
Abstract
We investigated possibility of predicting whether blooms, if they occur, would be formed of microcystin-producing cyanobacteria. DGGE analysis of 16S-ITS and mcyA genes revealed that only Planktothrix and Microcystis possessed mcy-genes and Planktothrix was the main microcystin producer. qPCR analysis revealed that the proportion of cells with mcy-genes in Planktothrix populations was almost 100%. Microcystin concentration correlated with the number of potentially toxic and total Planktothrix cells and the proportion of Planktothrix within all cyanobacteria, but not with the proportion of cells with mcy-genes in total Planktothrix. The share of Microcystis cells with mcy-genes was low and variable in time. Neither the number of mcy-possessing cells, nor the proportion of these cells in total Microcystis, correlated with the concentration of microcystins. This suggests that it is possible to predict whether the bloom in the Masurian Lakes will be toxic based on Planktothrix occurrence. Two species of toxin producing Planktothrix, P. agardhii and P. rubescens, were identified by phylogenetic analysis of 16S-ITS. Based on morphological and ecological features, the toxic Planktothrix was identified as P. agardhii. However, the very high proportion of cells with mcy-genes suggests P. rubescens. Our study reveals the need of universal primers for mcyA genes from environment.
Similar content being viewed by others

Introduction
Blooms of algae, including cyanobacteria, are one of the consequences of eutrophication1, 2. They create various problems, such as decreasing water transparency and species diversity of phytoplankton and other organisms, high values of production and respiration, high oxygen consumption and formation of anaerobic zones, accumulation of toxic hydrogen sulfide at the bottoms of reservoirs and the occurrence of unpleasant odors, and, finally, the presence of various toxins. The latter is particularly detrimental, because toxins produced by algae are dangerous to aquatic organisms and can be hazardous for human health and farm animals, who drink water from the reservoirs. In freshwater ecosystems, cyanobacteria constitute the main group of phytoplankton organisms responsible for toxic blooms3. Eutrophication and climate change, which involve the increase of air and water temperature and higher runoff from the catchment, bring about the occurrence of cyanobacterial blooms. The blooms take place more often, last longer and become toxic more often4, 5.
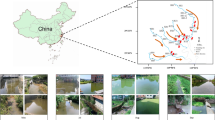
The Great Masurian Lake (GML) system in North-Eastern Poland, comprising several large lakes interconnected by canals, are used intensively for recreation (Fig. 1). The lakes are a holiday destination for hundreds of thousands of people and serve as the pillar of the local economy, which relies on tourism. The outflow of the lakes is connected to the Narew River, which is the main tributary of the Vistula River, a source of drinking water for Warsaw, and one of the last unregulated river systems in Europe.

Study area – the Great Masurian Lakes system. Arrows indicate lakes analyzed in this study. Map prepared in GIMP 2.8.16 software https://www.gimp.org/.
The cyanobacterial communities in the GML are diverse6,7,8,9,10 and often dominate phytoplankton. Most of the cyanobacterial species in these lakes are potentially toxic. Therefore, it is important to determine whether there are toxins in lake waters or if there is a risk that the cyanobacteria will produce toxins. The ability of a cyanobacterial community to produce toxins can be identified either by the direct analysis of toxins in the water or by the analysis of the genes involved in the biosynthesis of the toxins11. These so-called toxicity genes are responsible directly or indirectly for the production of various toxins (e.g. the mcy-gene cluster for microcystins; the sxt, cyr and ana genes for saxitoxin, cylindrospermopsin and anatoxin-a). In the case of microcystins (which are the most frequently-occurring freshwater toxins), genes from the mcy-gene cluster are routinely assayed3. Eight genes, mcyA, B, C, D, E, G, H, J, are always present and are essential for toxin production11.
Information about the number of cells with toxicity genes, without information of the share of potentially ‘toxic’ genotypes in a given taxon of cyanobacteria, allow the recognition that the community is presently formed by toxic cyanobacteria, but it is not enough to predict whether the bloom, if it occurs, will be toxic. Verification of the proportion of toxic cells requires precise tools, which make it possible to count potentially toxic cells and the overall number of cells from a particular taxon. Quantitative PCR (qPCR), using taxa-specific probes allows the estimation of both the total number of cells and the cell number from this taxon with a specific genotype ‒ in this case, possessing ‘toxicity genes’ in the genome. According to the hypothesis of Kurmayer et al.12, the share of genotypes in the environment bearing mcy genes is stable during the vegetation season. Therefore, on the basis of the proportion of potentially ‘toxic’ genotypes in spring, it should be possible to predict whether the bloom, if it occurs later during the season, will be toxic.
There are few studies that have investigated relationships between the proportion of mcy-genotypes in the population of cyanobacteria and the concentrations of microcystins in waters13. Even less is known about the risk of toxic cyanobacterial blooms and the concentrations of toxins in the GML. Mankiewicz et al.14 showed that Masurian waters are not free of microcystins and that their concentrations varied during one sampling from 4 to 12 µg L−1. Recently, genes from the mcy-gene cluster were detected in four of the Masurian Lakes during the entire vegetation season9.
The main objective of our study was to determine – on the basis of the share of cells with toxicity genes in the total number of given cyanobacteria taxon – whether we can predict if the bloom of cyanobacteria, if it occurs, will be toxic. The second objective of the study was to reveal the relationships between environmental parameters and the occurrence of cyanobacteria bearing toxicity genes, their contribution to the total cyanobacteria community, and microcystin (MC) concentrations in the studied lakes. To obtain our results, we investigated: (1) the biomass, structure and contribution of cyanobacteria in the phytoplankton of studied lakes; (2) which taxa are potentially toxic and are responsible for the toxic bloom; (3) if and when genotypes with toxicity genes from the mcy-gene cluster occur in the waters of the Great Masurian Lake system and to which cyanobacterial taxon they belong; (4) the number of cells with toxicity genes in the total number of a given taxon, and how this proportion changes over the season and within the two years of the study; and finally (5) if and what concentrations of microcystins accompany the potentially toxic cyanobacteria in these waters.
Results
During the study period, the trophic status of the studied lakes varied between advanced mesotrophy and eutrophy, with mean trophic state index (TSI) values between 47 ± 4 and 60 ± 4 (Table S1). The maximal and minimal values of TSI were noted in the spring. During summer, the differences between TSI in the studied lakes were smaller, but the respective lakes could still be classified within the same trophic status category. During the course of the study, the northern lakes – Mamry and Kisajno – represented the lowest trophic status, while lakes Niegocin and Tałtowisko could be characterized as slightly eutrophic; and Lake Śniardwy was characterized as eutrophic, and Mikołajskie, Bełdany and Tałty as highly eutrophic. The chlorophyll a concentrations in the studied lakes varied from 6 to over 40 µg L−1, and in five of the eight studied lakes, were above 20 µg L−1 during most of the vegetation season. In all studied lakes in the GML system, in 58 out of 64 samples, the PCR analyses revealed the presence of the mcyA, mcyD and mcyE genes (Table S2).
