for 58 passages. The evolved isolate rapidly catabolized sugars at high concentrations and subsequently produced ethanol with good yield. A 1.6-fold improvement of ethanol production was achieved in a medium containing 120 g/L of carbon substrate using the evolved strain, compared to the start strain. The analysis of transcriptome and intracellular solute pools suggested that the adaptive evolution altered the synthesis of some compatible solutes and activated the DNA repair system in the two Thermoanaerobacterium sp. evolved strains. Overall, the results indicated the potential of adaptive evolution as a simple and effective tool for the modification and optimization of industrial microorganisms.
Similar content being viewed by others


Introduction
Microbial fermentation is continuously developed to shorten fermentation time, lower costs, and increase product yields1. High-sugar-medium fermentation has several advantages, including improved device efficiency, enhanced product output per unit time, decreased heat and water consumption, and simplified downstream extraction processes2, 3. In bioethanol production, high-concentration mash fermentation techniques are primarily used because it can increase cell density, product concentration, and production rate2, 4. At 25% (w/v) sugar medium, more than 15% (v/v) of ethanol can be obtained by Saccharomyces cerevisiae, which exhibits good tolerance to sugars and metabolites3.
High substrate concentration leads to elevated osmotic pressure and can thus cause cell dehydration5, plasmolysis6, and even cell inactivation7. These effects results in extended lag phase, metabolic disturbance, low productivity, and stagnation of fermentation8, 9. Thus, bacteria resistant to hypertonic solution are necessary to economic industrial production. Unfortunately, most native microorganisms cannot tolerate extreme hyperosmotic environments. Thus, processes, such as domestication, which is a relatively simple and effective method to produce tolerant mutants, is frequently employed. Through adaptive evolution and process optimization, S. cerevisiae and Zymomonas mobilis can utilize more than 250 g/L of glucose for ethanol production2, 3, 5, 9. Furthermore, an engineered Escherichia coli strain was adaptively evolved for xylose resistance at 120 g/L xylose and production of d-lactate is 50% higher than that by a start strain under same conditions10.
In the present study, the tolerance and ethanol productivity of a genetically engineered Thermoanaerobacterium aotearoense SCUT27/Δldh 11 subjected to a 120 g/L sugar substrate was enhanced through adaptive evolution. This bacterium can produce ethanol with high yield from lignocellulosic biomass11, 12. Bacterial regulatory response against high substrate conditions was further investigated.
Results and Discussion
Adaptive evolution under 120 g/L sugar stress
A high initial sugar concentration during fermentation is favorable because it increases fermentation efficiency at low process water and energy conditions13. In the present study, two high-carbon-source concentrations (81 and 120 g/L of glucose and xylose at gram ratio of 2:1) were evaluated for the modified strain SCUT27/Δldh 11, 12 in 125 mL serum bottles. After 50-h cultivation, no apparent cell growth was observed at the initial substrate concentration of 120 g/L.
The strain was subjected to a gradually increasing sugar concentration to increase the survival or reproduction of ethanol-producing strain SCUT27/Δldh in a medium with high initial sugar concentration. Only the individuals capable of growing under controlled environments were selected and transferred (Fig. 1). The start sugar concentration was set at 81 g/L in the evolution experiments. In the initial 16 passages, the cells had a low growth rate (approximately 0.03 g/L · h). The growth rate then showed a shock-type increase and was stabilized at 0.10–0.14 g/L · h. Meanwhile, the survival cells adapted well to 100 g/L sugar medium. The cells were then inoculated into a medium containing 120 g/L of carbon source. Low growth rate and shock-type growth increase were observed again until the cells remained larger than 0.10 g/L · h for four passages. Finally, 0.2 mL of the 58-passage culture was plated on the DSMZ 640 agar plates and incubated at 50 °C for 3 days. A total of 10 single colonies were selected and transferred into serum bottles containing 120 g/L of sugar each. After five-time transfer, only one colony exhibited steady growth in the 120 g/L medium. This colony was designated as SCUT27/Δldh-G58 and subjected to further research.

Adaptive evolution of T. aotearoense SCUT27/Δldh to step-increasing sugar concentration medium. Sugar was mixed by glucose and xylose at a ratio of 2:1 (g:g) and its concentration was indicated by the grey background.
Fermentation characteristics of SCUT27/Δldh and G58
For the comparison between SCUT27/Δldh and G58 with regard to fermentation, the strains were cultured separately with 30, 81, and 120 g/L total sugar substrate in 125 mL serum bottles, and their growth profiles were monitored (Fig. 2). In the low-sugar medium (30 g/L), the growth profiles were roughly similar to those of the start strain SCUT27/Δldh and its derivative G58. The strain took approximately 8 h to reach the dry cell weight (DCW) of 0.5 g/L. After 82 h incubation, the final DCW of the SCUT27/Δldh was only 0.8 g/L when the substrate concentration was increased to 81 g/L. However, no apparent cell growth in SCUT27/Δldh was observed during the 60 h cultivation at 120 g/L initial sugar concentration. The metabolite compositions of T. aotearoense SCUT27/Δldh and G58 were detected after a 48 h incubation (see Supplementary Table S1). For the parent strain SCUT27/Δldh, the concentration and yield of ethanol decreased at increased initial sugar concentration, indicating that high-sugar level can markedly inhibit cell growth and extend lag time. However, the evolved strain G58 showed substantial tolerance to high substrate concentrations. No considerable inhibition of cell growth was observed even when the initial substrate concentration was increased to 120 g/L. The cell growth curves at different concentration media clearly demonstrated that the evolved strain gained resistance to 120 g/L sugar environments. Concurrently, no change was noted in the yield of ethanol production of SCUT27/Δldh-G58 subjected to different concentrations of carbon source.

Growth profiles of T. aotearoense SCUT27/Δldh and G58 in three different concentration mediums. (a) 30 g/L, (b) 81 g/L and (c) 120 g/L. Solid circles indicated the parent strain of SCUT27/Δldh; hollow circles represented the evolved SCUT27/Δldh-G58.
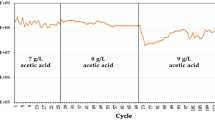
The strains were cultured in a 5 L fermenter containing 2 L of medium with 80 g/L of glucose and 40 g/L of xylose to investigate the fermentation characteristics of SCUT27/Δldh and G58. After inoculation, cell growth was observed immediately in the strain of SCUT27/Δldh-G58 (Fig. 3a). By contrast, approximately 75 h lag time was required to enable the SCUT27/Δldh to acclimate in environments subjected to high osmotic pressures. After 200 h fermentation, the final DCW values of SCUT27/Δldh and G58 were approximately 1.32 and 2.96 g/L, respectively. In addition, ethanol produced by strain G58 reached 36.2 g/L, which was 1.6-fold higher than that produced by SCUT27/Δldh (Fig. 3b). The glucose and xylose content in G58 were consumed instantaneously, and the consumption percentages were 78.5% and 97.5%, respectively (Fig. 3c). The sugar consumption results indicated that no significant carbon catabolite repression occurred in the mixed sugar medium and thus are consistent with our previous reports14. Meanwhile, for SCUT27/Δldh, only 52.4% of initial glucose and 76.5% of initial xylose were consumed. Low sugar utilization for SCUT27/Δldh was in accordance with its cell growth and metabolite production. Ethanol yields were 0.35 and 0.39 g/g for the start and resultant G58 strain, respectively. It should be noted that during the last 50 hours of fermentation, almost no changes of sugar consumption, cell growth or ethanol production were recorded for SCUT27/Δldh. However, the corresponding sugar depletion and ethanol accumulation rates of SCUT27/Δldh-G58 were remained as 0.19 ± 0.05 g/L/h and 0.17 ± 0.06 g/L/h, respectively. We believe that more ethanol could be produced for SCUT27/Δldh-G58 if the fermentation time was extended, without considering the ethanol inhibition on the bacterium. This study demonstrated that adaptive evolution can dramatically shorten the lag time and increase the final ethanol titer in 120 g/L sugar concentration media while eliciting minimal alteration in ethanol yield (Table 1).

Characteristic comparison of SCUT27/Δldh and G58 in 5-L fermenter containing 120 g/L sugars. (a) Cell growth curves. (b) Produced ethanol (circle) and acetic acid (triangle). (c) Residual sugar concentration including glucose (circle) and xylose (triangle). Solid symbols represented the SCUT27/Δldh, and the corresponding hollow ones indicated the SCUT27/Δldh-G58.
Strong tolerance to high-sugar medium enables cells to consume and use sugar rapidly, thereby rendering the strains competitive for industrial fermentation. Although genetic engineering is complex and varies in difficulty in different species, it is a direct approach that improves strain resistance to high osmotic pressures15,16,17. However, genetic manipulation in T. aotearoense SCUT27 is immature and unstable. Thus, transformation and competent cell preparation methods, such as electroporation11 and natural competence cell18, were employed by our group to transfer exogenous DNA into T. aotearoense SCUT27. However, only a few of these methods worked11, 19, and successful results were not reproduced. Another simple method of obtaining tolerant strains is performing long-term adaptation studies. Liang et al.20 employed adaptive evolution to increase the tolerance of C. thermocellum to cellobiose, reaching the highest concentration of 50 g/L after 13 weeks. They were able to produce 22.4 g/L of final ethanol by using 60 g/L of cellulose as substrate. E. coli can be cultured in a medium with gradually increasing ethanol content. Some studies reported that evolved strains have enhanced resistance to ethanol and decreased sensitivity to toxic aldehydes after 3 months of cultivation21, 22. In a previous study, T. aotearoense LA1002 was domesticated to adapt to high sugar concentration23. The results showed that the evolved strain can survive and ferment in 100 g/L of substrate (90 g/L glucose and 10 g/L fructose) for lactic acid production. The LA1002-G40 was used to produce lactic acid from mixed bakery waste hydrolysates, yielding 0.85 g/g. In the present study, resistance to sugar concentration was enhanced to 120 g/L over a 1.5-month period for T. aotearoense SCUT27/Δldh fermentation. Moreover, the resultant strain improved xylose utilization and ethanol productivity. Thus, this tolerance phenotype of SCUT27/Δldh-G58 has potential in lignocellulosic biomass fermentation.
Propagation stability of evolved strain
The phenotype stability of SCUT27/Δldh-G58 was examined by passing on from transfer to transfer in 120 g/L mixed sugar medium. After the strain was subcultured for 40 times, standard deviation values of different culture transfer in cell growth and metabolite spectrum were lower than 5%, indicating no significant change. This result showed that SCUT27/Δldh-G58 is resistant to high sugar conditions and thus can be used for ethanol production.
Transcriptome and intracellular solute pools analysis
Adaptive evolution enables SCUT27/Δldh-G58 to have a short lag phase, fast sugar consumption, and high ethanol production in a 120 g/L sugar medium. However, the regulatory mechanisms involved in the tolerance of SCUT27/Δldh-G58 to high-substrate medium remain unidentified. Thus, the total RNAs of SCUT27/Δldh and G58 were extracted for sequencing and transcriptome analysis. Concurrently, another adaptation-evolved strain of T. aotearoense LA1002-G40 for lactic acid production23 was analyzed. The RNAseq data have been deposited in the GenBank database under the accession number PRJNA385555 (SCUT27/Δldh vs G58) and PRJNA385757 (LA1002 vs G40).