Phytoplankton biomass, composition and structure
Total phytoplankton biomass varied between 0.6 ± 0.5 mg L−1 of wet weigh in Lake Niegocin in the spring and 5.7 ± 2.3 mg L−1 in Lake Tałty in the summer (Table S1). Bacillariophyceae and occasionally Cryptophyceae dominated among the eukaryotic phytoplankton in the studied lakes in the spring, while Dinophyceae prevailed among the eukaryotes during the summer. Cyclotella spp., Tabellaria flocculosa var. asterionelloides, Asterionella formosa and Aulacoseira granulata prevailed in the Bacillariophyceae community. Cryptophyceae, which dominated occasionally in a few of the lakes in the spring, were represented by Cryptomonas ovata and Rhodomonas lacustris. Ceratium hirundinella dominated among Dinophyceae and occasionally in the total phytoplankton biomass.
During the summer, cyanobacteria accounted on average for 41% ± 12 of the total phytoplankton biomass (Table S1), but the maximal share was as much as 82–91% of the total phytoplankton biomass (91% in Lake Mamry in August 2011; 83% in Lake Kisajno in July 2012; 82% in Lake Tałtowisko in August 2012). Microscopic analysis made the identification of 44 species of cyanobacteria possible. Among the cyanobacterial order Oscillatoriales, Pseudanabaena limnetica was the dominating species, prevailing in the biomass in most of the lakes. Planktolyngbya limnetica, Limnothrix redekei and Planktothrix agardhii and P. agardhii (var.) suspensa were also occasionally plentiful, constituting up to 50% of all Oscillatoriales. In less eutrophicated lakes in the spring and summer, and in the spring in more eutrophicated lakes, Chroococcales and Synechococcales contributed substantially to the cyanobacterial population. Among the Synechococcales, picocyanobacteria, represented by Synechococcus and Cyanobium, dominated in quantity and in the biomass, contributing between 60 and 95% to this group. From among the Chroococcales, Snowella lacustris, S. litoralis and Aphanocapsa spp. were noted in most of the lakes, mostly in July, while Microcystis aeruginosa was present in the northern, less eutrophicated lakes, and only occasionally occurred in the more eutrophic ones in the middle and southern part of the GML system. M. smithii, M. wesenbergii and M. ichtyoblabe were also noted on a few occasions.
From the 44 species of cyanobacteria identified in the samples, 16 were potentially microcystin-producing (including two species of Planktothrix: agardhii and agardhii (var.) suspensa, six species of Microcystis, Pseudanabaena limnetica, Snowella lacustris, Aphanocapsa spp., Dolichospermum flos-aquae and Limnothrix redekei). Six species were potential producers of neurotoxins or other unknown toxins (Table S3).
Phylogenetic analysis of DGGE-derived ITS and mcyA sequences – diversity of toxicity gene-bearing cyanobacteria vs. other cyanobacteria
Phylogenetic analysis of ITS sequences obtained from DGGE bands revealed that most of the OTUs could be assigned to known cyanobacterial genera and/or species. The tree (Fig. 2) shows that eight of the OTUs belong to the Cyanobium/Synechococcus clade and seven to the Dolichospermum/Aphanizomenon/Cuspidothrix clade. Within this clade, one sequence grouped with Cuspidothrix issatschenkoi sequences with high probability. Since C. issatschenkoi was detected by morphological analyses, it is possible that this sequence might come from this species. We were able to assign eight sequences to Leptolyngbyoideae and two others clustered with Romeria sp., though the phylogenetic position of Romeria was questionable on our tree. Five other sequences were assigned to Planktothrix and three to Microcystis. Therefore, all major groups of cyanobacteria identified microscopically were represented in the ITS-DGGE phylogenetic tree. However, neither Planktolyngbya nor Pseudanabaena were identified by phylogenetic analysis. This might be due to the so far unresolved phylogenetic questions concerning most cyanobacterial genera, including Leptolyngbya, Planktolyngbya and Pseudanabaena 15. Therefore, the sequences we have assigned to Leptolyngbyoideae could come from Planktolyngbya or even Pseudanabaena. Two sister clades, Planktothrix rubescens and P. agardhii, had relatively high statistical support and we were able to assign three sequences to P. agardhii and two to P. rubescens. Interestingly, P. agardhii was identified in microscopic analyses, while P. rubescens was not.

Phylogenetic tree obtained with partial 16S and ITS rRNA sequences corresponding to the sequences obtained from DGGE bands. Numbers at nodes represent branch frequencies. Evolutionary trees, involving 110 sequences, were estimated by the PhyML program49, using the Maximum Likelihood method based on the General Time Reversible model with a discrete Gamma distribution model for evolutionary rate differences among sites (4 categories (+G, Gamma shape parameter = 0.730)). Branch frequencies were calculated by aLRT test, according to Anisimova and Gascuel50. The tree with the highest log likelihood (−6266.296) is drawn to scale, with branch lengths measured in the number of substitutions per site. The rate variation model allowed for some sites to be evolutionarily invariable (+I, Proportion of invariant sites = 0.071).
Strikingly, the phylogenetic analysis of DGGE-derived mcyA sequences revealed much less variation. Only two genera: Planktothrix and Microcystis (Fig. 3) were identified. Altogether, eight different Planktothrix and four Microcystis sequences were identified. However, we were not able to assign them to a particular species of Planktothrix or Microcystis based on phylogenetic analysis of mcyA gene fragments.

Phylogenetic tree obtained with mcyA gene fragments corresponding to the sequences obtained from DGGE bands. Numbers at nodes represent branch frequencies. Evolutionary trees, involving 75 sequences, were estimated by the PhyML program49, using the Maximum Likelihood method based on the General Time Reversible model with a discrete Gamma distribution model for evolutionary rate differences among sites (4 categories (+G, Gamma shape parameter = 1.602)). Branch frequencies were calculated by aLRT test, according to Anisimova and Gascuel50. The tree with the highest log likelihood (−2148.459) is drawn to scale, with branch lengths measured in the number of substitutions per site. The rate variation model allowed for some sites to be evolutionarily invariable; however, no such sites were revealed (+I, Proportion of invariant sites = 0.000).
MC toxins by ELISA, HPLC and LC-MS
ELISA: MC concentrations in water, and total MC concentrations
Low concentrations of microcystins were detected in the studied lakes by ELISA during 2012 and 2013, despite the presence of all the three mcy genes in all of the lakes (Tables S2 and S4). The concentrations of MCs in 2012 varied from 0.1 µg L−1, which was at the detection level for the method, to 0.3 µg L−1. In 2013, the concentrations of microcystins in filtered water varied from 0.2 to 0.6 µg L−1. Analysis of microcystins in unfiltered water, performed after the sonication of cells, showed that total concentrations of MCs were from 2 to 20 times higher than those dissolved in water. Using ELISA, we did not detect even trace concentrations of microcystins in Lake Mamry, despite the presence of mcy-genes in most of the samples in this lake (except in May 2011). In September 2013, we did not detect microcystins dissolved in the water, only MCs present in the cells, with the exception of Lake Mikołajskie, in which the concentration of MCs in the water was 0.1 µg L−1 and the total concentration reached 2.1 µg L−1. These were also the highest concentrations measured by ELISA during the entire study (Table S4).
HPLC and LC-MS/MS
HPLC-DAD analysis of cyanobacterial material collected in 2012 and 2013 from the Great Masurian Lakes did not reveal the presence of cyanotoxins. Positive results were obtained when samples were analyzed using the LC-MS/MS method characterized by higher sensitivity. Altogether, seven MC variants were identified, with [Asp3]MC-RR being the most frequent (Table S5). In 2012, MCs were detected in five of the eight lakes. In Mikołajskie Lake, [Asp3]MC-RR was found in samples collected from May to August 2012. This demethylated microcystin was also present in samples from Lake Kisajno (in May and August), Lake Bełdany (in May and August) and Lake Śniardwy (in May).
The LC-MS/MS analyses of cyanobacterial samples collected in 2013 showed the presence of MCs in eight out of nine studied water bodies. No toxin was detected in Lake Mamry. In the eight lakes, the toxins were found almost in all samples, with the exception of samples collected in May from Lakes Tałtowisko and Tałty. Also, in that year, the [Asp3]MC-RR variant was the most common and was identified in all lakes for which positive results were obtained (Table S5).
Results of qPCR in environmental samples – the proportion of toxicity genes: mcyB vs. PC-IGS for Microcystis, and mcyA vs. 16S rRNA genes for Planktothrix
The qPCR analysis with hydrolysis probes showed that in the community of cyanobacteria, the number of Planktothrix cells calculated on the basis of a fragment of the 16S rRNA gene varied between 3.8 × 103 and 3.4 × 107 cells L−1 (Fig. 4a). In 2012, the highest numbers of cells were noted in May in Lake Mikołajskie, and in July in Lake Tałtowisko. In 2013, the highest abundance of Planktothrix cells was noted in July in Lake Ryńskie, and very high in July and September, in Lakes Tałtowisko and Mikołajskie, respectively. The mean cell number of Planktothrix was 3.1 × 106 cells L−1. The analysis of the mcyA gene showed similar values and the number of mcyA-possessing cells varied between 3.1 × 103 and 3.1 × 107 cells L−1, with a mean of 2.9 × 106 cells L−1. The highest numbers of cells with mcyA genes were detected in the same lakes and on the same dates as in the case of Planktothrix cells based on the 16S rRNA gene. Although the maximal numbers of Planktothrix cells and Planktothrix cells with toxicity genes were higher in 2013 than in 2012 the mean values did not differ statistically. In the case of Microcystis, the total cell number varied between 0.0, when the Microcystis cells with PC-IGS genes were not detected by qPCR analysis, and 1.2 × 106 L−1, when the mean cell number during the study period was 2.6 × 105 L−1, which is more than 10 times less than in the case of Planktothrix. The number of mcyB gene copies varied from 0.0 up to 7.0 × 105 L−1, but occasionally mcyB was not detected when PC-genes of Microcystis were found. Similar to Planktothrix, neither the total number of cells nor the number of cells with toxicity genes differed significantly during these two study years. The highest total number of Microcystis cells was noted in Lake Tałtowisko in July 2012, with very high numbers being found in Lake Śniardwy in July and August 2012. Unlike Planktothrix, the highest number of Microcystis cells with mcyB genes was not detected with the maximum total of Microcystis cells, but it was detected in Lake Mamry in July 2012 (Fig. 4b).

The total number of potentially toxic taxa – Microcystis (a) and Planktothrix (b), and the number of their toxic genotypes in the studied Great Masurian Lakes in 2012 and 2013, based on real-time PCR analyses. Mam – Lake Mamry, Tao – Lake Tałtowisko, Mik – Lake Mikołajskie, Ryn – Lake Ryńskie, Sni – Lake Śniardwy. Numbers denote month and year of the sampling.
Detailed analysis of each sample showed that the share of cells bearing mcyA-genes in the community of Planktothrix varied between 75 and 100%, with a mean value of 91% ± 8.5 (Fig. S4). Also, the mean percentage of cells with mcyA genes calculated for each of the study years was similar: 90 and 91% respectively. In each of the four lakes analyzed by qPCR (Mamry, Tałtowisko, Mikołajskie and Śniardwy), the percentage reached 100% at least once in each sampling season. The highest share was noted in all four lakes in the spring, in April or May, and the mean share in the spring for all the studied lakes was 97%. The lowest percentage was noted in Lake Tałtowisko in July 2012, and in Lake Mamry in August and September 2013. The mean shares for summer and autumn were 88 and 86%, respectively.
The share of toxic genotypes in the case of Microcystis varied widely, between 0 and 100% (Fig. S4), but on average it was only 9% ± 24, with a mean share of 2% in 2012, and 13% in 2013. We did not find any seasonal patterns in the share of Microcystis cells with mcyB genes in the total Microcystis cell number. The share varied irregularly in each lake, reaching seasonal maxima in May, July or September.
We also compared the number of Planktothrix and Microcystis cells bearing mcy-genes with counts of all cyanobacterial cells obtained in microscopic analyses. This share was very low in both years; in 2012, the summarized share of ‘toxic’ Planktothrix and Microcystis in the total number of cyanobacteria varied between 0.01 and 0.80%, and in 2013 between 0.02 and 5.89%. The mean values were 0.18 and 1.42%, respectively.