According to the analysis report of Genewiz, the total number of differential expression genes (|log(change fold)| ≥ 2.0, FDR ≤ 0.05) in LA1002-G40 was higher than that in SCUT27/Δldh-G58 (80 vs. 53, see Supplementary Tables S2 and S3). In SCUT27/Δldh-G58, 33 and 20 genes were upregulated and downregulated, respectively. In LA1002-G40, 52 genes were upregulated and 28 genes were downregulated. For the evaluation of the RNA sequencing results, change folds in the transcript abundance of selected genes were assessed via RT-qPCR. Both the RNA-seq and RT-qPCR results displayed the same direction (upregulation or downregulation) of differential genes (Table 2). Altered genes were enriched into 41 and 20 pathways in LA1002-G40 and SCUT27/Δldh-G58, respectively, as indicated by the function analysis results.
Enhancing UDP-glucose synthesis to improve intracellular osmotic pressure
The transcriptional profiles of SCUT27/Δldh versus G58 and LA1002 versus G40 both showed that several genes involved in galactose metabolism pathway (ko00052) were upregulated after adaptive evolution (Table 2). However, the related specific genes were not consistent in the two profiles. In the SCUT27/Δldh versus G58 profile, the differentially expressed genes included Tsac_1294, Tsac_1295, Tsac_0148, and Tsac_1296, which can facilitate the increase in the flux of lactose to UDP-galactose (Fig. 4a). Meanwhile, in the LA1002 versus G40 profile, the differentially expressed genes were Tsac_1476, Tsac_2540, and Tsac_2297, which are beneficial to metabolic flux improvement from glucose to UDP-glucose. Tsac_1296 and Tsac_0599 are isoenzymes upregulated in SCUT27/Δldh versus G58 and LA1002 versus G40 profiles, respectively. Notably, the significant downregulation of Tsac_1517 in LA1002 versus G40 profile may reduce consumption and maintain high trehalose levels in cells. As the direct precursor of trehalose synthesis24, the intracellular UDP-glucose concentration was determined. It was recorded as 0.0362 ± 0.0024% and 0.0199 ± 0.0017% of dry cell weight for SCUT27/Δldh-G58 and SCUT27/Δldh, respectively. Similarly, intracellular UDP-glucose was also increased by 1.88-fold from 0.0230 ± 0.0021% (LA1002) to 0.0432 ± 0.0038% (LA1002-G40) of dry cell weight. The improvement facts of intracellular UDP-glucose were consistent with the transcriptome analysis results, and partially supported the upregulation of intracellular trehalose levels in the evolved strain.

Summary of regulation profiles in T. aotearoense SCUT27/Δldh vs. G58 and LA1002 vs. G40. Purple symbols, up-regulated genes in SCUT27/Δldh-58; blue symbols, down-regulated genes in SCUT27/Δldh-G58; red symbols, up-regulated genes in LA1002-G40; green symbols, down-regulated genes in LA1002-G40; solid arrows, direct relations; dashed arrows, indirect relations. (a) Galactose metabolism pathway. (b) Amino acid accumulation pathway. (c) DNA repair regulation. (d) Glycerol accumulation pathway. (e) Flagellum assembly. UDP, Uridine diphosphate; sn-Glycerol-3P, Glycerophosphoric acid; PRPP, 5-Phospho-alpha-D-ribose 1-diphosphate; UMP, Uridine monophosphate; AIR, Aminoimidazole ribotide; CAIR, 1-(5-Phospho-D-ribosyl)-5-amino-4-imidazolecarboxylate; SAICAR, 1-(5′-Phosphoribosyl)-5-amino-4-(N-succinocarboxamide)-imidazole; IMP, Inosine 5′-phosphate; XMP, Xanthosine 5′-phosphate; GMP, Guanosine 5′-phosphate.
To respond to the extracellular environment changes, organisms constantly transmitted the external signals to control gene expression patterns so that cells could adjust the levels of several metabolites25. Among them, proper intracellular glucose partitioning plays a significant role in maintaining cellular homeostasis26. As the sole glucosyl donor for crucial metabolites, UDP-glucose locates at a strategic point in glucose partitioning24, 27. It is involved in the synthesis of many structural and storage polysaccharides, such as trehalose and β-glucan in yeast24, sucrose and cellulose in plant28, and glycogen in animals29. UDP-glucose pyrophosphorylase (UGPase, Tsac_1476, Tsac_2540) catalyzes the formation of UDP-glucose from glucose 1-phosphate (Glc-1P, Fig. 4) and UTP30. Previous studies demonstrated that the regulation of UGPase expression is related to stress response and long-term survival of yeast cells31. And the storage of carbohydrates, glycogen and trehalose, have been considered as the contribution of stress response32. In particular, trehalose plays an important role as a protectant of protein integrity against hyperosmotic, heat and oxidants stress33,34,35. Taking into consideration that UDP-glucose is the necessary supply of glucosyl for biosynthesis of other carbohydrates, the upregulation of Tsac_1476, Tsac_2540 expression and concentration of UDP-glucose in the evolved strain, is based on anticipated outcome.
Increasing amino acid concentration in cells
When exposed to hypertonic surroundings, cells accumulate amino acids to resist high-sugar conditions through several pathways36,37,38. In SCUT27/Δldh versus G58 profile, Tsac_2230, Tsac_2231, encoding an ABC transporter responsible for the organic acids produced in branched-chain amino acid catabolism (Fig. 4b), were significantly upregulated by 18.752- and 8.017-fold, respectively (Table 2). Similarly, genes related branched-chain amino acid metabolism in the LA1002 versus G40 profile, were also upregulated by 4.20- to 5.20-fold (Fig. 4b). Metabolomics study has demonstrated that 2-hydroxyisovalerate, 2-hydroxyisocaproate, and 2-hydroxy-3-methylvalerate were the most robustly increased metabolites in response to osmotic stress38. All three compounds are 2-hydroxy carboxylic acid derivatives of the branched-chain amino acids valine, leucine, and isoleucine, respectively, and ultimately exported from the cell by the ABC transporter39. Intracellular amino acid concentration measurement showed that no obvious changes in the branched-chain amino acids were detected (Table 3).