Relationships between environmental parameters and the occurrence of mcy-genes and MCs
The PCA showed that Factor 1 was responsible for 30.2% and Factor 2 for 21.9% of the variability (Fig. 5). Total cell number of Planktothrix (16S rRNA gene) and the number of Planktothrix cells with mcyA correlated positively with each other and with TSI and total Kjeldahl nitrogen (TKN), as well as negatively with Secchi depth (SD). The percentage of Planktothrix cells with mcyA correlated negatively with month of sampling, and air and water temperatures, while the total numbers of Planktothrix cells and Planktothrix cells with mcyA-genes were independent of temperature and time. Total Microcystis cell numbers (based on PC-IGS) correlated slightly with water temperature, while Microcystis cell numbers bearing toxicity genes (mcyB), and the percentage of these cells in total Microcystis (PC-IGS) did not show a correlation with any of the factors or environmental variables.

Principal component analysis (PCA) (Factors I and II) made on the loadings of environment variables and cyanobacterial parameters. SD – Secchi depth, time –month of sampling, air temp – air temperature, water temp – water temperature, chl a – chlorophyll a, TSI – Trophic State Index, TP – total phosphorus, PO4 – orthophosphates, Microc PG-IGS – total Microcystis (PC-IGS), Microc mcyB – potentially toxic Microcystis (mcyB), % tox Microc – share of potentially toxic Microcystis in total number of Microcystis, Plankt 16S rRNA – total Planktothrix (16S rRNA gene), Plankt mcyA – potentially toxic Planktothrix (mcyA), % tox Plankt – share of potentially toxic Planktothrix in total number of Planktothrix.
Spearman’s rank order correlation analysis revealed several significant (P < 0.05) relationships between the occurrences and share of potentially toxic taxa with microcystins (not analyzed in PCA), and confirmed relationships with environmental data (Table S9). The concentrations of MCs in the water, and total concentrations of MCs (in water and intracellular) measured by ELISA correlated positively with the total population of the Planktothrix community, based on qPCR, and with the number of mcyA-carrying cells. Interestingly, the MC concentrations (in the water and in total) did not correlate with the percentage of cells with mcyA in the total number of Planktothrix cells, but they were positively correlated with the share of the total number of Planktothrix cells (based on 16S) and of Planktothrix with the mcyA gene in the total number of cyanobacteria. Additionally, we found that total MCs correlated positively with TP (total phosphorous) and TKN, while total MCs correlated negatively with SD (Table S9). Air temperature significantly positively correlated with the numbers of Planktothrix from microscopic counts, while SD correlated negatively. The total Planktothrix cell number, based on qPCR and number of cells with mcy genes (qPCR-mcyA), correlated positively with the lakes’ TSI and chlorophyll a, and negatively with SD. The percentage of cells with mcyA in the total Planktothrix cell count was negatively correlated with month of sampling, the temperature of the air and water, and with NH4 +. Microcystis cell numbers obtained from qPCR analysis correlated positively with the temperature of the water.
Discussion
The presented results show that all studied Masurian lakes host potentially toxic cyanobacteria. Microscopic analysis revealed that 22, out of 44 identified cyanobacterial taxa, belonged to potentially toxic taxa, including 16 potentially microcystin-producing. Two of them – P. agardhii and M. aeruginosa – are the most common and potent microcystin producers among freshwater phytoplankton16.
Contrary to the high diversity of cyanobacteria occurring in these lakes (identified microscopically and by ITS-DGGE), the DGGE analysis of mcy genes revealed that only Planktothrix and Microcystis carried the mcyA genes. Altogether, we identified 13 different mcyA DGGE sequences, of which we assigned eight to Planktothrix and five to Microcystis, but the phylogenetic analysis did not allow us to identify the species of these potential MC producers.
The possible reason for not detecting mcyA genes in other species is that the primers we used turned out not to be universal. They were designed for main microcystin producers, Microcystis, Planktothrix and Dolichospermum 17 and were considered universal18. Some authors using these primers for environmental samples were able to amplify the mcyA gene fragment only from Microcystis and Planktothrix 13, 15. From other cyanobacterial taxa such as Dolichospermum (Anabaena)17, 19,20,21, Nostoc 19, Leptolyngbya 22 and Cylindrospermopsis raciborskii 23, mcyA sequences were obtained only from cultivated strains, except for one case of Nostoc isolated from a lichen24. The other possible reason we were not able to recognize the diversity of mcy-bearing cyanobacteria is that the mcyA sequences are not known for all taxa. The GenBank database includes mcyA sequences belonging to only ten genera: Dolichospermum (Anabaena), Cylindrospermopsis, Geitlerinema, Leptolyngbya, Mastigocladus, Microcystis, Nostoc, Phormidium, Planktothrix, Radiocystis, of which only three genera (Dolichospermum (Anabaena), Microcystis and Planktothrix) were found in the microscopic analyses of samples from GML. Thus, the low diversity of mcyA genes in GML is likely to be an underestimation, and we cannot rule out that, apart from Planktothrix and Microcystis, other potential microcystin producers were present there. However, in order to prove this, new primers, which would amplify the mcyA-gene fragments also of other taxa from environmental samples, should be designed.
During three years, the studied lakes were characterized by considerably high TSI (between 47 and 60) and chlorophyll a concentrations often exceeding or close to 20 µg L−1, which is considered the threshold level needed to classify the amount of phytoplankton as a bloom25. The mean share of cyanobacteria in the total phytoplankton biomass varied largely in the studied lakes, from 3 ± 1% to 70 ± 22% of the total phytoplankton biomass (Table S1). In our study, the mcy genes were accompanied by very low concentrations of microcystins (Table S4). Previous studies conducted in the GML system pointed to the presence of considerably high concentrations of microcystins (up to 12 µg L−1) in four lakes of the system14. [Asp3, dhb7]MC-RR was the most frequent microcystin variant in studied lakes, with [Asp3, Mdha7]MC-RR and MC-RR occurring more often than the other four variants found. [Asp3]MC-RR is known to be widely distributed in European bodies of water, including many Polish lakes12, 26,27,28 and frequently associated with P. agardhii populations16, 29. This variant was also detected by Mankiewicz et al.14 in four lakes within the GML system. Recently, [Asp3]MC-RR was detected in 11 out of 21 studied water bodies in Poland, and all 11 lakes were dominated by P. agardhii. P. rubescens, on the other hand, was found in only 3 of 238 studied lakes in Poland27. Populations of P. rubescens are also mostly characterized by production of [Asp3]MC-RR16.