In this study, several amino acids, Asp, Glu, Pro, Gly, His and Tyr were accumulate to high concentrations in response to osmotic stress (Table 3). Meanwhile, we found that the contents of intracellular Asp and Glu were the highest in the both evolved strains. It was reported that Glu was the direct biosynthetic precursor for Pro and Arg40. Similarly, Asp was the precursor of Lys and Thr, and Gly was the Cys synthesis precursor, respectively41. As important compatible solutes, glutamate, aspartate and glycine play important roles in physiological adaptation to high external osmolarity37, 42. Glycine addition can increase the survival rate of S. cerevisiae in a hyperosmotic medium43. Pro accumulation is the primary defense response by protecting membrane from damage when plants adapt to salt stress36, 42. Asp and Glu could increase unsaturated fatty acids synthesis, enhance lipid transport and improve cytomembrane fluidity44, 45. And the permeability and fluidity of cell membrane were related to the resistance to turgor pressure in A. pasteurians 46. All these supported that the increase in levels of Asp, Glu and Gly and their derivative amino acids was quantitatively most important.
Activation of DNA repair
DNA repair regulation was complex in the two transcriptional profiles (Fig. 4c). In the SCUT27/Δldh versus G58 profile, three genes and one gene were upregulated and downregulated, respectively. These genes were involved in nitrogen metabolism (ko00910), glutamate metabolism (ko00250), and purine metabolism (ko00230) according to KEGG Pathway database. All the regulation may lead to increased synthesis of xanthosine monophosphate (XMP), which is the precursor of guanosine monophosphate (GMP)47, 48. A similar phenomenon of gene regulation for GMP accumulation was observed in the LA1002 versus G40 profile.
Tsac_2588 was upregulated in both profiles. Tsac_2588 encodes inosine-5′-monophosphate dehydrogenase (IMPDH), which catalyzes the transformation of inosine monophosphate to XMP with NAD+. This reaction not only is the first step in GMP synthesis but also is the rate-limiting process49. IMPDH plays a major role in regulation of intercellular GMP, DNA, and RNA synthesis50; signal transduction51; and membrane glycoprotein synthesis52. The regulations in both profiles indicated the role of GMP in DNA repair initiation. Tsac_2813, another DNA repair-related gene, was upregulated in the LA1002 versus G40 profile and encoded single-strand binding proteins (SSBs), which can increase the specificity of DNA polymerase against heat treatment53 and prevent premature annealing54. They considerably affect DNA replication, repair, and recombination in bacteria55. As indicated by the comet assay results, DNA repair-related genes are activated, because high osmolality promotes DNA damage to nucleus pulposus cell56.
Other altered pathways in the two transcriptional profiles
In the SCUT27/Δldh versus G58 profile, glycerol consumption was inhibited through the downregulation of Tsac_1205 and Tsac_0493 (Fig. 4d). These genes encode the enzymes of glycerol-3-phosphate dehydrogenase (mtGDP), thereby catalyzing glycerophospholipid oxidation for subsequent glycolysis57, 58. The downregulation of mtGDP resulted in intracellular glycerol accumulation, which counteracts high substrate conditions59, 60.
In the LA1002 versus G40 profile, Tsac_0296, Tsac_1788, Tsac_1792, and Tsac_0296 encoding flagellar structure proteins were all upregulated more than four times according to the RNA sequencing results (Fig. 4e). Previous reports61, 62 emphasized that the genes involved in flagellation are activated by regulator genes, such as those in Salmonella enterica serovar Typhi, under hyperosmotic conditions.
Methods
Strains and culture conditions
Table 4 shows the bacteria used in this study. The engineered strain T. aotearoense SCUT27/Δldh, in which the gene encoding lactate dehydrogenase was deleted to increase ethanol yield, was obtained and preserved11. Normal cell culture was described in the references11, 12, 23, and 1 g/L of yeast extract was added into the medium during adaptive domestication to provide abundant amino acid sources. A total of 5 mL of stock culture was activated by transferring it into 125 mL of serum bottles, each of which contained 50 mL of fresh modified MTC medium and incubated at 55 °C and with a nitrogen gas headspace.
Adaptive evolution of T. aotearoense SCUT27/Δldh
T. aotearoense SCUT27/Δldh was first cultured in medium containing 81 g/L of sugar (glucose:xylose = 2:1, g:g) until the DCW reached approximately 0.7 g/L (approach to stationary phase). The saturated culture was then inoculated into fresh MTC media at the ratio of 1:10. The required time (t) for bacterium concentration to reach 0.7 g/L was recorded, and the specific growth rate (g/L/h) was calculated as cell density divided by the needed time. When the specific growth rate remained higher than 0.1 g/L · h at three consecutive times, sugar concentration was gradually enhanced from 102 g/L to 120 g/L. A total of 0.2 mL of culture sample was collected at passage 58 and plated on solid DSMZ 640 medium with 2% agar19. Clones were selected randomly after streak plating, and the selected clones were incubated at 55 °C for 3 days. Isolated single colonies were then transferred into separate tubes containing modified MTC medium with 120 g/L of sugars to verify the resistance of the clones against high osmotic pressure. The final isolated clone was designated as SCUT27/Δldh-G58.
Flask-fermentation analysis of SCUT27/Δldh and G58
For the investigation of the growth characteristics of SCUT27/Δldh and G58 cells, they were cultivated in 125 mL serum bottles containing different sugar concentrations (30, 81, and 120 g/L). The cells were sampled every 6 h, and cell density was measured. All experiments were performed in triplicate.