According to Kosol et al.30 MC-containing strains of P. agardhii and P. rubescens usually produced 1–2 microcystins, including one of the demethylated MC-RR variants that were found also in our samples. Although the two species of Planktothrix analyzed by these authors differed with respect to pigmentation, the production of chlorophyll a and anabaenopeptins’ per biomass, no significant differences in the MC profile or content of the toxins were observed, which would allow for differentiation between the species. Suda et al.31 reported that the similarity of 16 S rRNA gene sequence and fatty acid composition in P. agardhii and P. rubescens were also high. Analysis of Planktothrix strains from Lake Steinsfjorden in Norway revealed that representatives of both species could be classified as the same ecotypes and chemotypes, suggesting significant overlapping of the two species32. Some strains from the two species can also belong to the same mcyB genotypes, although genotypes consisting exclusively of one species were also recorded. These controversies regarding classification were attributed to frequent recombination events33.
In our case, Planktothrix, identified microscopically as P. agardhii, occurred in all studied lakes, although the proportion of this cyanobacterium in the total number of cyanobacterial cells was very low, and the number of Microcystis was even lower. The very low biomass of identified potentially toxic taxa in the overall cyanobacterial population, and the even lower percentage in the total phytoplankton biomass, may explain the very low concentrations of MCs in the studied lakes.
Planktothrix agardhii is known as a potent toxin producing cyanobacterium that dominates in many eutrophicated shallow lakes12, 26, 34, but can also occur in higher numbers in deep meso-eutrophic and deep eutrophic lakes7, 35. While studying phytoplankton in the GML system in the 70 s, 80 s and 90 s, Spodniewska6, 7, 36, 37 noted very high numbers and biomass of P. agardhii in the studied lakes. Massive blooms of P. agardhii were reported in 1991–1993 in Lake Niegocin, the lake included in the study8. At that time, Lake Niegocin was highly eutrophicated, but with the modernization of the water treatment plant in Giżycko in 1995, the trophic status of the lake gradually decreased38, as did the phytoplankton biomass and share of cyanobacteria in it8, 35. However, recent results suggest that the process of oligotrophication has slowed down, if not stopped, especially in the central and southern part of the GML system10, 38. In this respect, we cannot rule out a repeated increase of Planktothrix biomass in the coming years.
The qPCR analysis revealed that the population of Planktothrix present in the four lakes studied by this method was almost entirely toxic, and the number of all Planktothrix cells and toxic cells correlated with concentrations of microcystins (Table S9).
The high share of toxic genotypes in Planktothrix populations in GML is an interesting phenomenon, because studies elsewhere in Europe demonstrate that the proportion of toxic genotypes in P. agardhii is usually strikingly lower39. Such a high portion of toxic genotypes is characteristic in turn of the phycoerythrin-rich P. rubescens. Kurmayer et al.39 reported a very high proportion of toxicity genes in populations of P. rubescens, which dominated in deep and mesotrophic lakes. Analyzing PC-IGS and microcystin-encoding genes, these authors divided Planktothrix into two lineages: one containing green-pigmented strains, which lacked toxicity genes, and a second containing red-pigmented and green-pigmented strains. Within the second lineage, the green strains were characterized by a low percentage of genotypes with mcy-genes, while the red strains were composed almost solely of genotypes bearing mcy-genes39. Further study carried out in lakes, varying in depth, trophic status, and the presence of green (PC-rich) and red (PE-rich) Planktothrix, confirmed this division and the differences between these two lineages of Planktothrix 12, 33. The authors reported that the percentage of toxicity genes in populations of green strains occurring in shallow lakes varied between 7 and 40%, while for red strains occurring in deep, stratified, mesotrophic lakes, it was between 75 and 100%. The same study showed that the green strains occurring in deep, stratified lakes were also characterized by much lower proportions of mcy genes than red strains from deep lakes, and this share did not differ significantly from that noted for green strains in shallow lakes12. The presence of these two main lineages of Planktothrix, and a third lineage, consisting of green-pigmented strains from tropical regions, was recently confirmed in 138 strains and it was shown that MC and other peptide production depends on Planktothrix phylogeny and ecophysiological adaptations40.
The results of our study are relevant because the Planktothrix occurring in our lakes, which are stratified and eutrophic, contained almost 100% of toxic genotypes, which makes the population more similar to the red P. rubescens, thriving in deep, mesotrophic lakes, than to other green Planktothrix, characteristic of eutrophic, often shallow lakes. The phylogenetic affiliation of two DGGE-ITS sequences to P. rubescens agrees with the high share of toxicity genes. There is, however, an inconsistency with the pigmentation of this Planktothrix closely related to P. rubescens. In our study, we have not analyzed the phycocyanin and phycoerythrin operons. In microscopic analyses, this Planktothrix appeared green, and isolation of filamentous cyanobacteria did not produced red Planktothrix strains. Typical P. rubescens has also not been detected in earlier studies6,7,8, 36, 37. Therefore, our results indicate the presence of a strain of Planktothrix in GML, which, based on ITS analysis, is closely related to P. rubescens and is characterized by a very high share of toxicity genes, typical for P. rubescens. However, like P. agardhii, it is green and thrives in the epilimnion of these eutrophic lakes.
The hypothesis of Tooming-Klunderud et al.41 may shed some light on the subject. They analyzed the whole genomes of eight Planktothrix strains (four red and four green) isolated from two lakes in Norway. They found that strains of the same chemotype, producing the same oligopeptides, were closely related regardless of color. Furthermore, the authors suggested that the ancestral Planktothrix was green and obtained the whole PE gene cluster by horizontal gene transfer, similarly to Synechococcus 42. Based on phylogenetic analyses of the 16S rRNA gene and the PC-IGS operon, Jasser et al.43 hypothesized that strains of Synechococcus from one of the clades (clade M) present in GML were originally green. They proposed that subsequently, during the oligotrophication of these glacial lakes, the strains could have acquired red pigmentation from closely related, ubiquitous, PE-rich strains from another clade (clade B), which allowed them to adapt better to the changing environment. These two hypotheses could explain the nature of the green color in the Planktothrix occurring in GML closely related to P. rubescens.
Another phenomenon is the decrease in the proportion of genotypes containing mcyA genes in the summer. The share of toxic genotypes in P. rubescens is stable during the vegetation season and is close to 100%12, 44. This stability makes it possible to predict whether the cyanobacterial blooms, if they occur, will be toxic. We noted herein a slight, though statistically significant, decrease in the share of toxic genotypes in the summer, when the air and water temperature was higher. Unfortunately, we could not differentiate between P. agardhii and P. rubescens by qPCR. We can, however, hypothesize that the decrease of the percentage of cells with toxicity genes was the consequence of a growing second population of Planktothrix, a typical, green P. agardhii, characterized by no, or a low percentage of toxicity genes. P. agardhii was shown to take advantage of the increased temperature, in which it may even outcompete P. rubescens 45. In line with this, the PCA analysis in our study showed a negative correlation between the proportion of cells with toxicity genes, and the air and water temperature. This should be, however, further investigated, e.g. by using primers that differentiate between these two species or by metagenomic analysis of 16S rDNA and specific microcystin-encoding gene amplicons.