Propagating stability was analyzed every 10 passages after sequential passing on. The final cell density and ethanol production were monitored after incubation with 120 g/L of sugar for 24 h.
Batch fermentation in 5 L fermenter
Batch fermentation was carried out in a 5 L of BiostatB fermenter (B. Braun, Germany) containing 2 L of medium with 120 g/L of sugar (80 g/L of glucose and 40 g/L of xylose). The saturated cells of SCUT27/Δldh or G58 were inoculated into the fermenter at a ratio of 10% (v/v) and then incubated at 55 °C and stirred at 150 rpm. The pH of the fermentation broth was maintained at 6.5 through automatic addition of 5 M NaOH12. Residual sugars, ethanol, and organic acids were analyzed at specified intervals via high-performance liquid chromatography (HPLC).
Transcriptome sequencing and qRT-PCR analysis
Start strain of SCUT27/Δldh and LA1002 were cultured with 30 g/L mixed sugars (glucose:xylose = 2:1, g:g), and the evolved SCUT27/Δldh-G58 and LA1002-G40 were incubated with 120 g/L of carbon sources. When the cell density of OD600 reached 0.8–1.0 (in exponential phase), 1.0 OD cell was harvested via centrifugation (12000 rpm × 2 min) at 4 °C for RNA extraction and 200 OD cells for the analysis of intracellular compounds. Total RNA was extracted using RNAprep pure kit (for cell/bacteria, TIANGEN, Beijing, China) according to the manufacturer’s instructions. RNA sequencing and data analysis were performed by Genewiz (Suzhou, China). Given the imperfection and fragmentation of T. aotearoense SCUT27 genome, which has been recently updated, the genome annotation of T. saccharolyticum JW/SL-YS485 (CP003184) in NCBI was selected as reference sequence, because the similarity identities of these species is similar by 99%14.
Quantitative real time-PCR (qRT-PCR) was implemented according to standard protocol14. Primers were designed under the guidance of IDT website (http://www.idtdna.com/primerquest/Home/Index). Annealing temperature of primers was set in the range of 62–66 °C, and the length of amplicon was set at 100 bp (see Supplementary Table S4). cDNAs were reverse-transcribed using 1 μg of extracted RNA as template. PCR reactions were carried out in 400 nM of each specific primer, SYBR Premix Ex Taq II (TliRNaseH Plus), and 1 μL of diluted cDNA (~100 ng/μL) in a final volume of 20 μL. PCR reactions were run on LightCycler 96 (Roche, Basel, Switzerland) with 30-s incubation at 95 °C, followed by 45 cycles of 95 °C (5 s) and 60 °C (5 s). Sterile Milli-Q water and 16 s rRNA were designed as background control and internal reference, respectively. Three biological replicates were done for each gene, and the results were analyzed via LightCycler 96 SW 1.0 software (Roche).
Analytical methods
The residual sugars and fermentation products were analyzed via HPLC (Waters 2695, Milford, MA) equipped with an Aminex 87H column (Bio-Rad, Hercules, CA), and a refractive index detector (Waters 2414, Milford, MA). The mobile phase was 5 mM H2SO4 at a flow rate of 0.6 mL/min. The detector and column temperature were set at 40 and 60 °C, respectively. All the samples were passed through 0.22 μm filters before loading.
UDP-glucose and amino acids in SCUT27/Δldh and its derivatives were extracted and measured. Cells were collected and resuspended in 4 mL 20 mM phosphate buffer. After sonication by high-pressure cell disruptor (Constant Systems 01/40/AA, England), the supernatants were collected by centrifugation at 10,000 × g, 4 °C for 30 min and stored at −20 °C.
For amino acid assay, the samples were treated according to the Standard Press of China (NY/T 887–2010), and monitored using amino acid analyzer (L8900, Hitachi, Japan). For UDP-glucose determination, the collected liquid were gently mixed with a triple volume of 95% ethanol and centrifuged at 10,000 × g, 4 °C for 5 min for protein and polysaccharide removal63. The supernatants were dried at 55 °C and re-dissolved in water and analyzed by HPLC (Shimadzu CBM-20A, Kyoto, Japan) equipped with a Inertsil NH2 column (Shimadzu, Kyoto, Japan), and a diode array detector (Shimadzu SPD-M20A, Kyoto, Japan). The mobile phase was acetonitrile-0.125 M potassium phosphate buffer (60:40, v/v, pH = 3.6) at a flow rate of 1.0 mL/min. The column temperature was set at 30 °C, and the detection wavelength was 262 nm.
Bacterial DCW was determined through a linear correlation equation from the optical density at 600 nm [DCW(g/L) = 0.0371 + 0.3343 × OD600]19. OD600 was monitored using a spectrophotometer (Thermo Fisher Scientific GENESYS 10, Bremen, Germany).
Data Availability
All data supporting the conclusions of this article are included in this published article and its supplementary information files.
References
Weber, C. et al. Trends and challenges in the microbial production of lignocellulosic bioalcohol fuels. Appl. Microbiol. Biot 87, 1303–1315, doi:10.1007/s00253-010-2707-z (2010).
Bayrock, D. P. & Ingledew, W. M. Application of multistage continuous fermentation for production of fuel alcohol by very-high-gravity fermentation technology. J Ind Microbiol Biot 27, 87–93, doi:10.1038/sj.jim.7000167 (2001).
Bai, F. W., Anderson, W. A. & Moo-Young, M. Ethanol fermentation technologies from sugar and starch feedstocks. Biotechnol. Adv 26, 89–105, doi:10.1016/j.biotechadv.2007.09.002 (2008).
Bai, F. W., Chen, L. J., Zhang, Z., Anderson, W. A. & Moo-Young, M. Continuous ethanol production and evaluation of yeast cell lysis and viability loss under very high gravity medium conditions. J. Biotechnol 110, 287–293, doi:10.1016/j.jbiotec.2004.01.017 (2004).
Wang, H., Cao, S., Wang, W. T., Wang, K. T. & Jia, X. Very high gravity ethanol and fatty acid production of Zymomonas mobilis without amino acid and vitamin. J Ind Microbiol Biot 43, 1–11 (2016).
Schaber, J. et al. Biophysical properties of Saccharomyces cerevisiae and their relationship with HOG pathway activation. Eur Biophys J Biophy 39, 1547–1556, doi:10.1007/s00249-010-0612-0 (2010).
Skjerdal, O. T. et al. Changes in cell volume, growth and respiration rate in response to hyperosmotic stress of NaCl, sucrose and glutamic acid in Brevibacterium lactofermentum and Corynebacterium glutamicum. Appl. Microbiol. Biot 43, 1099–1106, doi:10.1007/bf00166932 (1995).
Varela, C., Agosin, E., Baez, M., Klapa, M. & Stephanopoulos, G. Metabolic flux redistribution in Corynebacterium glutamicum in response to osmotic stress. Appl. Microbiol. Biot 60, 547–555, doi:10.1007/s00253-002-1120-7 (2003).
Deesuth, O., Laopaiboon, P. & Laopaiboon, L. High ethanol production under optimal aeration conditions and yeast composition in a very high gravity fermentation from sweet sorghum juice by Saccharomyces cerevisiae. Ind. Crop Prod 92, 263–270 (2016).
Utrilla, J. et al. Engineering and adaptive evolution of Escherichia coli for d-lactate fermentation reveals GatC as a xylose transporter. Metab. Eng 14, 469–476 (2012).
Li, S. et al. High efficiency hydrogen production from glucose/xylose by the ldh-deleted Thermoanaerobacterium strain. Bioresour. Technol 101, 8718–8724, doi:10.1016/j.biortech.2010.06.111 (2010).
Lai, Z., Zhu, M., Yang, X., Wang, J. & Li, S. Optimization of key factors affecting hydrogen production from sugarcane bagasse by a thermophilic anaerobic pure culture. Biotechnol. Biofuels 7, 119–129 (2014).
Zhang, Q. et al. Substrate and product inhibition on yeast performance in ethanol fermentation. Energ. Fuel 29, 1019–1027, doi:10.1021/ef502349v (2015).
Zhu, M. Z., Lu, Y. P., Wang, J. F., Li, S. & Wang, X. N. Carbon catabolite repression and the related genes of ccpA, ptsH and hprK in Thermoanaerobacterium aotearoense. Plos One 10, doi:10.1371/journal.pone.0142121 (2015).
Warnecke, T. E., Lynch, M. D., Karimpour-Fard, A., Sandoval, N. & Gill, R. T. A genomics approach to improve the analysis and design of strain selections. Metab. Eng 10, 154–165, doi:10.1016/j.ymben.2008.04.004 (2008).
Gill, R. T., Wildt, S., Yang, Y. T., Ziesman, S. & Stephanopoulos, G. Genome-wide screening for trait conferring genes using DNA microarrays. Proc. Natl. Acad. Sci. USA 99, 7033–7038, doi:10.1073/pnas.102154799 (2002).
Alper, H. & Stephanopoulos, G. Global transcription machinery engineering: a new approach for improving cellular phenotype. Metab. Eng 9, 258–267, doi:10.1016/j.ymben.2006.12.002 (2007).
Shaw, A. J., Hogsett, D. A. & Lynd, L. R. Natural competence in Thermoanaerobacter and Thermoanaerobacterium species. Appl Environ Microbiol 76, 4713–4719, doi:10.1128/Aem.00402-10 (2010).
Yang, X. et al. Efficient production of l-lactic acid by an engineered Thermoanaerobacterium aotearoense with broad substrate specificity. Biotechnol. Biofuels 6, 124–135, doi:10.1186/1754-6834-6-124 (2013).
Tian, L. et al. Simultaneous achievement of high ethanol yield and titer in Clostridium thermocellum. Biotechnol. Biofuels 9, doi:10.1186/S13068-016-0528-8 (2016).
Yomano, L. P., York, S. W. & Ingram, L. O. Isolation and characterization of ethanol-tolerant mutants of Escherichia coli KO11 for fuel ethanol production. J. Ind. Microbiol. Biotechnol 20, 132–138 (1998).
Zaldivar, J., Martinez, A. & Ingram, L. O. Effect of selected aldehydes on the growth and fermentation of ethanologenic Escherichia coli. Biotechnol. Bioeng 65, 24–33 (1999).
Yang, X. F. et al. Valorisation of mixed bakery waste in non-sterilized fermentation for l-lactic acid production by an evolved Thermoanaerobacterium sp. strain. Bioresour. Technol 198, 47–54, doi:10.1016/j.biortech.2015.08.108 (2015).
Daran, J. M., Dallies, N., Thines, S. D., Paquet, V. & François, J. Genetic and biochemical characterization of the UGP1 gene encoding the UDP-glucose pyrophosphorylase from Saccharomyces cerevisiae. Eur J Biochem 233, 520–530, doi:10.1111/j.1432-1033.1995.520_2.x (1995).
Gasch, A. P. et al. Genomic expression programs in the response of yeast cells to environmental changes. Mol Biol Cell 11, 4241–4257, doi:10.1091/mbc.11.12.4241 (2000).
Parrou, J.-L. et al. Dynamic responses of reserve carbohydrate metabolism under carbon and nitrogen limitations in Saccharomyces cerevisiae. Yeast 15, 191–203, doi:10.1002/(SICI)1097-0061 (1999).