The statistical analyses (Spearman’s rank-order correlation) revealed that the abundance of Planktothrix and toxic Planktothrix genotypes correlated with concentrations of MCs in water, and total concentrations of MC (intracellular and in water). This finding suggests that Planktothrix was the main toxin producer in GML. The correlations of the total number of Planktothrix cells, as well as Planktothrix cells with toxicity genes to TKN and TSI, suggest that potential MC toxicity in the studied lakes may increase along with their further eutrophication. Considering the massive blooms of Planktothrix recorded in the 1980s and 90s, when the lakes were more eutrophicated, such blooms are possible in the future if the trophic status of the studied lakes increases again38. The Spearman’s rank-order correlation analysis demonstrated that MC concentrations correlated with the total number of Planktothrix cells and the number of cells with mcyA genes, but not with the proportion of ‘toxic’ cells. The latter result is rather puzzling, though it could be explained by the fact that the concentrations of MCs in the studied lakes were low. Thus, it is plausible that the percentage of cells with toxicity genes was above a threshold level when the concentrations of toxins became measurable, but the concentrations were too low to trace the relationships with the percentage of toxic genotypes. However, the results suggest that the very high proportion of genotypes containing mcyA genes in the Planktothrix population in GML during most of the season makes it possible to predict whether the bloom, if formed by Planktothrix, will be toxic. It also seems that the slight decrease in the share of ‘toxic’ genotypes noted in the summer does not exclude the possibility of forecasting it.
Based on present results, we cannot predict whether the bloom of Microcystis, if it occurs, will be toxic. We did not find any trends in the proportion of toxic genotypes within the total Microcystis cell number over time, or any significant relationships between Microcystis cells with and without mcyB genes, and environmental variables. However, as the numbers of Microcystis detected in this study by microscopic and qPCR analyses were low, it is possible that they were below the detection level for both methods. Also other studies suggest that changes in the share of toxic Microcystis genotypes are too irregular46, 47, and therefore, reliable prediction of toxic Microcystis blooms is difficult. On the other hand Kurmayer and Kutzenberger48 found the proportion of toxic genotypes in Microcystis to be very stable during the entire season, which could make such forecasting possible. However, Bevendorf et al.13 concluded that the microcystin-encoding genes are in general not good indicators of toxin occurrence in the environment, and that new studies should concentrate on measuring toxin gene expression.
Conclusions
Our study suggests that Planktothrix is the main genus responsible for microcystins occurring in the Great Masurian Lake system. Based on the results of qPCR analysis, we can predict that, if there were to be a Planktothrix bloom in GML, it would be toxic. We propose that this analysis is a good, and considerably easy, tool to predict the probability of the bloom being toxic not only in GML, but also in other waters in which Planktothrix occurs. The study also revealed that to detect potentially microcystin-producing species – other than Planktothrix and Microcystis – new primers for mcyA-gene should be designed for the environmental studies, as the ones, which were used routinely up till now are not universal.
As concerns the Great Masurian Lake, we expect that further eutrophication of the system may lead to an increase in Planktothrix biomass, as was already noted in the 1970s, 80s and 90s, and therefore the threat of a rise of toxic Planktothrix blooms in the GML system cannot be ruled out. However, because it is not clear which genotypes would increase along with the eutrophication of these lakes, it is important to regularly monitor the lakes with respect to the presence of potentially toxic genotypes that could be classified as P. rubescens or P. agardhii.
Materials and Methods
The study area and sampling
The study was conducted in The Great Masurian Lake system (GML), located in the North-Eastern part of Poland (54° N and 22° E). The lakes are connected with natural and man-made canals, and are spread North-South for over 100 km (Fig. 1). They are of glacial origin; three of them are moraine-dammed lakes, including the two largest lakes in Poland: Śniardwy and Mamry. The other five sampled lakes represent ribbon-type lakes. All the studied lakes are considerably deep, with summertime thermal stratification, which lasts from June to October. The lakes vary in trophic status from mesotrophic to highly eutrophic (Table S1), and are characterized by rich and mixed cyanobacterial communities9. In 2011, 2012 and 2013, eight lakes were sampled four times during the vegetation season: in the spring (April and/or May) before stratification, during the summer (July and August), and in the early autumn (September). Integrated samples were collected with a 2-L Limnos sampler at 0.5 m depth intervals, from the euphotic zone during the mixing period, and from the epilimnion during the summer stratification period.
Abiotic parameters characterizing the ecosystems were determined during each sampling in all the lakes, and the chlorophyll a concentration was measured. The entire phytoplankton community was analyzed in 2011 and 2012. In 2013, cyanobacterial communities were analyzed microscopically in the four lakes, which were studied by qPCR, in order to assess the share of cyanobacterial strains bearing mcy genes in the total number of cyanobacterial cells. The diversity of cyanobacteria was studied using DGGE analysis of the 16S-ITS gene fragment and the diversity of potentially toxic taxa based on a highly conserved domain in the mcyA gene. The sequences obtained from DGGE bands were used to build phylogenetic trees with 16S-ITS and mcyA gene reference sequences.
Four lakes (Mamry, Tałtowisko, Mikołajskie and Śniardwy) were sampled four times in 2012 and in 2013, in order to assess the percentage of toxicity genes (mcyA and mcyB) in the given taxon and in the overall communities of cyanobacteria. Lake Ryńskie, a highly eutrophic lake, was additionally sampled in July and August 2013 for qPCR analyses, and Lake Mikołajskie in June 2013. All methods are described in detail in the Supplementary Information.
References
Dokulil, M. T. & Teubner, K. Cyanobacterial dominance in lakes. Hydrobiologia 438, 1–12, doi:10.1023/A:1004155810302 (2000).
Hudnell, H. K. Proceedings of the interagency, international symposium on cyanobacterial harmful algal blooms (ISOC-HAB): state of the science and research needs. Adv Exp Med Biol 619, 1–924 (2008).
Merilouto, J. & Codd, G. A. Toxic: Cyanobacterial Monitoring and Cyanotoxin Analysis (Åbo Akademi University Press, Åbo, Finland 2005).
Paerl, H. W. & Huisman, J. Climate. Blooms like it hot. Science 320, 57–58, doi:10.1126/science.1155398 (2008).
Paerl, H. W. & Otten, T. G. Environmental science. Blooms bite the hand that feeds them. Science 342, 433–434, doi:10.1126/science1245276 (2013).