Yi, D.-G. & Huh, W.-K. PKA, PHO and stress response pathways regulate the expression of UDP-glucose pyrophosphorylase through Msn2/4 in budding yeast. FEBS Lett 589, 2409–2416, doi:10.1016/j.febslet.2015.07.015 (2015).
Kleczkowski, L. A., Geisler, M., Ciereszko, I. & Johansson, H. UDP-glucose pyrophosphorylase. An old protein with new tricks. Plant Physiol 134, 912–918, doi:10.1104/pp.103.036053 (2004).
Alonso, M. D., Lomako, J., Lomako, W. M. & Whelan, W. J. A new look at the biogenesis of glycogen. FASEB journal: official publication of the Federation of American Societies for Experimental Biology 9, 1126–1137 (1995).
Lamerz, A.-C., Haselhorst, T., Bergfeld, A. K., von Itzstein, M. & Gerardy-Schahn, R. Molecular cloning of the leishmania major UDP-glucose pyrophosphorylase, functional characterization, and ligand binding analyses using NMR spectroscopy. J Biol Chem 281, 16314–16322, doi:10.1074/jbc.M600076200 (2006).
Yi, D. G. & Huh, W. K. UDP-glucose pyrophosphorylase Ugp1 is involved in oxidative stress response and long-term survival during stationary phase in Saccharomyces cerevisiae. Biochem Biophys Res Commun 467, 657–663, doi:10.1016/j.bbrc.2015.10.090 (2015).
Francois, J. & Parrou, J. L. Reserve carbohydrates metabolism in the yeast Saccharomyces cerevisiae. FEMS Microbiol Rev 25, 125–145, doi:10.1016/s0168-6445(00)00059-0 (2001).
Mimura, H., Nagata, S. & Matsumoto, T. Concentrations and compositions of internal free amino acids in a halotolerant Brevibacterium sp. in response to salt stress. Biosci Biotechnol Biochem 58, 1874 (1994).
Rep, M., Krantz, M., Thevelein, J. M. & Hohmann, S. The transcriptional response of Saccharomyces cerevisiae to osmotic shock. J Biol Chem 275, 8290–8300, doi:10.1074/jbc.275.12.8290 (2000).
Benaroudj, N., Lee, D. H. & Goldberg, A. L. Trehalose accumulation during cellular stress protects cells and cellular proteins from damage by oxygen radicals. J Biol Chem 276, 24261–24267, doi:10.1074/jbc.M101487200 (2001).
Measures, J. C. Role of amino acids in osmoregulation of non-halophilic bacteria. Nature 257, 400, doi:10.1038/257398a0 (1975).
Glaasker, E., Konings, W. N. & Poolman, B. Osmotic regulation of intracellular solute pools in Lactobacillus plantarum. J. Bacteriol 178, 575–582 (1996).
Shellhammer, J. P. et al. Amino acid metabolites that regulate G protein signaling during osmotic stress. PLoS Genetics 13, e1006829, doi:10.1371/journal.pgen.1006829 (2017).
Hazelwood, L. A. et al. A new physiological role for Pdr12p in Saccharomyces cerevisiae: export of aromatic and branched-chain organic acids produced in amino acid catabolism. FEMS Yeast Research 6, 937–945, doi:10.1111/j.1567-1364.2006.00094.x (2006).
Aral, B. & Kamoun, P. The proline biosynthesis in living organisms. Amino Acids 13, 189–217, doi:10.1007/bf01372588 (1997).
Umbarger, H. E. Amino acid biosynthesis and its regulation. Annu Rev Biochem 47, 532–606, doi:10.1146/annurev.bi.47.070178.002533 (1978).
Hu, Y. et al. Effects of paclobutrazol on cultivars of Chinese bayberry (Myrica rubra) under salinity stress. Photosynthetica 55, 443–453, doi:10.1007/s11099-016-0658-z (2017).
Thomas, K. C., Hynes, S. H. & Ingledew, W. M. Effects of particulate materials and osmoprotectants on very-high-gravity ethanolic fermentation by Saccharomyces cerevisiae. Appl. Environ. Microbiol 60, 1519–1524 (1994).
Yin, H. S. et al. Effect of aspartic acid and glutamate on metabolism and acid stress resistance of Acetobacter pasteurianus. Microb Cell Fact 16, doi:10.1186/s12934-017-0717-6 (2017).
Stetten, M. R. Mechanism of the conversion of ornithine into proline and glutamic acid in vivo. The Journal of biological chemistry 189, 499–507 (1951).
Fozo, E. A. & Quivey, R. G. Shifts in the membrane fatty acid profile of Streptococcus mutans enhance survival in acidic environments. Appl Environ Microbiol 70, 929–936, doi:10.1128/aem.70.2.929-936.2004 (2004).
Hedstrom, L. IMP dehydrogenase: structure, mechanism, and inhibition. Chem. Rev 109, 2903–2928, doi:10.1021/cr900021w (2009).
Pimkin, M. & Markham, G. D. The CBS subdomain of inosine 5′-monophosphate dehydrogenase regulates purine nucleotide turnover. Mol. Microbiol 68, 342–359, doi:10.1111/j.1365-2958.2008.06153.x (2008).
Kerr, K. M., Cahoon, M., Bosco, D. A. & Hedstrom, L. Monovalent cation activation in Escherichia coli inosine 5′-monophosphate dehydrogenase. Arch Biochem Biophys 375, 131–137, doi:10.1006/abbi.1999.1644 (2000).
Dunkern, T. et al. Virtual and experimental high-throughput screening (HTS) in search of novel inosine 5′-monophosphate dehydrogenase II (IMPDH II) inhibitors. J Comput Aid Mol Des 26, 1277–1292, doi:10.1007/s10822-012-9615-5 (2012).
Markham, G. D., Bock, C. L. & Schalk-Hihi, C. Acid-base catalysis in the chemical mechanism of inosine monophosphate dehydrogenase. Biochemistry 38, 4433–4440, doi:10.1021/Bi9829579 (1999).