Spodniewska, I. Phytoplankton as the indicator of lake eutrophication. 2. Summer situation in 25 Masurian lakes in 1976. Ekol pol 27, 481–49 (1979).
Spodniewska, I. Ecological characteristics of lakes in north-eastern Poland versus their trophic gradient VI. The phytoplankton of 43 lakes. Ekol pol 31, 353–381 (1983).
Napiórkowska-Krzebietke, A. & Hutorowicz, A. Long-term changes of phytoplankton in Lake Niegocin, in the Masurian Lake Region, Poland. Oceanol Hydrobiol Stud 35, 209–226 (2006).
Bukowska, A., Bielczyńska, A., Karnkowska, A., Chróst, R. J. & Jasser, I. Molecular (PCR-DGGE) versus morphological approach: analysis of taxonomic composition of potentially toxic cyanobacteria in freshwater lakes. Aquat Biosyst 10, 2–11, doi:10.1186/2046-9063-10-2 (2014).
Napiórkowska-Krzebietke, A. Cyanobacterial bloom intensity in the ecologically relevant state of lakes – an approach to Water Framework Directive implementation. Oceanol Hydrobiol Stud 44, 97–108, doi:10.1515/ohs-2015-0010 (2015).
Kurmayer, R. & Christiansen, G. The genetic basis of toxin production in cyanobacteria. Freshw Rev 2, 31–50, doi:10.1608/FRJ-2.1.2 (2009).
Kurmayer, R., Schober, E., Tonk, L., Visser, P. M. & Christiansen, G. Spatial divergence in the proportions of genes encoding toxic peptide synthesis among populations of the cyanobacterium Planktothrix in European lakes. FEMS Microbiol Lett 317, 127–137, doi:10.1111/j.1574-6968.2011.02222.x (2011).
Beversdorf, L. J., Chaston, S. D., Miller, T. R. & McMahon, K. D. Microcystin mcyA and mcyE Gene Abundances Are Not Appropriate Indicators of Microcystin Concentrations in Lakes. PLoS ONE 10(5), e0125353, doi:10.1371/journal.pone.0125353 (2015).
Mankiewicz, J. et al. Hepatotoxic cyanobacterial blooms in the lakes of northern Poland. Environ Toxicol 20, 499–506, doi:10.1002/tox.20138 (2005).
Komárek, J., Kaštovský, J., Mareš, J. & Johansen, J. R. Taxonomic classification of cyanoprokaryotes (cyanobacterial genera) 2014, using a polyphasic approach (2014).
Fastner, J. et al. Characterization and diversity of microcystins in natural blooms and strains of the genera Microcystis and Planktothrix from German freshwaters. Arch Hydrobiol 145, 147–163 (1999).
Hisbergues, M., Christiansen, G., Rouhiainen, L., Sivonen, K. & Börner, T. PCR-based identification of microcystin-producing genotypes of different cyanobacterial genera. Arch Microbiol 180, 402–410, doi:10.1007/s00203-003-0605-9 (2003).
Xu, Y. et al. Non-microcystin producing Microcystis wesenbergii (Komárek) Komárek (Cyanobacteria) representing a main waterbloom-forming species in Chinese waters. Environ pollut 156, 162–167, doi:10.1016/j.envpol.2007.12.027 (2008).
Rantala, A. et al. Phylogenetic evidence for the early evolution of microcystin synthesis. Proc Natl Acad Sci USA 101, 568–573, doi:10.1073/pnas.0304489101 (2004).
Prasanna, R., Gupta, V., Natarajan, C. & Chaudhary, V. Bioprospecting for genes involved in the production of chitosanases and microcystin-like compounds in Anabaena strains. World J Microbiol Biotechnol 26, 717–724, doi:10.1007/s11274-009-0228-7 (2010).
Gupta, V. et al. Analyses of diversity among fungicidal Anabaena strains. J Appl Phycol, 1395–1405 doi:10.1007/s10811-012-9793-5 (2012).
Richardson, L. L. et al. The presence of the cyanobacterial toxin microcystin in black band disease of corals. FEMS Microbiol Lett 272, 182–187, doi:10.1111/j.1574-6968.2007.00751.x (2007).
Fathalli, A. et al. Genetic variability of the invasive cyanobacteria Cylindrospermopsis raciborskii from Bir M’cherga reservoir (Tunisia). Arch Microbiol 193, 595–604, doi:10.1007/s00203-011-0704-y (2011).
Oksanen, I. et al. Discovery of rare and highly toxic microcystins from lichen-associated cyanobacterium Nostoc sp. Appl Environ Microbiol 70, 5756–5763, doi:10.1128/AEM.70.10.5756-5763.2004 (2004).
Nebaeus, M. Algal water-blooms under ice-cover. Verh Internat Verein Limnol 22, 719–724 (1984).
Jacquet, S. et al. The proliferation of the toxic cyanobacterium Planktothrix rubescens following restoration of the largest natural French lake (Lac du Bourget). Harm Algae 4, 651–672, doi:10.1016/j.hal.2003.12.006 (2005).
Kobos, J. et al. Cyanobacteria and cyanotoxins in Polish freshwater bodies. Ocean Hydrobiol Stud 42, 358–378, doi:10.2478/s13545-013-0093-8ran (2013).
Rantala, A. et al. Detection of microcystin-producing cyanobacteria in Finnish lakes with genus-specific microcystin synthetase gene E (mcyE) PCR and associations with environmental factors. Appl Environ Microbiol 72, 6101–6110, doi:10.1128/AEM.01058-06 (2006).
Grabowska, M., Kobos, J., Toruńska-Sitarz, A. & Mazur-Marzec, H. Non-ribosomal peptides produced by Planktothrix agardhii from Siemianówka Dam Reservoir SDR (northeast Poland). Arch Microbiol 196, 697–707, doi:10.1007/s00203-014-1008-9 (2014).
Kosol, S., Schmidt, J. & Kurmayer, R. Variation in peptide net production and growth among strains of the toxic cyanobacterium Planktothrix spp. Eur J Phycol 44, 49–62, doi:10.1080/09670260802158659 (2009).
Suda, S. et al. Taxonomic revision of water-bloom-forming species of oscillatorioid cyanobacteria. Int J Syst Evol Microbiol 52, 1577–1595, doi:10.1099/00207713-52-5-1577 (2002).
Rohrlack, T. et al. Oligopeptide chemotypes of the toxic freshwater cyanobacterium Planktothrix can form sub-populations with dissimilar ecological traits. Limnol Oceanogr 53, 1279–1293, doi:10.4319/lo.2008.53.4.1279 (2008).