Raab, M. et al. In vitro effects of mycophenolic acid on the nucleotide pool and on the expression of adhesion molecules of human umbilical vein endothelial cells. Clin Chim Acta 310, 89–98, doi:10.1016/S0009-8981(01)00527-7 (2001).
Shi, H. Y. et al. Systematic functional comparative analysis of four single-stranded DNA-binding proteins and their affection on viral RNA metabolism. Plos One 8, doi:10.1371/journal.pone.0055076 (2013).
Chakraborty, K. & Bandyopadhyay, S. Dynamics of water around the complex structures formed between the KH domains of far upstream element binding protein and single-stranded DNA molecules. J. Chem. Phys 143, doi:10.1063/1.4927568 (2015).
Raghunathan, S., Ricard, C. S., Lohman, T. M. & Waksman, G. Crystal structure of the homo-tetrameric DNA binding domain of Escherichia coli single-stranded DNA-binding protein determined by multiwavelength x-ray diffraction on the selenomethionyl protein at 2.9-A resolution. Proc. Natl. Acad. Sci. USA 94, 6652–6657 (1997).
Mavrogonatou, E. & Kletsas, D. High osmolality activates the G1 and G2 cell cycle checkpoints and affects the DNA integrity of nucleus pulposus intervertebral disc cells triggering an enhanced DNA repair response. DNA Repair (Amst) 8, 930–943, doi:10.1016/j.dnarep.2009.05.005 (2009).
Gancedo, C., Gancedo, J. M. & Sols, A. Glycerol metabolism in yeasts. Pathways of utilization and production. Eur. J. Biochem 5, 165–172 (1968).
Wang, Y., Tao, F. & Xu, P. Glycerol dehydrogenase plays a dual role in glycerol metabolism and 2,3-butanediol formation in Klebsiella pneumoniae. J. Biol. Chem 289, 6080–6090, doi:10.1074/jbc.M113.525535 (2014).
Tan, H. W., Aziz, A. R. A. & Aroua, M. K. Glycerol production and its applications as a raw material: A review. Renew Sust Energ Rev 27, 118–127, doi:10.1016/j.rser.2013.06.035 (2013).
Nevoigt, E. & Stahl, U. Osmoregulation and glycerol metabolism in the yeast Saccharomyces cerevisiae. Fems Microbiol. Rev 21, 231–241, doi:10.1111/j.1574-6976.1997.tb00352.x (1997).
Sheng, X. M. et al. Mig-14 plays an important role in influencing gene expression of Salmonella enterica serovar Typhi, which contributes to cell invasion under hyperosmotic conditions. Res Microbiol 164, 903–912, doi:10.1016/j.resmic.2013.07.006 (2013).
Du, H. et al. RpoE may promote flagellar gene expression in Salmonella enterica serovar Typhi under hyperosmotic stress. Curr Microbiol 62, 492–500, doi:10.1007/s00284-010-9734-y (2011).
Dai, J., Guan, H. Q., Chen, S. W., Du, G. C. & Chen, J. Separation and determination of uridine diphosphoglucose (UDPGlu) and uridine diphosphogalactose (UDPGal)in the cell free extracts of Corynebacterium glutamicium by HPLC. Chinese Journal of Chromatography 21, 580–583 (2003).
Arora, R., Behera, S., Sharma, N. K. & Kumar, S. Augmentation of ethanol production through statistically designed growth and fermentation medium using novel thermotolerant yeast isolates. Renew Energ 109, 406–421, doi:10.1016/j.renene.2017.03.059 (2017).
Huerta, B. G. et al. Metabolic engineering and adaptive evolution of Escherichia coli KO11 for ethanol production through the Entner-Doudoroff and the pentose phosphate pathways. J Chem Technol Biot 92, 990–996, doi:10.1002/jctb.5138 (2017).
Shaw, A. J. et al. Metabolic engineering of a thermophilic bacterium to produce ethanol at high yield. Proc Natl Acad Sci USA 105, 13769–13774, doi:10.1073/pnas.0801266105 (2008).
Acknowledgements
We acknowledge the State Key Laboratory of Applied Microbiology South China, Guangdong Institute of Microbiology, for technical support and assistance with amino acid and UDP-glucose determination. This study was partially supported by research grants from the International Cooperation Research Program of Guangdong, China (2014A050503024), the Natural Science Foundation of Guangdong, China (2014A030313258), the Science and Technology Planning Project of Guangdong Province, China (2017A010105019), the Science and Technology Program of Guangzhou, China (201704020183) and the Fundamental Research Funds for the Central Universities, SCUT (2015ZZ108).
Author information
Authors and Affiliations
Contributions
M.Z. participated in the experiment design, performance, data analysis, and manuscript writing. W.F. carried out the intracellular solute measurements. Y.C. and Z.L. contributed with SCUT27/Δldh evolution and partial fermentation. X.Y. contributed with LA1002 evolution and analyzed the data. S.L. conceived the experiment, analyzed the data and critically revised the manuscript. X.W. coordinated the study and participated in supervision. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Competing Interests
The authors declare that they have no competing interests.
Additional information
Publisher's note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Zhu, M., Fan, W., Cha, Y. et al. Dynamic cell responses in Thermoanaerobacterium sp. under hyperosmotic stress. Sci Rep 7, 10088 (2017). https://doi.org/10.1038/s41598-017-10514-8
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-017-10514-8
This article is cited by
-
Kinetic study and model of fermentation parameters affected growth and xylitol production in bioreactor by Kluyveromyces marxianus ATCC 36,907
Biomass Conversion and Biorefinery (2023)
-
High production of valencene in Saccharomyces cerevisiae through metabolic engineering
Microbial Cell Factories (2019)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