Kurmayer, R. & Gumpenberger, M. Diversity of microcystin genotypes among populations of the filamentous cyanobacteria Planktothrix rubescens and Planktothrix agardhii. Mol Ecol 15, 3849–3861, doi:10.1111/j.1365-294X.2006.03044.x (2006).
Reynolds, C. S. The ecology of freshwater phytoplankton. (Cambridge Univ. Press, Cambridge and New York, 1984).
Napiórkowska-Krzebietke, A. et al. Changes in the trophic state of Lake Niegocin after the modernization of a local wastewater treatment plant. Limnol Rev 7, 153–159 (2007).
Spodniewska, I. Phytoplankton as indicator of lake eutrophication. 1. Summer situation in 34 Masurian lakes in 1973. Ekol pol 26, 53–70 (1978).
Spodniewska, I. Planktonic blue-green algae of lakes in north-eastern Poland. Ekol pol 34, 151–83 (1986).
Siuda, W., Kaliński, T., Kauppinen, E. & Chróst, R. J. Eutrophication of the southern part of Great Masurian Lakes in 1977–2011. Technologia wody 3, 48–63 (in Polish, 2014).
Kurmayer, R., Christiansen, G., Fastner, J. & Börner, T. Abundance of active and inactive microcystin genotypes in populations of the toxic cyanobacterium Planktothrix spp. Environ Microbiol 6, 831–841, doi:10.1111/j.1462-2920.2004.00626.x (2004).
Kurmayer, R., Blom, J. F., Deng, L. & Pernthaler, J. Integrating phylogeny, geographic niche partitioning and secondary metabolite synthesis in bloom-forming Planktothrix. ISME J 9, 909–921, doi:10.1038/ismej.2014.189 (2015).
Tooming-Klunderud, A. et al. From green to red: Horizontal gene transfer of the phycoerythrin gene cluster between Planktothrix strains. Appl Environ Microbiol 79, 6803–6812, doi:10.1128/AEM.01455-13 (2013).
Six, C. et al. Diversity and evolution of phycobilisomes in marine Synechococcus spp.: a comparative genomics study. Genome Biol R259, doi:10.1186/gb-2007-8-12-r259 (2007).
Jasser, I., Królicka, A. & Karnkowska-Ishikawa, A. A novel phylogenetic clade of picocyanobacteria from the Mazurian lakes (Poland) reflects the early ontogeny of glacial lakes. FEMS Microbiol Ecol 75, 89–98, doi:10.1111/j.1574-6941.2010.00990.x (2011).
Ostermaier, V., Schanz, F., Köster, O. & Kurmayer, R. Stability of toxin gene proportion in red-pigmented populations of the cyanobacterium Planktothrix during 29 years of re-oligotrophication of Lake Zürich. BMC Biol 10, 100–115, doi:10.1186/1741-7007-10-100 (2012).
Oberhaus, L., Briand, J. F., Leboulanger, C., Jaquet, S. & Humbert, J. F. Comparative effects of the quality and quantity of light and temperature on the growth of Planktothrix agardhii and P. rubescens. J Phycol 43, 1191–1199, doi:10.1111/j.1529-8817.2007.00414.x (2007).
Okello, W., Ostermaier, V., Portmann, C., Gademann, K. & Kurmayer, R. Spatial isolation favours the divergence in microcystin net production by Microcystis in Ugandan freshwater lakes. Water Res 44, 2803–2814, doi:10.1016/j.watres.2010.02.018 (2010).
Sabart, M. et al. Spatiotemporal variations in microcystin concentrations and in the proportions of microcystin-producing cells in several Microcystis aeruginosa populations. Appl Environ Microbiol 76, 4750–4759, doi:10.1128/AEM.02531-09 (2010).
Kurmayer, R. & Kutzenberger, T. Application of real-time PCR for quantification of microcystin genotypes in a population of the toxic cyanobacterium Microcystis sp. Appl Environ Microbiol 69, 6723–6730, doi:10.1128/AEM.69.11.6723–6730.2003 (2003).
Guindon, S. & Gascuel, O. A simple, fast and accurate algorithm to estimate large phylogenies by maximum likelihood. Syst Biol 52, 696–704, doi:10.1080/10635150390235520 (2003).
Anisimova, M. & Gascuel, O. Approximate likelihood-ratio test for branches: a fast, accurate, and powerful alternative. Syst Biol 55, 539–52, doi:10.1080/10635150600755453 (2006).
Acknowledgements
This study was supported by grants from the National Science Centre, Poland (N304 102240 and 2015/19/B/NZ9/00473). Experiments were carried out with the use of CePT infrastructure financed by the European Union – the European Regional Development Fund [Innovative economy 2007-13, Agreement POIG.02.02.00-14-024/08-00]. Field studies were performed in the Hydrobiological Station in Mikołajki, Nencki Institute of Experimental Biology of Polish Academy of Sciences. We thank also the European Co-operation in Science and Technology COST Action ES1105 ‘CYANOCOST’ for networking and knowledge-transfer support. We thank two anonymous Reviewers for comments, which greatly improved the paper.
Author information
Authors and Affiliations
Contributions
I.J. designed the research. A.B. and M.K. designed and carried out molecular analyses. A.B., T.K. and I.J. carried out the majority of analyses. H.M.-M. carried out the HPLC and LC-MS analyses. J.K. performed the phylogenetic analyses. I.J. and I.K.-S. compiled the data and performed statistical analyses. A.B., I.J. and J.K. wrote the manuscript. All authors discussed and reviewed the manuscript.
Corresponding author
Ethics declarations
Competing Interests
The authors declare that they have no competing interests.
Additional information
Publisher's note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Bukowska, A., Kaliński, T., Koper, M. et al. Predicting blooms of toxic cyanobacteria in eutrophic lakes with diverse cyanobacterial communities. Sci Rep 7, 8342 (2017). https://doi.org/10.1038/s41598-017-08701-8
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-017-08701-8
This article is cited by
-
Bottom sediments as an indicator of the restoration potential of lakes—a case study of a small, shallow lake under significant tourism pressure
Scientific Reports (2024)
-
Is salinity a driving factor for the phytoplankton community structure of a brackish shallow Mediterranean lake?
Hydrobiologia (2023)
-
Dynamic phytoplankton community structure in a subtropical reservoir during an extended drought, Central Texas, USA
Aquatic Sciences (2023)
-
Effects of Ginkgo biloba extract on growth, photosynthesis, and photosynthesis-related gene expression in Microcystis flos-aquae
Environmental Science and Pollution Research (2022)
-
Morphological, molecular, and biochemical study of cyanobacteria from a eutrophic Algerian reservoir (Cheffia)
Environmental Science and Pollution Research (2022)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
